Introduction
Infertility, a worldwide reproductive health
problem, affects 10–15% of couples (1). In particular, more than one-half of
the cases are due to male infertility, and 60–75% male infertility
cases are idiopathic, which is frequently the most difficult form
of infertility to treat (2). Male
infertility may result from a number of factors, including
azoospermia, oligospermia, asthenospermia, orchitis and varicoceles
(3). The pathogenesis of male
infertility is usually derived from genetic and environmental
factors. In terms of environmental factors, previous studies
suggested that chronic hypoxia induces male infertility (4,5). A
number of previous studies further suggested that hypoxia may
induce the apoptosis of spermatogenic cells and spermatogenesis in
mice and hypoxia-induced rat models of male infertility (6–8).
However, the underlying molecular mechanisms of this effect remain
to be elucidated.
MicroRNAs (miRNAs), a family of small non-coding
RNAs (~22 nt), serve an important role in mediating
post-transcriptional gene silencing by sequence-selective targeting
of mRNAs (9). It was demonstrated
that miRNAs are important regulators of cell growth,
differentiation, apoptosis, metabolism and tumorigenesis (10). Previously, numerous miRNAs have
been identified to be exclusively or preferentially expressed in
mice testes, suggesting the important role of miRNAs in
translational repression during spermatogenesis (11,12).
Previous studies demonstrated that certain miRNAs may be regulated
by hypoxia (6,13), and miRNA (miR)-210 is the most
induced miRNA under hypoxia of all the hypoxia-induced miRNAs
(14).
miR-210 targets, including E2F transcription factor
3, autophagy-related protein 7, iron-sulfur cluster scaffold
protein and Kruppel-like factor 7 (KLF7), have effects on cell
proliferation, autophagy, adenosine triphosphate metabolism and
angiogenesis (15–19). Among the miR-210 targets, KLF7 is a
member of the Kruppel-like factors (KLFs) family and, due to its
wide expression in a number of adult human tissues, is additionally
termed ubiquitous KLF (20). In
the KLFs family, 17 members (KLF1-KLF17) have been identified in
mammals, exerting effects on cell proliferation, differentiation
and apoptosis (21). It was
additionally demonstrated that chicken KLF7 inhibits preadipocyte
differentiation and promotes preadipocyte proliferation (22). However, the function of KLF7 in
hypoxia-induced apoptosis of GC-2spd (GC-2) cells has not been
fully elucidated. The present study investigated whether and how
miR-210 contributes to the apoptosis of spermatocytes by targeting
KLF7.
Materials and methods
Cell treatment and transfection
GC-2 cells (a mouse pachytene spermatocyte-derived
immortalized cell line) were obtained from the American Type
Culture Collection (Manassas, VA, USA) and cultured in RPMI 1640
medium (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) supplemented with 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.) and antibiotics (100 U/ml penicillin and 100 U/ml
streptomycin). The cells were subsequently subjected to hypoxia (1%
O2, 5% CO2, 10% H2 and 85%
N2) at 37°C in a hypoxia workstation
(InvivO2; The Baker Company, Inc., Sanford, ME, USA) for
12, 24, 48 or 72 h or incubated in normoxia (21% O2 and
5% CO2) as a control. RNA and protein isolation,
terminal deoxynucleotidyl-transferase-meditated dUTP nick end
labeling (TUNEL) staining and flow cytometry analysis were
performed on the GC-2 cells at all the indicated time points.
GC-2 cells were cultured until they reached 80%
confluence in six-well dishes, washed once with PBS and
subsequently transfected with small interfering (si)RNA with
sequences as follows: sense, 5′-GGGCCAUAUUCAUGUCUAUUU-3′ and
antisense, 5′-AUAGACAUGAAUAUGGCCCUU-3′ for mouse hypoxia-inducible
factor-1α (HIF-1α); sense, 5′-UUCUCCGAACGUGUCACGUTT-3′ and
antisense, 5′-ACGUGACACGUUCGGAGAATT-3′ as a negative control (NC;
all purchased from Guangzhou RiboBio Co., Ltd., Guangzhou, China).
siRNA was transfected at a final concentration of 100 nM. In
addition, full-length KLF7 cDNA fragments (20 ng; Guangzhou RiboBio
Co., Ltd.) were cloned into the pcDNA3.1-Myc-His vector between the
Kpn1 and Not1 restriction sites (Invitrogen; Thermo
Fisher Scientific, Inc.), generating pcDNA3.1-KLF7. The empty
pcDNA3.1 vector was used as the control. The GC2 cells were
cultured in the hypoxia workstation for 48 h or were transfected
with miR-210 mimics (0.4 nM) or inhibitors (0.4 nM), as well as
pcDNA3.1-KLF7 (50 nM) using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. The sequence of the miRNA-210 mimic was:
5′-CUGUGCGUGUGACAGCGGCUGA-3′. The sequence of the NC miRNA mimic
was: 5′-UUCUCCGAACGUGUCACGU-3′. The sequence of the anti-miR-210
inhibitor was: 5′-UCAGCCGCUGUCACACGCACAG-3′. The sequence of the NC
inhibitor was: 5′-CAGUACUUUUGUGUAGUACAA-3′. Subsequently, the cells
were cultured in the hypoxia workstation for 12, 24, 48 and 72 h.
Experiments were routinely performed in triplicate wells and
repeated three times. TUNEL assay. TUNEL assays were
performed to detect the apoptosis of GC-2 cells using TUNEL
fluorescent kit (Wuhan Boster Biological Technology, Ltd., Wuhan,
China) according to the manufacturer's protocol. Cells, subjected
to hypoxia or normoxia, and then fixed using 4% paraformaldehyde
for 1 h at 15–25°C. Cells were then permeabilized using 0.1% Triton
X-100 for 2 min on ice (2–8°C), followed by fluorescein
isothiocyanate (FITC)-labeled TUNEL staining (Roche Diagnostics
GmbH, Mannheim, Germany) for 1 h at 37°C. Following this, cells
were counterstained with 1 µg/ml 4′,6-diamidino-2-phenylindole
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) and subsequently
mounted in ProLong™ Gold Antifade Mountant (Invitrogen;
Thermo Fisher Scientific, Inc.). TUNEL-positive cells were imaged
under a fluorescence microscope and imaging system (AF6000; Leica
Microsystems GmbH, Wetzlar, Germany), and five fields from each
image were quantified (the number of green spots).
Apoptosis assays by flow
cytometry
Cells (1×106 cells/ml) were subjected to
hypoxia as above, washed twice with ice-cold PBS, and subsequently
stained with an Annexin V-FITC apoptosis detection kit (BD
Pharmingen; BD Biosciences, San Jose, CA, USA) according to the
manufacturer's protocol. The apoptosis incidence rate was analyzed
using a flow cytometer (FACSCalibur; BD Biosciences) within 1 h of
staining. Apoptotic cells were counted and presented as a
percentage of the total cell count for each sample. The percentage
of FITC-positive cells was analyzed using BD CellQuest™
software version 6.0 (BD Biosciences).
Luciferase assay
The putative miR-210-binding site in the 3′
untranslated region (UTR) of the KLF7-mRNA was identified using the
TargetScan database (www.targetscan.org). To construct a KLF7 3′UTR
luciferase reporter plasmid, the KLF7 3′UTR was amplified from
mouse genomic DNA. The 3′-UTR of KLF7, containing the predicted
wild-type (Wt) or mutated (Mut) binding sites of miR-210 were
amplified using mouse genomic DNA via PCR using DreamTaq DNA
Polymerase (Thermo Fisher Scientific, Inc.). The primers used were
as follows: KLF7-Wt 3′UTR construct primer forward,
5′-GCAGCCAATGTCCGAAGGA-3′, and reverse, 5′-GAGGACCCAATAAACAGG-3′;
KLF7-Mut primer forward,
5′-CCTCTGTGTGCATACATGTACACGCACACGTACACACACCCTCTCAC-3′, and reverse,
5′-GTGAGAGGGTGTGTGTACGTGTGCGTGTACATGTATGCACACAGAGG-3′. The
thermocycling conditions used were as follows: Initial denaturation
at 95°C for 2 min; followed by 40 cycles of denaturation at 95°C
for 30 sec, annealing at 60°C for 30 sec, extension at 72°C for 1
min and a final extension at 72°C for 5 min. The resulting purified
polymerase chain reaction (PCR) products were subsequently cloned
into the downstream region of the pGL3vector (Promega Corporation,
Madison, WI, USA). Mutant constructs with 6–7 mutated residues in
the predicted binding site were generated by site-directed
mutagenesis. Subsequently, 293T cells (Shanghai Aiyan Biological
Technology Co., Ltd., Shanghai, China) were cotransfected with the
pGL3 vectors containing the Wt or Mut 3′UTR luciferase reporter of
KLF7, a miR-210 mimics or a control using Lipofectamine®
2000, in addition to Renilla luciferase (pRL-TK Vector; Promega
Corporation) as a transfection efficiency control. After 48 h, the
luciferase signal was analyzed using the Dual-Luciferase Reporter
Assay kit (Promega Corporation) according to the manufacturer's
protocol. The results are presented as the relative luciferase
activity (firefly luciferase/Renilla luciferase). Each experiment
was repeated three times.
RNA isolation and reverse
transcription-quantitative (RT-q)PCR
miRNA was extracted from treated GC-2 cells using
the miRVana miRNA Isolation kit (Ambion; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. cDNA was
synthesized from 1 µg total RNA using a miRNA RT kit (Applied
Biosystems; Thermo Fisher Scientific, Inc.) at 37°C for 15 min and
85°C for 5 sec. The miR-210 Bulge-Loop TMmiR-210-3p RT-qPCR Primer
Set (Guangzhou RiboBio Co., Ltd.) was used: miR-210 forward primer,
5′-CTGTGCGTGT-3′ and miR-210 reverse primer,
5′-CATGATCAGCTGGGCCAAGATCAGCCG-3′; U6 forward primer,
5′-CTCGCTTCGGCAGCACA-3′ and U6 reverse primer,
5′-AACGCTTCACGAATTTGCGT-3′. The PCR protocol consisted of an
initial denaturation step at 95°C for 10 min, followed by 45 cycles
of denaturation at 95°C for 10 sec and primer annealing/extension
at 60°C for 60 sec. qPCR was performed using 1 µg cDNA and
SYBR-Green (Bio-Rad Laboratories, Inc., Hercules, CA, USA) in a
LightCycler rapid thermal cycler system (Roche Applied Science,
Penzberg, Germany). The data were standardized to U6. Relative
quantification of the target gene was conducted using the
comparative Cq (2−∆∆Cq) method (23).
Western blot analysis
Following treatment, proteins were lysed from GC-2
cells in ice-cold radioimmunoprecipitation assay lysis buffer
(Beyotime Institute of Biotechnology, Haimen, China), supplemented
with protease inhibitors for 1 h to extract total cell protein.
Following this, protein concentrations were determined using a
bicinchoninic acid kit (Thermo Fisher Scientific, Inc.). Equivalent
amounts of protein (20 µg) were separated using 10% SDS-PAGE and
electrotransferred onto a nitrocellulose membrane (Bio-Rad
Laboratories, Inc.). The membrane was blocked with 5% non-fat dry
milk in TBS containing 0.1% Tween (TBST) for 1 h at room
temperature. Membranes were subsequently incubated at 4°C overnight
with rabbit anti-caspase 3 antibody (1:1,000; cat. no. 9664),
rabbit anti-apoptosis regulator BAX (Bax) antibody (1:500; cat. no.
14796), rabbit anti-B-cell lymphoma 2 (Bcl-2; antibody (1:1,000;
cat. no. 3498; all Cell Signaling Technology, Inc., Danvers, MA,
USA), rabbit anti-HIF-1α antibody (1:1,000; cat. no. sc-10790;
Santa Cruz Biotechnology, Inc., Dallas, TX, USA), rabbit anti-KLF7
antibody (1:500; cat. no. ab197690; Abcam, Cambridge, UK) or rabbit
anti-β-actin antibody (1:5,000; cat. no. SAB5500001; Sigma-Aldrich;
Merck KGaA). The membranes were washed with TBST and incubated with
goat anti-rabbit horseradish peroxidase-conjugated secondary
antibodies (1:5,000; cat. no. P044801-2; Dako; Agilent
Technologies, Inc., Santa Clara, CA, USA) at room temperature for 1
h. Immunoblotted bands were visualized using an enhanced
chemiluminescence kit (GE Healthcare Life Sciences, Little
Chalfont, UK). The ChemiDoc Imaging System (Bio-Rad Laboratories,
Inc.) and Quantity One™ 1-D Analysis software version
4.6.7 (Bio-Rad Laboratories, Inc.) were used to scan the blots and
analyze the density of the blots.
Statistical analysis
Data are presented as the mean ± standard deviation,
and each experiment was repeated at least three times. Statistical
analyses were conducted using SPSS software version 19.0 (IBM
Corp., Armonk, NY, USA). Statistical significance was evaluated
using an unpaired two-tailed Student's t-test or by one-way
analysis of variance followed by Dunnett's test or the Least
Significant Difference test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Hypoxia treatment induces GC-2 cell
apoptosis
The effects of different hypoxia time periods on
GC-2 cell apoptosis were investigated using TUNEL and flow
cytometry assays (Fig. 1). The
apoptosis at 12, 24, 48 and 72 h of hypoxia are presented in
Fig. 1A and C. No difference was
observed in the number of apoptotic cells between the hypoxia group
at 12 h and the normal group; however, a significant increase in
cell apoptosis occurred after 24 h of hypoxia (P<0.01). The
results of the flow cytometry assay were consistent with the
results of TUNEL staining (Fig. 1B and
D). Hypoxia induced a significant increase in the miR-210
expression level compared with the normoxia group (Fig. 1E; P<0.01). Furthermore, the
HIF-1α protein expression levels were significantly increased after
12 h of hypoxia (Fig. 1F;
P<0.01).
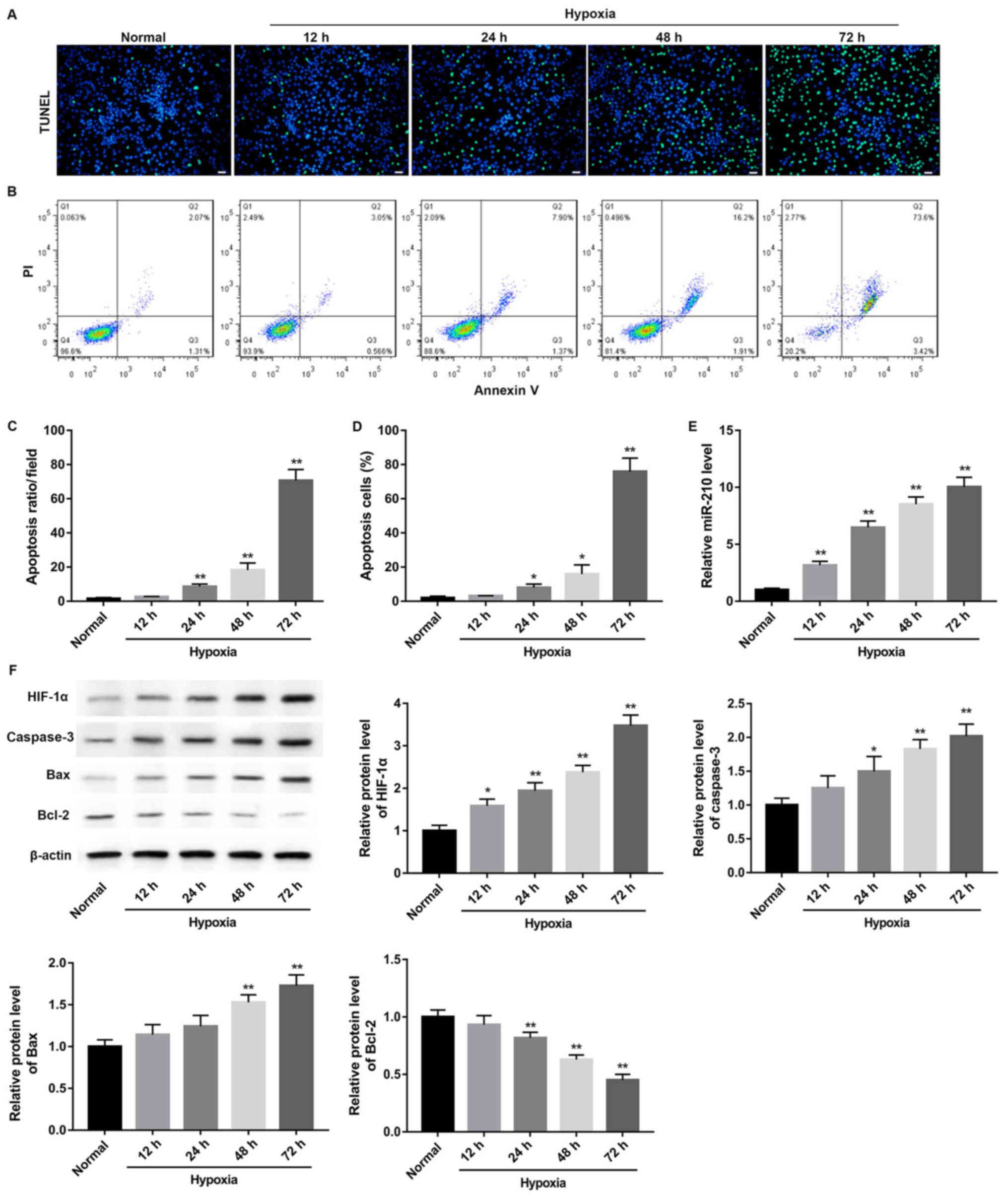 | Figure 1.Hypoxia induces apoptosis of GC-2
cells at different time points. (A) TUNEL staining results of GC-2
cells. Scale bar, 20 µm. (B) Representative graphs of flow
cytometry analysis. (C) Apoptosis rate of GC-2 cells was evaluated
by TUNEL staining. (D) Apoptotic GC-2 cells were measured by flow
cytometry. (E) Reverse transcription-quantitative polymerase chain
reaction analysis of miR-210 expression in mouse GC-2 cells
subjected to hypoxia for 12, 24, 48 and 72 h. (F) Western blot
analysis for HIF-1α, caspase-3, Bax and Bcl-2 protein expression in
mouse GC-2 cells subjected to hypoxia for 12, 24, 48 and 72 h.
*P<0.05, **P<0.01 vs. respective normal. miR, microRNA;
HIF-1α, hypoxia-inducible factor-1α; Bax, apoptosis regulator BAX;
Bcl-2, B-cell lymphoma 2; TUNEL, terminal
deoxynucleotidyl-transferase-meditated dUTP nick end labeling and
flow cytometry; PI, propidium iodide; GC-2, GC-2spd. |
Subsequently, the effect of different time periods
of hypoxia on the apoptosis pathway in GC-2 cells was determined by
measuring apoptosis-associated protein expression levels at 12, 24,
48 and 72 h following hypoxia. The results demonstrated that the
pro-apoptotic proteins caspase-3 and Bax were significantly
increased in the hypoxia group compared with the normoxia group in
GC-2 cells after 48 h (Fig. 1F;
P<0.01). However, the anti-apoptotic protein Bcl-2 was
significantly decreased after 24 h of hypoxia (Fig. 1F; P<0.01).
Hypoxia-induced HIF-1α mediates
apoptosis of GC-2 cells
A previous study suggested that HIF-1α is involved
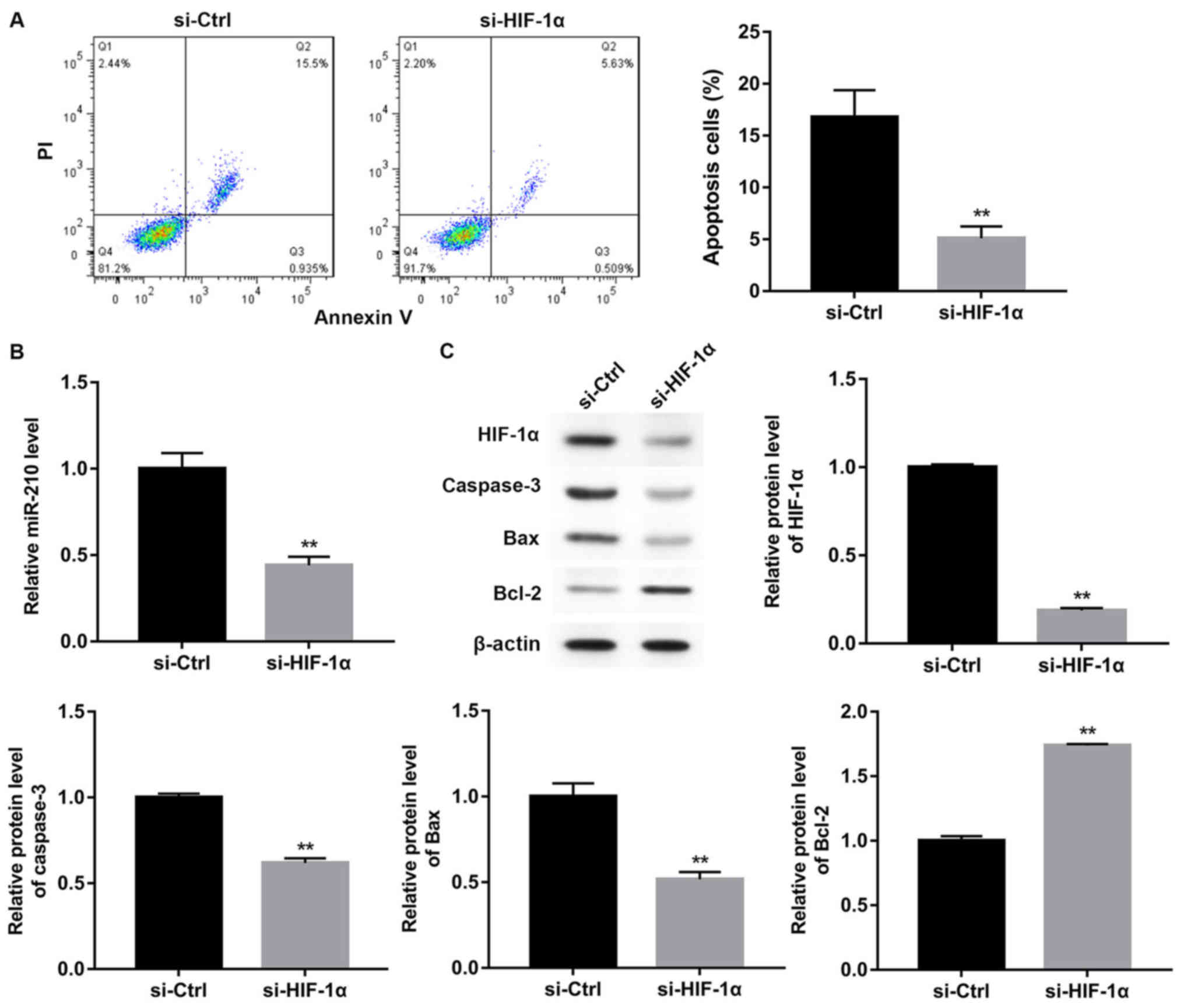
in mouse spermatocyte apoptosis during hypoxia (6). Therefore, whether HIF-1α silencing
affected apoptosis in GC-2 cells and whether the alteration of
miR-210 expression was induced by hypoxia was additionally
examined. The results demonstrated that the apoptosis of GC-2 cells
treated with HIF-1α silencing subjected to hypoxia for 48 h was
significantly decreased compared with the null vector group
(Fig. 2A; P<0.01).
Additionally, interference with HIF-1α-siRNA resulted in a
significant decrease in the expression of miR-210 (Fig. 2B; P<0.01), HIF-1α, caspase-3 and
Bax, and a significant increase in the expression of Bcl-2
(Fig. 2C; P<0.01).
Upregulation of miR-210 induces
apoptosis of GC-2 cells
A previous study suggested that miR-210 is
upregulated in the testes of patients with non-obstructive
azoospermia (24). In addition, it
was demonstrated that miR-210 serves an important role in cell
adaptation to hypoxia by regulating a HIF-1-dependent pathway
(25). Therefore, it was
hypothesized that alteration of miR-210 may affect the apoptosis
rate of GC-2 cells under hypoxic conditions. Therefore, the role of
miR-210 in the apoptosis of hypoxia-induced cells was examined. As
presented in Fig. 3A, transfection
of GC-2 cells with miR-210 mimics or a miR-210 inhibitor had high
transfection efficiency. The number of apoptotic GC-2 cells
transfected with the miR-210 mimics was significantly increased
compared with the mimic NC group under hypoxic conditions (Fig. 3B; P<0.01). Conversely, the
apoptosis of GC-2 cells was significantly decreased in the miR-210
inhibitor group (Fig. 3B;
P<0.01). Overexpression of miR-210 resulted in a significant
increase in the expression of caspase-3 and Bax, and a significant
decrease in the expression of Bcl-2 (Fig. 3C; P<0.01). Converse results were
observed for the suppression of miR-210 (Fig. 3C), suggesting that miR-210 is
involved in the apoptotic pathway of GC-2 cells following
hypoxia.
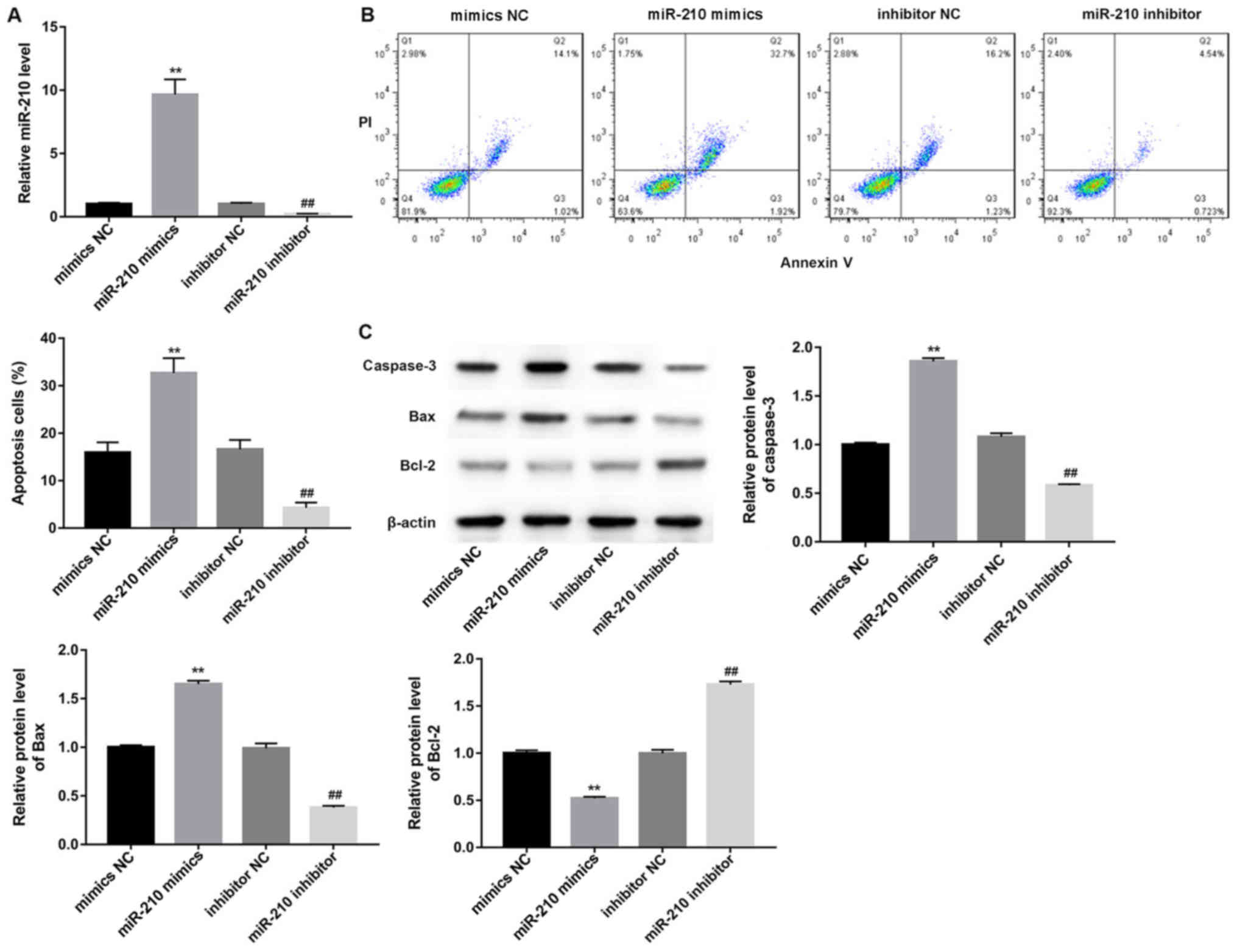 | Figure 3.Overexpression of miR-210 induces
apoptosis in GC-2 cells. (A) Measurement of transfection
efficiency. (B) Analysis of the rate of apoptotic cells transfected
with miR-210 mimics, mimics NC, miR-210 inhibitor and inhibitor NC
following hypoxic culture. (C) Analysis of apoptosis-associated
protein expression in GC-2 cells transfected with miR-210 mimics,
mimics NC, miR-210 inhibitor and inhibitor NC. **P<0.01 vs.
respective mimics NC; ##P<0.01 vs. respective
inhibitor NC. miR, microRNA; NC, negative control; PI, propidium
iodide; Bax, apoptosis regulator BAX; Bcl-2, B-cell lymphoma 2;
GC-2, GC-2spd. |
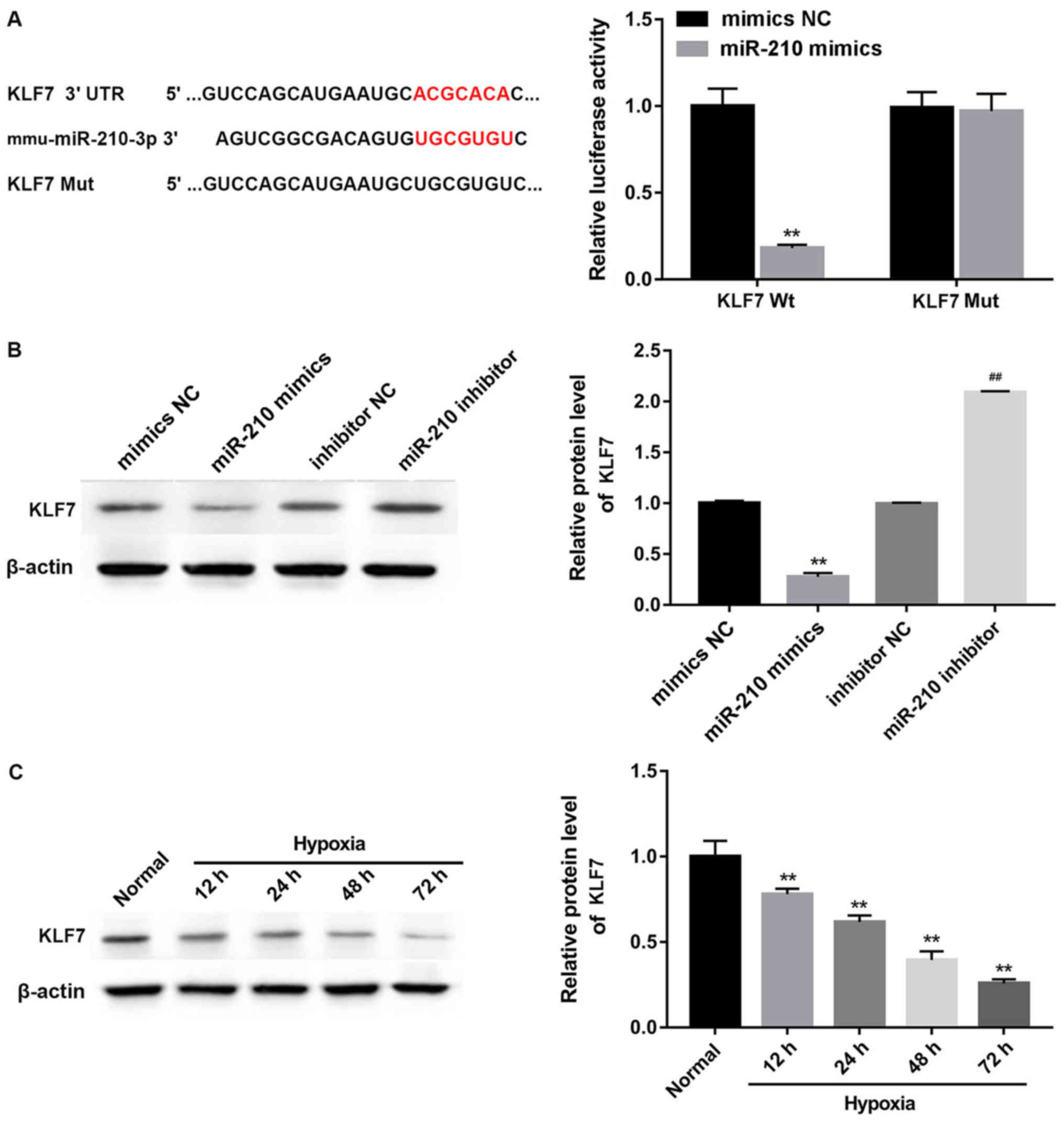
KLF7 is directly targeted by
miR-210
As the 3′UTR of the KLF7-mRNA has a putative
miR-210-binding site, identified using the TargetScan database
(www.targetscan.org), KLF7 was predicted
as a potential target of miR-210. To identify whether the KLF7 gene
was targeted by miR-210 directly in GC-2 cells, luciferase reporter
assays were performed. As presented in Fig. 4A, there was a significant decrease
in luciferase activity following cotransfection of GC-2 cells in
the KLF7 Wt group with the miR-210 mimics and the luciferase
reporter gene (P<0.01); however, the KLF7 Mut demonstrated no
difference in luciferase activity between the miR-210 mimics and
the mimics NC groups. Overexpression of miR-210 with the mimics
significantly decreased the expression of KLF7 (P<0.01);
however, treatment with an inhibitor of miR-210 significantly
increased KLF7 expression (Fig.
4B; P<0.01). In addition, the expression of KLF7 protein in
hypoxic GC-2 cells at each time point was significantly decreased
compared with the normal cells (Fig.
4C; P<0.01), suggesting that hypoxia significantly decreased
KLF7 protein expression in GC-2 cells.
miR-210 affects apoptosis in GC-2
cells by targeting KLF7
To further determine whether the altered miR-210 and
KLF7 expression levels are associated with GC-2 cell apoptosis, the
apoptosis of GC-2 cells transfected with the miR-210 mimics and/or
KLF7 mimics was assessed by flow cytometry. Under hypoxic
conditions, overexpression of miR-210 significantly increased
apoptosis, whereas cotransfection of the miR-210 mimics and the
KLF7 mimics in GC-2 cells significantly decreased the effects of
the miR-210 mimics on cell apoptosis (Fig. 5A; P<0.05). In addition, it was
identified that KLF7 overexpression had no effect on miR-210
compared with transfection of miR-210 mimics alone (Fig. 5B). The effect of hypoxia-induced
miR-210 on apoptosis-associated protein expression was detected by
targeting KLF7. The results demonstrated that the protein
expression of caspase-3 and Bax was significantly higher in the
GC-2 cells transfected with the miR-210 mimics compared with the
control cells (Fig. 5C;
P<0.01); however, upregulation of KLF7 with the KLF7 mimics
cotransfected with miR-210 mimics decreased the effects of the
miR-210 mimics on caspase-3 and Bax expression. Converse results
were observed for Bcl-2 and KLF7 expression (Fig. 5C).
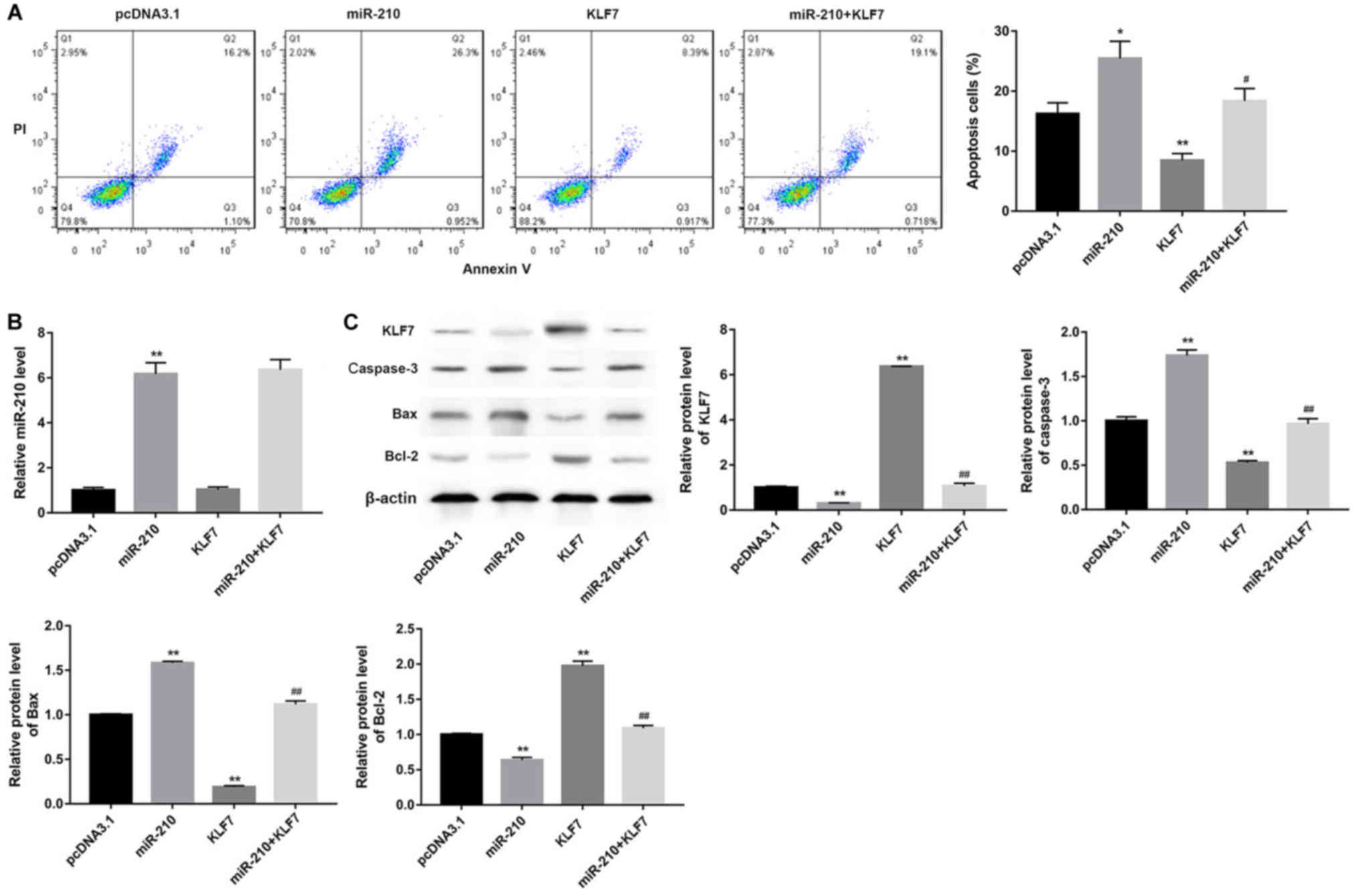 | Figure 5.miR-210 promotes apoptosis of GC-2
cell by targeting KLF7. GC-2 cells were transfected with pcDNA3.1,
miR-210 mimics, KLF7 mimics and miR-210 mimics + KLF7 mimics
following hypoxic for 48 h. (A) Analysis of apoptotic cells. (B)
Analysis of miR-210 expression. (C) Analysis of
apoptosis-associated protein expression. *P<0.05, **P<0.01
vs. respective pcDNA3.1; #P<0.05,
##P<0.01 vs. miR-210. miR, microRNA; KLF7,
Kruppel-like factor 7; PI, propidium iodide; Bax, apoptosis
regulator BAX; Bcl-2, B-cell lymphoma 2; GC-2, GC-2spd. |
Discussion
A number of previous studies suggested that hypoxia
is a signature of the tumor microenvironment and contributes to
proliferation of various types of cancer cells in humans (26–28).
However, previous studies on non-cancer cells demonstrated that
hypoxia may induce the apoptosis of cardiomyocytes, endothelial
cells, astrocytes and muscle cells (29–32).
Notably, hypoxia may induce the apoptosis of spermatogenic cells in
mice (6). Based on these
observations, a hypoxic model of GC-2 cells was established by
subjecting cells to hypoxia for different lengths of time. The
results of the present study demonstrated that hypoxia induced
apoptosis of GC-2 cells and promoted the expression of
pro-apoptotic proteins caspase-3 and Bax, while inhibiting the
expression of the anti-apoptotic protein Bcl-2 after 48 h of
hypoxia. The apoptosis of GC-2 cells additionally increased in a
time-dependent manner. These results suggested that the effect of
hypoxia on apoptosis-associated protein expression induced
apoptosis in GC-2 cells, which may result in the obstruction of
spermatogenesis.
HIF-1α serves a key role in the cellular adaptation
to hypoxia and ischemia (33). A
previous study demonstrated that HIF-1α is involved in the
regulation of hypoxia-induced apoptosis (34). Furthermore, it was observed the
inhibition of HIF-1α protected cells against apoptosis induced by
ischemia and hypoxia (35).
Previous studies suggested that miR-210 regulates response to
hypoxia in a HIF-dependent way (25,36,37).
In the GC-2 cell hypoxia model, it was identified that HIF-1α
protein expression levels significantly increased after 12 h of
hypoxia. Therefore, considering the effect of hypoxia on GC-2 cell
apoptosis and HIF-1α expression, in addition to the association
between HIF-1α and miR-210 under hypoxic condition, the present
study attempted to determine whether the silencing of HIF-1α
affected the apoptosis of GC-2 cells and the alteration of miR-210
expression induced by hypoxia.
The apoptosis of GC-2 cells with HIF-1α silencing
demonstrated a significant decrease in apoptosis compared with the
null vector group. Additionally, interference of HIF-1α by siRNA
resulted in a significant decrease in the miR-210, caspase-3 and
Bax expression levels and a significant increase in Bcl-2
expression. In addition, the apoptosis rate of GC-2 cells and the
expression level of pro-apoptotic proteins in the miR-210 mimics
group were significantly increased compared with the mimics NC
group under hypoxic culture conditions; however, the expression
level of anti-apoptotic Bcl-2 was significantly decreased. However,
data for depletion of miR-210 were in contrast with the results
demonstrating overexpression of miR-210. These results suggested
that hypoxia-induced HIF-1α and miR-210 were involved in the
apoptotic pathway of GC-2 cells.
KLFs regulate the expression of a number of genes in
a variety of cellular processes during embryonic development and in
adult cells (38). A previous
study on KLF7-null mice suggested that KLF7 is a key factor in the
development of the nervous system (39). Previous functional studies
demonstrated that KLF7 inhibits preadipocyte differentiation and
promotes pre-adipocyte proliferation (22,40).
In the present study, the luciferase reporter gene assay
demonstrated that KLF7 is a potential target of miR-210. Western
blot analysis additionally demonstrated that miR-210 was able to
suppress the expression of KLF7. Furthermore, the expression level
of KLF7 protein in hypoxic GC-2 cells decreased significantly
compared with the control cells at each time point. Under hypoxic
conditions, overexpression of miR-210 increased the apoptosis of
GC-2 cells and pro-apoptotic protein expression, whereas
cotransfection of the miR-210 mimics and the KLF7 mimics decreased
the effects of the transfection with miR-210 mimics. Converse
effects for Bcl-2 and KLF7 were observed for caspase-3 and Bax.
These data suggested that the effects of hypoxia-induced miR-210
expression on apoptosis-associated protein function were via the
targeting of KLF7 in GC-2 cells.
In conclusion, the results of the present study
suggested that the hypoxia-induced apoptosis of GC-2 cells is
mediated by the targeting of KLF7 by miR-210. These results may aid
the understanding of the underlying mechanisms of hypoxia-induced
GC-2 cell apoptosis, and provide insight for the development of
impaired spermatogenesis.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
Key Discipline of Medicine of Jiangsu Province (grant no.
ZDXKA2016012), the Suzhou Key Medical Center (grant nos.
SZXK2015020 and SZZXJ201501), the Suzhou Government (grant nos.
LCZX201502 and SYS201754) and the National Natural Science
Foundation of China (grant no. 81300537).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
X-DW and J-XL contributed to the study conception.
J-XL, X-DW and FX designed the study. The clinical studies were
conducted by JZ and BW, and the experimental studies were conducted
by R-QT and X-LC. BW acquired the data and performed the analysis.
Y-YZ conducted the statistical analysis. J-XL, JZ and R-QT edited
the manuscript. Y-YZ and FX edited the manuscript. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Inhorn MC and Patrizio P: Infertility
around the globe: New thinking on gender, reproductive technologies
and global movements in the 21st century. Hum Reprod Update.
21:411–426. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Okada H, Tajima A, Shichiri K, Tanaka A,
Tanaka K and Inoue I: Genome-wide expression of azoospermia testes
demonstrates a specific profile and implicates ART3 in genetic
susceptibility. PloS Genet. 4:e262008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wu H, Sun L, Wen Y, Liu Y, Yu J, Mao F,
Wang Y, Tong C, Guo X, Hu Z, et al: Major spliceosome defects cause
male infertility and are associated with nonobstructive azoospermia
in humans. Proc Natl Acad Sci USA. 113:4134–4139. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Verratti V, Berardinelli F, Giulio CD,
Bosco G, Cacchio M, Pellicciotta M, Nicolai M, Martinotti S and
Tenaglia R: Evidence that chronic hypoxia causes reversible
impairment on male fertility. Asian J Androl. 10:602–606. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Torres M, Laguna-Barraza R, Dalmases M,
Calle A, Pericuesta E, Montserrat JM, Navajas D, Gutierrez-Adan A
and Farré R: Male fertility is reduced by chronic intermittent
hypoxia mimicking sleep apnea in mice. Sleep. 37:1757–1765. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yin J, Bing N, Liao WG and Gao YQ:
Hypoxia-induced apoptosis of mouse spermatocytes is mediated by
HIF-1α through a death receptor pathway and a mitochondrial
pathway. J Cell Physiol. 233:1146–1155. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Reyes JG, Farias JG, Henríquez-Olavarrieta
S, Madrid E, Parraga M, Zepeda AB and Moreno RD: The hypoxic
testicle: Physiology and pathophysiology. Oxid Med Cell Longev.
2012:9292852012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liao W, Cai M, Chen J, Huang J, Liu F,
Jiang C and Gao Y: Hypobaric hypoxia causes deleterious effects on
spermatogenesis in rats. Reproduction. 139:1031–1038. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Stark A, Bushati N, Jan CH, Kheradpour P,
Hodges E, Brennecke J, Bartel DP, Cohen SM and Kellis M: A single
Hox locus in Drosophila produces functional microRNAs from opposite
DNA strands. Genes Dev. 22:8–13. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Marignol L, Coffey M, Lawler M and
Hollywood D: Hypoxia in prostate cancer: A powerful shield against
tumour destruction? Cancer Treat Rev. 34:313–327. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kotaja N and Sassone-Corsi P: The
chromatoid body: A germ-cell-specific RNA-processing centre. Nat
Rev Mol Cell Biol. 8:85–90. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ro S, Park C, Sanders KM, McCarrey JR and
Yan W: Cloning and expression profiling of testis-expressed
microRNAs. Dev Biol. 311:592–602. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Crosby ME, Devlin CM, Glazer PM, Calin GA
and Ivan M: Emerging roles of microRNAs in the molecular responses
to hypoxia. Curr Pharm Des. 15:3861–3866. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Huang X, Ding L, Bennewith KL, Tong RT,
Welford SM, Ang KK, Story M, Le QT and Giaccia AJ:
Hypoxia-inducible mir-210 regulates normoxic gene expression
involved in tumor initiation. Mol Cell. 35:856–867. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Giannakakis A, Sandaltzopoulos R, Greshock
J, Liang S, Huang J, Hasegawa K, Li C, O'Brien-Jenkins A, Katsaros
D, Weber BL, et al: miR-210 links hypoxia with cell cycle
regulation and is deleted in human epithelial ovarian cancer.
Cancer Biol Ther. 7:255–264. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang C, Zhang ZZ, Yang W, Ouyang ZH, Xue
JB, Li XL, Zhang J, Chen WK, Yan YG and Wang WJ: MiR-210
facilitates ECM degradation by suppressing autophagy via silencing
of ATG7 in human degenerated NP cells. Biomed Pharmacother.
93:470–479. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee DC, Romero R, Kim JS, Tarca AL,
Montenegro D, Pineles BL, Kim E, Lee J, Kim SY, Draghici S, et al:
miR-210 targets iron-sulfur cluster scaffold homologue in human
trophoblast cell lines: Siderosis of interstitial trophoblasts as a
novel pathology of preterm preeclampsia and
small-for-gestational-age pregnancies. Am J Pathol. 179:590–602.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
White K, Lu Y, Annis S, Hale AE, Chau BN,
Dahlman JE, Hemann C, Opotowsky AR, Vargas SO, Rosas I, et al:
Genetic and hypoxic alterations of the microRNA-210-ISCU1/2 axis
promote iron-sulfur deficiency and pulmonary hypertension. EMBO Mol
Med. 7:695–713. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gao Y, Cao Q, Lu L, Zhang X, Zhang Z, Dong
X, Jia W and Cao Y: Kruppel-like factor family genes are expressed
during Xenopus embryogenesis and involved in germ layer formation
and body axis patterning. Dev Dyn. 244:1328–1346. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Matsumoto N, Laub F, Aldabe R, Zhang W,
Ramirez F, Yoshida T and Terada M: Cloning the cDNA for a new human
zinc finger protein defines a group of closely related Krüppel-like
transcription factors. J Biol Chem. 273:28229–28237. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
James-Zorn C, Ponferrada VG, Jarabek CJ,
Burns KA, Segerdell EJ, Lee J, Snyder K, Bhattacharyya B, Karpinka
JB, Fortriede J, et al: Xenbase: Expansion and updates of the
Xenopus model organism database. Nucleic Acids Res. 41:D865–D870.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang Z, Wang H, Sun Y, Li H and Wang N:
Klf7 modulates the differentiation and proliferation of chicken
preadipocyte. Acta Biochim Biophys Sin (Shanghai). 45:280–288.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tang D, Huang Y, Liu W and Zhang X:
Up-regulation of microRNA-210 is associated with spermatogenesis by
targeting IGF2 in male infertility. Med Sci Monit. 22:2905–2910.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kelly TJ, Souza AL, Clish CB and
Puigserver P: A hypoxia-induced positive feedback loop promotes
hypoxia-inducible factor 1alpha stability through miR-210
suppression of glycerol-3-phosphate dehydrogenase 1-like. Mol Cell
Biol. 31:2696–2706. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Feng Y, Zhu M, Dangelmajer S, Lee YM,
Wijesekera O, Castellanos CX, Denduluri A, Chaichana KL, Li Q,
Zhang H, et al: Hypoxia-cultured human adipose-derived mesenchymal
stem cells are non-oncogenic and have enhanced viability, motility,
and tropism to brain cancer. Cell Death Dis. 5:e15672014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Salama R, Masson N, Simpson P, Sciesielski
LK, Sun M, Tian YM, Ratcliffe PJ and Mole DR: Heterogeneous effects
of direct hypoxia pathway activation in kidney cancer. PloS One.
10:e01346452015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
de Motta LL, Ledaki I, Purshouse K, Haider
S, De Bastiani MA, Baban D, Morotti M, Steers G, Wigfield S,
Bridges E, et al: The BET inhibitor JQ1 selectively impairs tumour
response to hypoxia and downregulates CA9 and angiogenesis in
triple negative breast cancer. Oncogene. 36:122–132. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li R, Geng HH, Xiao J, Qin XT, Wang F,
Xing JH, Xia YF, Mao Y, Liang JW and Ji XP: miR-7a/b attenuates
post-myocardial infarction remodeling and protects H9c2
cardiomyoblast against hypoxia-induced apoptosis involving Sp1 and
PARP-1. Sci Rep. 6:290822016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Taraseviciene-Stewart L, Kasahara Y, Alger
L, Hirth P, Mc Mahon G, Waltenberger J, Voelkel NF and Tuder RM:
Inhibition of the VEGF receptor 2 combined with chronic hypoxia
causes cell death-dependent pulmonary endothelial cell
proliferation and severe pulmonary hypertension. FASEB J.
15:427–438. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ma YL, Zhang LX, Liu GL, Fan Y, Peng Y and
Hou WG: N-Myc downstream-regulated gene 2 (Ndrg2) is involved in
ischemia-hypoxia-induced astrocyte apoptosis: A novel target for
stroke therapy. Mol Neurobiol. 54:3286–3299. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang Q, Fan K, Wang P, Yu J, Liu R, Qi H,
Sun H and Cao Y: Carvacrol induces the apoptosis of pulmonary
artery smooth muscle cells under hypoxia. Eur J Pharmacol.
770:134–146. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kalakech H, Tamareille S, Pons S,
Godin-Ribuot D, Carmeliet P, Furber A, Martin V, Berdeaux A, Ghaleh
B and Prunier F: Role of hypoxia inducible factor-1α in remote limb
ischemic preconditioning. J Mol Cell Cardiol. 65:98–104. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Janke K, Brockmeier U, Kuhlmann K,
Eisenacher M, Nolde J, Meyer HE, Mairbäurl H and Metzen E: Factor
inhibiting HIF-1 (FIH-1) modulates protein interactions of
apoptosis-stimulating p53 binding protein 2 (ASPP2). J Cell Sci.
126:2629–2640. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Huang X, Yang K, Zhang Y, Wang Q and Li Y:
Quinolinic acid induces cell apoptosis in PC12 cells through
HIF-1-dependent RTP801 activation. Metab Brain Dis. 31:435–444.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
McCormick RI, Blick C, Ragoussis J,
Schoedel J, Mole DR, Young AC, Selby PJ, Banks RE and Harris AL:
miR-210 is a target of hypoxia-inducible factors 1 and 2 in renal
cancer, regulates ISCU and correlates with good prognosis. Br J
Cancer. 108:1133–1142. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mutharasan RK, Nagpal V, Ichikawa Y and
Ardehali H: microRNA-210 is upregulated in hypoxic cardiomyocytes
through Akt- and p53-dependent pathways and exerts cytoprotective
effects. Am J Physiol Heart Circ Physiol. 301:H1519–H1530. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kaczynski J, Cook T and Urrutia R: Sp1-
and Krüppel-like transcription factors. Genome Biol. 4:2062003.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Laub F, Aldabe R, Friedrich V Jr, Ohnishi
S, Yoshida T and Ramirez F: Developmental expression of mouse
Krüppel-like transcription factor KLF7 suggests a potential role in
neurogenesis. Dev Biol. 233:305–318. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kawamura Y, Tanaka Y, Kawamori R and Maeda
S: Overexpression of Kruppel-like factor 7 regulates adipocytokine
gene expressions in human adipocytes and inhibits glucose-induced
insulin secretion in pancreatic beta-cell line. Mol Endocrinol.
20:844–856. 2006. View Article : Google Scholar : PubMed/NCBI
|