Introduction
Osteoporotic fractures are increasingly gaining
attention due to their high incidence rates and economic burden
(1). Osteoporosis is a
pathological condition characterized by a decrease in bone mineral
density or bone mass, which can reduce bone strength and lead to
fractures. Clinical osteoporosis is classified into two types: Type
I, or postmenopausal osteoporosis; and type II, i.e., osteoporosis
in the elderly (1,2). Bone homeostasis depends on the
synergistic activities of osteoclasts and osteoblasts, and any
imbalance between bone formation and resorption may lead to a
number of diseases, including osteoporosis, Paget's bone disease
and osteodystrophy (3). Apart from
decreased bone mass, osteoporosis is associated with a slower rate
of bone differentiation following an increase in the number of
adipocytes and osteoclasts, decreased osteoblasts and enhanced bone
resorption (4,5). In clinical terms, the prevention of
osteoporosis is particularly significant and includes resistance
exercises to increase bone density, adequate calcium in the diet,
and calcium supplement medications (6). In recent years, vitamin K2 (VK2) has
attracted attention as an auxiliary drug for preventing
osteoporosis (7).
VK2 is a fat-soluble vitamin that is structurally
similar to other members of the same family, including
phylloquinone, menaquinones and menadione (7). It is a critical factor in the blood
clotting process (8–10), and epidemiological studies have
demonstrated that a lack of VK2 may lead to osteoporosis and
osteoarthritis in older individuals (10,11).
In addition, VK2 has been clinically utilized to prevent
osteoporosis, and likely exerts its protective effects by promoting
osteoblast differentiation and mineralization (12–14).
However, the precise mechanism remains to be fully elucidated
(7).
Increasing evidence in recent years has implicated
autophagy in the development of osteoporosis. Autophagy is an
essential process that maintains homeostasis in eukaryotic cells by
breaking down and eliminating damaged proteins and organelles
(15,16). When a cell is subjected to external
stress, it may initiate autophagy to adjust to the associated
stimulus by degrading cytoplasmic substances to provide energy for
cell survival (17,18). Autophagy has a critical role in
physiological conditions, and is associated with certain diseases,
including cancer, diabetes and leukemia. A lack of autophagy in
osteoblasts can decrease mineralizing capacity, and induce an
imbalance in the population of osteoblasts and osteoclasts,
resulting in a low bone mass phenotype (19). The aim of the present study was to
verify whether VK2 is able to promote the differentiation and
mineralization of osteoblasts by inducing autophagy.
Materials and methods
Chemical reagents
α-Modification minimum essential medium (α-MEM) was
purchased from HyClone (GE Healthcare Life Sciences, Logan, UT,
USA). Fetal bovine serum (FBS) was acquired from Clark Bioscience
(Claymont, DE, USA). The MTT Cell Proliferation and Cytotoxicity
Assay kit was purchased from Nanjing Keygen Biotech Co., Ltd.
(Nanjing, China). The Alkaline Phosphatase (ALP) Assay kit was
purchased from Nanjing Jiancheng Bioengineering Institute (Nanjing,
China). VK2, vitamin C (VC), and dexamethasone (Dex) were acquired
from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany).
β-Glycerophosphate and alizarin red were purchased from Beijing
Solarbio Science & Technology Co., Ltd. (Beijing, China). The
autophagy inhibitor 3-methyladenine (3-MA) and agonist rapamycin
were purchased from Selleck Chemicals (Houston, TX, USA). Rabbit
anti- microtubule associated protein 1 light chain 3-α (LC3)b
antibody was purchased from Abcam (Cambridge, UK; cat. no.
ab192890). Rabbit anti-β-actin antibody and goat anti-rabbit IgG
antibody were purchased from BIOSS (Beijing, China; cat. nos.
bs-0061R and bs-0295G, respectively). Anti-beclin 1 antibody was
purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA;
cat. no. 3738S) and Cy3-conjugated Affinipure Goat Anti-Rabbit IgG
(H+L) was obtained from Wuhan Sanying Biotechnology (Wuhan, China;
cat. no. SA00009-2).
Cell culture and differentiation
MC3T3-E1 is a preosteoblast cell line. It may be
used to study differential and mineral induction in an experimental
system that mimics a bone-like extracellular matrix (ECM)
environment. The appearance of the ECM is crucial to the bone
formation process. The mouse cranial osteoblast (MC3T3-E1 Subclone
14; American Type Culture Collection CRL-2594) cell line was
obtained from Cell Bank of the Type Culture Collection of the
Chinese Academy of Sciences (Shanghai, China). A subclone with high
differentiation potential was derived from the parental cell line
for studying critical events during osteoblast differentiation and
mineralization. The cells were seeded in 15 cm2 flasks
in α-MEM containing 1% antibiotics and 10% FBS (complete medium),
and the medium was changed every 2 days. Cells were passaged when
they reached 90% confluence. For osteogenic differentiation, the
osteoblasts were first seeded in 6-well plates at a density of
1×104 cells/well in complete medium and cultured for 3
days until the cells reached 70% confluence. To initiate the
differentiation, 3 ml complete medium supplemented with Dex
(10−7 M), β-glycerophosphate (10 mM) and VC (50 µg/ml)
was added to each well, and the differentiation medium was replaced
every 2 days.
Cell viability assay
The cell viability assay was performed to analyze
the possible cytotoxic effects of different concentrations of VK2
(10−8−10−3 M) on MC3T3-E1 cells. The MC3T3-E1
osteoblasts were seeded into 96-well plates. When 70% confluence
was achieved, cells were treated with VK2 at different
concentrations (10−8−10−3 M) for different
durations (1–5 days). At least six replicated wells per sample were
prepared. The cell viability was measured at 48 h using the MTT
Cell Proliferation and Cytotoxicity Assay kit, according to the
manufacturer's protocol.
Quantitative assay of ALP
activity
MC3T3-E1 osteoblasts were plated in 6-well plates at
a density of 2.5×105 cells/well and treated with
different concentrations (10−5, 10−6 and
10−7 M) of the drugs (3MA, VK2 and 3MA+VK2; untreated
controls were also included) for 24 h during the differentiation
protocol, and harvested after 1, 3, 5 and 7 days. Total protein was
extracted from the cells using radioimmunoprecipitation assay
(RIPA) buffer [300 mM NaCl, 50 mM Tri-HCL (pH 7.6), 0.5% Triton
X-100, 2 mM phenylmethylsulfonyl fluoride] and quantified using the
bicinchoninic acid (BCA) assay. ALP activity was measured using a
fluorescence detection kit per the manufacturer's protocol. The
standard curve was plotted using p-nitrophenol, and the ALP
activity of each sample was normalized to the total protein
concentration.
Mineralization analysis
MC3T3-E1 osteoblasts were treated with the different
drugs as described above, during the 7-day differentiation. On the
7th day, the supernatant was discarded, and the cells were washed
twice with PBS and fixed with 4% paraformaldehyde at room
temperature (20°C) for 5 min. Calcium nodes were stained with 0.1%
alizarin red at 37°C for 1 h. For quantitative analysis, cells were
destained with ethylpyridinium chloride for 30 min at room
temperature and transferred to a 96-well plate to measure the
absorbance at 550 nm using a microplate reader.
Western blotting
MC3T3-E1 osteoblasts were seeded into 100 mm dishes
and cultured in complete α-MEM to 70% confluency, following which
the cells were treated with the different drugs as described above.
VK2 was used at the optimal concentration of 10−6 M (1
µM). Total proteins were extracted following lysis of the cells in
RIPA buffer at 4°C for 1 h. The lysates were centrifuged at 20,000
× g for 30 min, and the protein concentration was determined using
a BCA protein quantification kit. Equal amounts of protein (30 µg)
were loaded into each well of a 7.5–15% SDS-PAGE gel and separated
by electrophoresis. The separated protein bands were transferred to
a polypropylene fluoride membrane and, following incubation
overnight at 4°C with primary antibody against LC3b (1:2,000),
beclin 1 (1:1,000) and β-actin (1:2,000), respectively. Next, the
membrane was incubated with goat anti-rabbit secondary antibody
(1:5,000) for 2 h at room temperature. The specific bands were
visualized using an enhanced chemiluminescence detection system
(MF-ChemiBIS 3.2, DNR Bio-Imaging Systems, Ltd., Neve Yamin,
Israel) and imaged with an Alpha Imager HP (ProteinSimple, San
Jose, CA, USA). The band density was quantified using the ImageJ
image processing program (National Institutes of Health, Bethesda,
MD, USA. software version 1.5b).
Immunofluorescence assay
MC3T3-E1 osteoblasts were seeded in 24-well plates
and upon reaching 50% confluency, were treated with 3-MA, VK2 and
VK2+3MA for 1 h. The cells were washed twice with PBS, fixed with
4% paraformaldehyde at room temperature for 15 min, and
permeabilized with 0.5% Triton X-100 for 2 min at room temperature.
After blocking for 2 h with 5% bovine serum albumin (cat. no.
ST023-50 g, Beyotime Institute of Biotechnology, Shanghai, China)
at room temperature, the cells were incubated overnight with
anti-LC3b antibody (1:200) at 4°C. After 1 h incubation with
Cy3-conjugated secondary antibody (1:3,000), the cells were
counterstained with DAPI for 10 min at room temperature. The
stained cells were imaged using confocal fluorescence microscopy
(magnification, ×200, DS-U3 Nikon Eclipse CI; Nikon Corporation,
Tokyo, Japan).
Monodansylcadaverine (MDC)
staining
The cells were seeded in 24-well plates at a density
of 4.0×104 cells/well, and VK2 (1 µM) and the autophagy
inhibitor 3-MA (0.5 mM) were added. The supernatant was removed and
after washing the cells once, they were stained with MDC (a
fluorescent marker of autophagic vacuoles) for 45 min at 37°C. The
stained cells were imaged using an Olympus fluorescence microscope
(magnification, ×200, IX71; Olympus Corporation, Tokyo, Japan).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
MC3T3-E1 osteoblasts were seeded at a density of
5×104 cells/well in a 6-well plate for 3 days until the
cells reached 70% confluence. The cells were treated with the
different drugs as described, and total RNA was extracted using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA), according to the manufacturer's protocol.
A total of ~1 µg of RNA was used in the reverse transcription
reaction using RevertAid First Strand cDNA Synthesis Kit (Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocols.
For gene-specific primed cDNA synthesis, the reaction was conducted
for 60 min at 42°C. Then, for random hexamer primed synthesis,
incubate for 5 min at 25°C. RT was terminated by heating at 70°C
for 5 min. Real-time PCR was performed using the SYBR Green PCR
Master Mix (Applied Biosystems; Thermo Fisher Scientific, Inc.) on
the ABI 7500 Fast Real-Time PCR System (Applied Biosystems 7500
System Sequence Detection System, software version 2.6.2; Thermo
Fisher Scientific, Inc.). The PCR conditions were as follows:
Initial denaturation at 95°C for 15 min, followed by 40 cycles of
denaturation at 94°C for 20 sec, annealing at 60°C for 30 sec and
extension at 72°C for 60 sec. The primer sequences were as follows:
Runt related transcription factor 2 (Runx2) forward,
5′-CCCTGAACTCTGCACCAAGT-3′; Runx2 reverse,
5′-TGGAGTGGATGGATGGGGAT-3′; osteocalcin (OCN) forward,
5′-AGCAGCTTGGCCCAGACCTA-3′; OCN reverse,
5′-TAGCGCCGGAGTCTGTTCACTAC-3′; β-actin forward,
5′-TTCGTTGCCGGTCCACACCC-3′; and β-actin reverse,
5′-GCTTTGCACATGCCGGAGCC-3′.
Statistical analysis
All experiments were repeated at least three times
and the results are expressed as the mean ± standard deviation.
Statistical analysis was performed using SPSS 13.0 software (SPSS,
Inc., Chicago, IL, USA). Different groups were compared by one-way
analysis of variance followed by Tukey's multiple comparisons test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
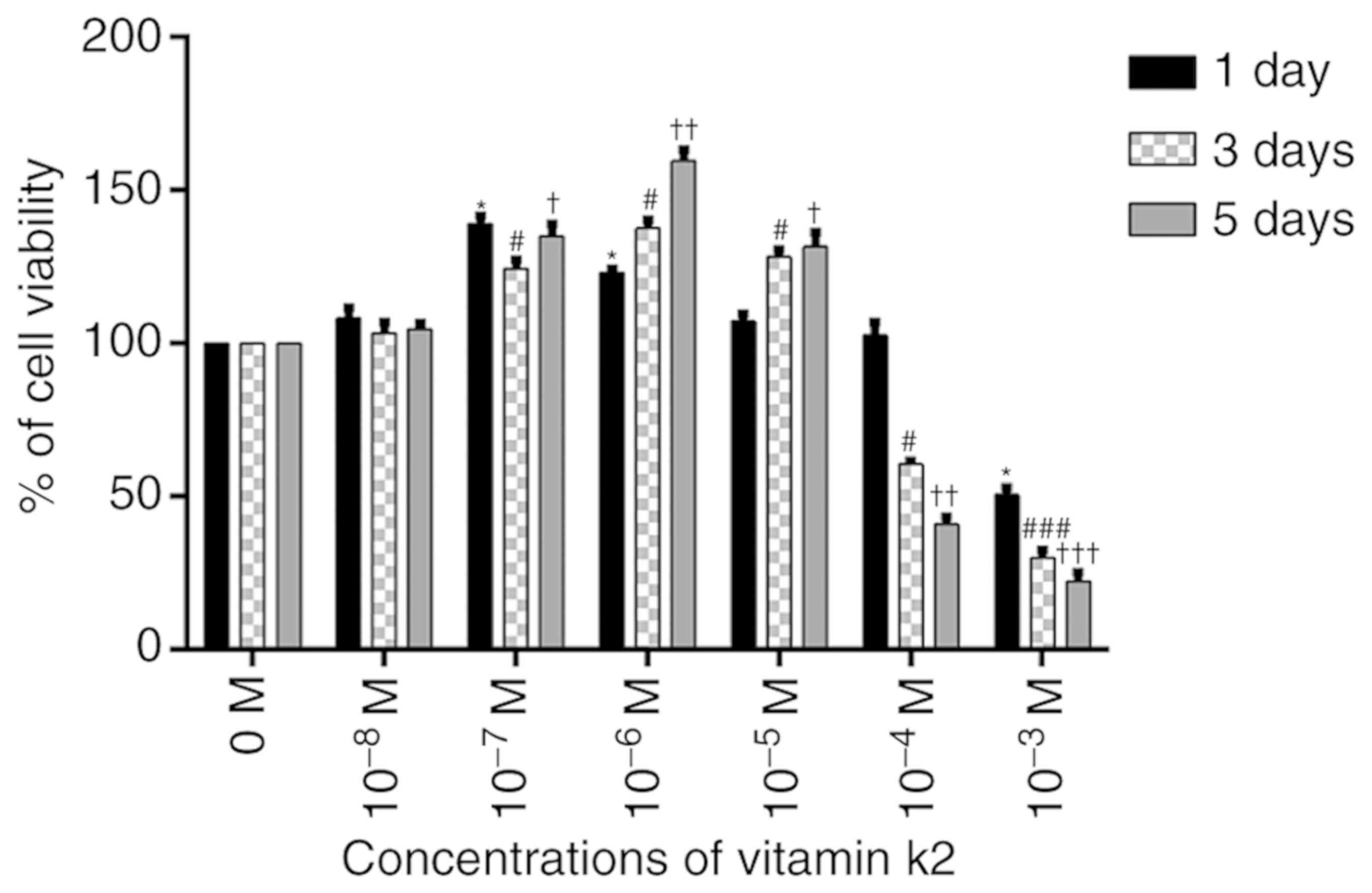
VK2 is not cytotoxic to MC3T3-E1 cells
at concentrations below 10−5 M
An MTT assay was used to analyze the possible
cytotoxic effects of different concentrations of VK2
(10−8−10−3 M) on MC3T3-E1 cells. The cells
were stimulated for different durations (1–5 days). No cytotoxicity
was observed at concentrations below 10−5 M, but cell
viability was reduced in a dose-dependent manner at concentrations
above 10−5 M. Furthermore, compared with the control
group, 10−8 M VK2 did not have a significant effect on
the induction of differentiation and mineralization. Therefore, all
subsequent experiments were performed with VK2 at concentrations
between 10−7 and 10−5 M (Fig. 1).
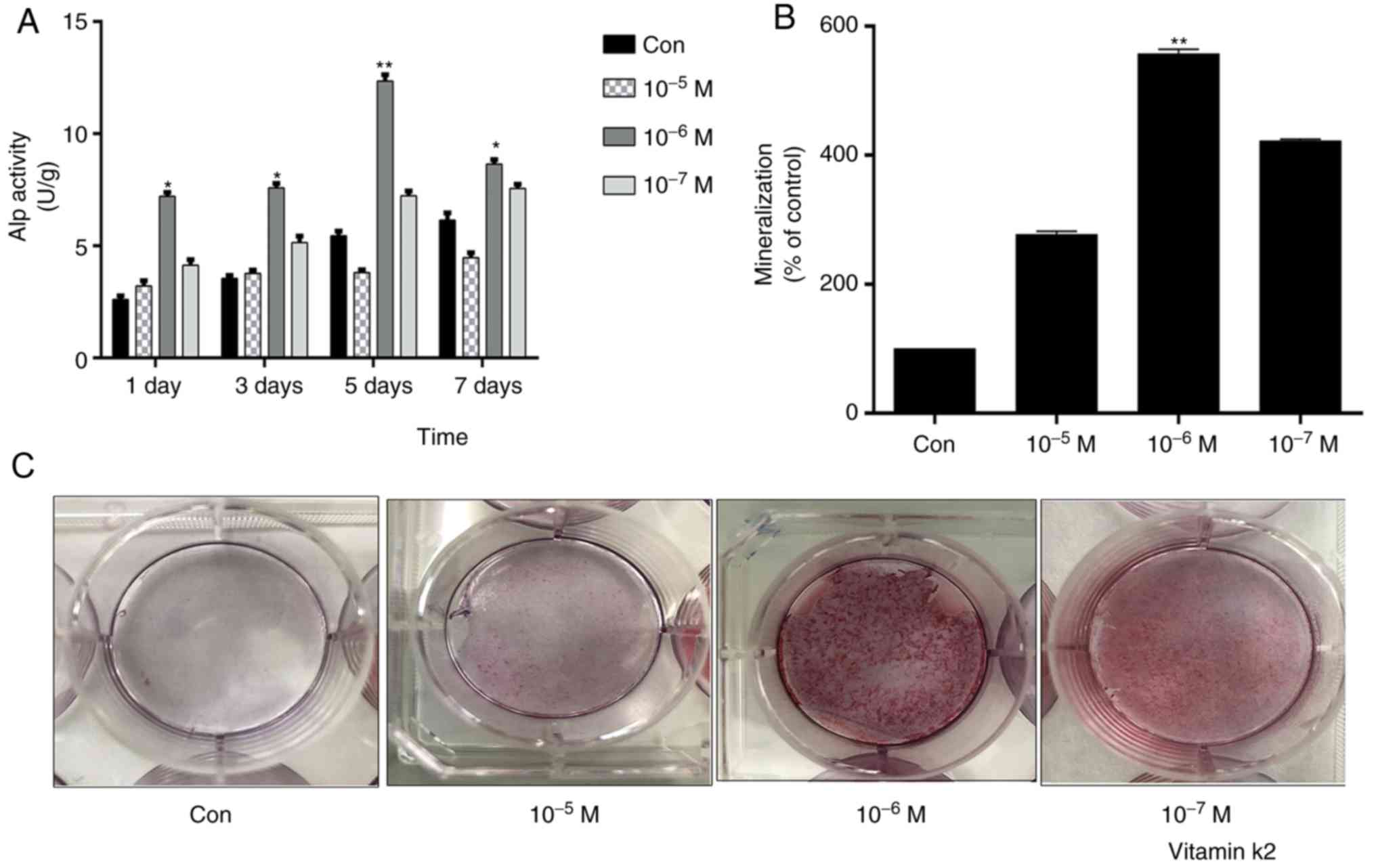
VK2 enhances ALP activity and promotes
MC3T3-E1 osteoblast differentiation and mineralization
To determine the optimal concentration of VK2 that
promotes MC3T3-E1 osteoblast differentiation and mineralization, an
ALP assay and alizarin red staining were performed, respectively,
following treatment of the cells with
10−7−10−5 M VK2 for 1–7 days. Treatment with
1 µM VK2 significantly increased ALP activity, while other
concentrations were not notably effective. In addition, maximum ALP
activity with 1 µM VK2 was observed on the 5th day (Fig. 2A). Bone formation is accompanied by
osteogenic differentiation and mineralization, i.e., calcium
deposition. MC3T3-E1 osteoblasts treated with VK2 (1 µM) for 7 days
exhibited significant calcium deposition, as detected by alizarin
red staining (Fig. 2B and C).
VK2 stimulates autophagy in MC3T3-E1
osteoblasts
To analyze the effects of VK2 on autophagy in
MC3T3-E1 osteoblasts, the cells were treated with VK2 (1 µM) for
0.5–1.5 h (each day for 7 days), various autophagy markers,
including LC3 and Beclin-1, were detected on days 1, 3, 5 and 7 of
differentiation. The ratio of LC3II/LC3I is frequently analyzed to
determine the extent of autophagy. Treatment with VK2 for 0.5–1.5 h
steadily increased LC3I conversion rates in the osteoblasts on days
1, 3, 5 and 7 (Fig. 3A). In
addition, VK2 treatment for 1 h significantly increased the
conversion of LC3I at 1, 3, and 5 days (Fig. 3B-D). Furthermore, the LC3II/LC3I
ratio and Beclin-1 levels were higher on day 5, corresponding to
the strongest ALP activity (Fig.
3G). Autophagy was also evaluated using immunofluorescence and
MDC staining assays. The number of fluorescent bodies and the
intensity increased with VK2 and decreased following treatment with
3-MA+VK2. MDC is a marker for autolysosomes; the results
demonstrated that the VK2 group had stronger fluorescence, and the
fluorescence intensity of the 3-MA+VK2 group was markedly reduced
(Fig. 3H). The inhibitory effect
of 3-MA on VK2-induced autophagy was confirmed with the
significantly decreased LC3II/LC3I ratio in cells treated with
3-MA, and significantly higher ratio in cells treated with
rapamycin. Notably, the conversion rate of LC3II/LC3I in the
VK2+3-MA group was lower compared with the VK2 group (Fig. 4A). These results demonstrated that
VK2 stimulated autophagy in MC3T3-E1 osteoblasts.
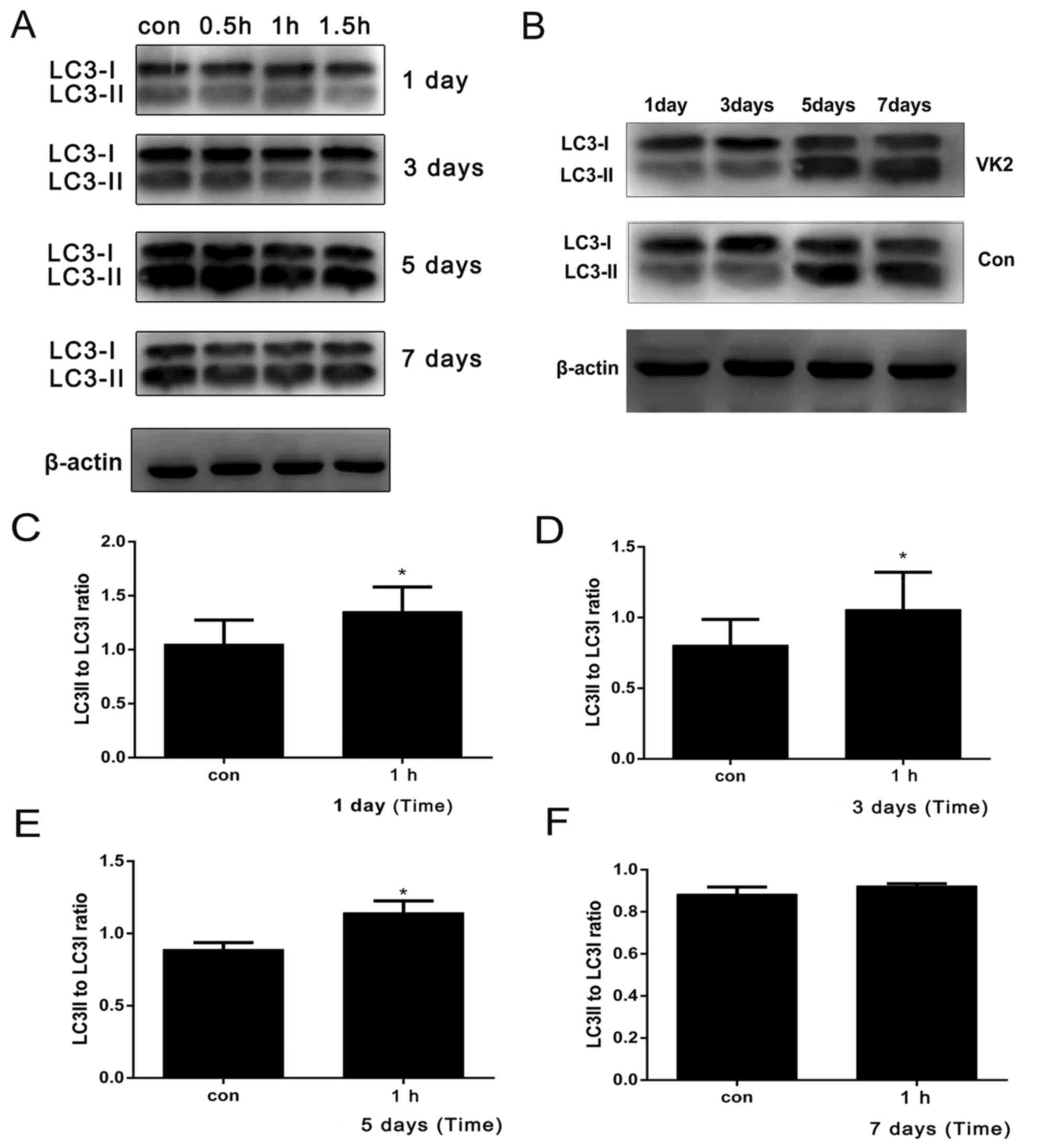 | Figure 3.VK2 stimulates autophagy in MC3T3-E1
osteoblasts. The degree of autophagy was evaluated on the basis of
LC3I to LC3II conversion (LC3II/LC3I). (A) Treatment with 1 µM VK2
for 0.5–1.5 h steadily increased LC3I conversion rates in MC3T3-E1
osteoblasts over (B) 1, 3, 5 and 7 days. Compared with the
untreated control cells, those treated with VK2 for 1 h exhibited a
significantly higher degree of autophagy at (C) 1, (D) 3 and (E) 5
days, although not at (F) 7 days. (G) Higher LC3II/LC3I conversion
rates were observed on day 5 after 0.5, 1 and 1.5 h of treatment
compared with the untreated controls. (H) To further verify the
effect of vitamin K2 on autophagy in MC3T3-E1 osteoblasts, cells
were stained in situ with fluorescence labeled anti-LC3b. The
number of fluorescent bodies and the fluorescence intensity
increased with VK2 and decreased following treatment with 3-MA+VK2
compared with the untreated controls. (I) MDC is a marker for
autolysosomes. MDC staining was used to confirm the abundance of
autophagic vacuoles in VK2-treated cells. In the control group,
weak and diffuse MDC staining was observed throughout the
cytoplasm, and very little punctate staining. In VK2-treated cells,
the MDC staining was visibly enhanced, and its distribution pattern
was altered from diffuse to punctate accumulation. The punctate
staining in the 3-MA+VK2 treatment group was markedly reduced.
Scale bars, 50 µm. The experimental data are expressed as the mean
± standard deviation. Significance analysis of the experimental
data for each group was performed using one-way analysis of
variance and Tukey's multiple comparisons test. *P<0.05,
**P<0.01 vs. respective control group. VK2, vitamin K2; ALP,
alkaline phosphatase; 3-MA, 3-methyladenine; LC3, microtubule
associated protein 1 light chain 3-α; Con, control; MDC,
monodansylcadaverine. |
 | Figure 4.Association between autophagy and
differentiation in MC3T3-E1 osteoblasts. (A) To further verify the
inhibitory effect of 3-MA on autophagy mediated by VK2, the western
blotting results demonstrated that the conversion rate of
LC3II/LC3I in cells treated with the autophagy inhibitor 3-MA was
significantly decreased, while that of rapamycin was significantly
higher. The conversion rate of LC3II/LC3I in VK2+3-MA group was
lower compared with the VK2 group. **P<0.01 vs. control group
and ‡P<0.05 vs. VK2 group. (B) Compared with the
untreated control cells, the 3-MA treated osteoblasts exhibited a
significant inhibition of ALP activity, and the ALP activity of the
VK2+3-MA treated cells was significantly lower compared with the
VK2-treated cells. *P<0.05 vs. control group at 3 days;
#P<0.05 vs. control group at 5 days. (C) The mRNA
expression levels of LC3II, ALP, OCN and Runx2 in VK2-treated cells
increased. *P<0.05 vs. LC3II control; #P<0.05 vs.
ALP control; ††P<0.01 vs. Runx2 control;
‡‡P<0.01 vs. OCN control. (D) In addition, alizarin
red staining indicated that 3-MA markedly inhibited the osteogenic
differentiation induced by VK2. The experimental data are expressed
as the mean ± standard deviation. Significance analysis of the
experimental data for each group was performed using one-way
analysis of variance and Tukey's multiple comparisons test.
*P<0.05, **P<0.01 vs. control group. VK2, vitamin K2; 3-MA,
3-methyladenine; Ra, rapamycin; Con, control; ALP, alkaline
phosphatase; Runx2, runt related transcription factor 2; OCN,
osteocalcin; LC3, microtubule associated protein 1 light chain
3-α. |
3-MA inhibits the osteogenic
differentiation and mineralization induced by VK2
To confirm the role of autophagy in VK2-induced
osteogenic differentiation and mineralization, the ALP assay and
alizarin red staining were performed on cells treated additionally
with 3-MA. The 3-MA treated osteoblasts (3-MA+VK2 group) exhibited
significantly reduced ALP activity compared with the VK2 group
(Fig. 4B). In addition, 3-MA
slightly inhibited the mRNA expression of osteogenic
differentiation markers compared with the control (Fig. 4C). 3-MA inhibited the
mineralization induced by VK2 (Fig.
4D). These results demonstrated that autophagy induced by VK2
stimulation is involved in osteogenic differentiation and
mineralization.
Discussion
MC3T3-E1 osteoblasts were used to study the
mechanism of VK2-mediated amelioration of osteoporosis symptoms, as
this cell line is routinely used to study the osteogenic
characteristics of bone differentiation and mineralization in
vitro. Biomarkers including ALP, RUNX2 and OCN are upregulated
in the bone matrix during osteogenic differentiation and
mineralization, and are thus used to assess the these processes in
in vitro models (20,21).
Osteoblasts were stimulated with different concentrations of VK2,
and osteogenic differentiation and mineralization were evaluated by
ALP activity and alizarin red staining, respectively; the
concentration of 1 µM induced the most significant effect on
osteoblast differentiation. The present results are consistent with
previous studies that have correlated VK2 with increased osteoblast
differentiation and mineralization (22–24).
However, the specific mechanism of VK2 in promoting bone metabolism
has not yet been elucidated. It was demonstrated that VK2 induced
autophagy during osteoblast differentiation and mineralization
using the markers LC3 and beclin 1. Studies have increasingly
demonstrated a role for autophagy in the occurrence and development
of osteoporosis. Studies on human genome-wide association data
report that autophagy-related genes are associated with
osteoporosis (15,25). In addition, the degree of autophagy
decreases along with the expression of LC3-II, beclin 1 and unc-51
like autophagy activating kinase 1, while apoptosis and p62
expression increases during osteoporosis (16,26,27).
Decreased autophagy likely promotes osteoporosis by increasing
oxidative stress (28,29). Furthermore, certain studies have
demonstrated that autophagy is associated with the differentiation
of osteoblasts. It has been demonstrated that autophagy-associated
proteins are involved in osteoblast differentiation and bone
formation, and other studies have reported that autophagy affects
osteoblast mineralization and bone cell network structure (30,31).
Finally, mouse knockout models of autophagy genes illustrate a
significant decrease in trabecular bone volume, trabecular number
and trabecular thickness (30,32).
The present study also confirmed that VK2 may stimulate autophagy
in MC3T3 cells to promote differentiation and mineralization.
The use of MC3T3 cells is the primary limitation of
the present study; these cells are appropriate for the purpose of
this research and numerous studies have used MC3T3-E1 to assess
differentiation and mineralization (33–36).
However, there are limitations associated with performing
experiments with a single cell line. The present results may be
confirmed by using an additional cell line, such as mesenchymal
stem cells, and additional factors may be used to evaluate
autophagy in the future, including other types of autophagy
inhibitors to exclude off-target effects, and in vivo
experiments. Finally, the specific pathways associated with
autophagy were completely explored. Therefore, more in-depth
mechanistic studies are required.
In conclusion, the present findings indicated that
VK2 promotes autophagy during the differentiation and
mineralization of osteoblasts, which may be a potential therapeutic
target for osteoporosis.
Acknowledgements
Not applicable.
Funding
This study was supported by the Science and
Technology Project of Shenyang (grant no. F16-102-4-00).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
WL and QL conceived and designed the study. WL, SZ
and YL performed the experiments. WL and JL analyzed the data. WL
was a major contributor in writing the manuscript. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Tella SH and Gallagher JC: Prevention and
treatment of postmenopausal osteoporosis. J Steroid Biochem Mol
Biol. 142:155–170. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Glaser DL and Kaplan FS: Osteoporosis.
Definition and clinical presentation. Spine (Phila Pa 1976). 22 (24
Suppl):12S–16S. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Edwards MH, Dennison EM, Aihie Sayer A,
Fielding R and Cooper C: Osteoporosis and sarcopenia in older age.
Bone. 80:126–130. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Diab DL and Watts NB: Postmenopausal
osteoporosis. Curr Opin Endocrinol Diabetes Obes. 20:501–509. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Clarke BL: Anti-sclerostin antibodies:
Utility in treatment of osteoporosis. Maturitas. 78:199–204. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Levine JP: Identification, diagnosis, and
prevention of osteoporosis. Am J Manag Care. 17 (Suppl
6):S170–S176. 2011.PubMed/NCBI
|
|
7
|
Palermo A, Tuccinardi D, D'Onofrio L,
Watanabe M, Maggi D, Maurizi AR, Greto V, Buzzetti R, Napoli N,
Pozzilli P and Manfrini S: Vitamin K and osteoporosis: Myth or
reality? Metabolism. 70:57–71. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nelsestuen GL, Zytkovicz TH and Howard JB:
The mode of action of vitamin K. Identification of
gamma-carboxyglutamic acid as a component of prothrombin. J Biol
Chem. 249:6347–6350. 1974.PubMed/NCBI
|
|
9
|
Stenflo J, Fernlund P, Egan W and
Roepstorff P: Vitamin K dependent modifications of glutamic acid
residues in prothrombin. Proc Natl Acad Sci USA. 71:2730–2733.
1974. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Berkner KL: The vitamin K-dependent
carboxylase. Annu Rev Nutr. 25:127–149. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hauschka PV, Lian JB and Gallop PM: Direct
identification of the calcium-binding amino acid,
gamma-carboxyglutamate, in mineralized tissue. Proc Natl Acad Sci
USA. 72:3925–3929. 1975. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Coutu DL, Wu JH, Monette A, Rivard GE,
Blostein MD and Galipeau J: Periostin, a member of a novel family
of vitamin K-dependent proteins, is expressed by mesenchymal
stromal cells. J Biol Chem. 283:17991–18001. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rubinacci A: Expanding the functional
spectrum of vitamin K in bone. Focus on: ‘Vitamin K promotes
mineralization, osteoblast to osteocyte transition, and an
anti-catabolic phenotype by {gamma}-carboxylation-dependent and
-independent mechanisms’. Am J Physiol Cell Physiol.
297:C1336–1338. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Atkins GJ, Welldon KJ, Wijenayaka AR,
Bonewald LF and Findlay DM: Vitamin K promotes mineralization,
osteoblast-to-osteocyte transition, and an anticatabolic phenotype
by {gamma}-carboxylation-dependent and -independent mechanisms. Am
J Physiol Cell Physiol. 297:C1358–1367. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pierrefite-Carle V, Santucci-Darmanin S,
Breuil V, Camuzard O and Carle GF: Autophagy in bone: Self-eating
to stay in balance. Ageing Res Rev. 24:206–217. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Parzych KR and Klionsky DJ: An overview of
autophagy: Morphology, mechanism, and regulation. Antioxid Redox
Signal. 20:460–473. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
White E, Mehnert JM and Chan CS:
Autophagy, Metabolism, and Cancer. Clin Cancer Res. 21:5037–5046.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mizushima N and Levine B: Autophagy in
mammalian development and differentiation. Nat Cell Biol.
12:823–830. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mizushima N, Levine B, Cuervo AM and
Klionsky DJ: Autophagy fights disease through cellular
self-digestion. Nature. 451:1069–1075. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yamaguchi M: The role of regucalcin in
bone homeostasis: Involvement as a novel cytokine. Integr Biol
(Camb). 6:258–266. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Raouf A and Seth A: Ets transcription
factors and targets in osteogenesis. Oncogene. 19:6455–6463. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yamaguchi M and Weitzmann MN: Vitamin K2
stimulates osteoblastogenesis and suppresses osteoclastogenesis by
suppressing NF-κB activation. Int J Mol Med. 27:3–14.
2011.PubMed/NCBI
|
|
23
|
Ichikawa T, Horie-Inoue K, Ikeda K,
Blumberg B and Inoue S: Vitamin K2 induces phosphorylation of
protein kinase A and expression of novel target genes in
osteoblastic cells. J Mol Endocrinol. 39:239–247. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang YL, Yin JH, Ding H, Zhang W, Zhang
CQ and Gao YS: Protective effect of VK2 on glucocorticoid-treated
MC3T3-E1 cells. Int J Mol Med. 39:160–166. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nollet M, Santucci-Darmanin S, Breuil V,
Al-Sahlanee R, Cros C, Topi M, Momier D, Samson M, Pagnotta S,
Cailleteau L, et al: Autophagy in osteoblasts is involved in
mineralization and bone homeostasis. Autophagy. 10:1965–1977. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen K, Yang YH, Jiang SD and Jiang LS:
Decreased activity of osteocyte autophagy with aging may contribute
to the bone loss in senile population. Histochem Cell Biol.
142:285–295. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pan F, Liu XG, Guo YF, Chen Y, Dong SS,
Qiu C, Zhang ZX, Zhou Q, Yang TL, Guo Y, et al: The
regulation-of-autophagy pathway may influence Chinese stature
variation: Evidence from elder adults. J Hum Genet. 55:441–447.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jilka RL, Weinstein RS, Parfitt AM and
Manolagas SC: Quantifying osteoblast and osteocyte apoptosis:
Challenges and rewards. J Bone Miner Res. 22:1492–1501. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li L, Tan J, Miao Y, Lei P and Zhang Q:
ROS and Autophagy: Interactions and molecular regulatory
mechanisms. Cell Mol Neurobiol. 35:615–621. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu F, Fang F, Yuan H, Yang D, Chen Y,
Williams L, Goldstein SA, Krebsbach PH and Guan JL: Suppression of
autophagy by FIP200 deletion leads to osteopenia in mice through
the inhibition of osteoblast terminal differentiation. J Bone Miner
Res. 28:2414–2430. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xi G, Rosen CJ and Clemmons DR: IGF-I and
IGFBP-2 stimulate AMPK activation and autophagy, which are required
for osteoblast differentiation. Endocrinology. 157:268–281. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chang KH, Sengupta A, Nayak RC, Duran A,
Lee SJ, Pratt RG, Wellendorf AM, Hill SE, Watkins M, Gonzalez-Nieto
D, et al: p62 is required for stem cell/progenitor retention
through inhibition of IKK/NF-kB/Ccl4 signaling at the bone marrow
macrophage-osteoblast niche. Cell Rep. 9:2084–2097. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu Y, Huang L, Hao B, Li H, Zhu S, Wang
Q, Li R, Xu Y and Zhang X: Use of an osteoblast overload damage
model to probe the effect of icariin on the proliferation,
differentiation and mineralization of MC3T3-E1 cells through the
Wnt/β-catenin signalling pathway. Cell Physiol Biochem.
41:1605–1615. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kim EC, Kim TH, Jung JH, Hong SO and Lee
DW: Enhanced osteogenic differentiation of MC3T3-E1 on
rhBMP-2-immobilized titanium via click reaction. Carbohydr Polym.
103:170–178. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
He Y, Wang S, Mu J, Dai L, Zhang Z, Sun Y,
Shi W and Ge D: Synthesis of polypyrrole nanowires with positive
effect on MC3T3-E1 cell functions through electrical stimulation.
Mater Sci Eng C Mater Biol Appl. 71:43–50. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mu W and Wang Z, Ma C, Jiang Y, Zhang N,
Hu K, Li L and Wang Z: Metformin promotes the proliferation and
differentiation of murine preosteoblast by regulating the
expression of sirt6 and oct4. Pharmacol Res. 129:462–474. 2018.
View Article : Google Scholar : PubMed/NCBI
|