Introduction
The uterine menstrual cycle consists of four
consecutive phases: Menstrual, proliferative, secretory, and
ischemic. During the uterine proliferative phase, the thickness of
the endometrium increases as a result of stimulation by estrogen
produced by the ovaries. Continuous estrogen stimulation of the
uterus has been known to cause excess proliferation of the
functional layer of the endometrium, which proceeds to the
development of endometrial hyperplasia or atypical endometrial
hyperplasia (1), leading to
infertility. Estrogen, as well as epidermal growth factor (EGF) and
transforming growth factor (TGF) α, are involved in the
proliferation of the endometrium. In normal and hyperplastic
endometria, endometrial glands positive for TGFα are generally
positive for estrogen receptor α (ERα) (2).
Peroxisome proliferator-activated receptors (PPARs)
are members of the nuclear hormone receptor family. They are
ligand-induced transcription factors that regulate the
transcription of target genes. PPARα, which is highly expressed in
hepatocytes, cardiomyocytes, enterocytes, and kidney proximal
tubule cells, is moderately expressed in the uterine glands, cervix
(3,4) and ovarian follicular cells (4,5).
ERα, a nuclear receptor, is activated by estrogens secreted from
follicular lutein cells and binds to estrogen response elements
(EREs) in the promoters of target genes involved in proliferation
of the endometrium functional layer (6). Estrogens can modulate other nuclear
receptor signaling pathways, such as that of PPARs (7). ERα binds to the peroxisome
proliferator response element (PPRE) to interact with the ERE of
the genes, and this transactivation by ERα is inhibited by PPARs
through competition for ERE binding (8).
Astragalus mongholicus var.
membranaceaus Bunge. (synonym A. membranaceaus)
belongs to the family Leguminosae and is distributed throughout
Mongolia, Russia, Kazakhstan, China, Japan, and Korea. The roots of
A. mongholicus, which is used as a traditional medicine in
Japan, is a part of Kampo medicine and indicated for tonic,
cardiotonic, hidroschesis, diuresis, and hypotensive effects. The
root has been reported to possess a wide range of various
biological activities, such as anti-inflammatory properties
(9), antiviral activity, immune
modulation, antineoplastic activity, enhancement of cardiovascular
function (10), anti-tumor
activity (11). Chemical
constituents have studied a lot, and main compounds are determined
as astragalosides (12,13) and isoflavones (14). Astragaloside IV reduces Aβ
production in Alzheimer's disease (15) and has anticancer activities in
breast cancer (16), hepatic
cancer (17), and lung cancer
(18).
In our previous report, a water extract of
Astragalus root (AsR) exhibited PPARα ligand activity, and AsR
decreased renal fatty acid level through PPARα expressed in
proximal tubular epithelial cells, suggesting that AsR prevents
renal damage by fatty acid overload (19). Therefore, we performed an
experiment using mice to determine whether AsR has the likelihood
of improving the function of the reproductive organs through its
PPARα agonistic activity.
Materials and methods
Extract of crude drug for PPARα ligand
test
All crude drugs were purchased from Tochimoto
Tenkaidou Co., Ltd. (Osaka, Japan). Each crude drug was refluxed
for 1 h with 5-fold methanol. The solution was filtered, and the
solvent was removed in a rotary evaporator (EYELA N-1100; Tokyo
Rikakikai Co., Ltd., Tokyo, Japan) under reduced pressure to obtain
the methanol extract.
PPARα agonistic activity by
enzyme-linked immunosorbent assay (ELISA)
PPARα agonistic activity was measured by an ELISA
kit (Enbio RCAS for PPARα; Fujikura Kasei Co., Ltd., Ibaraki,
Japan) according to the manufacturer's protocol. The ligand
activity was calculated as (A-C)/(B-C) ×100, where A is the
absorbance of the sample, B is the absorbance of the positive
control (0.5 mM bezafibrate), and C that of the blank (no
sample).
Comparison among PPARα, PPARγ and ERα
agonistic activities by ELISA
Bezafibrate (Wako Pure Chemical Industries, Ltd.,
Osaka, Japan), β-estradiol (Wako Pure Chemical Industries, Ltd.),
formononetin (Tokyo Chemical Industry Co., Ltd., Tokyo, Japan),
astragaloside IV (Carbosynth Ltd., Berkshire, UK) were used for
ELISA. Cyclic AMP response element binding protein (CRBP) binding
protein (CBP; Bioss Antibodies, Woburn, MA, USA) was immobilized in
the plastic wells at 4°C for 24 h. After washing the wells, 3% skim
milk was added to each well as a blocking reagent. The sample
solution, antigen (PPARα, human recombinant, Fujikura Kasei Co.,
Ltd.; PPARγ, human recombinant; Prospec-Tany TechnoGene Ltd.,
Ness-Ziona, Israel), antibody (PPARα, rabbit polyclonal, GeneTex
Inc., Irvine, CA, USA; PPARγ, rabbit polyclonal, Bio-Rad
Laboratories, Inc., Hercules, CA, USA; ERα, rabbit polyclonal;
Signalway Antibody LLC, College Park, MD, USA) and IgG antibody
conjugated alkaline phosphatase (PPARα and ERα, Human IgG; Bethyl
Laboratories, Inc., Montgomery, TX, USA; PPARγ, Rabbit IgG; Bio-Rad
Laboratories, Inc.) were added to an individual well and incubated
for 60 min at 37°C. Uterus of 13-week-old female ICR strain mice
was used as a source of ERα. The supernatant of homogenized uterus
in phosphate-buffered saline (PBS) was measured the total amount of
protein by a protein assay kit (TaKaRa BCA Protein Assay kit;
Takara Bio, Inc., Otsu, Japan), and 1 µg protein was added to each
well. After washing the wells, p-nitrophenyl phosphate
(SIGMAFAST™ p-nitrophenyl phosphate tablets;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was used as the
substrate for alkaline phosphatase to develop color. After shaking
the plate in a dark place, the absorbance of the solution in each
well was measured at 405 nm. When the absorbance of the sample well
was greater than that of the control well (without addition of
extracts), the sample was deemed to have the agonistic
activities.
Extract of A. mongholicus root for
animal test
A. mongholicus root was purchased from
Tochimoto Tenkaidou Co., Ltd. The dried, chopped-root (5.0 g) was
boiled in 600 ml distilled water, and the final volume reduced by
half. The solution was filtered and concentrated in a rotary
evaporator under reduced pressure at 40°C and then freeze-dried.
The extract was stored at 4°C until use. The yield of the extract
was 1.49±0.13/5.0 g of the chopped root.
Animals
Female SPF/ICR mice were purchased from Charles
River Laboratories, Japan Inc. (Yokohama, Japan), and housed and
maintained under standardized conditions of temperature (25±1°C)
and humidity (55±5%) in a light cycle room (light from 07:00 a.m.
to 07:00 p.m.; dark from 07:00 p.m. to 07:00 a.m.). The mice were
allowed to acclimatize for a week. All experiments were approved by
The Animal Experimental Committee of Tohoku Medical and
Pharmaceutical University, and experimental procedures were
conducted in accordance with the ethical guidelines of the
University.
Female 13-week-old adult mice were divided randomly
into three groups, with five mice in each group. The control, AsR
and bezafibrate groups were fed a standard powdered chow (CE-2;
CLEA Japan Inc., Tokyo, Japan) or chow containing 5% AsR or 0.1%
bezafibrate (Wako Pure Chemical Industries, Ltd.) for 56 days. The
body weight and food intake were measured randomly. Blood samples
were collected from the tail vein and centrifuged at 6,000 × g for
10 min to obtain sera. Visceral adipose tissue (VAT), mammary
glands, uterus, ovary and liver were collected after euthanasia by
ether anesthesia. These tissues were weighed, and the tissues of on
day 56 were prepared for measurements.
Female 20-week-old adult mice (n=5) were
treated with subcutaneous injection of 17β-estradiol at a dose of
0.5 mg/kg body weight once-daily for eight days. When mice were 21
weeks old, blood samples were collected from the tail vein and
centrifuged at 6,000 × g for 10 min to obtain sera. Uteri and
ovaries were collected after euthanasia by ether anesthesia. These
tissues were weigheds and prepared for each measurement.
Quantitative analysis of PPARα
mRNA
Total RNA and mRNA extraction, reverse transcription
and real-time PCR were performed using kits and according to the
manufacturers' protocols. Total RNA was extracted using Nucleo Spin
RNA kit (Macherey-Nagel GmbH & Co. KG, Düren, Germany). mRNA
was isolated from total RNA using the Oligotex-dT30 <Super>
mRNA Purification kit (Takara Bio, Inc.). The mRNA was
reverse-transcribed using Reverse transcription-PrimeScript RT
reagent kit with gDNA Eraser (Takara Bio, Inc.), and cDNA was
generated. To quantify PPARα-mRNA expression, the synthesized cDNA
fragments were amplified using Premix Ex Taq (Takara Bio, Inc.),
and primers and probes for PPARα (assay ID, Mm00440939_m1) and
GAPDH (assay ID, Mm99999915_g1), used as an internal control in
TaqMan Gene Expression Assays (Applied Biosystems; Thermo Fisher
Scientific, Inc., Waltham, MA, USA), were used for qPCR. The qPCR
conditions were as follows: Premix Ex Taq (probe qPCR, 2X conc.),
10 µl; Taqman gene expression assay (premixing primer and prove,
20X conc.), 1 µl; ROX reference dye (50X conc.), 0.4 µl; sample,
2.0 µl; dH2O, 6.6 µl. qPCR was performed on a
StepOnePlus Real-Time PCR system (Applied Biosystems; Thermo Fisher
Scientific, Inc.) with the following PCR cycling conditions:
Holding stage, 20 sec. 95°C; cycling stage, 40X (1 sec 95°C, 20 sec
60°C). The data was analyzed by comparative Cq method using StepOne
Software version 2.3 (Applied Biosystems; Thermo Fisher Scientific,
Inc.). The expression level of PPARα-mRNA was represented by
2−ΔΔCq (20).
Blood estradiol level
Estradiol concentrations in sera were determined
according to the manufacturer's protocol using the Estradiol EIA
Kit (Diagnostic Systems Laboratories, Inc., Webster, TX, USA).
Expression level of PPARα, ERα and
mitochondrial 2,4-dienoyl-CoA reductase (mDECR)
PPARα, ERα and mDECR expression levels were checked
by ELISA using 96-well plastic plates. The uterus, ovary and liver
were homogenized in PBS (uterus: 75 mg/ml; ovary: 10 mg/ml; liver:
100 mg/ml) and centrifuged at 13,000 × g for 15 min at 4°C to
obtain supernatants. The supernatant was measured the total amount
of protein by a protein assay kit (TaKaRa BCA Protein Assay kit;
Takara Bio, Inc.). ELISA was performed as follows. The supernatant
was immobilized in each well at 37°C for 1 h after cAMP response
element binding protein (CRBP; Bioss Antibodies) had been
immobilized at 4°C for 24 h. 3% skim milk was used as blocking
reagent, and Rabbit anti-human polyclonal PPARα antibody (1:500,
44509; GeneTex Inc.), Rabbit polyclonal ERα antibody (1:500, 11071;
Signalway Antibody LLC) or Rabbit anti-human polyclonal DECR1
antibody (1:500, 109608; GeneTex Inc.) was immobilized at 37°C for
1 h as the primary antibody. Goat anti-human polyclonal
ALP-conjugated IgG (1:40, A80-219AP; Bethyl Laboratories, Inc.) or
Goat anti-rabbit polyclonal ALP-conjugated IgG (1:40, 170-6518;
Bio-Rad Laboratories Inc.) was used as a secondary antibody.
p-Nitrophosphate (Sigma-Aldrich; Merck KGaA) was used as a
color reagent and 0.1 M EDTA as a stop reagent. Absorbance was
measured at 405 nm. PPARα, ERα and mDECR expression levels were
represented as a relative ratio of the absorbance compared with the
control group.
Statistical analysis
All data are expressed as the mean ± standard
deviation. The Mann-Whitney U test and ANOVA (Dunnett's test) were
performed using the SigmaStat version 2.03 (Systat Software, Inc.,
San Jose, CA, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
PPARα agonistic activity of crude
drugs
As shown in Table
I, a methanol extract of AsR had the strongest PPARα agonistic
activity among the 15 crude drugs.
 | Table I.PPARα agonistic activities of crude
drugs. |
Table I.
PPARα agonistic activities of crude
drugs.
| Crude drug
name | Representative of
original plant source | Yield of extract
(mg/g) | PPARα ligand
activity (%) |
|---|
| Astragalus
Root | Astragalus
mongholicus |
20.0 | 83.45±4.88 |
| Zinger | Zingiber
officinale |
69.9 | 82.46±11.03 |
| Saposhnikovia Root
and Rhizome | Saposhnikovia
divaricata |
9.6 | 73.64±8.37 |
| Sinomenium Stem and
Rhizome | Sinomenium
acutum |
23.3 | 72.87±15.10 |
| Cnidium
Rhizome | Cnidium
officinale | 174.5 | 63.83±20.82 |
| Japanese Angelica
Root | Angelica
acutiloba |
63.1 | 63.45±11.99 |
| Peony Root | Paeonia
lactiflora | 135.7 | 38.65±3.98 |
| Atractylodes
Rhizome | Atractylodes
japonica |
75.5 | 34.54±17.52 |
| Schizonepeta
Spike | Schizonepeta
tenuifolia |
78.5 | 27.67±5.63 |
| Jujube | Zizyphus
jujuba | 489.3 | 23.58±3.14 |
| Mentha Herb | Mentha
arvensis |
68.8 | NA |
| Glycyrrhiza | Glycyrrhiza
uralensis | 169.6 | NA |
| Rhubarb | Rheum
palmatum | 298.2 | NA |
| Scutellaria
Radix | Scutellaria
baicalensis |
92.0 | NA |
| Ephedra Herb | Ephedra
sinica |
88.4 | NA |
| Bezafibrate |
| – | 78.18±2.97 |
Comparison among PPARα, PPARγ and ERα
agonistic activities
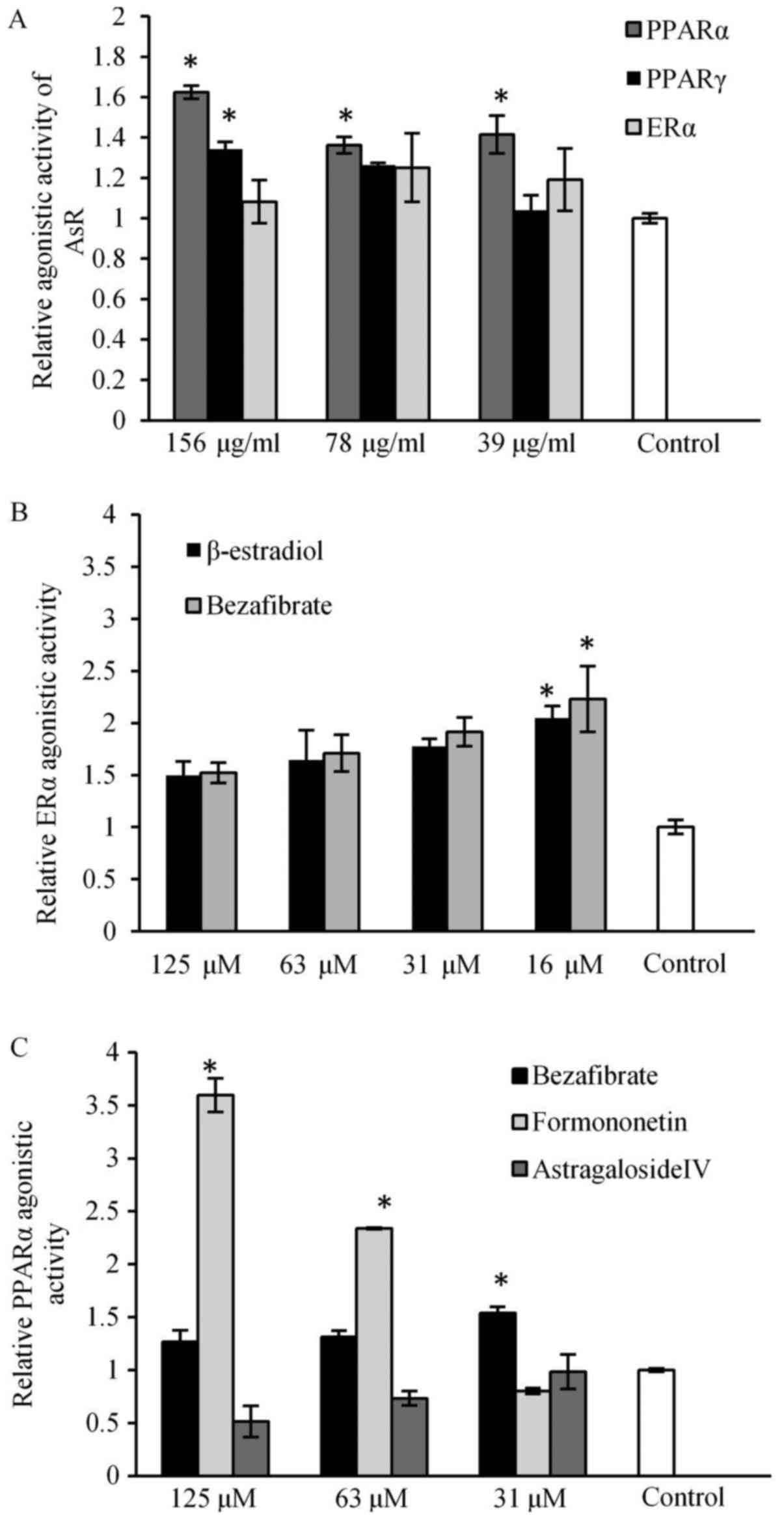
PPARα agonistic activity of AsR was storonger than
PPARγ agonistic activity, and AsR had weak agonistic activity on
ERα (Fig. 1A). Bezafibrate showed
significantly strong potent ERα agonistic activity (Fig. 1B). Formononetin and astragaloside
IV, major ingredients in AsR, were compared with their PPARα
agonistic activities. Formononetin showed PPARα agonistic activity
in a dose-dependent manner, bezafibrate showed significantly strong
PPARα agonistic activity (Fig.
1C).
Body weight and cumulative food
intake
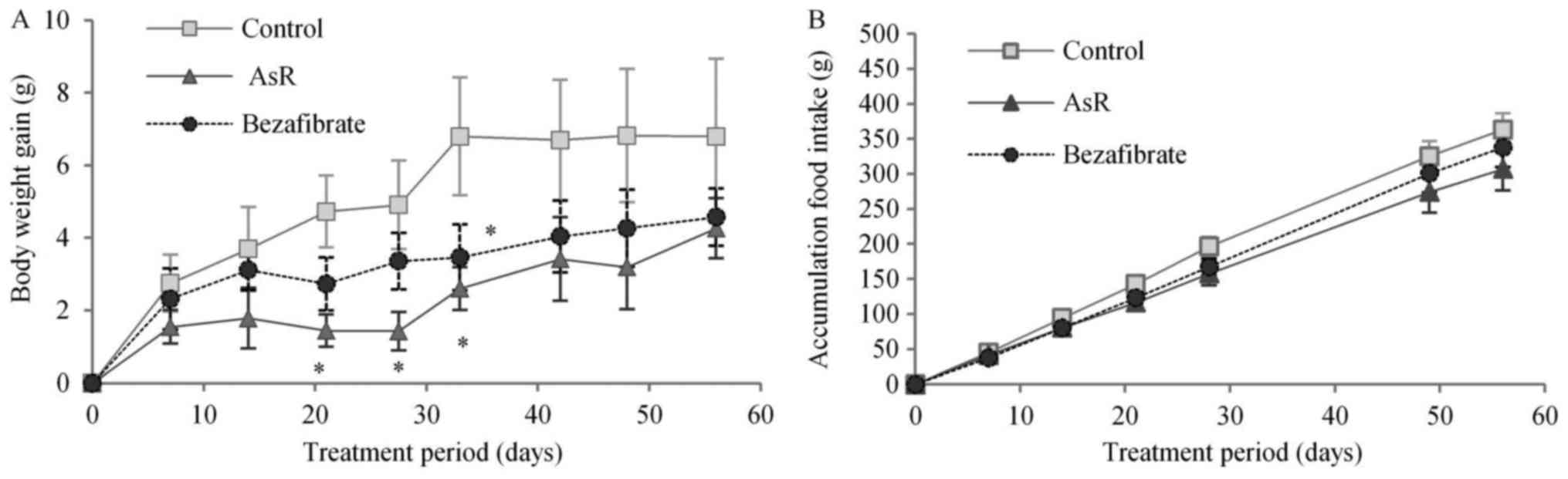
The mice fed bezafibrate and AsR-supplements had
significantly lower body weights compared to controls: AsR, at days
21, 27 and 33; bezafibrate, at day 33 (Fig. 2A). Across all the groups, no
differences were observed in cumulative food intake from day 0
(Fig. 2B).
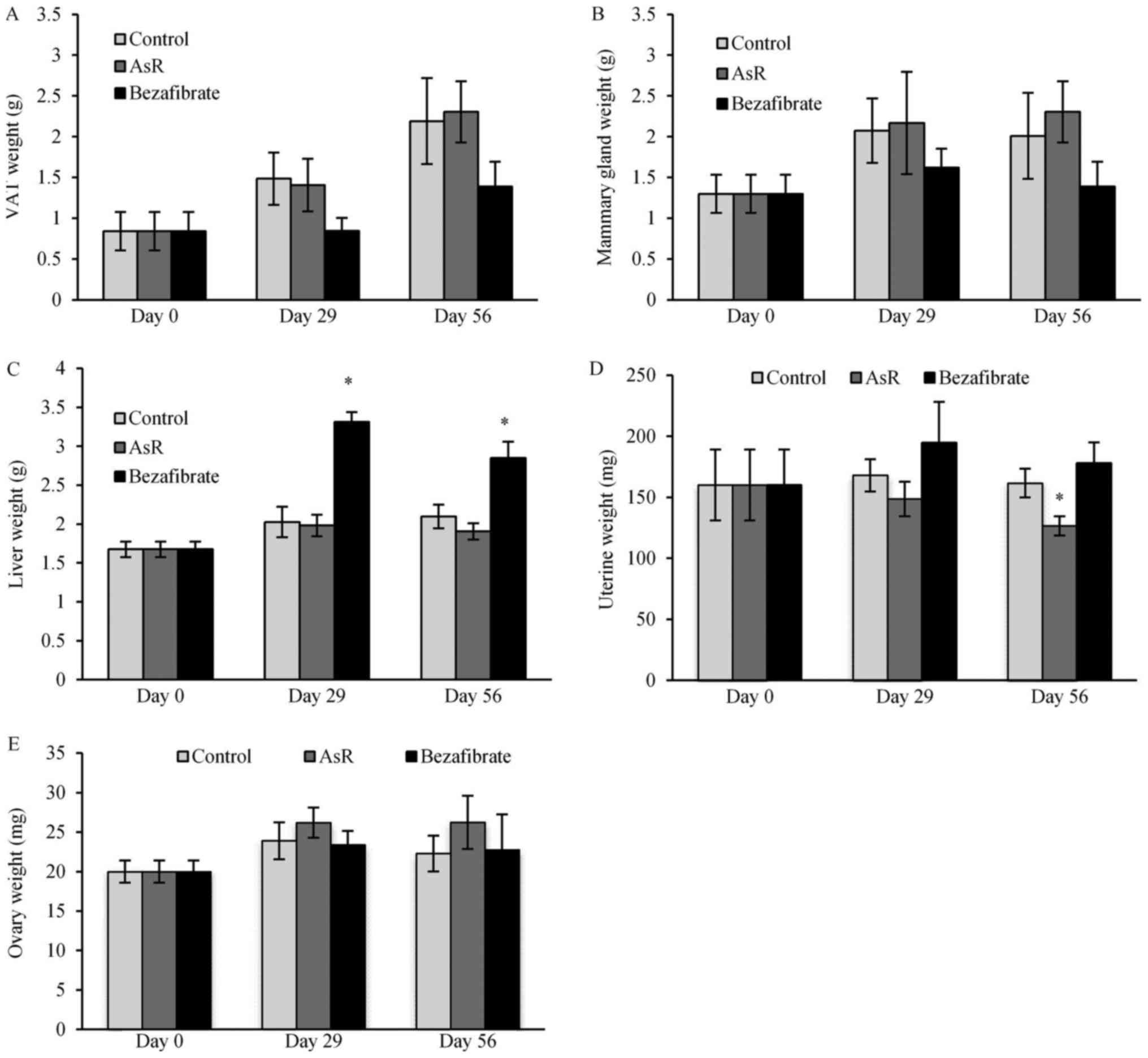
Organ weight
Mice with the bezafibrate treatment showed a trend
toward decreased VAT weight (Fig.
3A) and mammary gland weight (Fig.
3B). Bezafibrate resulted in significantly higher liver weight
on days 29 and 56 (Fig. 3C).
Regarding the reproductive organs, the uterine weights decreased
gradually during AsR treatment: 160±29.05, 148.69±14.13 and
126.64±7.91 mg at days 0, 29 and 56, respectively, but a
significant decrease was detected only on day 56 (Fig. 3D). The uterine weight of mice in
the bezafibrate treatment group showed an increasing trend through
the experiment. The uterine weight of estradiol-treated mice was
significantly higher compared to the control group (Fig. 3D). Ovary weights tended to increase
with AsR treatment (Fig. 3E).
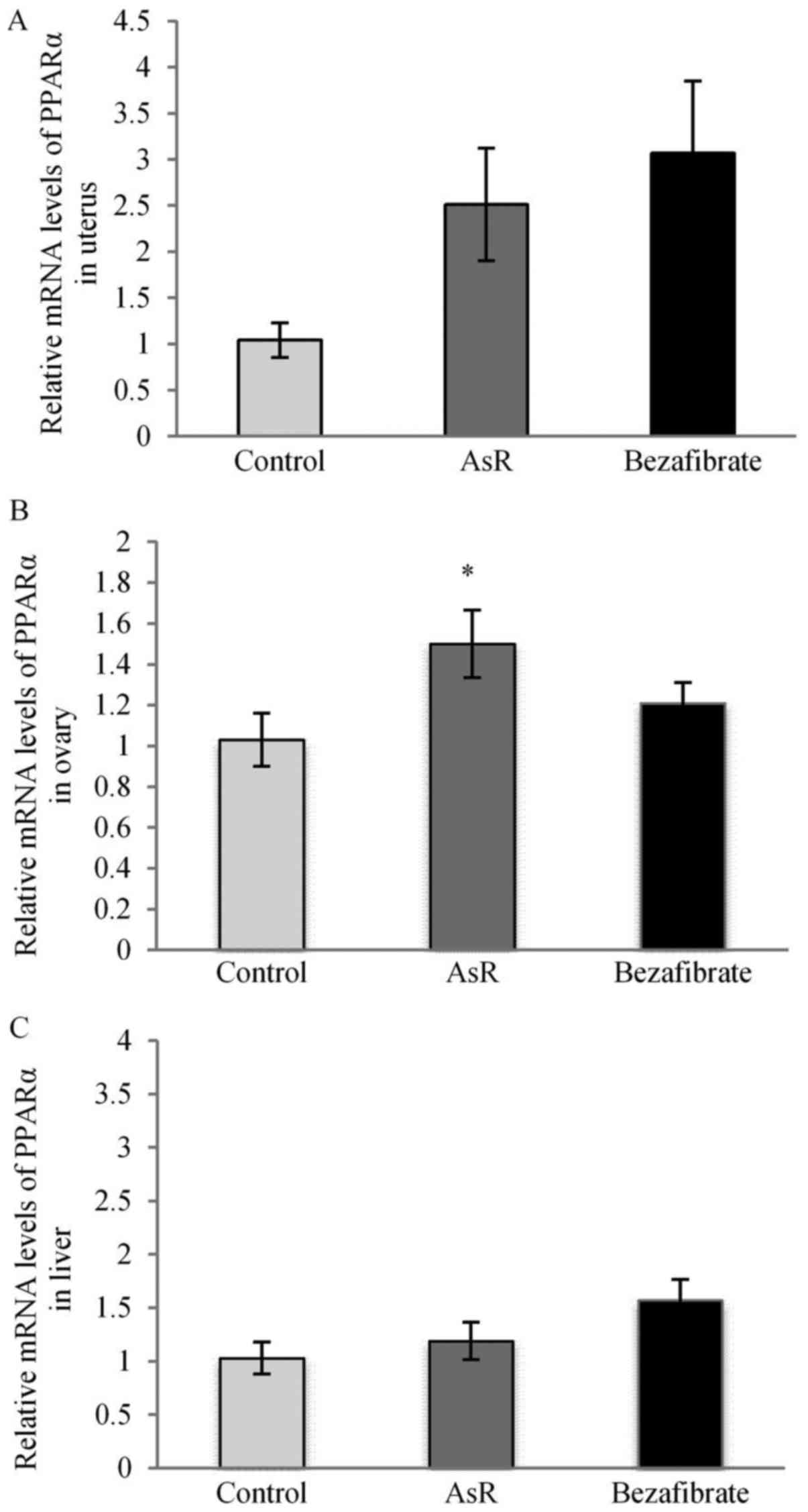
PPARα-mRNA expression levels of
organs
We evaluated PPARα-mRNA expression levels in the
liver and reproductive organs using the comparative Ct method. The
AsR group showed a trend toward increased the PPARα-mRNA expression
levels (by 2.51-fold) in the uterus (Fig. 5A) and significantly increased the
PPARα-mRNA expression levels (by 1.47-fold) in the ovary (Fig. 5B) compared to the control group.
The bezafibrate group showed a trend toward increased the
PPARα-mRNA expression level in the uterus (Fig. 5A) and in the liver (Fig. 5C).
Estradiol levels in serum
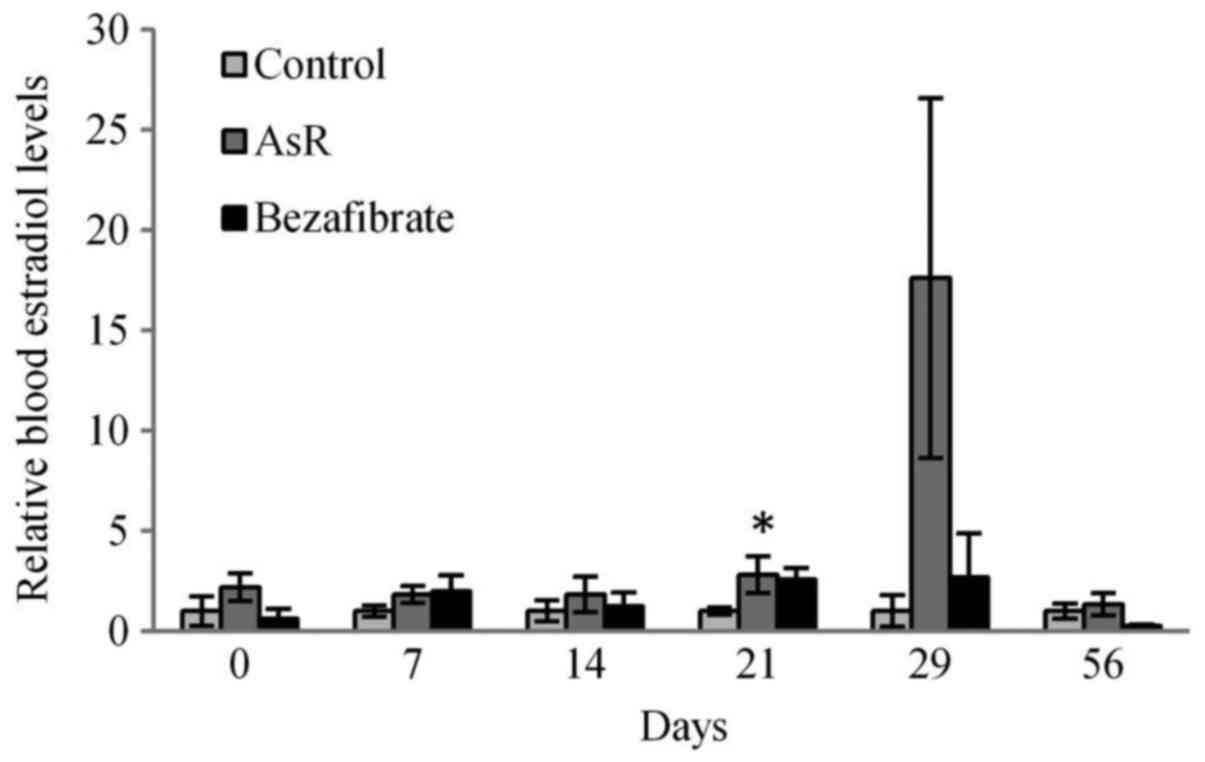
In Fig. 6,
blood-circulating estradiol levels in AsR-treated mice were
significantly higher on day 21, and AsR showed an unstable increase
of blood estradiol levels on day 29.
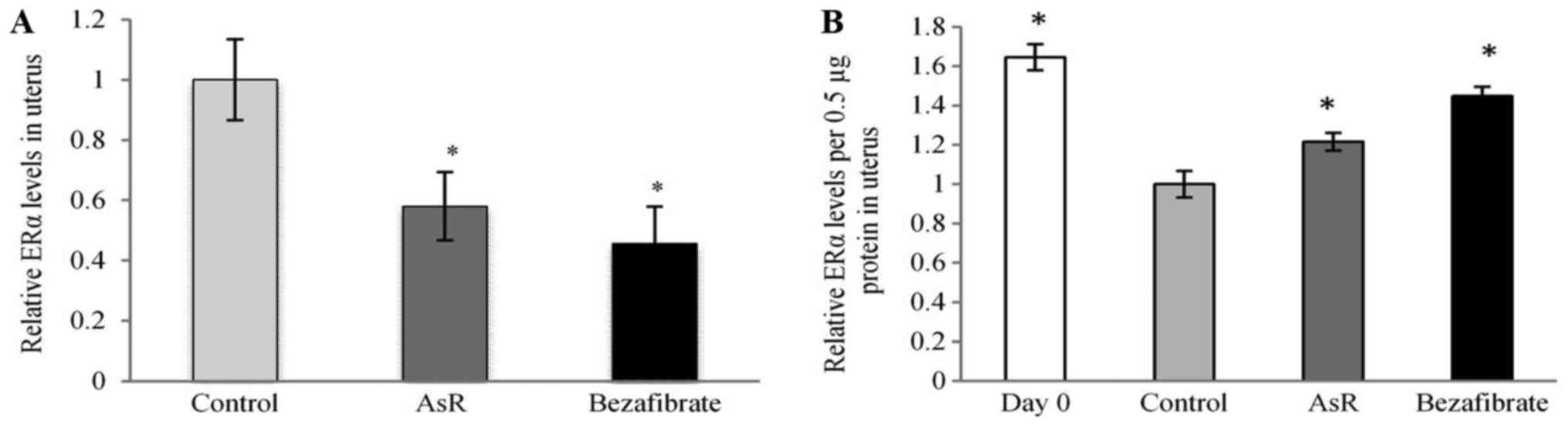
ERα expression level in uterus
The expression levels of uterine ERα were
significantly lower in the AsR- and bezafibrate-treated groups (by
approximately 42 and 55%, respectively) compared to the control
group (Fig. 7A). In Fig. 7B, ERα expression levels per 0.5 µg
protein in the uterus were significantly increased in the AsR- and
bezafibrate-treated groups compared with the control group.
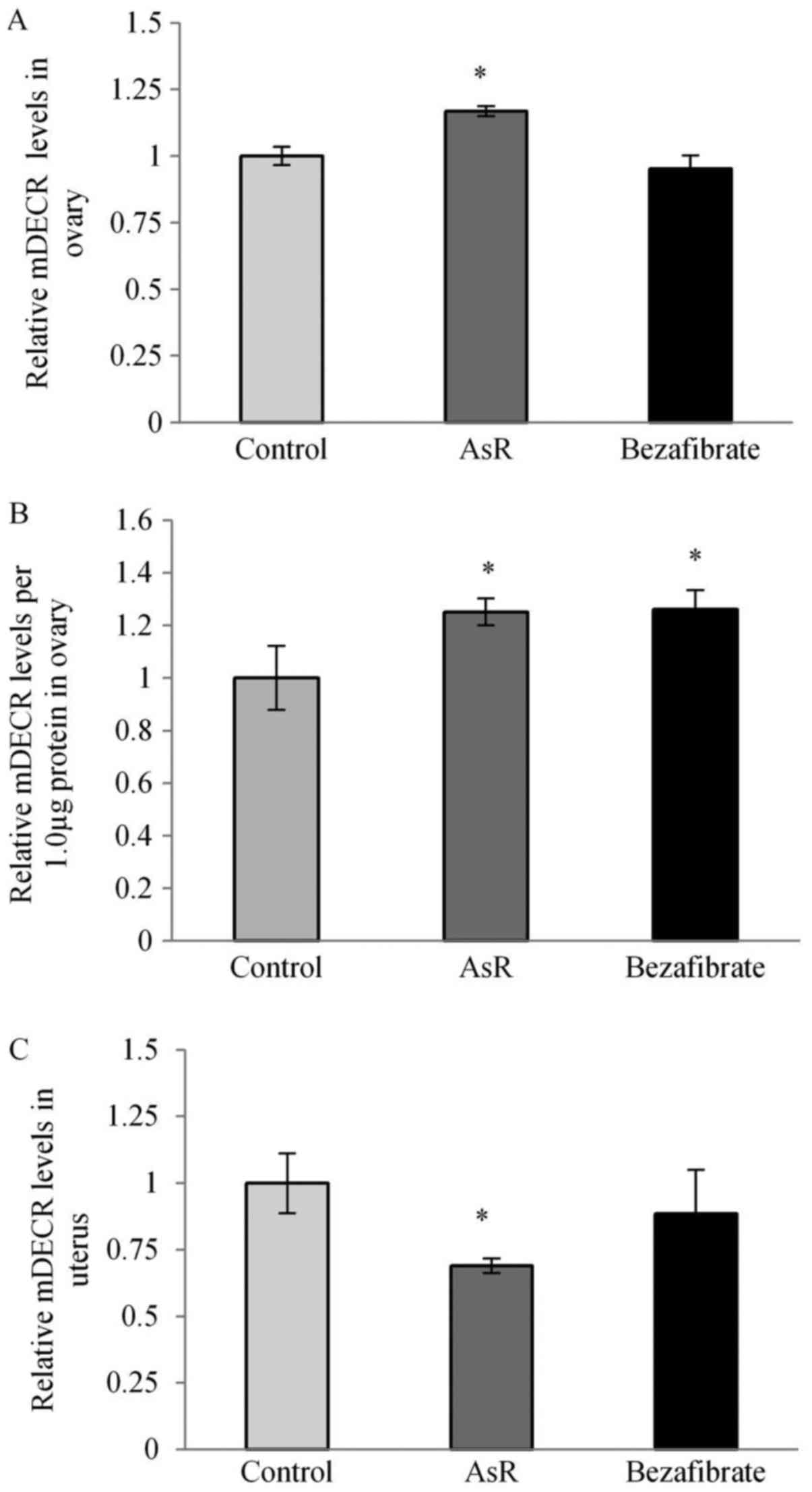
mDECR expression levels in ovary and
uterus
The expression levels of ovarian mDECR was
significantly higher in the AsR-treated group (Fig. 8A). In Fig. 8B, mDECR expression levels per 1.0
µg protein in the ovary were significantly increased in AsR- and
bezafibrate-treated groups. In the uterus, the mDECR expression
level was significantly lower in the AsR-treated group (Fig. 8C).
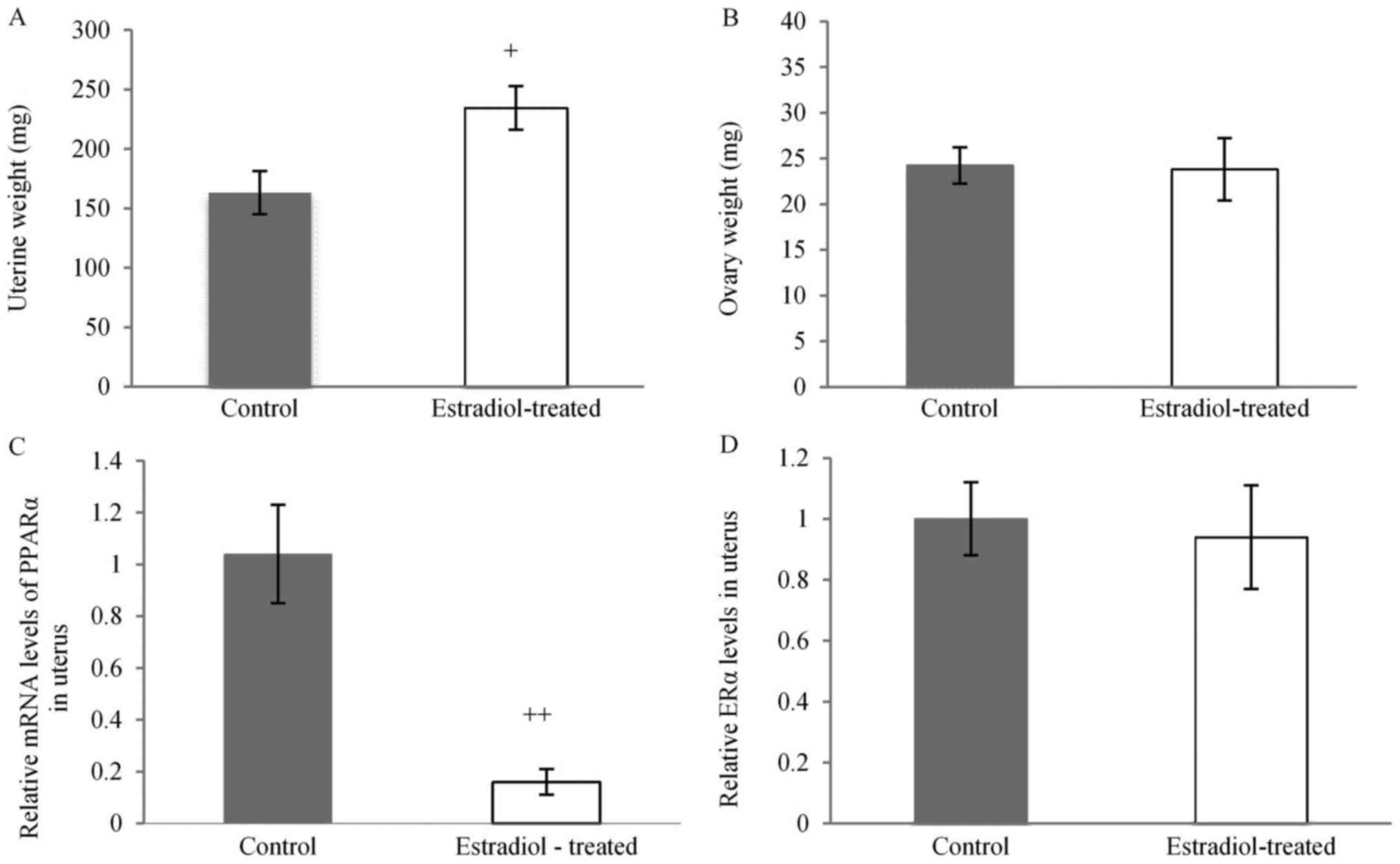
Estradiol-treated mice
The uterine weight of estradiol-treated mice was
significantly higher compared to the control group (Fig. 4A). There is no effect on the ovary
weight of estradiol-treated mice (Fig.
4B). Moreover, the expression levels in the uterus treated with
estradiol was significantly lower compared to the control group
(Fig. 4C). 17β-estradiol-treated
mice had no effect to uterine ERα expression level (Fig. 4D).
Discussion
AsR influenced ovarian proliferation
as a consequence of upregulation of mitochondrial β-oxidation
Fatty acids provide acetyl-CoA through an
enzyme-catalyzed reaction termed β-oxidation, which mainly occurs
in mitochondria and to a lesser extent in peroxisomes. Saturated
fatty acids are easily processed by β-oxidation, but unsaturated
fatty acids are problematic for β-oxidation, and several auxiliary
enzymes are required to generate conformations suitable for
oxidation. Among these auxiliary enzymes, DECR can process the
unsaturated fatty acid trans-2-cis-4-dienoyl-CoA to
trans-3-enoyl-CoA, which is isomerized to a conformation suitable
for β-oxidation. DECR activity is present in mitochondria and
peroxisomes in mammals. There are two mitochondrial isoforms and at
least one peroxisomal isoform of DECR in rats (21,22).
AsR significantly increased the expression of ovarian mDECR
(Fig. 8A) in parallel with
day-dependent increases of ovarian weight (Fig. 3E). The TCA cycle, which produces
ATP as cell energy from acetyl-CoA provided by β-oxidation, is
present in the mitochondria, but it is absent in the peroxisome.
These findings suggest that AsR can produce cellular energy by
increasing mitochondrial β-oxidation. Additionally, PPARα
expression levels increase during follicle development (23). AsR significantly increased the
expression level of ovarian PPARα-mRNA (Fig. 5B). AsR is thought to contribute to
follicle development. Moreover, AsR significantly increased blood
estradiol levels (Fig. 6).
Estradiol-producing cells of the corpus luteum have three
characteristic features: Lipid droplets, smooth endoplasmic
reticulum and mitochondria, which contain the enzymes involved in
the synthesis of estradiol. Follicle stimulating hormone (FSH)
regulates estradiol secretion from ovaries, but mitochondria
proliferation caused by increasing ovarian weight may be related to
the amount of estradiol synthesis in the ovary.
Bezafibrate did not significantly affect ovarian
PPARα-mRNA expression levels or ovarian weight. Bezafibrate is
known to strongly activate human PPARα, whereas bezafibrate
activates murine PPARγ twice as much as PPARα (24). Bezafibrate-treated mice showed a
trend toward a decrease in VAT and mammary gland weight. These data
suggest that bezafibrate promotes white adipose differentiation by
activating murine PPARγ. PPARγ agonistic activity of AsR was weaker
than PPARα agonistic activity in vitro (Fig. 1A) and AsR did not decrease VAT
weight of mice. Therefore, AsR might have no effective agonistic
activity for PPARγ. Bezafibrate increased ovarian mDECR per protein
amount (Fig. 8B), but did not
affect to ovarian mass (Fig. 3E).
PPARγ activation by bezafibrate may prevent ovarian
proliferation.
Mitochondrial dysfunction has been implicated in
cellular senescence in ovarian aging (25). AsR might improve ovarian
dysfunction by upregulating mitochondrial β-oxidation and PPARα
expression.
AsR attenuates the proliferative
action of the uterus
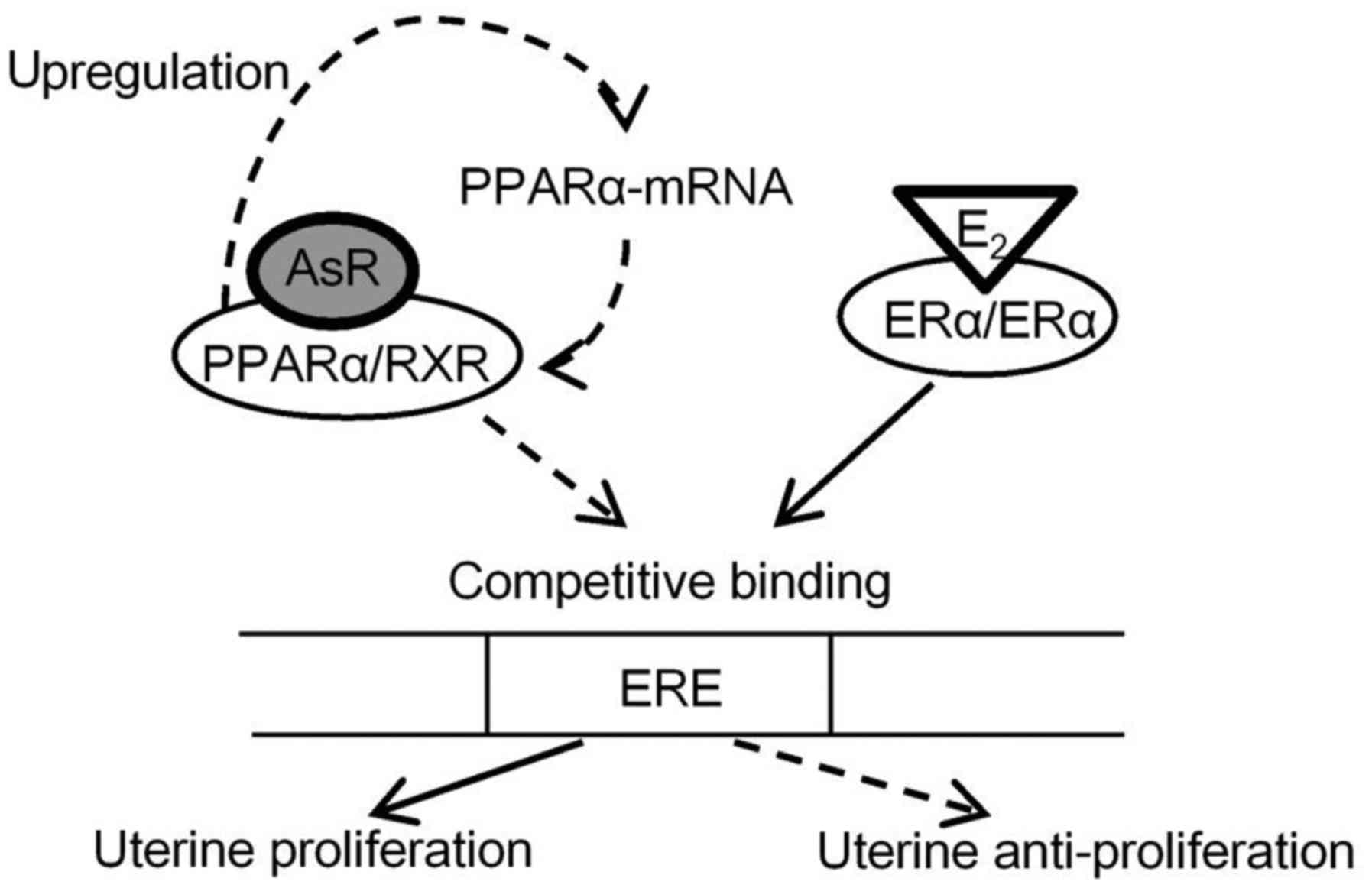
PPARα forms heterodimers with retinoic X receptor
(RXR), binds to PPREs in the promoter of target genes, and can bind
to diverse hormone responsive elements, such as the ERE. ERα binds
to the PPRE sequence to interact with ERE, and this transactivation
by ERα is inhibited by PPARs/RXRs thorough competition for binding
to EREs (8). It has also been
reported that PPARα and ERα share the ability to bind to the AGGTCA
half-site, which occurs as palindrome and as a direct repeat in ERE
and PPRE sequences, respectively (26). Mice treated with estradiol by
subcutaneous injection had significantly decreased expression of
uterine PPARα-mRNA (Fig. 4C). It
is apparent that a negative cross-talk exists between PPARα and ERα
activation or expression, and hence increased uterine PPARα-mRNA
expression occurred in mice treated with AsR (Fig. 5A) resulted in diminished uterine
ERα expression levels (Fig. 7A).
ERα is needed for complete EGF response leading to proliferation of
the endometrium, and ERα has been observed in the endometrium of
endometriosis patients (27).
Fenofibrate, PPARα agonist, was reported that it was influenced to
attenuate of the uterine weight (28). As shown in Fig. 7B, ERα expression level per 0.5 µg
protein in the uterus was significantly higher than that of
control. Fenofibrate, a PPARα agonist, are increased levels of ERα
in randomly selected section of uterus accompanied by decreased in
mitosis and cell proliferation in myometrial cells, stromal cells,
glandular epithelium and luminal epithelium, and also increased
β-catenin in glandular epithelium and luminal epithelium to protect
from the formation of precancerous changes (28). AsR showed weak ERα agonistic
activity and strong on PPARα (Fig.
1A). It is thought that these agonistic activities by AsR might
lead to a decrease of uterine mass. As shown in Fig. 9, AsR affects the molecular
mechanism in PPARα and ERα, AsR might prevent estrogen-dependent
endometrial hyperplasia by downregulating ERα expression through
its PPARα agonistic activity and PPARα proliferation.
Fig. 5A shows that
bezafibrate treatment resulted in increased PPARα-mRNA expression
levels, but also resulted in a trend toward increased uterine
weight (Fig. 3D). The PPARγ
agonist rosiglitazone supports the development of
estrogen-dependent endometrial hyperplasia, and increases the
uterine weight because the PPARγ agonist enhances proliferative and
morphogenetic estrogen action (28). Bezafibrate is thought to act on
murine PPARγ in the uterus. AsR, on the other hand, showed PPARγ
agonistic activity weaker than that of PPARα (Fig. 1A). It is thought that AsR would
activate more strong to uterine PPARα than PPARγ.
In our previous report, AsR increases the
consumption of fatty acids through PPARα expressed in PTECs
(19). In ovary, AsR contributes
the production of energy through the consumption of fatty acids in
mitochondria. Conversely, AsR is not involved in the consumption of
fatty acids in uterine mitochondria (Fig. 8C), suggesting that in the uterine
decreased of the mass by PPARα agonist were suppressed the energy
production by mitochondria.
The diseases of endometrial hyperplasia (EH) and
atypical endometrial hyperplasia (AEH) occur due to continuous
estrogen-stimulation and continuous progesterone-reduction. AEH
increases the risk of endometrial carcinoma, but EH is not
associated with risk of endometrial carcinoma (29,30).
As shown in Fig. 1C, formononetin
that is one of the main constituents of AsR showed strong agonistic
activity on PPARα. Formononetin might prevent uterine proliferative
action by continuous estrogen stimulation as a PPARα activator and
PPARα proliferator. Moreover, AsR increases the secretion of
estradiol caused by ovarian proliferation. Our findings raise the
possibility that AsR is likely to contribute to the improvement of
infertility caused by endometrial hyperplasia and ovarian
dysfunction. In recent studies, oogonial stem cells promote ovarian
regeneration and sustain ovarian function (31,32).
In our research in progress, the ovariectomized mice fed a 10%
AsR-included diet showed rearrangements of the ovary in 4 of 5
tested animals after exposure for 49 days. It is hoped that AsR may
be used as regenerative medicine to treat women suffering from
ovarian failure or extirpation.
Acknowledgements
The authors would like to thank to all the members
of JICA M-JEED project in Mongolia, B. Orkhon, a student on the
doctoral course, obtained financial supports from this project. We
also would like to thank Nobuyoshi Kadowaki and Moe Hayashi,
undergraduates of Tohoku Medical and Pharmaceutical University, for
their efforts in the animal experiments.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during current
study are available from the corresponding author on reasonable
request.
Authors' contributions
BO analyzed and KK designed the research, analyzed
the data and prepared the manuscript. BJ and KS contributed in
guiding research and interpreting the data. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
All experiments were approved by The Animal
Experimental Committee of Tohoku Medical and Pharmaceutical
University, and experimental procedures were conducted in
accordance with the ethical guidelines of the University.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Llorens MA, Bermejo MJ, Salcedo MC, Charro
AL and Puente M: Epidermal growth factor receptors in human breast
and endometrial carcinomas. J Steroid Biochem. 34:505–509. 1989.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Niikura H, Sasano H, Kaga K, Sato S and
Yajima A: Expression of epidermal growth factor family proteins and
epidermal growth factor receptor in human endometrium. Hum Pathol.
27:282–289. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Houston KD, Copland JA, Broaddus RR,
Gottardis MM, Fischer SM and Walker CL: Inhibition of proliferation
and estrogen receptor signaling by peroxisome
proliferator-activated receptor gamma ligands in uterine leiomyoma.
Cancer Res. 63:1221–1227. 2003.PubMed/NCBI
|
|
4
|
Braissant O, Foufelle E, Scotto C, Dauça M
and Wahli W: Differential expression of peroxisome
proliferator-activated receptors (PPARs): Tissue distribution of
PPAR-alpha, -beta, and -gamma in the adult rat. Endocrinology.
137:354–366. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhao Y, Tan YS, Strynar MJ, Perez G,
Haslam SZ and Yang C: Perfluorooctanoic acid effects on ovaries
mediate its inhibition of peripubertal mammary gland development in
Balb/c and C57Bl/6 mice. Reprod Toxicol. 33:563–576. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mitchell DC and Ing NH: Estradiol
stabilizes estrogen receptor messenger ribonucleic acid in sheep
endometrium via discrete sequence elements in its 3′-untranslated
region. Mol Endocrinol. 17:562–574. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bonofiglio D, Gabriele S, Aquila S,
Catalano S, Gentile M, Middea E, Giordano F and Andó S: Estrogen
receptor alpha binds to peroxisome proliferator-activated receptor
response element and negatively interferes with peroxisome
proliferator-activated receptor gamma signaling in breast cancer
cells. Clin Cancer Res. 11:6139–6147. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mu YM, Yanase T, Nishi Y, Takayanagi R,
Goto K and Nawata H: Combined treatment with specific ligands for
PPARgamma: RXR nuclear receptor system markedly inhibits the
expression of cytochrome P450arom in human granulosa cancer cells.
Mol Cell Endocrinol. 181:239–248. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang QH, Han N, Dai N, Wang X and Ao W:
Anti-inflammatory effects and structure elucidation of two new
compounds from Astragalus membranaceus (Fisch) Bge. var.
mongholicus (Bge) Hsiao. J Mol Struc. 1074:284–288. 2014.
View Article : Google Scholar
|
|
10
|
No authors listed: Astragalus
membranaceus. Monograph. Altern Med Rev. 8:72–77. 2003.PubMed/NCBI
|
|
11
|
Chen J, Ge B, Wang Y, Ye Y, Zeng S and
Huang Z: Biochanin A promotes proliferation that involves a
feedback loop of microRNA-375 and estrogen receptor alpha in breast
cancer cells. Cell Physiol Biochem. 35:639–646. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tohda C, Tamura T, Matsuyama S and Komatsu
K: Promotion of axonal maturation and prevention of memory loss in
mice by extracts of Astragalus mongholicus. Br J Pharmacol.
149:532–541. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kitagawa I, Wang HK, Saito M, Takagi A and
Yoshikawa M: Saponin and sapogenol. XXXV. Chemical constituents of
Astragali radix, the root of Astragalus membranaceus BUNGE. (2).
Astragalosides I, II and IV, Acetylastragaloside I and
isoastragalosides I and II. Chem Pharm Bull. 31:698–708. 1983.
View Article : Google Scholar
|
|
14
|
Luo HL, Zhong J, Ye FY, Wang Q, Ma YM, Liu
P, Zhang H, Sunc MY and Jiang J: A systematic quality control
method of Huangqi decoction: Simultaneous determination of eleven
flavonoids and seven triterpenoid saponins by UHPLC-MS. Anal
Methods. 6:4593–4601. 2014. View Article : Google Scholar
|
|
15
|
Wang X, Wang Y, Hu JP, Yu S, Li BK, Cui Y,
Ren L and Zhang LD: Astragaloside IV, a natural PPARγ agonist,
reduces Aβ production in Alzheimer's disease through inhibition of
BACE1. Mol Neurobiol. 54:2939–2949. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jiang K, Lu Q, Li Q, Ji Y, Chen W and Xue
X: Astragaloside IV inhibits breast cancer cell invasion by
suppressing Vav3 mediated Rac1/MAPK signaling. Int Immunopharmacol.
42:195–202. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang PP, Xu DJ, Huang C, Wang WP and Xu
WK: Astragaloside IV reduces the expression level of P-glycoprotein
in multidrug-resistant human hepatic cancer cell lines. Mol Med
Rep. 9:2131–2137. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cheng X, Gu J, Zang M, Yuan J, Zhao B,
Jiang J and Jia X: Astragaloside IV inhibits migration and invasion
in human lung cancer A549 cells via regulating PKC-α-ERK1/2-NF-κB
pathway. Int Immunopharmacol. 23:304–313. 2010. View Article : Google Scholar
|
|
19
|
Kobayashi K, Matsuyama W, Arai Y, Koizumi
S, Shimizu T, Tomioka R and Sasaki K: Boiogito increases the
metabolism of fatty acids in proximal tubular cells through
peroxisome proliferators-activated receptor (PPAR) α agonistic
activity. Biol Pharm Bull. 39:143–147. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCt method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dommes V, Baumgart C and Kunau WH:
Degradation of unsaturated fatty acids in peroxisomes. Existence of
a 2,4-dienoyl-CoA reductase pathway. J Biol Chem. 256:8259–8262.
1981.PubMed/NCBI
|
|
22
|
Hakkola EH, Autio-Harmainen HI, Sormunen
RT, Hassinen IE and Hiltunen JK: The known purified mammalian
2,4-dienoyl-CoA reductases are mitochondrial isoenzymes. J
Histochem Cytochem. 37:1863–1867. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rak-Mardyła A and Drwal E: In vitro
interaction between resistin and peroxisome proliferator-activated
receptor γ in porcine ovarian follicles. Reprod Fertil Dev.
28:357–368. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Willson TM, Brown PJ, Sternbach DD and
Henke BR: The PPARs: From orphan receptors to drug discovery. J Med
Chem. 43:527–550. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang T, Zhang M, Jiang Z and Seli E:
Mitochondrial dysfunction and ovarian aging. Am J Reprod Immunol.
77:e126512017. View Article : Google Scholar
|
|
26
|
Keller H, Givel F, Perroud M and Wahli W:
Signaling cross-talk between peroxisome proliferator-activated
receptor/retinoid X receptor and estrogen receptor through estrogen
response elements. Mol Endocrinol. 9:794–804. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Dassen H, Punyadeera C, Delvoux B,
Schulkens I, Marchetti C, Kamps R, Klomp J, Dijcks F, de Goeij A,
D'Hooghe T, et al: Olfactomedin-4 regulation by estrogen in the
human endometrium requires epidermal growth factor signaling. Am J
Pathol. 177:2495–2508. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gunin AG, Bitter AD, Demakov AB, Vasilieva
EN and Suslonova NV: Effects of peroxisome proliferator activated
receptors-alpha and -gamma agonists on estradiol-induced
proliferation and hyperplasia formation in the mouse uterus. J
Endocrinol. 182:229–239. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kurman RJ, Kaminski PF and Norris HJ: The
behavior of endometrial hyperplasia. A long-term study of
‘untreated’ hyperplasia in 170 patients. Cancer. 56:403–412. 1985.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lindahl B and Willén R: Spontaneous
endometrial hyperplasia. A prospective, 5 year follow-up of 246
patients after abrasio only, including 380 patients followed-up for
2 years. Anticancer Res. 14:2141–2146. 1994.PubMed/NCBI
|
|
31
|
Truman AM, Tilly JL and Woods DC: Ovarian
regeneration: The potential for stem cell contribution in the
postnatal ovary to sustained endocrine function. Mol and Cell
Endocrinol. 445:74–84. 2017. View Article : Google Scholar
|
|
32
|
Erler P, Sweeney A and Monaghan JR:
Regulation of injury-induced ovarian regeneration by activation of
Oogonial stem cells. Stem Cells. 35:236–247. 2017. View Article : Google Scholar : PubMed/NCBI
|