Introduction
Glioma is the most common type of primary brain
tumor, with an annual incidence of ~6.6/100,000 individuals in the
USA (1). Approximately half of all
glioma cases are classified as glioblastoma, which is the most
malignant type of brain tumor (1).
Despite current treatment standards involving maximal safe
resection, followed by radiotherapy with concomitant and adjuvant
temozolomide (TMZ) (2), the median
survival of patients with glioblastoma is only 14–17 months
(2,3). As a recognized disease with high
heterogeneity, survival variability has been observed in cases of
glioblastoma with similar clinical and histological features
(4). Traditional
clinicopathological factors, including histological grade, age and
Karnofsky performance status (KPS), do not appear to be sufficient
for precise outcome prediction (3,4).
However, with the characterization of specific genetic alterations
using advanced sequencing and microarray technologies, several
genes and multi-gene expression-based molecular signatures have
been discovered. These have since been used for the identification
of novel glioblastoma subtypes, and may also have better prognostic
significance (3–5).
Proteins are primary functional effectors in cells
that represent the vast majority of prognostic biomarkers currently
used for glioma (3). Over 68% of
the human genome is transcribed to long noncoding RNAs (lncRNAs), a
class of transcripts >200 nucleotides in length that lack
protein-coding ability. These have emerged as important regulators
of tissue physiology and disease processes, including tumorigenesis
(6). lncRNAs can regulate the
expression levels of oncogenes or tumor suppressors through various
mechanisms, including chromatin modification, transcriptional
control and post-transcriptional processing, and affect various
aspects of cellular homeostasis (7). The expression pattern of lncRNAs is
highly tissue- and cell type-specific, a characteristic that may be
applied for accurate molecular cancer subclassification and outcome
prediction (7). A successfully
established example is the lncRNA prostate cancer antigen 3, which
is currently used for early diagnosis and treatment response
surveillance in prostate cancer (8).
Reports have revealed that lncRNAs are aberrantly
expressed in glioma tissue relative to normal brain tissue
(9,10). Some lncRNAs have been implicated in
glioma proliferation, apoptosis, motility and stemness maintenance
(11,12). However, the detailed mechanisms and
clinical application of most lncRNAs remain largely unknown. By
virtue of the constant updating of gene annotation databases, such
as GENCODE and RefSeq, more gene probes from previously published
microarray studies are likely to be repurposed as lncRNAs,
providing novel information for investigating the expression
profiles and prognostic value of lncRNAs in cancer.
In the present study, the updated gene annotations
were adopted to conduct lncRNA profiling on publicly available
microarray datasets of glioma, and a four-lncRNA signature was
identified to be able to predict the outcome of glioblastoma.
Insulin-like growth factor binding protein 7-antisense 1
(IGFBP7-AS1), one of the survival-associated lncRNAs, was further
investigated by bioinformatics analysis and functional
experiments.
Materials and methods
Glioma datasets preparation
Gene expression profiles generated by the Affymetrix
HG-U133 Plus 2.0 microarray platform (Affymetrix; Thermo Fisher
Scientific, Inc.) and corresponding clinical data were downloaded
from the publicly available Gene Expression Omnibus (GEO;
www.ncbi.nlm.nih.gov/geo) and the
Repository for Molecular Brain Neoplasia Data (Rembrandt) project
(13). The Rembrandt database is
available at the Georgetown Database of Cancer web portal
(https://gdoc.georgetown.edu). Rembrandt
database (13) includes 227 grade
IV glioblastomas, 84 grade III gliomas, 99 grade II gliomas and 28
normal brain (NB) samples, according to the world health
organization grading system (14).
GEO dataset GSE4290 (15) includes
81 glioblastomas, 31 grade III gliomas, 45 grade II gliomas and 23
NB. All these samples were used for lncRNA expression analysis. To
avoid interference from different treatment regimens on the
analysis of survival-associated lncRNAs, only glioblastoma samples
from patients who received post-operative radiation with or without
chemotherapy were analyzed, including 108 samples from the GEO
dataset GSE16011 (4), 69 from the
GEO dataset GSE7696 (16) and 78
from Rembrandt database. The clinical characteristics of the 255
patients with glioblastoma used for survival analysis were
described in Table SI.
Microarray data processing and
analysis
The raw Affymetrix microarray expression data were
processed and normalized using the Robust Multichip Average
algorithm (17). The custom Chip
Definition File was used to reorganize probes to Entrez Gene
identifiers (version 21; National Center for Biotechnology
Information, National Institutes of Health) and exclude inaccurate
or wrongly annotated probes (18).
To avoid systematic error across different experiments, each
dataset was standardized independently by the Z-score method to
transform the expression of each gene into having a mean of 0 and a
standard deviation of 1 (19).
Similar to previous methods (9),
genes on microarray platforms were identified as lncRNAs according
to their RefSeq transcript ID (National Center for Biotechnology
Information, National Institutes of Health) and Ensembl gene ID
(20) from the updated NetAffx
annotation files (HG-U133_Plus_2 Annotations, Release 36;
Affymetrix; Thermo Fisher Scientific, Inc.) and then filtered by
removing pseudogenes, ribosomal RNAs, transfer RNAs, microRNAs
(miRNAs) and other short noncoding RNAs. Finally, a gene list
containing 1,895 lncRNAs was created for further analysis. Among
these lncRNAs, 1,495 transcripts were annotated on RefSeq and
Ensembl databases and 400 transcripts were annotated selectively on
RefSeq (Table SII).
Hierarchical clustering analysis was performed to
divide patients based on multi-lncRNA expression levels using
centered correlation metrics and the average linkage method. Gene
set enrichment analysis (GSEA) was implemented according to lncRNA
median expression, with the gene sets from the Molecular Signatures
Database (software.broadinstitute.org/gsea) (21).
Cell culture and transient
transfection
The human glioma cell lines U87 (cat. no. HTB-14,
glioblastoma from unknown origin) and U251 were purchased from the
Chinese Academy of Sciences. The cells were cultured in Dulbecco's
modified Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific,
Inc.) containing 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.), 100 U/ml penicillin and 50 µg/ml streptomycin,
and were incubated at 37°C in an atmosphere containing 5%
CO2.
The small interfering RNA (siRNA) targeting
IGFBP7-AS1 and nonspecific negative control (NC) sequence were
synthesized by Shanghai GenePharma Co., Ltd. Cell transient
transfection was performed using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) 24 h after seeding
cells, at 70% confluency. Transfection complexes were prepared
according to the manufacturer's protocol and added to the cells to
a final oligonucleotide concentration of 100 nM. The transfection
medium was replaced 6 h post-transfection. Cells were incubated for
2 days prior to subsequent experimentation. The siRNA sequences are
presented in Table I.
 | Table I.Small interfering RNA sequences of
long noncoding RNA IGFBP7-AS1 and nonspecific NC. |
Table I.
Small interfering RNA sequences of
long noncoding RNA IGFBP7-AS1 and nonspecific NC.
| Gene symbol | Positive-sense
strand (5′-3′) | Antisense strand
(5′-3′) |
|---|
| IGFBP7-AS1 |
CUAAGUUUCUGGAAGAUAAAG |
UUAUCUUCCAGAAACUUAGAA |
| NC |
UUCUCCGAACGUGUCACGUTT |
ACGUGACACGUUCGGAGAATT |
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was extracted using RNAiso
Plus® (Takara Bio, Inc.) according to the manufacturer's
protocol. Total RNA (1 µg) was used to synthesize cDNA using the
PrimeScript® RT reagent kit with gDNA Eraser (both from
Takara Bio, Inc.) according to the manufacturer's protocol. Then, 1
µl cDNA was used to determine the quantification cycle (Cq) value
of each sample using the SYBR® Premix Ex Taq II kit
(Takara Bio, Inc.) in a LightCycler 480 system (Roche Applied
Science). GAPDH served as the reference gene. Relative gene
expression was calculated using the 2−ΔΔCq method
(22). The PCR primer sequences
are in Table II.
| Table II.Primer sequences of IGFBP7-AS1 and
the reference gene GAPDH. |
Table II.
Primer sequences of IGFBP7-AS1 and
the reference gene GAPDH.
| Gene symbol | Forward primer
(5′-3′) | Reverse primer
(5′-3′) |
|---|
| IGFBP7-AS1 |
GGAAAGCTCTTCCTGACCCA |
CCTGCTAATCTCAGGCAGCA |
| GAPDH |
CCCATCACCATCTTCCAGGAG |
GTTGTCATGGATGACCTTGGC |
Cell viability assay
Cells were seeded into 96-well plates at a density
of 1,000-1,500 cells/well, for 1–5 days. The cell viability in each
well was assessed at 1, 2, 3, 4 and 5 days using the Cell Counting
kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.) according to
the manufacturer's protocol. Absorbance was measured at 450 nm to
calculate the number of viable cells.
Wound healing and Transwell
assays
A scratch was made on 80% confluent monolayer cell
cultures, and culture medium was replaced with serum-free DMEM. The
healing process was monitored for 24 h, and the percentage of wound
closure was calculated using ImageJ software (version 1.51;
National Institutes of Health).
In total, 5×104 cells in 200 µl
serum-free DMEM medium were seeded onto the upper chamber of
Transwell apparatus (Costar; Corning, Inc.) with a Matrigel-coated
membrane. A total of 600 µl DMEM containing 10% FBS was added to
the lower chamber as a chemoattractant. After a 12-h incubation at
37°C in a 5% CO2. atmosphere, the cells that invaded the
lower chamber through the membrane were fixed with 4% methanol for
30 min at room temperature, stained with 0.1% crystal violet for 15
min at room temperature and counted under a light microscope
(magnification, ×100).
Statistical analysis
SPSS (version 20.0; IBM Corp.) and GraphPad Prism
(version 7.0; GraphPad Software Inc.) were used for statistical
analysis. ANOVA was performed for multiple comparisons followed by
Dunnett's post-hoc test. Data are presented as the mean ± SD, where
appropriate. Student's t-test was used to compare two groups.
χ2 test was used to compare qualitative variables.
Survival-related lncRNAs were screened via univariate Cox
regression analysis using Biometric Research Branch-Array Tools
version 4.5.0 (National Center Institute, National Institutes of
Health) and a permutation test method with 10,000 permutations
(23). The selected lncRNAs were
analyzed using a multivariate Cox regression analysis in the
training set and a risk score formula was established including
each identified lncRNA, and weighted using the estimated regression
coefficients in the multivariate model (24,25).
Survival differences were evaluated by the Kaplan-Meier method, and
curves were compared using a log-rank test. Time-dependent receiver
operating characteristic (ROC) curves were created with the
R-package ‘survivalROC’ and Nearest Neighbor Estimation method
(26). P<0.05 was considered to
indicate a statistically significant difference.
Results
Four lncRNAs, tumor protein
P73-antisense 1 (TP73-AS1), IGFBP7-AS1, PAX interacting protein
1-antisense 2 (PAXIP1-AS2) and long intergenic non-protein coding
RNA 672 (LINC00672), are associated with glioblastoma outcomes
Among the 159 glioblastoma patients in the GSE16011
dataset, 108 patients received post-operative radiation without
chemotherapy. These 108 patients were used as a training set to
screen survival-associated lncRNAs. A panel of four lncRNAs
strongly associated with overall survival was identified via
univariate Cox regression analysis, when the permutation was
P<0.005, as listed in Table
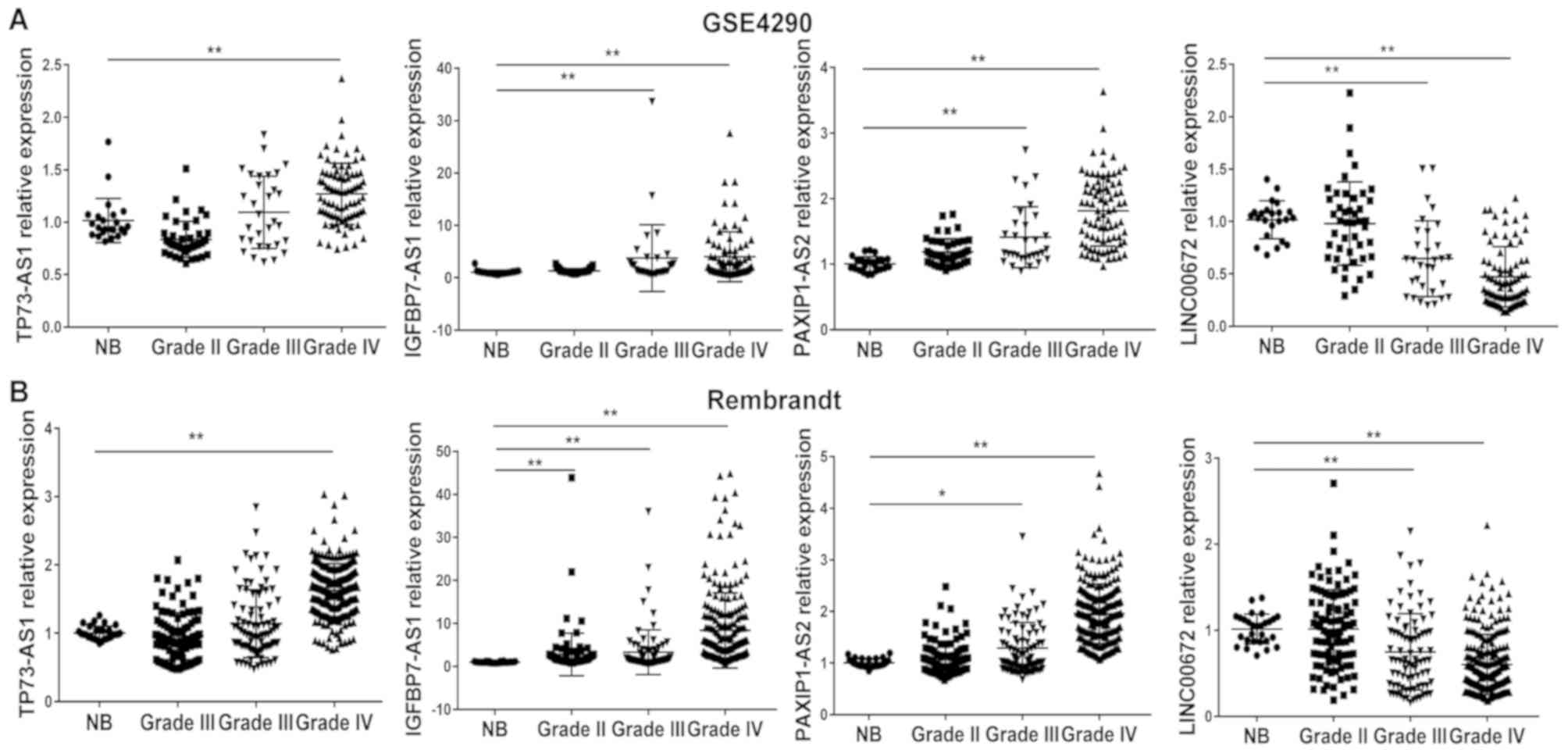
III. In addition, the upregulation of lncRNAs with a positive
regression coefficient (TP73-AS1, IGFBP7-AS1 and PAXIP1-AS2), and
the downregulation of lncRNAs with a negative coefficient
(LINC00672) were observed in glioblastoma tissues compared with in
normal brain tissues in the GSE4290 and Rembrandt datasets
(Fig. 1).
| Table III.Long noncoding RNAs associated with
overall survival of 108 cases of glioblastoma in the training
set. |
Table III.
Long noncoding RNAs associated with
overall survival of 108 cases of glioblastoma in the training
set.
| Gene symbol | Permutation
P-value | Hazard ratio | Coefficient | Ensembl ID | Entrez Gene ID | RefSeq ID |
|---|
| TP73-AS1 | 0.001 | 3.229 | 1.172 |
ENSG00000227372 | 57212 | NR_033708/ |
|
|
|
|
|
|
| NR_033709/ |
|
|
|
|
|
|
| NR_033710/ |
|
|
|
|
|
|
| NR_033711/ |
|
|
|
|
|
|
| NR_033712 |
| IGFBP7-AS1 | <0.001 | 1.447 | 0.369 |
ENSG00000245067 | 255130 | NR_034081 |
| PAXIP1-AS2 | <0.001 | 2.537 | 0.931 |
ENSG00000214106 | 100132707 | NR_024476/ |
|
|
|
|
|
|
| NR_024477 |
| LINC00672 | 0.003 | 0.605 | −0.502 |
ENSG00000263874 | 100505576 | NR_038847 |
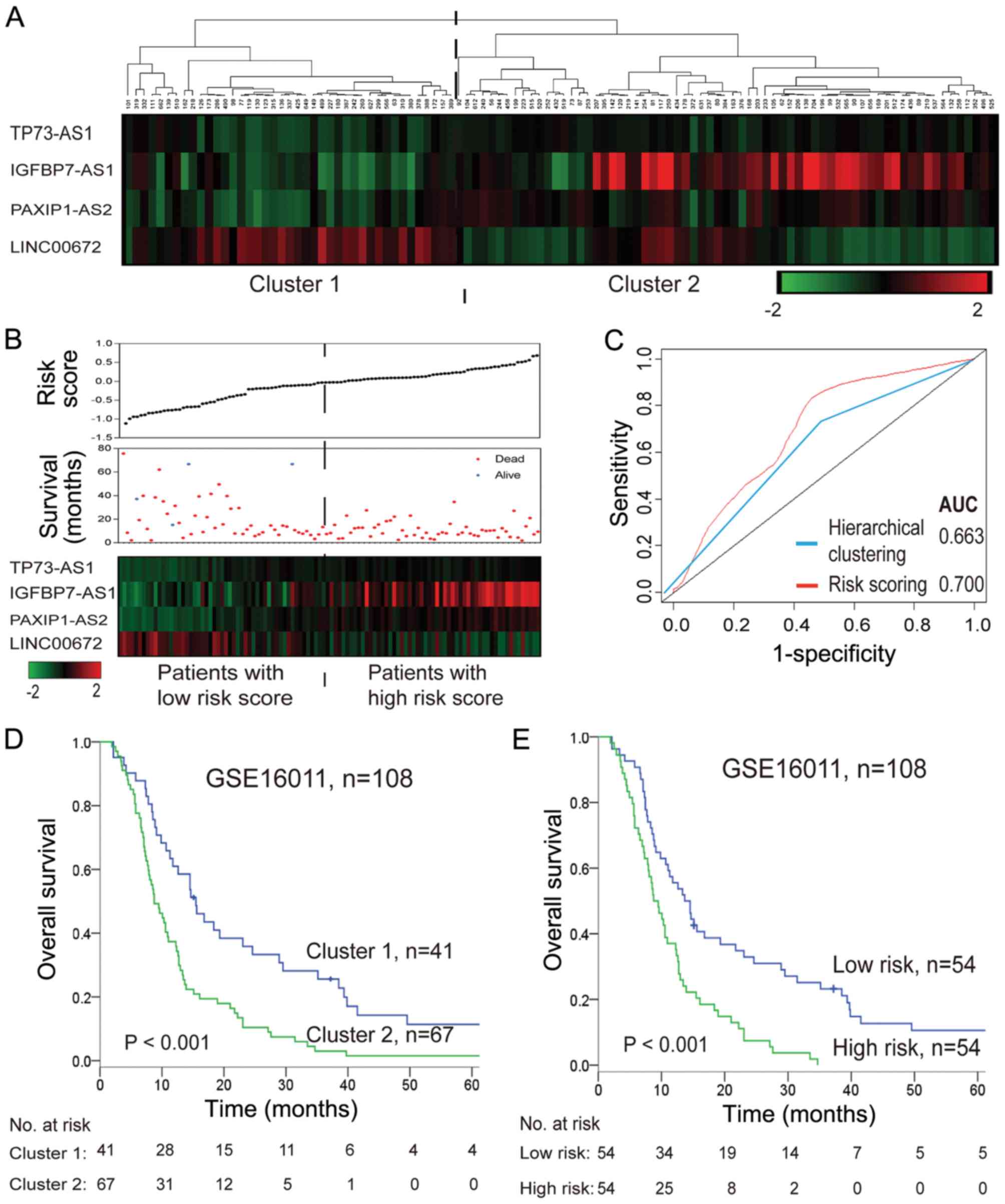
As shown in Fig. 2A and
D, hierarchical clustering with these four survival-associated
lncRNAs subdivided patients in the training set into two clusters
with distinct survival rates (median 8.76 months vs. 15.48 months).
The four identified lncRNAs were integrated into a molecular
signature using a risk score formula to weigh their prognostic
effects. The risk score was calculated as follows: Risk score =
(0.397× expression value of TP73-AS1) + (0.191× expression value of
IGFBP7-AS1) + (0.468× expression value of PAXIP1-AS2) + (−0.129×
expression value of LINC00672). The risk score of each patient was
calculated, and then patients were divided into a high-risk or
low-risk group according to the median risk score (−0.0254) of the
training set. Patients in the high-risk group exhibited
significantly shorter survival times than those in the low-risk
group (median 9.48 months vs. 13.68 months; log-rank P<0.001;
Fig. 2B and E). The correlation of
the risk score with overall survival was also significant when
assessed in the univariate model (Table IV). A larger area under the curve
in time-dependent ROC analysis was achieved using the risk scoring
method, illustrating that this constituted a better algorithm for
predicting outcomes, with good sensitivity and specificity compared
with hierarchical clustering (Fig.
2C).
| Table IV.Univariate and multivariate Cox
regression analyses for overall survival of patients with
glioblastoma in public datasets. |
Table IV.
Univariate and multivariate Cox
regression analyses for overall survival of patients with
glioblastoma in public datasets.
| A, GSE16011
(training set, n=108) |
|---|
|
|---|
|
| Univariate
model | Multivariate
model |
|---|
|
|
|
|
|---|
| Variables | HR | 95% CI | P-value | HR | 95% CI | P-value |
|---|
| lncRNA-risk
score | 2.716 | 1.712–4.308 |
<0.001a | 1.906 | 1.124–3.230 | 0.017a |
| Sex | 0.856 | 0.565–1.298 |
0.465 | 0.841 | 0.552–1.282 | 0.421 |
| Age | 1.033 | 1.016–1.051 |
<0.001a | 1.019 | 0.999–1.039 | 0.057 |
| KPS | 0.980 | 0.967–0.994 |
0.004a | 0.984 | 0.970–0.998 | 0.029a |
|
| B, Entire
independent dataset (n=147) |
|
|
| Univariate
model | Multivariate
model |
|
|
|
|
|
Variables | HR | 95% CI | P-value | HR | 95% CI | P-value |
|
| lncRNA-risk
score | 2.560 | 1.586–4.133 |
<0.001a | 2.450 | 1.539–3.901 |
<0.001a |
| Sex | 0.893 | 0.610–1.307 |
0.560 | 1.030 | 1.011–1.050 | 0.658 |
| Age | 1.029 | 1.011–1.048 |
0.002a | 0.917 | 0.625–1.346 | 0.002a |
|
| C, GSE7696
dataset (n=69) |
|
|
| Univariate
model | Multivariate
model |
|
|
|
|
|
Variables | HR | 95% CI | P-value | HR | 95% CI | P-value |
|
| lncRNA-risk
score | 3.709 | 1.463–9.403 | 0.006a | 2.600 | 0.899–7.520 |
0.042a |
| Sex | 0.887 | 0.491–1.602 | 0.690 | 0.540 | 0.457–1.507 | 0.540 |
| Age | 1.041 | 1.008–1.075 | 0.015a | 1.026 | 0.989–1.063 | 0.167 |
|
| D, Rembrandt
dataset (n=78) |
|
|
| Univariate
model | Multivariate
model |
|
|
|
|
|
Variables | HR | 95% CI | P-value | HR | 95% CI | P-value |
|
| lncRNA-risk
score | 2.197 | 1.216–3.969 | 0.009a | 2.451 | 1.335–4.501 | 0.004a |
| Sex | 0.950 | 0.575–1.571 | 0.842 | 1.189 | 0.706–2.002 | 0.516 |
| Age | 1.031 | 1.007–1.055 | 0.011a | 1.028 | 1.003–1.053 | 0.027a |
| KPS | 0.994 | 0.988–1.000 | 0.060 | 0.993 | 0.987–1.000 | 0.043a |
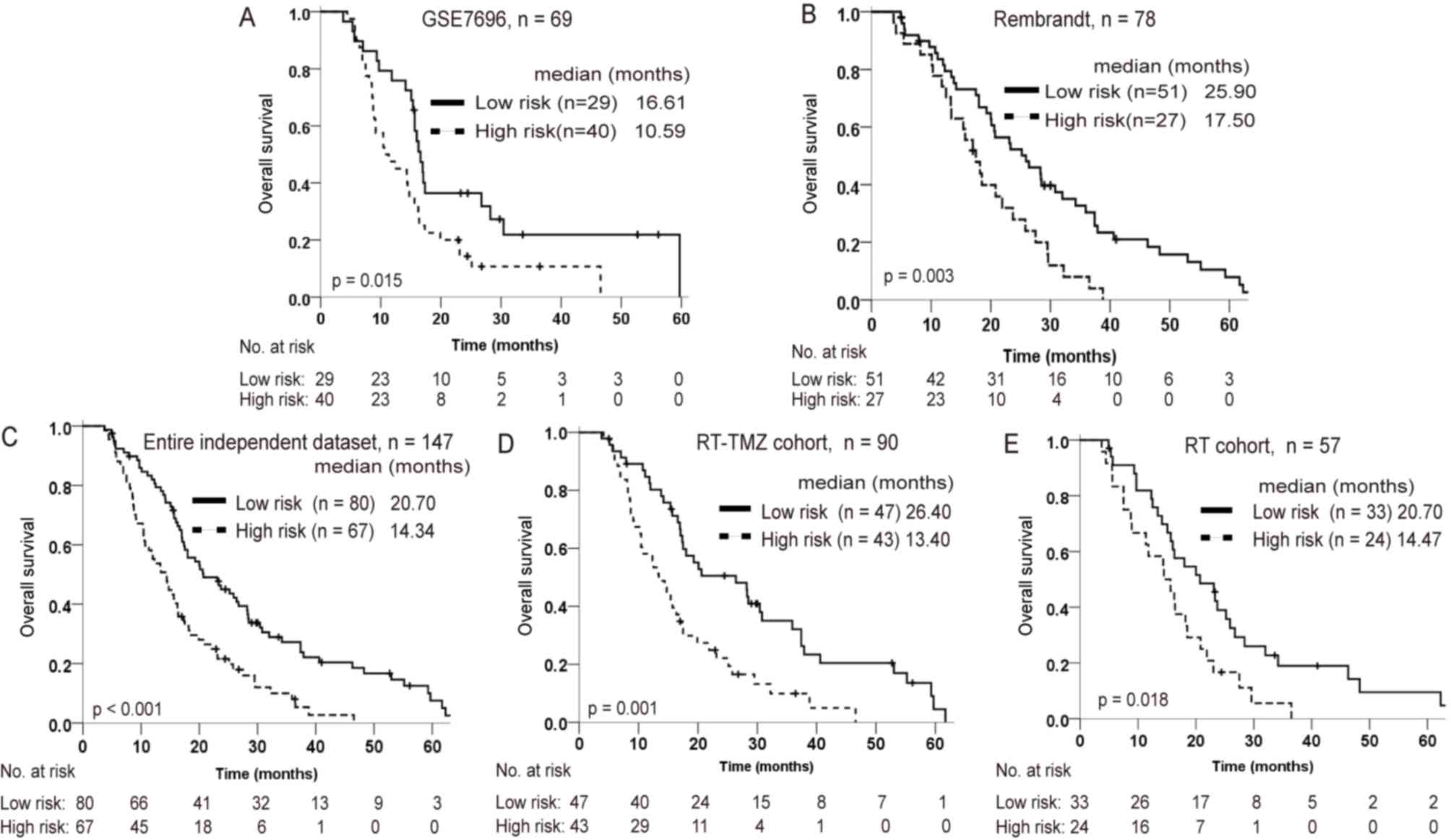
Further validation of the four-lncRNA
signature in two independent datasets
From the GSE7696 and Rembrandt datasets, 69 and 78
patients with glioblastoma, who had received postoperative
radiation, respectively, were considered as independent datasets
for validating the prognostic value of the lncRNA signature. Of
these, a total of 57 patients accepted radiotherapy only (RT
cohort), and 90 patients accepted radiotherapy combined with TMZ
chemotherapy (RT-TMZ cohort). Using the same cutoff point for the
lncRNA-risk score as for the training set, patients in the GSE7696
and Rembrandt datasets could be also divided into distinct survival
subgroups (Fig. 3A and B). Risk
score-based subclassification of the entire independent cohort
(both the GSE7697 and Rembrandt patients) had similar results,
whether the patients accepted TMZ treatment or not. Furthermore,
for patients with a high-risk score, radiation combined with TMZ
did not appear to significantly prolong the survival time relative
to radiotherapy alone (Fig.
3C-E).
Prognostic value of the four-lncRNA
signature is independent of conventional clinicopathological
factors
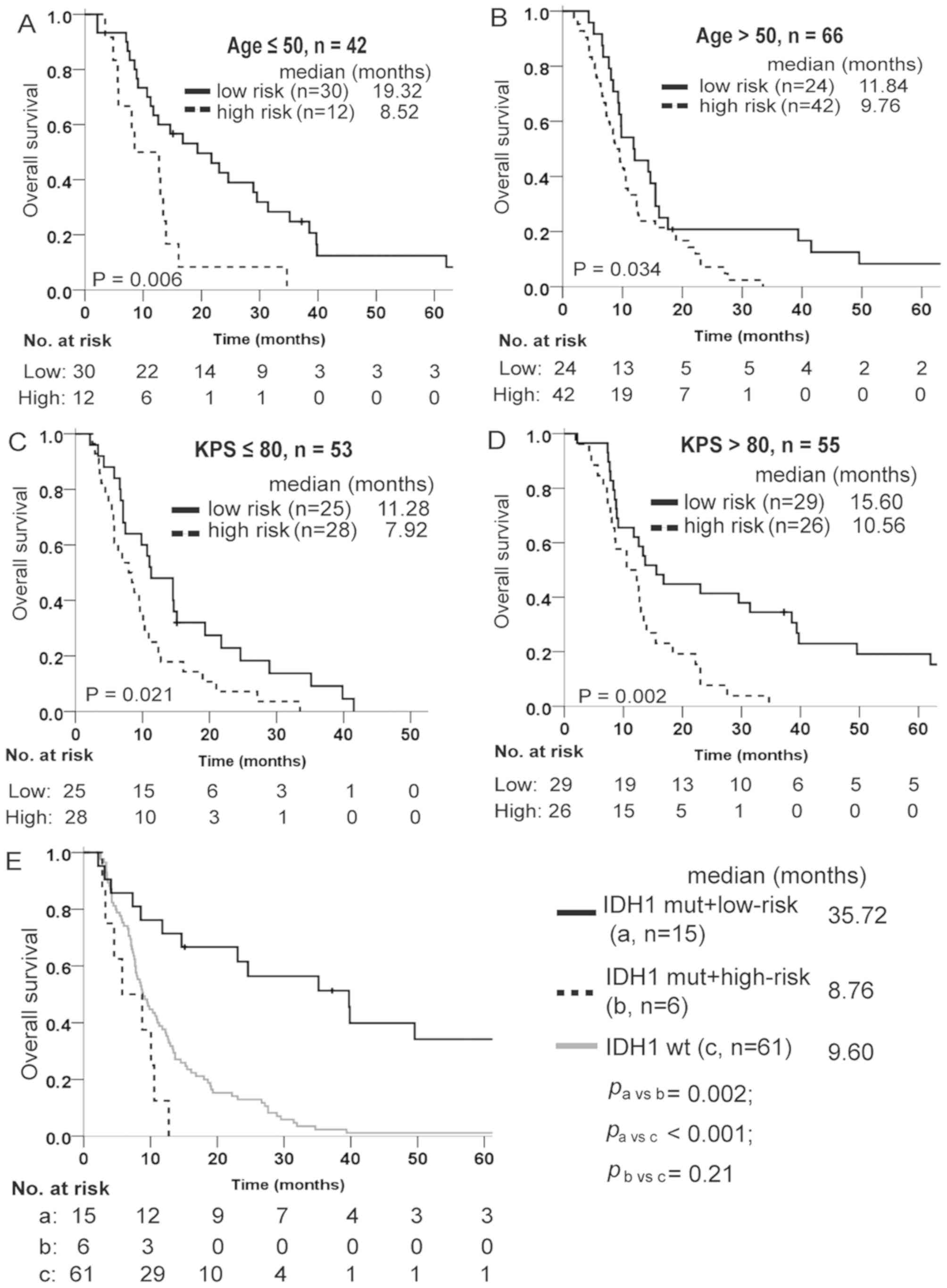
Multivariate Cox regression analysis was adopted to
estimate whether the prognostic value of the lncRNA signature was
independent of the clinical characteristics of patients with
glioblastoma. As shown in Table
IV, the lncRNA-risk score remained significantly associated
with overall survival when adjusted by age and KPS, when available,
in every cohort. Data stratification analysis was also performed to
assess the prognostic value of this signature with respect to the
same clinical factors. For this, patients in the training set were
first stratified into a younger group (age ≤50) and an older group
(age >50), then into a poor performance group (KPS ≤80) and a
good performance group (KPS >80). The results indicated that the
lncRNA signature remained a useful prognostic indicator within each
age and KPS stratum (Fig.
4A-D).
The isocitrate dehydrogenase 1 (IDH1) mutation is
one of the most valuable biomarkers for glioma diagnosis and
prognosis. In the GSE16011 dataset, the lncRNA signature could
predict different outcomes of glioblastoma carrying the IDH1
mutation. Patients exhibiting mutations in IDH1 gene and high-risk
signature exhibited a poor survival, similarly to those with
wild-type IDH1 (Fig. 4E).
Knockdown of IGFBP7-AS1 inhibits the
viability and invasion of U87 and U251 glioma cells
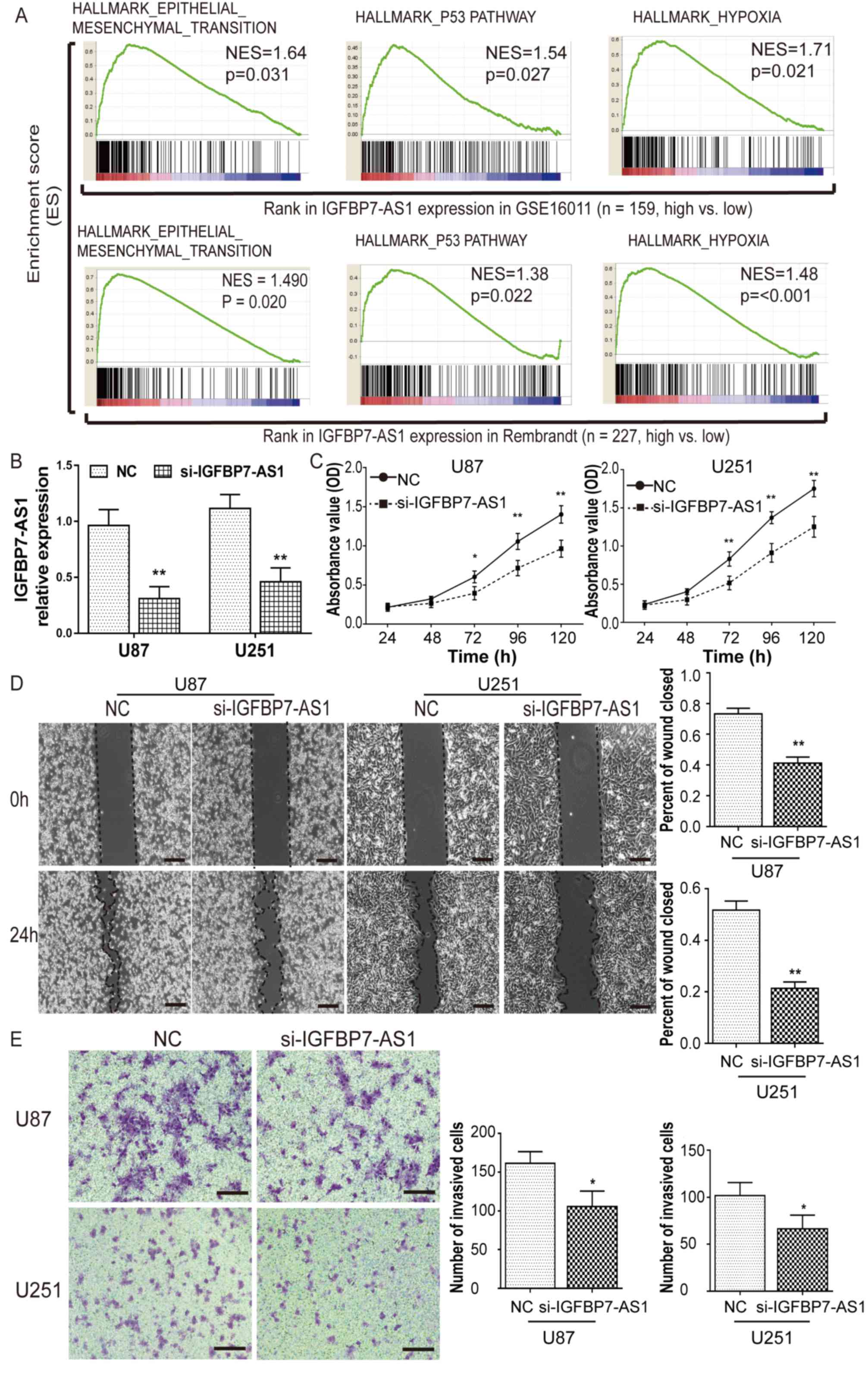
As the oncogenic function of TP73-AS1 in glioma has
been reported in recent studies (27–29),
GSEA was performed to identify the biological processes and
pathways of IGFBP7-AS1, PAXIP1-AS2 and LINC00672 lncRNAs involved
in glioma pathogenesis by dividing glioblastoma cases from the
GSE16011 and Rembrandt datasets into high- and low-expression
groups according to the median lncRNA expression. The results
demonstrated that the ‘epithelial-mesenchymal transition’, ‘p53
pathway’ and ‘hypoxia’ gene sets were significantly enriched in
glioblastoma samples with high IGFBP7-AS1 expression relative to
those with low expression (Fig.
5A). IGFBP7-AS1 expression was also the most upregulated of the
four prognostic lncRNAs in glioblastoma compared with NB (Fig. 1). Therefore, the role of IGFBP7-AS1
with regards to glioma viability and invasion was further assessed
following the transfection of U87 and U251 glioma cell lines with a
siRNA targeting IGFBP7-AS1 (Fig.
5B). IGFBP7-AS1 siRNA markedly reduced U87 and U251 cell
viability compared with in NC-transfected cells, as determined
using a CCK-8 assay (Fig. 5C).
After knockdown of IGFBP7-AS1, a wound-healing assay showed that
the wound closure percentage was visibly reduced, and a Transwell
assay revealed that the number of cells that invaded through the
Matrigel matrix was significantly decreased (Fig. 5D and E). These results further
demonstrated that IGFBP7-AS1 may serve a role in the regulation of
glioma cell migration and invasion.
Discussion
lncRNAs are a new class of noncoding gene regulators
involved in cancer pathogenesis and prognosis (6,7).
Although analyses of global expression patterns and functional
characterization of lncRNAs have been performed, the molecular
mechanisms and clinical applications of lncRNAs in the context of
glioblastoma remain unclear (9–12).
Previous studies demonstrated that lncRNA expression profiles can
be obtained by mining existing microarray data, as thousands of
probes on commonly used arrays are likely to be repurposed as
lncRNAs (9,30). Moreover, the number of probes
available to map lncRNAs is increasing rapidly due to the updating
of lncRNA databases, such as GENCODE, in order to provide a more
comprehensive documentation of sequence information, functional
annotation and expression profiles (31). By combining the microarray mining
method and updated gene annotations, the present study identified
1,895 lncRNAs on the Affymetrix platform. To the best of our
knowledge, more than half of these genes were not previously
annotated as lncRNAs; therefore, the present study provided novel
information for investigating lncRNA profiles in cancer, and a
four-lncRNA signature was identified with potential for predicting
outcomes in glioblastoma.
The current standard treatment for glioblastoma is
maximal safe resection followed by radiotherapy and TMZ
chemotherapy. Postoperative radio-chemotherapy confers better
prognosis than surgery alone, or than radiation without
chemotherapy (2). To avoid
interference from different treatment regimens, only the patients
who had received postoperative radiotherapy were chosen for
survival analysis in the training set. The results revealed that
four lncRNAs were significantly associated with overall survival in
108 cases of glioblastoma. Upregulation of TP73-AS1, IGFBP7-AS1 and
PAXIP1-AS2, with a positive regression coefficient, and
downregulation of LINC00672, with a negative coefficient, was
observed in glioblastoma samples relative to normal brain tissue.
The expression trends were also associated with the malignancy
grades of glioma, indicating their potential use in glioma
diagnosis and prognosis.
Based on the expression levels of the four lncRNAs,
patients with glioblastoma in the training set could be divided
into two groups with distinct survival by both hierarchical
clustering and a risk scoring method. Risk scoring is a widely used
and effective method to establish a multi-gene expression-based
molecular signature to predict outcomes of various types of cancer
(24,25). By performing ROC analysis, risk
scoring was determined as a better estimator of overall survival,
with better sensitivity and specificity compared with hierarchical
clustering. Overall, patients in the training set with high-risk
lncRNA signatures exhibited poorer survival. The reproducibility of
this signature was validated in additional independent datasets,
including patients who received radiotherapy alone, and those
treated with radiation combined with TMZ. The results indicated
that the prognostic value of the lncRNA signature in cases of
glioblastoma was not restricted by different treatment options. In
addition, the prognostic value of this signature was independent of
well-known clinical prognostic factors, including age and KPS,
further attesting to its potential usefulness in clinical practice.
It is worth noting that, compared with radiotherapy alone,
postoperative radiation combined with TMZ may have prolonged the
survival of patients with glioblastoma, who exhibited a low-risk
lncRNA signature. For high-risk patients, TMZ failed to improve
prognosis. This observation implies that the roles of these lncRNAs
in glioma progression may be associated with treatment
resistance.
IDH1 and IDH2 mutations were previously identified
as key biomarkers for molecular pathology and outcome prediction in
glioblastoma. The R132H mutation in the IDH1 gene is the most
frequent type of IDH mutation (14,32).
Mutations in IDH mainly occur in lower grade glioma and recurrent
glioblastoma, which have improved survival rates compared with
primary glioblastoma. In primary glioblastoma, patients exhibiting
IDH mutations also possess better survival rates than patients
without mutation (32). IDH
mutation can induce global DNA hypermethylation and deregulate
genes, including lncRNAs, inhibiting cellular differentiation,
which is involve in tumor initiation and progression (32,33).
These genetic findings promoted the emergence of therapeutic
approaches aimed to target the mutant form of IDH in glioma and
other types of cancer (32). In
the GSE16011 dataset, the expression levels of the four prognostic
lncRNAs were significantly different between glioblastoma cases
with and without the IDH1 mutation. A correlation between these
lncRNA levels and the IDH mutation has also been reported in a
RNA-sequencing report (10).
Furthermore, in the present study, IDH1 mutant glioblastoma with a
high-risk lncRNA signature exhibited poor survival, which was
comparable with IDH wild type glioblastoma. This may have clinical
implications for identifying at-risk patients, among those with IDH
mutant glioblastoma, who may therefore require more intensive
treatment, such as IDH-targeted therapy in clinical trials
(32). These results also
indicated the possibility that these lncRNAs are involved in IDH
mutation-associated glioma tumorigenesis. Further studies are
needed to elucidate the relationship between the IDH mutation and
lncRNA deregulation.
The functions of lncRNAs are closely associated with
their expression level, as they do not encode proteins (30). The expression of TP73-AS1 is
increased in a variety of malignancies, including hepatocellular
carcinoma, breast cancer and glioblastoma, and the upregulation of
TP73-AS1 also predicts the poor prognosis of cancer (28,34,35).
TP73-AS1 may combine with miRNAs to regulate glioma growth,
apoptosis and invasion (27–29).
To the best of our knowledge, the remaining three lncRNAs were
identified as survival predictors in glioma for the first time in
the present study. However, in endometrial cancer, LINC00672 has
been reported to be downregulated and involved in p53-mediated gene
suppression and malignant progression (36). With respect to IGFBP7-AS1 and
PAXIP1-AS2, no functional studies on cancer have been reported so
far, to the best of our knowledge. IGFBP7-AS1 and PAXIP1-AS2 are
the antisense transcripts of IGFBP7 and PAXIP1 genes, respectively.
IGFBP7 has been reported to inhibit insulin-like growth factor
signaling, and induce senescence and apoptosis in hepatocellular
carcinoma (37), whereas PAXIP1
has been reported to participate in DNA damage repair and
chemosensitivity in lung cancer (38). It is worth noting that antisense
lncRNAs could affect cellular homeostasis by interacting with their
positive sense gene, and regulating their expression and function.
For instance, the natural antisense transcript of zinc finger E-box
binding homeobox 2 (ZEB2) interacts with ZEB2 mRNA to modulate
alternative splicing, enhance protein translation and promote tumor
invasion (39). However, their
exact functions in glioma are still unclear. GSEA was thus
performed, and it revealed that ‘p53 pathway’ and
‘epithelial-mesenchymal transition’ were enriched in glioblastoma
cases with higher IGFBP7-AS1 expression. p53 is a vital gene,
involved in regulation of the cell cycle and tumor growth (40). Epithelial-mesenchymal transition
(EMT) is a complex cellular process contributing to the switch of
epithelial cells into motile mesenchymal cells, which promotes the
invasion and metastasis of epithelial tumors (41). Notably, reports have described an
EMT-like phenomenon in glioma, and its association with tumor
invasiveness and poorer prognosis (41,42).
The present results revealed that IGFBP7-AS1 may affect glioma cell
survival by regulating tumor growth and migration. Moreover,
IGFBP7-AS1 expression was the most upregulated of the four
prognostic lncRNAs in the glioblastoma cases analyzed, which
indicated that IGFBP7-AS1 may be comparatively more important and
more deserving of research focus. Further functional experiments
demonstrated that knockdown of IGFBP7-AS1 reduced the viability,
migration and invasion of U87 and U251 glioma cells, providing a
mechanistic explanation for the prognostic ability of IGFBP7-AS1.
IGFBP7-AS1 may therefore constitute a novel target for future
molecular therapy development.
In conclusion, a combination of microarray mining
and gene re-annotation may be considered an effective method to
investigate the expression profiles of lncRNAs in cancer. A novel
four-lncRNA signature was identified as a composite biomarker for
glioblastoma outcome prediction. The role of IGFBP7-AS1 in
influencing glioma cell viability, migration and invasion was
demonstrated with further in vitro experiments. The present
study provided a feasible method and novel information for
understanding the functions of lncRNAs in glioma pathogenesis and
prognosis.
Supplementary Material
Supporting Data
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was funded by The National Natural
Science Foundation of China (grant no. 81372692) and National Key
Clinical Specialist Construction Program of China.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
DL and LY designed this study and wrote the
manuscript. DL analyzed and interpreted the datasets. JL and HL
performed the cell experiments. SQ analyzed the data. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ostrom QT, Gittleman H, Xu J, Kromer C,
Wolinsky Y, Kruchko C and Barnholtz-Sloan JS: CBTRUS statistical
report: Primary brain and other central nervous system tumors
diagnosed in the united states in 2009–2013. Neuro Oncol. 18 (Suppl
5):v1–v75. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Stupp R, Mason WP, van den Bent MJ, Weller
M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn
U, et al: Radiotherapy plus concomitant and adjuvant temozolomide
for glioblastoma. N Engl J Med. 352:987–996. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Reifenberger G, Wirsching HG,
Knobbe-Thomsen CB and Weller M: Advances in the molecular genetics
of gliomas-implications for classification and therapy. Nat Rev
Clin Oncol. 14:434–452. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gravendeel LA, Kouwenhoven MC, Gevaert O,
de Rooi JJ, Stubbs AP, Duijm JE, Daemen A, Bleeker FE, Bralten LB,
Kloosterhof NK, et al: Intrinsic gene expression profiles of
gliomas are a better predictor of survival than histology. Cancer
Res. 69:9065–9072. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Verhaak RGW, Hoadley KA, Purdom E, Wang V,
Qi Y, Wilkerson MD, Miller CR, Ding L, Golub T, Mesirov JP, et al:
Integrated genomic analysis identifies clinically relevant subtypes
of glioblastoma characterized by abnormalities in PDGFRA, IDH1,
EGFR, and NF1. Cancer Cell. 17:98–110. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Iyer MK, Niknafs YS, Malik R, Singhal U,
Sahu A, Hosono Y, Barrette TR, Prensner JR, Evans JR, Zhao S, et
al: The landscape of long noncoding RNAs in the human
transcriptome. Nat Genet. 47:199–208. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Huarte M: The emerging role of lncRNAs in
cancer. Nat Med. 21:1253–1261. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hessels D and Schalken JA: The use of PCA3
in the diagnosis of prostate cancer. Nat Rev Urol. 6:255–261. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang X, Sun S, Pu JK, Tsang AC, Lee D,
Man VO, Lui WM, Wong ST and Leung GK: Long non-coding RNA
expression profiles predict clinical phenotypes in glioma.
Neurobiol Dis. 48:1–8. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Reon BJ, Anaya J, Zhang Y, Mandell J,
Purow B, Abounader R and Dutta A: Expression of lncRNAs in
low-grade gliomas and glioblastoma multiforme: An in silico
analysis. PLoS Med. 13:e10021922016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang XQ and Leung GK: Long non-coding
RNAs in glioma: Functional roles and clinical perspectives.
Neurochem Int. 77:78–85. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang X, Kiang KM, Zhang GP and Leung GK:
Long non-coding RNAs dysregulation and function in glioblastoma
stem cells. Noncoding RNA. 1:69–86. 2015.PubMed/NCBI
|
|
13
|
Madhavan S, Zenklusen JC, Kotliarov Y,
Sahni H, Fine HA and Buetow K: Rembrandt: Helping personalized
medicine become a reality through integrative translational
research. Mol Cancer Res. 7:157–167. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Louis DN, Perry A, Reifenberger G, von
Deimling A, Figarella-Branger D, Cavenee WK, Ohgaki H, Wiestler OD,
Kleihues P and Ellison DW: The 2016 world health organization
classification of tumors of the central nervous system: A summary.
Acta Neuropathol. 131:803–820. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sun LX, Hui AM, Su Q, Vortmeyer A,
Kotliarov Y, Pastorino S, Passaniti A, Menon J, Walling J, Bailey
R, et al: Neuronal and glioma-derived stem cell factor induces
angiogenesis within the brain. Cancer Cell. 9:287–300. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Murat A, Migliavacca E, Gorlia T, Lambiv
WL, Shay T, Hamou MF, de Tribolet N, Regli L, Wick W, Kouwenhoven
MC, et al: Stem cell-related ‘self-renewal’ signature and high
epidermal growth factor receptor expression associated with
resistance to concomitant chemoradiotherapy in glioblastoma. J Clin
Oncol. 26:3015–3024. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Irizarry RA, Hobbs B, Collin F,
Beazer-Barclay YD, Antonellis KJ, Scherf U and Speed TP:
Exploration, normalization, and summaries of high density
oligonucleotide array probe level data. Biostatistics. 4:249–264.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dai M, Wang P, Boyd AD, Kostov G, Athey B,
Jones EG, Bunney WE, Myers RM, Speed TP, Akil H, et al: Evolving
gene/transcript definitions significantly alter the interpretation
of GeneChip data. Nucleic Acids Res. 33:e1752005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cheadle C, Vawter MP, Freed WJ and Becker
KG: Analysis of microarray data using z score transformation. J Mol
Diagn. 5:73–81. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hubbard T, Barker D, Birney E, Cameron G,
Chen Y, Clark L, Cox T, Cuff J, Curwen V, Down T, et al: The
Ensembl genome database project. Nucleic Acids Res. 30:38–41. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Subramanian A, Tamayo P, Mootha VK,
Mukherjee S, Ebert BL, Gillette MA, Paulovich A, Pomeroy SL, Golub
TR, Lander ES and Mesirov JP: Gene set enrichment analysis: A
knowledge-based approach for interpreting genome-wide expression
profiles. Proc Natl Acad Sci USA. 102:15545–15550. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Simon R, Lam A, Li MC, Ngan M, Menenzes S
and Zhao Y: Analysis of gene expression data using BRB-ArrayTools.
Cancer Inform. 3:11–17. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lossos IS, Czerwinski DK, Alizadeh AA,
Wechser MA, Tibshirani R, Botstein D and Levy R: Prediction of
survival in diffuse large-B-cell lymphoma based on the expression
of six genes. N Engl J Med. 350:1828–1837. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang XQ, Sun S, Lam KF, Kiang KM, Pu JK,
Ho AS, Lui WM, Fung CF, Wong TS and Leung GK: A long non-coding RNA
signature in glioblastoma multiforme predicts survival. Neurobiol
Dis. 58:123–131. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Heagerty PJ, Lumley T and Pepe MS:
Time-dependent ROC curves for censored survival data and a
diagnostic marker. Biometrics. 56:337–344. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xiao S, Wang R, Wu X, Liu W and Ma S: The
long noncoding RNA TP73-AS1 interacted with miR-124 to modulate
glioma growth by targeting inhibitor of apoptosis-stimulating
protein of p53. DNA Cell Biol. 37:117–125. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang R, Jin H and Lou F: The long
non-coding RNA TP73-AS1 interacted with miR-142 to modulate brain
glioma growth through HMGB1/RAGE pathway. J Cell Biochem.
119:3007–3016. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang JB, Liu FH, Chen JH, Ge HT, Mu LY,
Bao HB and Lin ZG: Identifying survival-associated modules from the
dysregulated triplet network in glioblastoma multiforme. J Cancer
Res Clin Oncol. 143:661–671. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Du Z, Fei T, Verhaak RG, Su Z, Zhang Y,
Brown M, Chen Y and Liu XS: Integrative genomic analyses reveal
clinically relevant long noncoding RNAs in human cancer. Nat Struct
Mol Biol. 20:908–913. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lagarde J, Uszczynska-Ratajczak B,
Carbonell S, Pérez-Lluch S, Abad A, Davis C, Gingeras TR, Frankish
A, Harrow J, Guigo R and Johnson R: High-throughput annotation of
full-length long noncoding RNAs with capture long-read sequencing.
Nat Genet. 49:1731–1740. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Dang L, Yen K and Attar EC: IDH mutations
in cancer and progress toward development of targeted therapeutics.
Ann Oncol. 27:599–608. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Turcan S, Rohle D, Goenka A, Walsh LA,
Fang F, Yilmaz E, Campos C, Fabius AW, Lu C, Ward PS, et al: IDH1
mutation is sufficient to establish the glioma hypermethylator
phenotype. Nature. 483:479–483. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li S, Huang Y, Huang Y, Fu Y, Tang D, Kang
R, Zhou R and Fan XG: The long non-coding RNA TP73-AS1 modulates
HCC cell proliferation through miR-200a-dependent HMGB1/RAGE
regulation. J Exp Clin Cancer Res. 36:512017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yao J, Xu F, Zhang D, Yi W, Chen X, Chen G
and Zhou E: TP73-AS1 promotes breast cancer cell proliferation
through miR-200a-mediated TFAM inhibition. J Cell Biochem.
119:680–690. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li W, Li H, Zhang L, Hu M, Li F, Deng J,
An M, Wu S, Ma R, Lu J and Zhou Y: Long non-coding RNA LINC00672
contributes to p53 protein-mediated gene suppression and promotes
endometrial cancer chemosensitivity. J Biol Chem. 292:5801–5813.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Akiel M, Guo C, Li X, Rajasekaran D,
Mendoza RG, Robertson CL, Jariwala N, Yuan F, Subler MA, Windle J,
et al: IGFBP7 deletion promotes hepatocellular carcinoma. Cancer
Res. 77:4014–4025. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jhuraney A, Woods NT, Wright G, Rix L,
Kinose F, Kroeger JL, Remily-Wood E, Cress WD, Koomen JM, Brantley
SG, et al: PAXIP1 potentiates the combination of WEE1 inhibitor
AZD1775 and platinum agents in lung cancer. Mol Cancer Ther.
15:1669–1681. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Beltran M, Puig I, Peña C, García JM,
Alvarez AB, Peña R, Bonilla F and de Herreros AG: A natural
antisense transcript regulates Zeb2/Sip1 gene expression during
snail1-induced epithelial-mesenchymal transition. Genes Dev.
22:756–769. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Suzuki K and Matsubara H: Recent advances
in p53 research and cancer treatment. J Biomed Biotechnol.
2011:9783122011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kahlert UD, Nikkhah G and Maciaczyk J:
Epithelial-to-mesenchymal(-like) transition as a relevant molecular
event in malignant gliomas. Cancer Lett. 331:131–138. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Pećina-Šlaus N, Kafka A, Varošanec AM,
Marković L, Krsnik Ž, Njirić N and Mrak G: Expression patterns of
Wnt signaling component, secreted frizzled-related protein 3 in
astrocytoma and glioblastoma. Mol Med Rep. 13:4245–4251. 2016.
View Article : Google Scholar : PubMed/NCBI
|