Introduction
Breast cancer is the most common cancer in women
worldwide; it accounts for ~25% of all female malignancies and its
prevalence is higher in developed countries (1). Breast cancer is the second leading
cause of cancer-associated mortality among women in the world
(1,2). Current therapeutic strategies for
breast cancer, which include surgery, chemotherapy and
radiotherapy, may lack efficacy due to a high risk of relapse, poor
patient response and the emergence of drug resistance (3). This supports the requirement to
understand the genetic and biochemical factors underlying the
uncontrolled cell proliferation in breast cancer, in order to
develop novel therapies.
In breast cancer tissues, the overexpression of
cyclin-dependent kinases (CDKs) and underexpression of tumor
suppressor protein p53 is frequently observed (4). Simultaneously, a number of cell cycle
regulatory proteins are downregulated, including the CDK
inhibitors, p21, p27 and p57 (5–8).
Targeting these molecules may be effective in breast cancer therapy
(5), and natural products that
target these molecules are particularly attractive as they are
likely to have high therapeutic potential and less likely to induce
adverse effects (9,10). Plants are an excellent source of
bioactive natural compounds (7,11–13),
and polyphenolic compounds from plants frequently exert multiple
therapeutic effects (14–16). The polyphenolic phytochemical
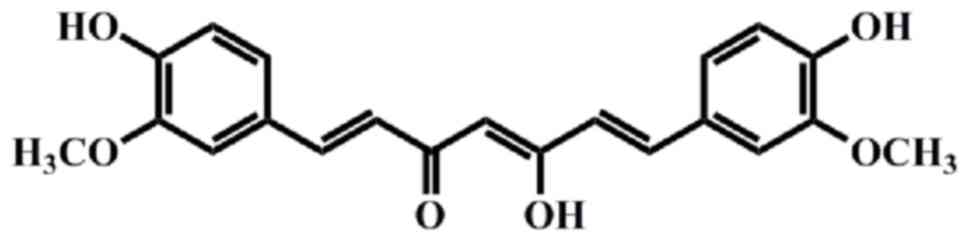
curcumin
[1,7-bis(4-hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione;
Fig. 1], isolated from the
powdered rhizome of Curcuma longa L. (Zingiberaceae)
(17,18), interacts with numerous biological
targets, including inflammatory mediators, growth factors, enzymes,
carrier proteins, metal ions, tumor suppressors, transcription
factors, oncoproteins and cellular nucleic acids (19–21).
Discovered in 1815 by Vogel and Pelletier as a yellow pigment
(4), curcumin has been consumed
for >2,000 years in Asian countries, due to its various
medicinal properties against human diseases, including cancer and
auto-immune diseases (10,17,22–29).
In the present report, the molecular targets of
curcumin and its potential benefits as a drug for breast cancer
therapy are critically reviewed.
Metabolism
One of the limitations in the use of curcumin as a
therapeutic agent is its rapid metabolism. Following absorption,
the double bonds in the heptadienedione chain are reduced, leading
to the production of a series of active metabolites (30).
Alcohol dehydrogenase reduces curcumin to tetra- and
hexahydrocurcumin in the liver, and an unidentified microsomal
enzyme leads to the formation of di- and octa-hydrocurcumin
(31). Hexahydrocurcuminol,
hexahydrocurcumin, tetrahydrocurcumin, dihydrocurcumin and their
glucuronide and sulfate conjugates have been detected in
hepatocytes (31). Curcumin and
its reduced metabolites undergo glucuronidation and are converted
into curcumin glucuronide and curcumin sulfate (32).
Bioavailability
Curcumin has very low water solubility and a high
oil-water partition coefficient. Its water solubility is low at
acidic and neutral pH; whereas, curcumin is soluble at an alkaline
pH (33). However, the compound
decomposes rapidly in alkaline media, with a half-life in the range
of a few min. Curcumin photodegrades in organic solvents (33). In total, ~80% of a typical oral
dose of curcumin passes unaltered through the gastrointestinal
tract, and the majority of the absorbed compound ends up
metabolized in the intestinal mucosa and liver (33). These properties limit the
applications of curcumin as a bioactive agent.
To overcome the limitations of poor solubility,
researchers have examined various strategies. Creating complexes of
curcumin with cyclodextrins may significantly improve its water
solubility and its stability under alkaline conditions, although
such complexation decreases the photostability of curcumin
(33,34). Stability of curcumin against
alkaline hydrolysis may additionally be improved by encapsulating
it in micelles composed of cationic surfactants, including cetyl
trimethylammonium bromide and dodecyl trimethylammonium bromide
(33,34).
Emulsion-based delivery systems have been
demonstrated to stabilize active ingredients and increase their
bioavailability (35), and the
same is true for curcumin; conjugating it with phosphatidylcholine
increases its bioavailability five-fold. Mono-polyethylene
glycolylation of curcumin produces pro-drugs that are stable in
buffer at a physiological pH and readily release curcumin into
human plasma (36).
Curcumin derivatization has additionally led to
promising drug compounds. Based on structure-activity studies of
the tautomeric forms of curcumin, the diketone system was modified
to generate two curcumin analogs, benzyloxime and isoxazole
(37). These analogs have
demonstrated much greater antitumor potency against MCF-7 breast
cancer cells and multidrug-resistant transfected MCF-7 cells
(37). Furthermore, these curcumin
analogs potently reduce expression of B-cell lymphoma-extra large
(Bcl-xL), B cell lymphoma 2 (Bcl-2) and cyclooxygenase-2 in the two
cell lines (38).
Curcumin polymers (polycurcumins) have high drug
loading efficiency and may be used as backbone-type conjugates to
stabilize and solubilize curcumin in water (39). Tang et al (39) prepared high-molecular-weight
curcumin polycurcumins through condensation polymerization of
curcumin. Polyacetal-based polycurcumin is highly cytotoxic to
MCF-7 breast cancer cell lines and to SKOV-3 intraperitoneal
xenograft tumors (40,41). This condensation approach protected
curcumin from hydrolysis at all pH values examined, and from
ultraviolet degradation. Loading curcumin into mixed polymeric
micelles improved its oral bioavailability ~55-fold (42).
Molecular targets of curcumin for breast
cancer therapy
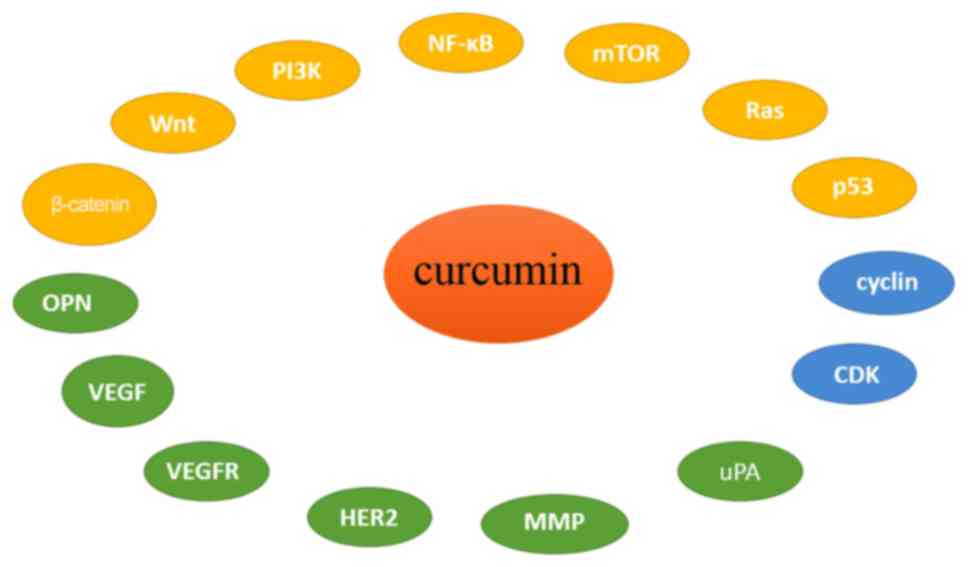
Curcumin inhibits breast cancer cell proliferation
by the following mechanisms: i) Inducing cell cycle arrest and
p53-dependent apoptosis; ii) altering expression of signaling
proteins, including Ras, phosphatidylinositol-3-kinase (PI3K),
protein kinase B (Akt), mammalian target of rapamycin (mTOR) and
Wnt/β-catenin; iii) downregulating transcription factors; and iv)
inhibiting tumor growth and angiogenesis (Fig. 2).
Effects of curcumin on CDK/cyclin
complexes
CDKs are serine/threonine kinases that control cell
cycle progression by forming a complex with their respective cyclin
partners (43,44). Altered expression of CDKs,
overexpression of cyclins and loss of expression of CDK inhibitors
are frequently observed in malignant cells (44). Dysregulated CDK activity provides
cancer cells with a selective growth advantage. In this way,
dysregulated overexpression of cyclin D1 triggers progression of
aggressive breast cancer (45).
Previous studies in mammary epithelial carcinoma cells suggest that
curcumin inhibits cell cycle progression by blocking the
association of cyclin D1 with CDK4, thus reducing cyclin D1
activity (46,47). In MCF-7 breast cancer cells,
curcumin reduces cell proliferation by arresting cells in
G1 phase. The drug achieves this arrest by stimulating
the proteosomal degradation of cyclin E and upregulating CDK
inhibitors, p53, p21 and p27; the addition of specific proteosomal
inhibitors suppresses these effects of curcumin (48). Cyclin E is a nuclear protein that
serves an important role in G1/S progression by
interacting with its catalytic partner, CDK2, and by interacting
with the retinoblastoma (Rb) protein (49,50).
It appears likely that the anti-proliferative effects of curcumin
are due to proteasome-mediated downregulation of cyclin E and
upregulation of CDK inhibitors (51).
The anti-proliferative effects of curcumin appear to
be selective to cells overexpressing CDK 2. In mammary carcinoma
cells, curcumin induces p53-dependent apoptosis and causes
G2 phase arrest. However, in normal human mammary cells,
curcumin causes G0 cell cycle arrest by blocking the
association between CDK 4 and CDK 6, and inhibiting the
phosphorylation of Rb (52). In
this way, curcumin also prevents the initiation of p53-mediated
apoptosis, which occurs only in cells arrested in G2
phase (52).
Effects of curcumin on the p53
pathway
The p53 protein is one of the most important tumor
suppressor proteins, regulating a wide range of cellular processes,
including cell proliferation, DNA damage and apoptosis (53). It is encoded by the tumor
protein p53 gene, which is frequently mutated in numerous types
of human cancer (53), leading to
loss of cell proliferative control, DNA check points and DNA repair
mechanisms. As a result, cancer cells become immortal. Restoring
the function of p53 is an attractive therapeutic strategy in cancer
therapy (54).
Curcumin induces apoptosis in breast cancer cells
via p53-dependent and -independent pathways. For instance, curcumin
arrests the cell cycle and induces p53-dependent apoptosis in MCF-7
breast cancer cells (55).
Notably, curcumin exerts no anti-proliferative effects on MDAH041
cells lacking p53 or on TR9-7 cells that express p53 at low levels;
rather, its effects are most notable in p53-expressing TR9-7 and
MCF-7 cells. Expression of the pro-apoptotic protein apoptosis
regulator Bax (Bax) is also higher in curcumin-treated MCF-7 cells.
These results suggest that curcumin exerts its anti-proliferative
effects via p53-dependent and p53-independent pathways (55,56).
Targets of curcumin in Ras
signaling
Ras is a small transmembrane protein belonging to
the large GTPase family of enzymes that hydrolyze guanosine
triphosphate in order to transduce signals inside the cell
(57). Mammalian cells have three
Ras proteins (K-, H- and N-Ras), each of which serves a different
function (57). Blocking oncogenic
Ras signaling is an attractive strategy in cancer therapy.
Curcumin has been extensively studied for its
effects on oncogenic Ras signaling pathways. In MCF-10A human
breast epithelial cells transformed using H-Ras, curcumin induces
reactive oxygen species production, which downregulates activity of
matrix metalloproteinase (MMP)-2 and Bcl-2 and upregulates the
activity of Bax and caspase-3 (58).
Potentially acting through a similar mechanism,
curcumin arrests Ras-transfected HAG-1 human adenocarcinoma cells
in G2/M phase by inducing expression of extracellular
signal-regulated kinase 1/2 and Bax, and reducing expression of
Bcl-xL. These results suggest that curcumin may be a potent therapy
against Ras-overexpressing cancer (59). Preclinical studies in animals, and
ultimately clinical trials, are required to clarify the therapeutic
effect of curcumin in Ras-induced cancer.
Targets of curcumin in PI3K/Akt/mTOR
signaling
PI3Ks are a family of lipid kinases that
phosphorylate inositol phospholipids and generate the secondary
messenger phosphatidylinositol-3,4,5-trisphosphate in the plasma
membrane (60). PI3K interacts
with Akt to trigger the latter's translocation inside the
cytoplasm. Activated Akt interacts with a number of substrates to
perform numerous functions in cell survival, cell cycle progression
and cell growth (60).
Constitutive expression of PI3K and Akt, in addition to silencing
of phosphatase and tensin homolog and glycogen synthase kinase 3β
(GSK3β), are frequently observed in a number of human malignancies.
Therefore, PI3K/Akt-mediated signaling is an attractive target in
cancer chemotherapy (61,62).
Cancer cells survive for a prolonged time by
activating survival pathways involving PI3K, Akt and mTOR, in
addition to anti-apoptotic pathways involving Bcl-2. Targeting
survival and apoptosis pathways is likely to be essential for
controlling highly metastatic breast cancer. Curcumin on its own
weakly stimulates apoptosis in breast cancer cells; however,
combining it with the PI3K-specific inhibitor LY294002 stimulates
apoptosis more strongly (63,64).
The authors of these previous studies hypothesized that the PI3K
obstruction overcomes the oncogenic expression of Bcl-2. Further
studies are required to verify whether curcumin may inhibit
PI3K/Akt/mTOR signaling in breast cancer cells and identify the
mechanism(s) involved.
Targets of curcumin in Wnt/β-catenin
signaling
Wnts are a family of secreted glycoproteins that
regulate multiple signaling pathways through β-catenin-dependent
and -independent mechanisms (65–67).
Wnts serve a crucial role in development, survival and metabolism.
Inappropriate regulation and hyperactivation of Wnt/β-catenin
signaling have been implicated in numerous human malignancies.
Overexpression of β-catenin leads to constitutive activation of
cell proliferation (68), and
tumor cells downregulate the tumor suppressor GSK3β, which limits
the activity of β-catenin by triggering its ubiquitin-mediated
proteosomal degradation. Therefore, targeting the Wnt/β-catenin
signaling pathway is an attractive approach in cancer therapy
(69,70).
In MCF-7 and MDA-MB-231 cells, curcumin arrests the
cell cycle in G2/M cells by modulating Wnt/β-catenin
signaling. In these cells, curcumin upregulates GSK3β and causes
loss of nuclear β-catenin. Loss of nuclear β-catenin results in a
loss of its downstream target cyclin D1 (71). This suggests that, at least in
MCF-7 and MDA-MB-231 cells, the antitumor effects of curcumin are
due to abrogation of Wnt/β-catenin signaling (71).
Targets of curcumin among nuclear
factor-κB (NF-κB) transcription factors
NF-κB is a family of transcription factors that are
involved in the immune response and inflammation. Gene expression
profiling studies suggest that the NF-κB pathway is a key regulator
in triple-negative breast cancer (TNBC), with activation of NF-κB
signaling strongly implicated in the pathogenesis of specific TNBCs
(72–74). Cytoplasmic NF-κB is bound to a
group of inhibitory proteins known as inhibitors of NF-κB (IκB);
accumulation of non-phosphorylated IκB prohibits the translocation
of NF-κB from cytoplasm to nucleus, resulting in inactivation of
NF-κB and its downstream targets (74). NF-κB promotes the transcription of
numerous key regulators of cancer invasion and progression,
including cytokines, chemokines, cell adhesion molecules and
inducible pro-inflammatory enzymes (74). In addition, NF-κB has been
postulated to be a useful marker of the epithelial-mesenchymal
transition (EMT) and invasiveness in breast cancer (19).
A number of previous studies suggest that curcumin
inhibits NF-κB expression and therefore additional downstream
signaling pathways, ultimately leading to the silencing of
inflammatory cytokines, including chemokine (C-X-C motif) ligand
(CXCL)1 and CXCL2 (19); and to
alterations in the expression of MMP-9, urokinase plasminogen
activator (uPA), uPA receptor, intercellular adhesion molecule 1
and chemokine receptor 4 (3,72,75).
In this manner, curcumin is likely to inhibit the growth and
invasion of breast cancer, in part, by downregulating NF-κB
signaling pathways.
Curcumin may modulate the expression of NF-κB target
genes (76,77), which include Bcl-2, ornithine
decarboxylase (ODC) and c-myc, which are associated with apoptosis
or cell survival (78). For
example, ODC is the rate-limiting enzyme in polyamine biosynthesis
and curcumin has been demonstrated to suppress ODC activity and
inhibit cell proliferation (79).
Activation of the NF-κB/Bcl-2 pathway is associated with drug
resistance in cancer cells (80).
Accumulating evidence suggests that targeting NF-κB
to inhibit cell growth and reverse EMT may be a novel therapeutic
strategy in breast cancer.
Targets of curcumin in tumor
angiogenesis
Angiogenesis is the normal physiological mechanism
by which novel blood vessels are formed from pre-existing blood
vessels. It occurs during embryogenesis, menstruation and wound
healing (81). Angiogenesis in
tumors is crucial for cancer progression. Tumor cells procure
nutrients for their uncontrolled growth through tumor angiogenesis
(81). Tumor cells constitutively
produce pro-angiogenic factors, including vascular endothelial
growth factor (VEGF) and basic fibroblast growth factor, which
curcumin may inhibit in order to modulate tumor angiogenesis
(81,82).
Curcumin inhibits angiogenesis and growth of breast
cancer tumors implanted into nude mice. These effects are
associated with downregulated expression of a number of VEGF
isomers, including VEGF-A, VEGF-C and VEGF receptor 2, in addition
to decreased microvessel density (83). These results are in agreement with
other previous studies demonstrating that suppression of VEGF
function inhibits breast tumor growth (83,84).
In nude mice, which were implanted with MDA-MB-231 tumors and
treated with osteopontin (OPN; additionally termed secreted
phosphoprotein 1) to stimulate angiogenesis, curcumin blocked
NF-κB/cyclic AMP-dependent transcription factor ATF-4 binding and
prevented OPN-induced upregulation of VEGF (85). This suggests that curcumin acts as
a potent anti-angiogenic agent in regulating OPN-induced tumor
angiogenesis in breast cancer.
Potential risks and adverse side effects of
curcumin
Curcumin causes blood thinning, which may decrease
blood flow and increase the risk of ischemic stroke (86). It may also inhibit the ability of
chemotherapeutics to induce production of reactive oxygen species
and block the c-Jun NH2-terminal kinase pathway. In fact, curcumin
may exert pro-oxidant effects, similar to numerous other
anti-oxidants (87).
Curcumin significantly inhibits
cyclophosphamide-induced regression of human breast cancer
xenografts in mice (88,89). In cultures of MCF-7, MDA-MB-231 and
BT-474 human breast cancer cells, curcumin may inhibit the ability
of camptothecin, mechlorethamine and doxorubicin to induce
apoptosis by ≤70% (18,90). Curcumin may also serve as an iron
chelator to inhibit hypoxia inducible factor-α prolyl hydroxylase
activity (91). Therefore, further
research is urgently required to establish whether patients with
breast cancer undergoing chemotherapy should limit their intake of
curcumin.
Conclusion
The available evidence suggests that curcumin, a
polyphenolic compound derived from the dietary spice turmeric, is a
non-toxic, highly promising natural anti-oxidant that exerts
anticancer effects by targeting multiple molecules and pathways. By
affecting different targets, curcumin modulates numerous cancer
hallmarks, including cell proliferation, cancer signaling pathways,
transcription factors and tumor angiogenesis. Curcumin may have
applications as a novel drug in the near future to control various
diseases, particularly breast cancer.
The clinical use of curcumin is limited by its poor
bioavailability; however, specific novel derivatives have been
prepared that may improve patient responses. Research is in
progress on nanotechnology-based formulations and delivery systems
to improve curcumin pharmacokinetics. Possibilities include
encapsulating curcumin into polymeric or lipid micelles, or
liposomes, and combining or conjugating curcumin to ligands or
antibodies that may target cancer cell receptors or other epitopes.
Novel curcumin analogs and nanotechnology-based formulations may
overcome the limitations of oral administration of curcumin.
Acknowledgements
Not applicable.
Funding
Supported by the Henan Science and Technology
Innovation Team, Investigation on Plant Resources in Dabie
Mountains and the study and utilization of active components of
special plants (grant no. 2017083), Nanhu Scholars Program for
Young Scholars of Xinyang Normal University (Xinyang, China; grant
no. 2018001).
Availability of data and materials
Not applicable.
Authors' contributions
XS conceived and designed the article. MZ and ED
read the literature and collated the appropriate information. XS
and YL wrote the paper.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
Bcl-xL
|
B-cell lymphoma-extra large
|
|
CDK
|
cyclin-dependent kinase
|
|
CXCL
|
chemokine (C-X-C motif) ligand
|
|
GSK3β
|
glycogen synthase kinase-3β
|
|
mTOR
|
mammalian target of rapamycin
|
|
NF-κB
|
nuclear factor-κB
|
|
OPN
|
osteopontin
|
|
ODC
|
ornithine decarboxylase
|
|
PI3K
|
phosphatidylinositol-3-kinase
|
|
TNBC
|
triple-negative breast cancer
|
|
uPA
|
urinary plasminogen activator
|
|
VEGF
|
vascular endothelial growth factor
|
References
|
1
|
Garcia-Aranda M and Redondo M: Protein
kinase targets in breast cancer. Int J Mol Sci. 18(pii): E25432017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sun YS, Zhao Z, Yang ZN, Xu F, Lu HJ, Zhu
ZY, Shi W, Jiang J, Yao PP and Zhu HP: Risk factors and preventions
of breast cancer. Int J Biol Sci. 13:1387–1397. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ye JC and Formenti SC: Integration of
radiation and immunotherapy in breast cancer - Treatment
implications. Breast. 38:66–74. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kasi PD, Tamilselvam R, Skalicka-Woźniak
K, Nabavi SF, Daglia M, Bishayee A, Pazoki-Toroudi H and Nabavi SM:
Molecular targets of curcumin for cancer therapy: An updated
review. Tumour Biol. 37:13017–13028. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Williams GH and Stoeber K: The cell cycle
and cancer. J Pathol. 226:352–364. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fulda S: Modulation of apoptosis by
natural products for cancer therapy. Planta Med. 76:1075–1079.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tan W, Lu J, Huang M, Li Y, Chen M, Wu G,
Gong J, Zhong Z, Xu Z, Dang Y, et al: Anti-cancer natural products
isolated from chinese medicinal herbs. Chin Med. 6:272011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Demain AL and Vaishnav P: Natural products
for cancer chemotherapy. Microb Biotechnol. 4:687–699. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yuan R, Hou Y, Sun W, Yu J, Liu X, Niu Y,
Lu JJ and Chen X: Natural products to prevent drug resistance in
cancer chemotherapy: A review. Ann N Y Acad Sci. 1401:19–27. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Messner DJ, Robinson T and Kowdley KV:
Curcumin and turmeric modulate the tumor-promoting effects of iron
in vitro. Nutr Cancer. 69:481–489. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hou J and Cui HL: In Vitro antioxidant,
antihemolytic and anticancer activity of the carotenoids from
halophilic archaea. Curr Microbiol. 75:266–271. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nabavi SM, Marchese A, Izadi M, Curti V,
Daglia M and Nabavi SF: Plants belonging to the genus Thymus as
antibacterial agents: from farm to pharmacy. Food Chem.
173:339–347. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang HL, Korivi M, Lin MK, Chang HC, Wu
CR, Lee MS, Chen WT and Hseu YC: Antihemolytic and antioxidant
properties of pearl powder against 2,2′-azobis(2-amidinopropane)
dihydrochloride-induced hemolysis and oxidative damage to
erythrocyte membrane lipids and proteins. J Food Drug Anal.
25:898–907. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hirapara H, Ghori V, Anovadiya A, Baxi S
and Tripathi C: Effects of ethanolic extract of Jasminum
grandiflorum Linn. Flowers on wound healing in diabetic Wistar
albino rats. Avicenna J Phytomed. 7:401–408. 2017.PubMed/NCBI
|
|
15
|
Nomura EC, Rodrigues MR, da Silva CF, Hamm
LA, Nascimento AM, de Souza LM, Cipriani TR, Baggio CH and Werner
MF: Antinociceptive effects of ethanolic extract from the flowers
of Acmella oleracea (L.) R.K. Jansen in mice. J Ethnopharmacol.
150:583–589. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Nabavi SM, Habtemariam S, Nabavi SF,
Sureda A, Daglia M, Moghaddam AH and Amani MA: Protective effect of
gallic acid isolated from Peltiphyllum peltatum against sodium
fluoride-induced oxidative stress in rat's kidney. Mol Cell
Biochem. 372:233–239. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fadus MC, Lau C, Bikhchandani J and Lynch
HT: Curcumin: An age-old anti-inflammatory and anti-neoplastic
agent. J Tradit Complement Med. 7:339–346. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Banik U, Parasuraman S, Adhikary AK and
Othman NH: Curcumin: The spicy modulator of breast carcinogenesis.
J Exp Clin Cancer Res. 36:982017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shishodia S, Sethi G and Aggarwal BB:
Curcumin: Getting back to the roots. Ann N Y Acad Sci.
1056:206–217. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Aggarwal BB, Kumar A and Bharti AC:
Anticancer potential of curcumin: Preclinical and clinical studies.
Anticancer Res. 23:363–398. 2003.PubMed/NCBI
|
|
21
|
Han X, Deng S, Wang N, Liu Y and Yang X:
Inhibitory effects and molecular mechanisms of tetrahydrocurcumin
against human breast cancer MCF-7 cells. Food Nutr Res.
60:306162016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu JL, Pan YY, Chen O, Luan Y, Xue X,
Zhao JJ, Liu L and Jia HY: Curcumin inhibits MCF-7 cells by
modulating the NF-κB signaling pathway. Oncol Lett. 14:5581–5584.
2017.PubMed/NCBI
|
|
23
|
Zheng J, Zhou Y, Li Y, Xu DP, Li S and Li
HB: Spices for prevention and treatment of cancers. Nutrients.
8(pii): E4952016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang Y, Yu J, Cui R, Lin J and Ding X:
Curcumin in treating breast cancer: A review. J Lab Autom.
21:723–731. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Imran M, Ullah A, Saeed F, Nadeem M,
Arshad MU and Suleria HAR: Cucurmin, anticancer, & antitumor
perspectives: A comprehensive review. Crit Rev Food Sci Nutr.
58:1271–1293. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ko EY and Moon A: Natural products for
chemoprevention of breast cancer. J Cancer Prev. 20:223–231. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hossain DM, Bhattacharyya S, Das T and Sa
G: Curcumin: The multi-targeted therapy for cancer regression.
Front Biosci (Schol Ed). 4:335–355. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhou H, Beevers CS and Huang S: The
targets of curcumin. Curr Drug Targets. 12:332–347. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Huang G, Xu Z, Huang Y, Duan X, Gong W,
Zhang Y, Fan J and He F: Curcumin protects against collagen-induced
arthritis via suppression of BAFF production. J Clin Immunol.
33:550–557. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Schneider C, Gordon ON, Edwards RL and
Luis PB: Degradation of curcumin: From mechanism to biological
implications. J Agric Food Chem. 63:7606–7614. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jamil QUA, Jaerapong N, Zehl M,
Jarukamjorn K and Jäger W: Metabolism of curcumin in human breast
cancer cells: Impact of sulfation on cytotoxicity. Planta Med.
83:1028–1034. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Vera-Ramirez L, Perez-Lopez P,
Varela-Lopez A, Ramirez-Tortosa M, Battino M and Quiles JL:
Curcumin and liver disease. Biofactors. 39:88–100. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Marin E, Briceño MI, Torres A and
Caballero-George C: New curcumin-loaded chitosan nanocapsules: In
vivo evaluation. Planta Med. 83:877–883. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Feng T, Wei Y, Lee RJ and Zhao L:
Liposomal curcumin and its application in cancer. Int J
Nanomedicine. 12:6027–6044. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tamvakopoulos C, Dimas K, Sofianos ZD,
Hatziantoniou S, Han Z, Liu ZL, Wyche JH and Pantazis P: Metabolism
and anticancer activity of the curcumin analogue,
dimethoxycurcumin. Clin Cancer Res. 13:1269–1277. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Venkateswarlu S, Ramachandra MS and
Subbaraju GV: Synthesis and biological evaluation of
polyhydroxycurcuminoids. Bioorg Med Chem. 13:6374–6380. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Nagaraju GP, Aliya S, Zafar SF, Basha R,
Diaz R and El-Rayes BF: The impact of curcumin on breast cancer.
Integr Biol (Camb). 4:996–1007. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Labbozzetta M, Notarbartolo M, Poma P,
Maurici A, Inguglia L, Marchetti P, Rizzi M, Baruchello R, Simoni D
and D'Alessandro N: Curcumin as a possible lead compound against
hormone-independent, multidrug-resistant breast cancer. Ann N Y
Acad Sci. 1155:278–283. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Tang H, Murphy CJ, Zhang B, Shen Y, Van
Kirk EA, Murdoch WJ and Radosz M: Curcumin polymers as anticancer
conjugates. Biomaterials. 31:7139–7149. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Catania A, Barrajón-Catalán E, Nicolosi S,
Cicirata F and Micol V: Immunoliposome encapsulation increases
cytotoxic activity and selectivity of curcumin and resveratrol
against HER2 overexpressing human breast cancer cells. Breast
Cancer Res Treat. 141:55–65. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Bayet-Robert M and Morvan D: Metabolomics
reveals metabolic targets and biphasic responses in breast cancer
cells treated by curcumin alone and in association with docetaxel.
PLoS One. 8:e579712013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Patil S, Choudhary B, Rathore A, Roy K and
Mahadik K: Enhanced oral bioavailability and anticancer activity of
novel curcumin loaded mixed micelles in human lung cancer cells.
Phytomedicine. 22:1103–1111. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Frouin I, Montecucco A, Biamonti G,
Hübscher U, Spadari S and Maga G: Cell cycle-dependent dynamic
association of cyclin/Cdk complexes with human DNA replication
proteins. EMBO J. 21:2485–2495. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
John PC, Mews M and Moore R: Cyclin/Cdk
complexes: Their involvement in cell cycle progression and mitotic
division. Protoplasma. 216:119–142. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Villegas SL, Darb-Esfahani S, von
Minckwitz G, Huober J, Weber K, Marmé F, Furlanetto J, Schem C,
Pfitzner BM, Lederer B, et al: Expression of cyclin D1 protein in
residual tumor after neoadjuvant chemotherapy for breast cancer.
Breast Cancer Res Treat. 168:179–187. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Lamb R, Lehn S, Rogerson L, Clarke RB and
Landberg G: Cell cycle regulators cyclin D1 and CDK4/6 have
estrogen receptor-dependent divergent functions in breast cancer
migration and stem cell-like activity. Cell Cycle. 12:2384–2394.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Peurala E, Koivunen P, Haapasaari KM,
Bloigu R and Jukkola-Vuorinen A: The prognostic significance and
value of cyclin D1, CDK4 and p16 in human breast cancer. Breast
Cancer Res. 15:R52013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Aggarwal BB, Banerjee S, Bharadwaj U, Sung
B, Shishodia S and Sethi G: Curcumin induces the degradation of
cyclin E expression through ubiquitin-dependent pathway and
up-regulates cyclin-dependent kinase inhibitors p21 and p27 in
multiple human tumor cell lines. Biochem Pharmacol. 73:1024–1032.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Doostan I, Karakas C, Kohansal M, Low KH,
Ellis MJ, Olson JA Jr, Suman VJ, Hunt KK, Moulder SL and Keyomarsi
K: Cytoplasmic cyclin E mediates resistance to aromatase inhibitors
in breast cancer. Clin Cancer Res. 23:7288–7300. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Mazumder S, DuPree EL and Almasan A: A
dual role of cyclin E in cell proliferation and apoptosis may
provide a target for cancer therapy. Curr Cancer Drug Targets.
4:65–75. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Keyomarsi K, Tucker SL, Buchholz TA,
Callister M, Ding Y, Hortobagyi GN, Bedrosian I, Knickerbocker C,
Toyofuku W, Lowe M, et al: Cyclin E and survival in patients with
breast cancer. N Engl J Med. 347:1566–1575. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Choudhuri T, Pal S, Das T and Sa G:
Curcumin selectively induces apoptosis in deregulated cyclin
D1-expressed cells at G2 phase of cell cycle in a p53-dependent
manner. J Biol Chem. 280:20059–20068. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Kandoth C, McLellan MD, Vandin F, Ye K,
Niu B, Lu C, Xie M, Zhang Q, McMichael JF, Wyczalkowski MA, et al:
Mutational landscape and significance across 12 major cancer types.
Nature. 502:333–339. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Muller PA and Vousden KH: Mutant p53 in
cancer: New functions and therapeutic opportunities. Cancer Cell.
25:304–317. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Fan H, Liang Y, Jiang B, Li X, Xun H, Sun
J, He W, Lau HT and Ma X: Curcumin inhibits intracellular fatty
acid synthase and induces apoptosis in human breast cancer
MDA-MB-231 cells. Oncol Rep. 35:2651–2656. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Choudhuri T, Pal S, Agwarwal ML, Das T and
Sa G: Curcumin induces apoptosis in human breast cancer cells
through p53-dependent Bax induction. FEBS Lett. 512:334–340. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Adjei AA: Blocking oncogenic Ras signaling
for cancer therapy. J Natl Cancer Inst. 93:1062–1074. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Kim MS, Kang HJ and Moon A: Inhibition of
invasion and induction of apoptosis by curcumin in
H-ras-transformed MCF10A human breast epithelial cells. Arch Pharm
Res. 24:349–354. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Ono M, Higuchi T, Takeshima M, Chen C and
Nakano S: Differential anti-tumor activities of curcumin against
Ras- and Src-activated human adenocarcinoma cells. Biochem Biophys
Res Commun. 436:186–191. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Ettl T, Schwarz-Furlan S, Haubner F,
Müller S, Zenk J, Gosau M, Reichert TE and Zeitler K: The
PI3K/AKT/mTOR signalling pathway is active in salivary gland cancer
and implies different functions and prognoses depending on cell
localisation. Oral Oncol. 48:822–830. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Chen WC, Lai YA, Lin YC, Ma JW, Huang LF,
Yang NS, Ho CT, Kuo SC and Way TD: Curcumin suppresses
doxorubicin-induced epithelial-mesenchymal transition via the
inhibition of TGF-β and PI3K/AKT signaling pathways in
triple-negative breast cancer cells. J Agric Food Chem.
61:11817–11824. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Carnero A, Blanco-Aparicio C, Renner O,
Link W and Leal JF: The PTEN/PI3K/AKT signalling pathway in cancer,
therapeutic implications. Curr Cancer Drug Targets. 8:187–198.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Akkoc Y, Berrak Ö, Arisan ED, Obakan P,
Çoker-Gürkan A and Palavan-Ünsal N: Inhibition of PI3K signaling
triggered apoptotic potential of curcumin which is hindered by
Bcl-2 through activation of autophagy in MCF-7 cells. Biomed
Pharmacother. 71:161–171. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Yu S, Shen G, Khor TO, Kim JH and Kong AN:
Curcumin inhibits Akt/mammalian target of rapamycin signaling
through protein phosphatase-dependent mechanism. Mol Cancer Ther.
7:2609–2620. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Zou YF, Xie CW, Yang SX and Xiong JP: AMPK
activators suppress breast cancer cell growth by inhibiting
DVL3-facilitated Wnt/β-catenin signaling pathway activity. Mol Med
Rep. 15:899–907. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Pohl SG, Brook N, Agostino M, Arfuso F,
Kumar AP and Dharmarajan A: Wnt signaling in triple-negative breast
cancer. Oncogenesis. 6:e3102017. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Kwon YJ, Leibovitch BA, Bansal N, Pereira
L, Chung CY, Ariztia EV, Zelent A, Farias EF and Waxman S: Targeted
interference of SIN3A-TGIF1 function by SID decoy treatment
inhibits Wnt signaling and invasion in triple negative breast
cancer cells. Oncotarget. 8:88421–88436. 2016.PubMed/NCBI
|
|
68
|
Ma X, Zhao X, Yan W, Yang J, Zhao X, Zhang
H, Hui Y and Zhang S: Tumor-infiltrating lymphocytes are associated
with β-catenin overexpression in breast cancer. Cancer Biomark.
21:639–650. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Roy A, Ansari SA, Das K, Prasad R,
Bhattacharya A, Mallik S, Mukherjee A and Sen P: Coagulation factor
VIIa-mediated protease-activated receptor 2 activation leads to
β-catenin accumulation via the AKT/GSK3β pathway and contributes to
breast cancer progression. J Biol Chem. 292:13688–13701. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Gao S, Ge A, Xu S, You Z, Ning S, Zhao Y
and Pang D: PSAT1 is regulated by ATF4 and enhances cell
proliferation via the GSK3β/β-catenin/cyclin D1 signaling pathway
in ER-negative breast cancer. J Exp Clin Cancer Res. 36:1792017.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Prasad CP, Rath G, Mathur S, Bhatnagar D
and Ralhan R: Potent growth suppressive activity of curcumin in
human breast cancer cells: Modulation of Wnt/beta-catenin
signaling. Chem Biol Interact. 181:263–271. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Poma P, Labbozzetta M, D'Alessandro N and
Notarbartolo M: NF-kB is a potential molecular drug target in
triple-negative breast cancers. OMICS. 21:225–231. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Pires BR, Mencalha AL, Ferreira GM, de
Souza WF, Morgado-Diaz JA, Maia AM, Corrêa S and Abdelhay ES:
NF-kappaB is involved in the regulation of EMT genes in breast
cancer cells. PLoS One. 12:e01696222017. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Park YH: The nuclear factor-kappa B
pathway and response to treatment in breast cancer.
Pharmacogenomics. 18:1697–1709. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Stanic Z: Curcumin, a compound from
natural sources, a true scientific challenge-a review. Plant Foods
Hum Nutr. 72:1–12. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Murray-Stewart T and Casero RA: Regulation
of polyamine metabolism by curcumin for cancer prevention and
therapy. Med Sci (Basel). 5(pii): E382017.PubMed/NCBI
|
|
77
|
Oyagbemi AA, Saba AB and Ibraheem AO:
Curcumin: From food spice to cancer prevention. Asian Pac J Cancer
Prev. 10:963–967. 2009.PubMed/NCBI
|
|
78
|
Leu TH and Maa MC: The molecular
mechanisms for the antitumorigenic effect of curcumin. Curr Med
Chem Anticancer Agents. 2:357–370. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Gafner S, Lee SK, Cuendet M, Barthélémy S,
Vergnes L, Labidalle S, Mehta RG, Boone CW and Pezzuto JM: Biologic
evaluation of curcumin and structural derivatives in cancer
chemoprevention model systems. Phytochemistry. 65:2849–2859. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Qian C, Li P, Yan W, Shi L, Zhang J, Wang
Y, Liu H and You Y: Downregulation of osteopontin enhances the
sensitivity of glioma U251 cells to temozolomide and cisplatin by
targeting the NF-κB/Bcl2 pathway. Mol Med Rep. 11:1951–1955. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Shehzad A, Qureshi M, Anwar MN and Lee YS:
Multifunctional curcumin mediate multitherapeutic effects. J Food
Sci. 82:2006–2015. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Arablou T and Kolahdouz-Mohammadi R:
Curcumin and endometriosis: Review on potential roles and molecular
mechanisms. Biomed Pharmacother. 97:91–97. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Carroll CE, Ellersieck MR and Hyder SM:
Curcumin inhibits MPA-induced secretion of VEGF from T47-D human
breast cancer cells. Menopause. 15:570–574. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Saberi-Karimian M, Katsiki N, Caraglia M,
Boccellino M, Majeed M and Sahebkar A: Vascular endothelial growth
factor: An important molecular target of curcumin. Crit Rev Food
Sci Nutr. 1–14. 2017.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Chakraborty G, Jain S, Kale S, Raja R,
Kumar S, Mishra R and Kundu GC: Curcumin suppresses breast tumor
angiogenesis by abrogating osteopontin-induced VEGF expression. Mol
Med Rep. 1:641–646. 2008.PubMed/NCBI
|
|
86
|
DeBusk RM: Dietary supplements and
cardiovascular disease. Curr Atheroscler Rep. 2:508–514. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Kawanishi S, Oikawa S and Murata M:
Evaluation for safety of antioxidant chemopreventive agents.
Antioxid Redox Signal. 7:1728–1739. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Di Martino RM, Luppi B, Bisi A, Gobbi S,
Rampa A, Abruzzo A and Belluti F: Recent progress on curcumin-based
therapeutics: A patent review (2012–2016). Part I: Curcumin. Expert
Opin Ther Pat. 27:579–590. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Di Martino RMC, Bisi A, Rampa A, Gobbi S
and Belluti F: Recent progress on curcumin-based therapeutics: A
patent review (2012–2016). Part II: Curcumin derivatives in cancer
and neurodegeneration. Expert Opin Ther Pat. 27:953–965. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Somasundaram R, Jacob L, Swoboda R, Caputo
L, Song H, Basak S, Monos D, Peritt D, Marincola F, Cai D, et al:
Inhibition of cytolytic T lymphocyte proliferation by autologous
CD4+/CD25+ regulatory T cells in a colorectal carcinoma patient is
mediated by transforming growth factor-beta. Cancer Res.
62:5267–5272. 2002.PubMed/NCBI
|
|
91
|
Strofer M, Jelkmann W, Metzen E,
Brockmeier U, Dunst J and Depping R: Stabilisation and knockdown of
HIF-two distinct ways comparably important in radiotherapy. Cell
Physiol Biochem. 28:805–812. 2011. View Article : Google Scholar : PubMed/NCBI
|