Introduction
PM2.5 is a particle of diameter less than 2.5 µm
common in air pollution. It can be found in engine exhaust gas, and
consists mainly of sulfur oxides (SOx) and nitrogen oxides (NOx).
Because of its small size, the PM2.5 particle can penetrate into
the trachea and lungs by inhalation, and it is considered to cause
damage to the respiratory systems (1). Meanwhile particles of diameter less
than 100 nm are called nanoparticles. Nanoparticles in PM2.5 were
reported to effectively induce ROS production and inflammatory
reactions in the body (2).
A model of PM2.5 nanoparticles can be prepared
employing titanium dioxide (TiO2). TiO2
particles are classified into Rutile, Anatase, and Brookite types
by their crystal structure. Rutile and Anatase types are often used
in sunscreen, food additives and photo-catalytic agents. Moreover,
when added to cultured human fibroblasts or lung cancer cells,
Anatase-type nanoparticles induced cell death about 100 times more
effectively than the Rutile type (3). The size of insoluble nanoparticles
should be important for each biological effect (4). Recently, Tada-Oikawa et al
(5) reported that the Anatase type
TiO2 nanoparticle of 50 nm diameter called A50
effectively induced IL-1β production in differentiated human
monocytic leukemia cells. A50 showed stronger activity than other
tested TiO2 nanoparticles with a different crystal type
or different size of particles.
We previously designed and synthesized
dehydroxymethyl-epoxyqinomicin (DHMEQ) based on the structure of
antibiotic epoxyqinomicin C as an inhibitor of NF-κB (6,7). It
binds to the specific cysteine residues of NF-κB components,
including p65, RelB, cRel, and p50, to inhibit their DNA binding
(8,9). It causes irreversible inhibition of
NF-κB when added to the cultured cells (10), since it binds to the cysteine
residue covalently (11). It was
shown to ameliorate various animal models of inflammation and
cancer without showing any side effect (12,13).
Previously, it was shown to inhibit lipopolysaccharide
(LPS)-induced inflammatory cytokine secretions in mouse monocyte
leukemia RAW264.7 cells (14) and
mouse primary culture macrophages (15). Recently, we reported that DHMEQ
inhibited IL-6 and MCP-1 expressions in primary culture human
peritoneal mesothelioma cells, either with or without stimulation
(16).
In the present study we looked for the signaling
inhibitors of low molecular weight that inhibit TiO2
nanoparticle-induced production of IL-1β. As a result, we found
that DHMEQ inhibited the A50-induced IL-1β production in
differentiated human monocytic leukemia THP-1 cells without
toxicity, possibly by inhibition of transcription alone.
Materials and methods
Materials
The TiO2 particles were purchased from
mkNano, Mississauga, ON, Canada. A50 (anatase type TiO2
with a primary diameter of 50 nm) was prepared as reported
previously (5). DHMEQ was
synthesized in our laboratory as previously described (17). Cytochalasin D is an inhibitor of
phagocytosis, and it was purchased from Wako Pure Chemical
Industries, Ltd., (Osaka, Japan). We used KCl (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) to increase the extracellular potassium
ion concentration. We purchased a caspase-1 inhibitor, zYVAD-FMK,
from Abcam (Cambridge, UK). N-acetylcysteine was used for
anti-oxidant which was obtained from Wako Pure Chemical Industries,
Ltd.
Cell culture and viability
measurement
Human monocytic leukemia THP-1 cells (RIKEN
BioResource Center, Tsukuba, Japan) were cultured in RPMI 1640
medium supplemented with 10% (v/v) fetal bovine serum and
penicillin/streptomycin at 37°C in a humidified incubator with 5%
CO2. THP-1 cells were seeded at 1.5×104
cells/well onto 96-well plates and differentiated into
macrophage-like cells using PMA. The cells were treated with 0.1
µg/ml phorbol 12-myristate 13-acetate (PMA; Sigma-Aldrich; Merck
KGaA) for 72 h. Cell viability was evaluated by trypan blue
(Sigma-Aldrich; Merck KGaA) exclusion assay.
Measurement of IL-1β production
THP-1 macrophages were exposed to different
concentrations of the suspended A50 particles for 24 h at 37°C. The
cell culture supernatant was collected and stored at −20°C until
cytokine analysis. IL-1β concentration in the medium was measured
by ELISA kit (BioLegend, Inc., San Diego, CA, USA) as described
previously (5). The absorbance was
measured at 450 nm by the micro plate reader (Bio-Rad Laboratories,
Inc., Hercules, CA, USA).
RNA isolation and semi-quantitative
(q)-PCR
Total RNA from culture cells was extracted using
TRIzol reagent (Thermo Fisher Scientific, Inc., Waltham, MA, USA).
Reverse transcription was carried out at 37°C for 120 min with
High-Capacity cDNA Reverse Transcription kit (Thermo Fisher
Scientific, Inc., Waltham, MA, USA). The prepared cDNA was used for
PCR amplification with Taq DNA polymerase (Toyobo Life Science,
Osaka, Japan). The number of PCR cycles for each product was
determined after the confirmation of amplification efficacy. After
the determination of PCR cycles and annealing temperatures, sqPCR
was carried out. The linear primers used in this study and
experimental conditions are as follows: IL-1β,
5′-TGAGCTCGCCAGTGAAATGA-3′ (forward) and 5′-AACACGCAGGACAGGTACAG-3′
(reverse), 24 cycles, 55°C; NLRP3, 5′-GGAGGAGGACTTCGTGCAAA-3′
(forward) and 5′-CCCGGCAAAAACTGGAAGTG-3′ (reverse), 26 cycles,
55°C, and β-actin, 5′-CTTCTACAATGAGCTGCGTG-3′ (forward) and
5′-TCATGAGGTAGTCAGTCAGG-3′ (reverse), 21 cycles, 58°C. The PCR
products were electrophoresed on 2% agarose gels, stained with
ethidium bromide, and visualized with a UV illuminator.
Measurement of NF-κB activity
THP-1 cells were seeded at 3.0×106
cells/dish onto 60 mm dishes and differentiated to macrophages
using PMA as described above. THP-1 macrophage cells were treated
with DHMEQ for 2 h then added A50 for 4 h. The nuclear extracts
were prepared as described before (18) using Nuclear Extract kit (Active
Motif Japan, Tokyo, Japan). The cells were washed with ice-cold PBS
containing phosphatase inhibitors and added to the same solution.
The cells were removed from the dish by scraping and transferred to
a conical tube, then centrifuged for 10 min at 200 × g. The cell
pellets were used as the total cell extracts. They were resuspended
in 500 µl of 1X Hypotonic Buffer and incubated on ice for 15 min.
After the incubation, they were centrifuged for 30 sec at 14,000 ×
g. The pellets were used for the preparation of nuclear fraction.
They were resuspended in 30 µl of 1X Complete Lysis Buffer and
incubated on ice for 30 min on a rocking platform shaker at 150
rpm. After the incubation on ice the lysate was centrifuged for 10
min at 14,000 × g. The resulting supernatant was used for the
nuclear extract.
Then, the DNA binding activity of NF-κB in nuclear
extracts was measured with the TransAM NF-κB p65 Transcription
Factor Assay kit (Active Motif Japan) according to the
manufacturer's instructions.
Measurement of caspase-1 activity
THP-1 cells were seeded at 1.5×106
cells/dish onto 60 mm dishes and differentiated to macrophages
using PMA as described above. THP-1 macrophages were treated with
DHMEQ for 2 h, and then A50 was added. The cells were cultured for
24 h. They were collected by centrifugation at 250 × g for 10 min
after the detachment. The supernatant was removed and discarded,
while the cell pellet was lysed by the addition of 50 µl Lysis
Buffer. The cell lysate was incubated on ice for 10 min and
centrifuged at 10,000 × g for 1 min. The supernatant was used for
the measurement of caspase-1 activity. To evaluate the caspase-1
activity, Caspase-1/ICE Colorimetric Assay Kit (R&D Systems,
Inc., Minneapolis, MN, USA) was used, and it was used as described
in the manufacturer's instructions.
Statistical analysis
All results were presented as the mean ± standard
deviation. The significance of differences among groups was
analyzed by one-way analysis of variance followed by Dunnett's post
hoc test using Excel software v.2013 (Microsoft Corporation,
Redmond, WA, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
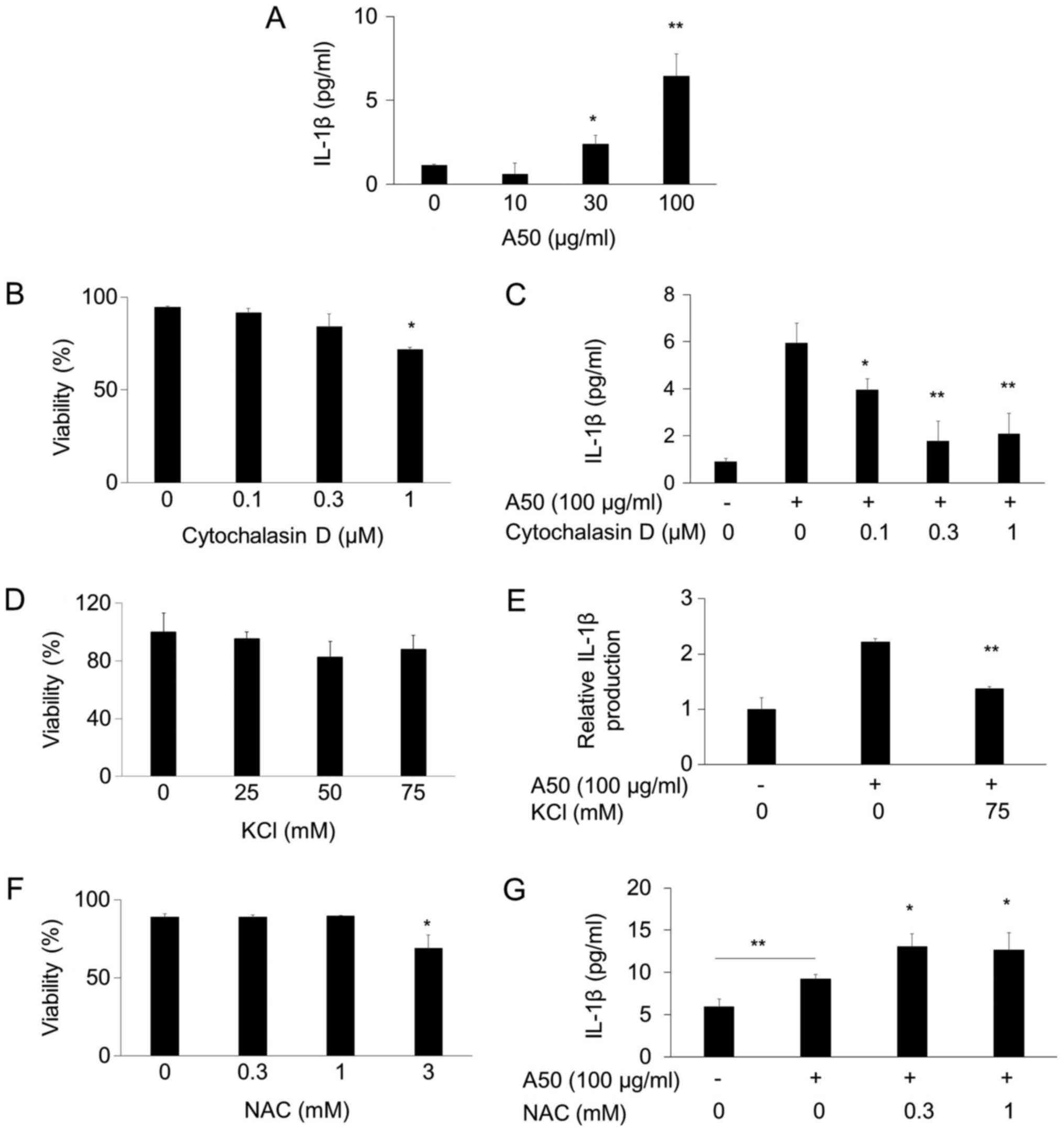
Inhibition of A50-induced IL-1β
production by cytochalasin D and extracellular potassium ion
We evaluated the IL-1β secretion by measuring the
amount in medium using ELISA. As reported before (5), A50 induced IL-1β production in
PMA-treated THP-1 cells (Fig. 1A).
Cytochalasin D was not toxic at 1 µM (Fig. 1B), and inhibited the IL-1β
production at 0.1–1 µM (Fig. 1C).
Thus, it is likely that A50 is incorporated into the cells by
phagocytosis. K+ efflux was reported to be necessary for
IL-1β maturation (19). We found
that extracellular potassium ion inhibited the increase of IL-1β
secretion by A50 (Fig. 1D and E).
Thus, it is likely that K+ efflux is essential for the
A50-induced IL-1β secretion. On the other hand, N-acetylcysteine
did not lower and rather increased the A50-induced IL-1β production
at the nontoxic concentrations (Fig.
1F and G).
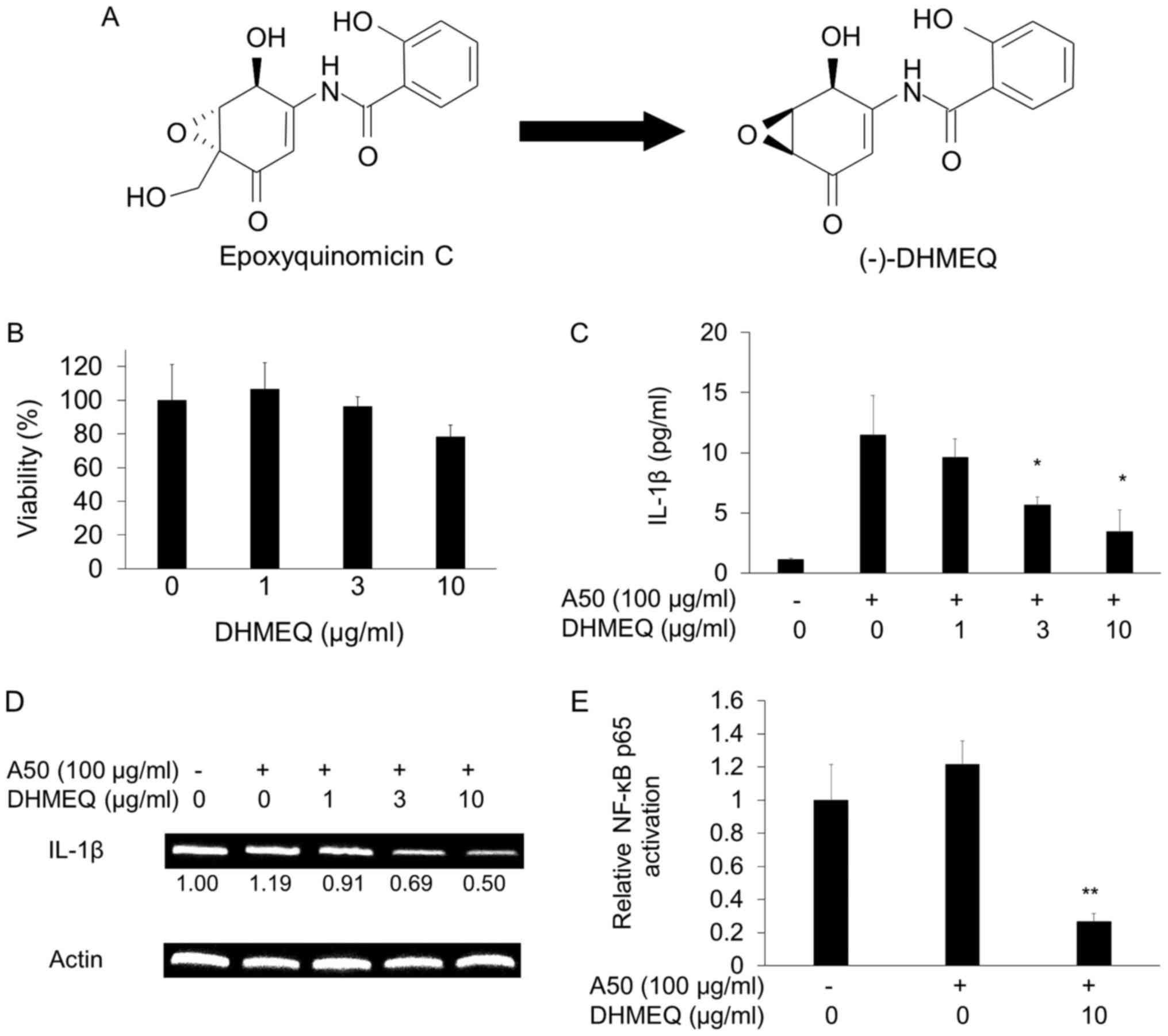
Inhibition of A50-induced IL-1β
production by DHMEQ
DHMEQ (Fig. 2A)
showed no toxicity below 10 µg/ml (Fig. 2B). It was shown to inhibit
A50-induced IL-1β secretion at 3–10 µg/ml (Fig. 2C). Then, we studied the effect of
IL-1β mRNA expression. A50 did not enhance the mRNA expression, but
DHMEQ clearly inhibited the expression (Fig. 2D). Next, we confirmed the
inhibition of NF-κB. As shown in Fig.
2E, A50 did not significantly increase the cellular NF-κB
activity. But DHMEQ clearly lowered the NF-κB activity in 2 h
(Fig. 2E).
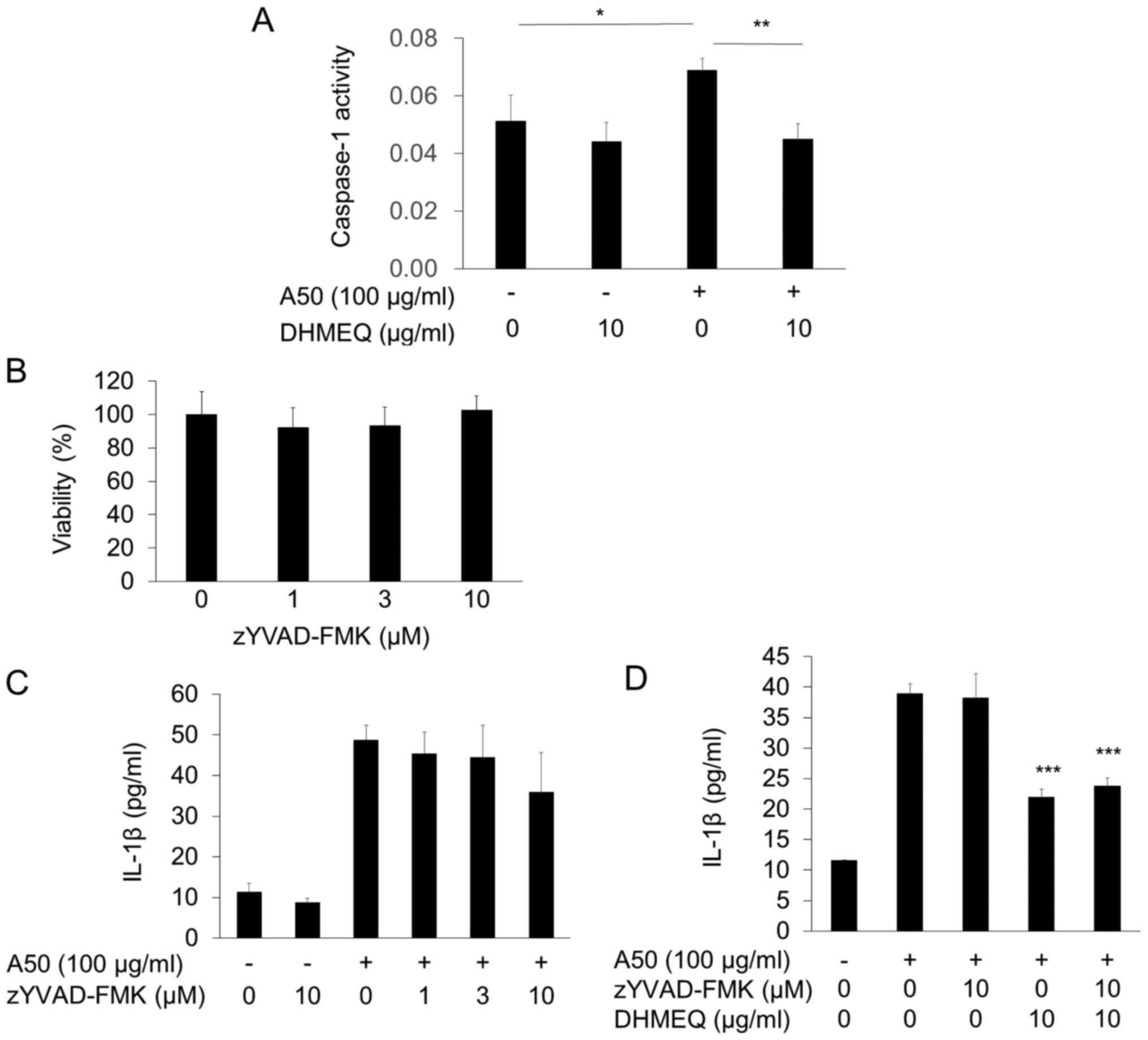
Effect of caspase-1 inhibitor on
A50-induced IL-1β secretion
IL-1β maturation is often activated by
inflammasome/caspase-1 mechanism. We found that cellular caspase-1
activity was significantly increased by A50, which was inhibited by
DHMEQ (Fig. 3A). NLRP3 mRNA
expression was not increased by A50, but inhibited by DHMEQ (data
not shown). To study the functional involvement of caspase-1
decrease, we employed zYVAD-FMK. It did not lower the viability
even at 10 µM (Fig. 3B).
Unexpectedly, the inhibitor did not influence the A50-induced IL-1β
secretion at 1–10 µM (Fig. 3C).
When added with DHMEQ, it did not show any influence again
(Fig. 3D). Thus, it is unlikely
that caspase-1 decrease is involved in the mechanism for inhibition
of IL-1β secretion by DHMEQ. Therefore, inhibition of pro-IL-1β
mRNA transcription by DHMEQ would be sufficient to inhibit
A50-induced K+ efflux-dependent IL-1β secretion
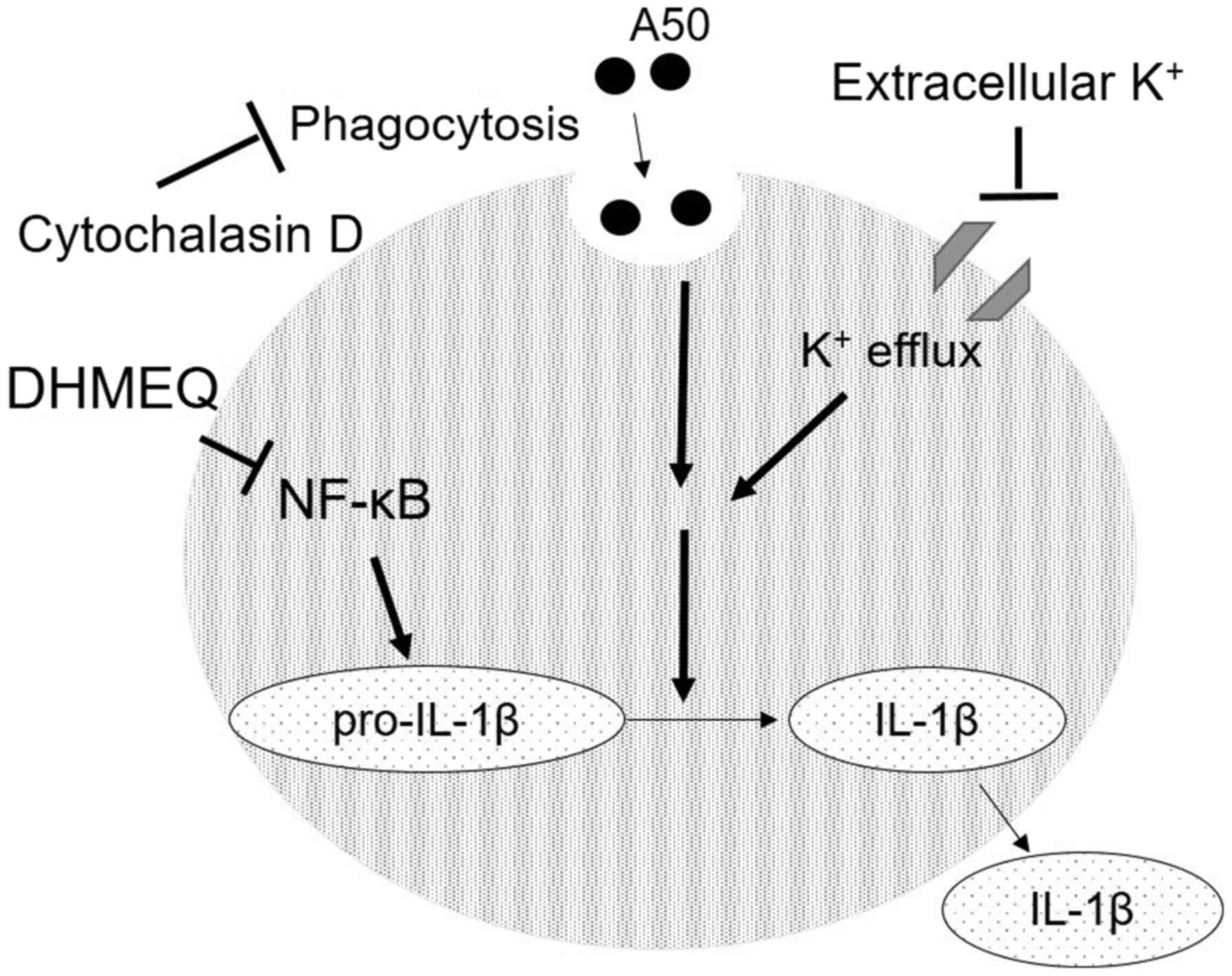
(Fig. 4).
Discussion
The TiO2 model may be too much simplified
as a model of PM2.5. But so far, it is considered to be a suitable
model, since TiO2 nanoparticles have been shown to
induce lung inflammation in mice (20) and IL-1β formation in macrophages
(5).
We found that A50-induced IL-1β secretion was
inhibited by a phagocytosis inhibitor, cytochalasin D.
K+ efflux was also found to be essential for the
efficacy of A50. But clinical use of cytochalasin D and KCl are
known to be difficult. On the other hand, DHMEQ has been widely
used for the suppression of various inflammation and neoplastic
disease models in animals. Therefore, it may be useful for the
treatment of PM2.5-induced inflammation in the trachea and
lungs.
We employed TiO2 nanoparticle of 50 nm
called A50, which is slightly smaller than general microorganisms.
Therefore, it is likely to be incorporated into the mammalian cells
by phagocytosis to stimulate cellular activity. In fact, the
efficacy of A50 was inhibited by cytochalasin D, which itself
inhibits phagocytosis (Fig.
1C).
A50 did not increase the NF-κB activity (Fig. 2D). This may be because the THP-1
cells are treated with PMA for 3 days to obtain THP-1 macrophages.
PMA is known to activate NF-κB (21). However, DHMEQ clearly inhibited the
basal NF-κB activity (Fig. 2D).
Thus, the basal NF-κB activity in THP-1 macrophages should be
necessary for the A50 activity on cytokine production. DHMEQ should
inhibit cellular NF-κB in the nonspecific manner on upstream
signaling, since it was shown to inhibit the cellular NF-κB in
unstimulated adult T-cell leukemia cells (22) and chronic lymphocytic leukemia
cells (23). The nonspecific
inhibition is due to the mechanism that DHMEQ inhibits the final
process of NF-κB activation, which is NF-κB-DNA binding (8,9). The
PMA-treated THP-1 cells possess limitation for the mechanistic
study with DHMEQ, since NF-κB activity is already activated by PMA.
Then, it is important to use PMA-untreated cells. Since intact
THP-1 cells were reported to be nonresponsive to Anatase-type
TiO2 nanoparticles (24), we have tried to use mouse monocytic
leukemia RAW264.7 cells that are LPS-sensitive without
differentiation. However, they were not responsive to A50 on IL-1β
production. The mechanism of A50 downstream signaling has not been
fully elucidated. In future, we should screen A50-sensitive
macrophage-like cells from other sources including primary culture
of macrophages to study the downstream signaling.
We measured secretion of IL-1β by ELISA. The
secretion should depend on the mRNA expression and
post-translational maturation. The former is known to be dependent
on NF-κB. In fact, it was inhibited by DHMEQ, as shown in Fig. 2C. On the other hand,
inflammasome/caspase-1 mechanisms is known to be necessary for the
maturation of IL-1β from its precursor protein (25). This system is a main signaling
pathway in pyroptosis, a programmed cell death (26). One of the main components of
inflammasome is NLRP3. We found that although A50 did not increase
NLRP3 expression, DHMEQ lowered its mRNA expression (data not
shown). Moreover, expression of a downstream component caspase-1
was activated by A50 and inhibited by DHMEQ (Fig. 3A). Inflammasome/caspase-1 axis to
activate IL-1β production is known to be dependent on K+
efflux (19). In fact, we found
that extracellular K+ ion inhibited the A50-induced
IL-1β production (Fig. 1E).
Therefore, we studied the functional involvement of caspase-1 using
its chemical inhibitor zYVAD-FMK. We used this inhibitor at 1–10 mM
as shown in Fig. 3B, because it
was reported to be used at below 10 mM (24). However, zYVAD-FMK did not inhibit
A50-induced IL-1β production with or without DHMEQ. Therefore, it
is unlikely that inhibition of caspase-1 is involved in the
mechanism of inhibition by DHMEQ in our experimental system.
N-acetylcysteine did not inhibit A50-induced IL-1β
production in our study (Fig. 1G),
although Tada-Oikawa et al (5) reported that A50 induced ROS
production in THP-1 macrophages. Moreover, Yazdi et al
(27) demonstrated that the
chemical ROS scavenger diminished IL-1β production triggered by
TiO2 nanoparticles in THP-1 macrophages, suggesting that
IL-1β production is caused by ROS in TiO2
nanoparticle-treated cells. On the other hand, other reports showed
that ROS was not essential for IL-1β production via the Nlrp3
inflammasome (28,29). ROS might not be the main
contributing factor of particle-induced IL-1β production in THP-1
macrophages because the ROS level was increased to some extent
after the exposure to all TiO2 particles (5). Our results indicated that ROS may not
be functionally involved in the A50-induced IL-1β production. In
total, it is still controversial whether ROS is involved in the
mechanism of TiO2 nanoparticle-induced IL-1β
production.
DHMEQ was shown to be nontoxic in animals in many
in vivo experiments. DHMEQ ointment inhibits the atopic
dermatitis model in mice (30,31),
and is now being developed for the treatment of skin inflammation.
Meanwhile it inhibited the LPS-induced sepsis model in animal
experiments by suppression of tumor necrosis factor (TNF)-α
production (32). DHMEQ also
inhibited allergic inflammation and airway remodeling in murine
models of asthma (33). Therefore,
it may be possible that DHMEQ can protect the patient from
PM2.5-caused inflammation in the trachea and lungs.
Acknowledgements
Not applicable.
Funding
The present study was financially supported in part
by JSPS Kakenhi (grant no. 17K01967), the AMED grant (grant no.
JP18fk0310118)), and the Aichi Medical University Research Unit
Fund.
Availability of data and materials
All data generated during the present study are
included in this published article.
Authors' contributions
HF performed the majority of experiments. KU, HF and
TU designed the experiments. KI and AI prepared the A50
(TiO2) suspension. NK, STO, SI, NM, YN and TK
contributed to the design of the methodology. HF and KU analyzed
the data and wrote the paper.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The Department of Molecular Target Medicine is a
fund-donated laboratory. It is supported financially by Shenzhen
Wanhe Pharmaceutical Company (Shenzhen, China), Meiji Seika Kaisha,
Ltd (Tokyo, Japan), Fukuyu Hospital (Nisshin, Japan), and Brunese
Co., Ltd (Nagoya, Japan).
References
|
1
|
Brook RD, Franklin B, Cascio W, Hong Y,
Howard G, Lipsett M, Luepker R, Mittleman M, Samet J, Smith SC Jr,
et al: Air pollution and cardiovascular disease: A statement for
healthcare professionals from the expert panel on population and
prevention science of the American heart association. Circulation.
109:2655–2671. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nel AE, Mädler L, Velegol D, Xia T, Hoek
EM, Somasundaran P, Klaessig F, Castranova V and Thompson M:
Understanding biophysicochemical interactions at the nano-bio
interface. Nat Mater. 8:543–557. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Morimoto Y and Tanaka I: Effects of
nanoparticles on humans. Sangyo Eiseigaku Zasshi. 50:37–48.
2008.(In Japanese). View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Adachi T, Takahara K, Taneo J, Uchiyama Y
and Inaba K: Particle size of latex beads dictates IL-1β production
mechanism. PLoS One. 8:e684992013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tada-Oikawa S, Ichihara G, Fukatsu H,
Shimanuki Y, Tanaka N, Watanabe E, Suzuki Y, Murakami M, Izuoka K,
Chang J, et al: Titanium dioxide particle type and concentration
influence the inflammatory response in Caco-2 cells. Int J Mol Sci.
17:5762016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Matsumoto N, Ariga A, To-e S, Nakamura H,
Agata N, Hirano S, Inoue J and Umezawa K: Synthesis of NF-kappaB
activation inhibitors derived from epoxyquinomicin C. Bioorg Med
Chem Lett. 10:865–869. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ariga A, Namekawa J, Matsumoto N, Inoue J
and Umezawa K: Inhibition of tumor necrosis factor-alpha-induced
nuclear translocation and activation of NF-kappaB by
dehydroxymethylepoxyquinomicin. J Biol Chem. 277:24625–24630. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yamamoto M, Horie R, Takeiri M, Kozawa I
and Umezawa K: Inactivation of NF-kappaB components by covalent
binding of (−)-dehydroxymethylepoxyquinomicin to specific cysteine
residues. J Med Chem. 51:5780–5788. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Takeiri M, Horie K, Takahashi D, Watanabe
M, Horie R, Simizu S and Umezawa K: Involvement of DNA binding
domain in the cellular stability and importin affinity of NF-κB
component RelB. Org Biomol Chem. 10:3053–3059. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shimada C, Ninomiya Y, Suzuki E and
Umezawa K: Efficient cellular uptake of the novel NF-kappaB
inhibitor (−)-DHMEQ and irreversible inhibition of NF-kappaB in
neoplastic cells. Oncol Res. 18:529–535. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kozawa I, Kato K, Teruya T, Suenaga K and
Umezawa K: Unusual intramolecular N->O acyl group migration
occurring during conjugation of (−)-DHMEQ with cysteine. Bioorg Med
Chem Lett. 19:5380–5382. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Umezawa K: Peritoneal NF-κB as a possible
molecular target for suppression of various cancers and
inflammation. ForumImmun Dis Ther. 4:63–77. 2013. View Article : Google Scholar
|
|
13
|
Lin Y, Ukaji T, Koide N and Umezawa K:
Inhibition of late and early phases of cancer metastasis by NF-κB
inhibitor DHMEQ derived from microbial bioactive metabolite
epoxyquinomicin: A review. Int J Mol Sci. 19:7292018. View Article : Google Scholar :
|
|
14
|
Suzuki E and Umezawa K: Inhibition of
macrophage activation and phagocytosis by a novel NF-kappaB
inhibitor, dehydroxymethylepoxyquinomicin. Biomed Pharmacother.
60:578–586. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Suzuki E, Sugiyama C and Umezawa K:
Inhibition of inflammatory mediator secretion by (−)-DHMEQ in mouse
bone marrow-derived macrophages. Biomed Pharmacother. 63:351–358.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sosińska P, Baum E, Maćkowiak B,
Staniszewski R, Jasinski T, Umezawa K and Bręborowicz A: Inhibition
of NF-kappaB with dehydroxyepoxiquinomicin modifies function of
human peritoneal mesothelial cells. Am J Transl Res. 8:5756–5765.
2016.PubMed/NCBI
|
|
17
|
Suzuki Y, Sugiyama C, Ohno O and Umezawa
K: Preparation and biological activities of optically active
dehydroxymethylepoxyquinomicin, a novel NF-κB inhibitor.
Tetrahedron. 60:7061–7066. 2004. View Article : Google Scholar
|
|
18
|
Andrews NC and Faller DV: A rapid
micropreparation technique for extraction of DNA-binding proteins
from limiting numbers of mammalian cells. Nucleic Acids Res.
19:24991991. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pétrilli V, Papin S, Dostert C, Mayor A,
Martinon F and Tschopp J: Activation of the NALP3 inflammasome is
triggered by low intracellular potassium concentration. Cell Death
Differ. 14:1583–1589. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hussain S, Vanoirbeek JA, Luyts K, De
Vooght V, Verbeken E, Thomassen LC, Martens JA, Dinsdale D, Boland
S, Marano F, et al: Lung exposure to nanoparticles modulates an
asthmatic response in a mouse model. Eur Respir J. 37:299–309.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bomsztyk K, Rooney WJ, Iwasaki T, Rachie
AN, Dower KS and Sibley HC: Evidence that interleukin-1 and phorbol
esters activate NF-kappaB by different pathways: Role of protein
kinase C. Cell Regul. 2:329–335. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Watanabe M, Ohsugi T, Shoda M, Ishida T,
Aizawa S, Maruyama-Nagai M, Utsunomiya A, Koga S, Yamada Y,
Kamihira S, et al: Dual targeting of transformed and untransformed
HTLV-1-infected T-cells by DHMEQ, a potent and selective inhibitor
of NF-kappaB, as a strategy for chemoprevention and therapy of
adult T cell leukemia. Blood. 106:2462–2471. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Horie R, Watanabe M, Okamura T, Taira M,
Shoda M, Motoji T, Utsunomiya A, Watanabe T, Higashihara M and
Umezawa K: DHMEQ, a new NF-kappaB inhibitor, induces apoptosis and
enhances fludarabine effects on chronic lymphocytic leukemia cells.
Leukemia. 20:800–806. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Morishige T, Yoshioka Y, Tanabe A, Yao X,
Tsunoda S, Tsutsumi Y, Mukai Y, Okada N and Nakagawa S: Titanium
dioxide induces different levels of IL-1beta production dependent
on its particle characteristics through caspase-1 activation
mediated by reactive oxygen species and cathepsin B. Biochem
Biophys Res Commun. 392:160–165. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Youm YH, Nguyen KY, Grant RW, Goldberg EL,
Bodogai M, Kim D, D'Agostino D, Planavsky N, Lupfer C, Kanneganti
TD, et al: The ketone metabolite β-hydroxybutyrate blocks NLRP3
inflammasome-mediated inflammatory disease. Nat Med. 21:263–269.
2015. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Fink SL and Cookson BT: Apoptosis,
pyroptosis, and necrosis: Mechanistic description of dead and dying
eukaryotic cells. Infect Immun. 73:1907–1916. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yazdi AS, Guarda G, Riteau N, Drexler SK,
Tardivel A, Couillin I and Tschopp J: Nanoparticles activate the
NLR pyrin domain containing 3 (Nlrp3) inflammasome and cause
pulmonary inflammation through release of IL-1α and IL-1β. Proc
Natl Acad Sci USA. 107:19449–19454. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Van de Veerdonk LF, Smeekens PS, Joosten
BAL, Kullberg JB, Dinarello AC, van der Meer MWJ and Netea GM:
Reactive oxygen species-independent activation of the IL-1β
inflammasome in cells from patients with chronic granulomatous
disease. Proc Natl Acad Sci USA. 107:3030–3033. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Iyer SS, He Q, Janczy JR, Elliott EI,
Zhong Z, Olivier AK, Sadler JJ, Knepper-Adrian V, Han R, Qiao L, et
al: Mitochondrial cardiolipin is required for Nlrp3 inflammasome
activation. Immunity. 39:311–323. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hamasaka A, Yoshioka N, Abe R, Kishino S,
Umezawa K, Ozaki M, Todo S and Shimizu H: Topical application of
dehydroxymethylepoxyquinomicinimproves allergic inflammation via
NF-kappaB inhibition. J Allergy Clin Immunol. 126:400–403. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jiang X, Lan Y, Wei B, Dai C, Gu Y, Ma J,
Liu X, Umezawa K and Zhang Y: External application of NF-κB
inhibitor DHMEQ suppresses development of atopic dermatitis-like
lesions induced with DNCB/OX in BALB/c mice. Immunopharmacol
Immunotoxicol. 39:157–164. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Shimo T, Adachi Y, Umezawa K, Okigaki M,
Takaya J, Taniuchi S, Ikehara S and Kaneko K:
Dehydroxymethylepoxyquinomicin (DHMEQ) can suppress tumour necrosis
factor-α production in lipopolysaccharide-injected mice, resulting
in rescuing mice from death in vivo. Clin Exp Immunol. 166:299–306.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shimizu K, Konno S, Ozaki M, Umezawa K,
Yamashita K, Todo S and Nishimura M: Dehydroxymethylepoxyquinomicin
(DHMEQ), a novel NF-kappa B inhibitor, inhibits allergic
inflammation and airway remodelling in murine models of asthma.
Clin Exp Allergy. 42:1273–1281. 2012. View Article : Google Scholar : PubMed/NCBI
|