Introduction
Epilepsy is a common cerebral disease characterized
by interictal epileptiform discharge, which affects nearly 9
million people in China, among whom ~25% (>2.2 million) are
considered to have refractory epilepsy (1). Previous studies have demonstrated
that the development of temporal lobe epilepsy (TLE) is associated
with both abnormal structure and dysfunction of the brain, such as
hippocampal neuron regeneration, mossy fiber sprouting, synaptic
reconstruction and astrocyte proliferation (2–4).
Developing an effective treatment for TLE remains an important
research focus.
The concept of the ‘purine receptor’ has been used
to describe cell membrane adenosine receptors and adenosine
triphosphate (ATP) receptors (5).
ATP is an energy currency unit that is essential for cell function
and organization of biological signaling molecules (6). According to the International
Association of Pharmacology classification of receptors and drug
names, receptors that are sensitive to extracellular adenosine are
referred to as P1 receptors, whereas those that are sensitive to
extracellular nucleotides are referred to as P2 receptors. P2
purinergic receptors are subclassified into two types, i.e., P2X
and P2Y. P2X is an ATP ligand-gated ion channel. When extracellular
ATP binds to the P2X receptor, the P2X channel opens, allowing
cations to pass through the cell membrane (7–10).
The P2X7 receptor (P2X7R) is abundantly distributed
in the cerebrum (11), including
the solitary nucleus, hippocampus, habenular nucleus and substantia
nigra pars compacta (12). In
addition, it performs numerous physiological functions, such as
regulation of neurotransmitter release, stimulation of
proinflammatory cytokines and induction of cell damage and
apoptosis (13,14). However, whether P2X7R is involved
in the development of TLE remains unknown. Therefore, the aim of
the present study was to assess the expression of P2X7R in rat and
human brain tissue samples from epileptic and non-epileptic
subjects using immunohistochemical techniques, in order to
elucidate the role of P2X7R in epilepsy, particularly its possible
implications in the pathogenesis of TLE.
Materials and methods
Animals
A total of 90 male Sprague-Dawley rats (age, 2
months; weight, 200–250 g) were raised under controlled conditions
(temperature 24–25°C, humidity 50–60% and a 12-h light/dark cycle).
The animals had ad libitum access to food and water. All
procedures were implemented according to the guidelines of the
Animal Care Committee of Huazhong University of Science and
Technology and the study protocol was approved by the Ethics
Committee of Huazhong University of Science and Technology.
Drugs
The following drugs were used in the present study:
Pilocarpine hydrochloride (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany), lithium chloride (Sigma-Aldrich; Merck KGaA), atropine
sulfate (Shanghai Harvest Pharmaceutical Co., Ltd., Shanghai,
China), anti-P2RX7 antibody (Abcam, Cambridge, UK), anti-glial
fibrillary acidic protein (GFAP) antibody (Abcam), anti-glutamate
(GLU) antibody (Sigma-Aldrich; Merck KGaA) and diazepam (Tianjin
Jinyao Group Co., Ltd., Tianjin, China). Lithium chloride and
pilocarpine hydrochloride were dissolved in physiological saline
(0.9% sodium chloride).
Animal model of TLE
First, the rats were administered an intraperitoneal
(IP) injection of lithium chloride (127 mg/kg). After 18 h,
atropine sulfate (1 mg/kg, IP) was injected, followed by
pilocarpine hydrochloride (15 mg/kg, IP) after a further 30 min.
The seizure severity was graded according to Racine's standard
classification as follows: Stage 1, movement of the mouth and face;
stage 2, nodding of the head; stage 3, clonus of the unilateral
forelimb; stage 4, clonus and rearing of the bilateral forelimbs;
and stage 5, stage 4 seizure with episodes of falling. Pilocarpine
hydrochloride injection (15 mg/kg, IP) was repeated every 30 min if
there was no seizure, or if seizure activity did not reach stage 4
on the Racine scale. The maximum dose of pilocarpine hydrochloride
was 60 mg/kg. When status epilepticus persisted for 1 h, it was
terminated by an injection of diazepam (10 mg/kg, IP). The present
experiment only used rats graded at stage 4 or 5.
Behavioral observation
A total of 2 weeks after the onset of status
epilepticus, all rats underwent uninterrupted video monitoring
throughout the daytime to observe whether any spontaneous repeated
seizures occurred over 7 weeks. Behavioral parameters, including
latency period, frequency of spontaneous recurrent seizure,
duration of chronic epilepsy and mortality were analyzed during the
entire study period.
Electroencephalogram monitoring
Rats were anesthetized with 10% chloral hydrate (0.3
ml/100 g, IP) to undergo an electroencephalography (EEG). The rats
were fixed and two electrodes were inserted underneath the scalp on
both sides in the temporal region (0.65 cm in front of the
connection of the external ear gate and 0.4 cm beside the center
line). The reference electrode was inserted underneath the scalp at
the frontal pole midpoint (1.2 cm in front of the external ear
gate). EEG waveforms filtered <0.53 Hz and >30 Hz were
subjected to analog-to-digital conversion by a dynamic
electroencephalography device (Beijing Symtop Instrument Co., Ltd.,
Beijing, China). When the rats entered the spontaneous recurrent
seizure period, they underwent EEG monitoring for 4 h once per week
to record EEG waveforms and spike-wave discharges. Some rats with
spontaneous recurrent seizures were injected with brilliant blue G
(BBG) or saline for 2 weeks and EEG waveforms were recorded 30 min
before and 60 min after the injection.
Animal grouping and treatment
This study used a total of 90 male Sprague-Dawley
rats, among which 6 were given an IP injection of saline solution
to serve as the control group, whereas the remaining 84 were used
to establish models of epilepsy. A total of ~21 rats had no
epileptic seizures, among which 5 were randomly selected to
comprise the no seizure group. The remaining 63 rats were
eventually affected by status epilepticus, among which 5 were
randomly selected to comprise the acute seizure group. A total of
13 rats died after the onset of status epilepticus, 8 rats died
during the latency period, 9 rats were in the spontaneous recurrent
seizures group and 28 rats were in a chronic phase of spontaneous
recurrent seizures. These 28 rats were randomly divided into two
groups: The BBG group (21 rats) and the saline group (7 rats). The
BBG group was further randomly divided into 3 subgroups (7 rats
each) that were separately administered IP injections of BBG at
different doses (50, 100 or 200 mg/kg) for 2 weeks.
Patients with TLE
From June 2017 to June 2018, a total of 25 patients
with TLE (15 men and 10 women; age range, 18–55 years) from the
Wuhan Union Hospital in China were included in the present study.
All the patients had characteristic EEGs and representative
clinical features (mean number of seizures, 5.5/month), and were on
the highest dose of ≥3 antiepileptic drugs (AEDs), such as valproic
acid, oxcarbazepine, ethosuximide, lamotrigine, and clonazepam. All
patients underwent presurgical assessment including medical history
review, detailed neurological examination and ictal or normal EEG
studies; they also underwent magnetic resonance imaging (MRI),
video EEG, and sphenoidal electrode EEG prior surgery. All brain
MRI scans demonstrated no progressive disease in the central
nervous system. All patients underwent surgery to remove the
epileptogenic zone in the temporal neocortex. For comparison
purposes, from June 2017 to June 2018, 6 patients with head trauma
(3 men and 3 women; age range, 12–60 years) whose neocortices were
histologically normal and who had no history of epilepsy or use of
AEDs were examined. All procedures adhered to the conduct of
research involving human subjects established by the National
Institutes of Health of China and the Committee on Human Research
of Huazhong University of Science and Technology. All subjects
provided written informed consent to participate in this study.
Tissue processing
The rats were anesthetized with 10% chloral hydrate
(0.3 ml/100 g, IP) and then quickly perfused with 4%
paraformaldehyde in PBS. After embedding in paraffin, the brain
tissue was sliced into 4-µm sections. Similarly, 4-µm sections were
prepared from the 4% paraformaldehyde-fixed and paraffin-embedded
human brain tissue samples.
Immunohistochemical staining
First, rat hippocampal sections were incubated with
5% bovine serum albumin (Cell Signaling Technology, Inc., Danvers,
MA, USA) in PBS for 30 min at room temperature. Subsequently, the
sections were incubated with anti-P2X7 immunoglobulin G (cat. no.
ab48871; 1:100), anti-GLU IgG (cat. no. G9282; 1:100), or anti-GFAP
IgG (cat. no. ab33922; 1:100) overnight at 4°C. The sections were
washed in PBS for three times for 10 min each time and then
incubated with an Alexa® 594-conjugated secondary
antibody (cat. no. ab150160; 1:100; Abcam) for 40 min at 4°C. Then,
the sections were visualized with 3,3′-diaminobenzidine for 10 min
at room temperature in the dark, rinsed in distilled water and
dehydrated in a graded ethanol series. Negative control sections
were incubated with PBS instead of the primary antibody. Images
were captured via fluorescence microscopy at the same magnification
(×200) and grayscale was measured using Image-Pro Plus 6.0 software
(Media Cybernetics, Inc., Rockville, MD, USA).
Western blotting
The tissues were harvested, and total cell lysates
were prepared using a protein extraction kit (Wuhan Servicebio
Technology Co., Ltd., Wuhan, China), following the manufacturer's
protocol. Protein concentrations were quantified using a
bicinchoninic acid kit (Beyotime Institute of Biotechnology,
Haimen, China). Subsequently, 50 µg samples were boiled in
gel-loading buffer and separated by 10% SDS-PAGE. The proteins were
transferred to polyvinylidene difluoride membranes (Bio-Rad
Laboratories, Inc., Hercules, CA, USA) and blocked at 37°C with 5%
skimmed milk powder in Tris-buffered saline for 1 h. Then, the
blots were incubated for 6 h at 37°C with anti-P2RX7 (cat. no.
ab48871; 1:1,000; Abcam) and β-actin (cat. no. ab8226; 1:2,000;
Abcam) antibodies independently. The membranes were then incubated
for 1 h at 37°C with goat anti-rat horseradish
peroxidase-conjugated secondary antibody (cat. no. ab150160; 1:100;
Abcam). Chemiluminescent signals were detected using the Enhanced
Chemiluminescent Plus kit (KPL, Inc., Gaithersburg, MD, USA) and
the signal intensity was measured by densitometry analysis using
the Versa-Doc™ Imaging system (version 4.0; Bio-Rad
Laboratories, Inc.). β-actin was used as the internal control to
normalize the samples.
RNA extraction and polymerase chain
reaction (PCR)
Hippocampi from rats in each group were treated with
TRIzol® (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) reagent to extract total RNA following the
manufacturer's recommended protocol. Subsequently, genomic DNA was
digested using an RNase-free DNase. A total of 4–6 independent RNA
samples were used in duplicate in reverse
transcription-quantitative (RT-q) PCR analysis. NanoDrop 2000
(Thermo Fisher Scientific, Inc.) was used to detect RNA
concentration. Then, cDNA was transcribed from a total of 500 ng
DNase I-treated RNA using a cDNA reverse transcription kit (DP304;
Tiangen Biotech Co., Ltd., Beijing, China). Primers used in the
study were as follows, P2X7R primers: 5′-GGCGGGGTGACGAAGTTAGTA-3′
(forward) and 5′-TTTGTGGGTCCATCCATCCTT-3′ (reverse); β-actin
primers 5′-TGCTATGTTGCCCTAGACTTCG-3′ (forward) and
5′-GTTGGCATAGAGGTCTTTACGG-3′ (reverse). The temperature protocol
was as follows: 37°C 15 min and 95°C 5 min. qPCR analysis was
performed on an ABI Prism 7500 sequence detection system (Applied
Biosystems; Thermo Fisher Scientific, Inc.) using Thunderbird SYBR
qPCR mix (Toyobo Life Science, Osaka, Japan) in 20 µl of reaction
mixture. The thermocycling conditions were as follows: Initial
denaturation at 95°C for 5 min, followed by 33 cycles of 95°C (15
sec) and 60°C (1 min). Data were analyzed using Sequence Detection
Software 1.4 (Applied Biosystems; Thermo Fisher Scientific, Inc.).
β-actin was selected as the endogenous reference and data were
analyzed using the 2−ΔΔCq method (15).
Statistical analysis
All values were presented as the mean ± standard
deviation of three separate experiments and were analyzed by the
one-way analysis of variance, which was used to compare multiple
groups, followed by a Bonferroni post-hoc test. An unpaired
two-tailed Student's t-test was employed to compare the levels of
P2X7 receptor and GFAP and GLU in patients with or without
intractable epilepsy. Statistical analysis was performed using SPSS
software, version 19.0 (IBM Corp., Armonk, NY, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
Behavioral analysis
Rats injected with pilocarpine hydrochloride
exhibited several behavioral events (data not shown), including
stereotypical masticatory movements, hypokinesia, head nodding and
wet-dog shakes. This behavior rapidly progressed to generalized
limbic seizures recurring every 1–3 min and finally culminating in
status epilepticus. A total of 4 rats (4.4%) died when injected
with the drug and 63 (73%) eventually developed status epilepticus.
Behavioral observations via video surveillance revealed that 28
rats (56%) were in a chronic phase of spontaneous recurrent
seizures. Interestingly, one-third reduction in the frequency of
spontaneous recurrent seizures was found in the BBG-treated
group.
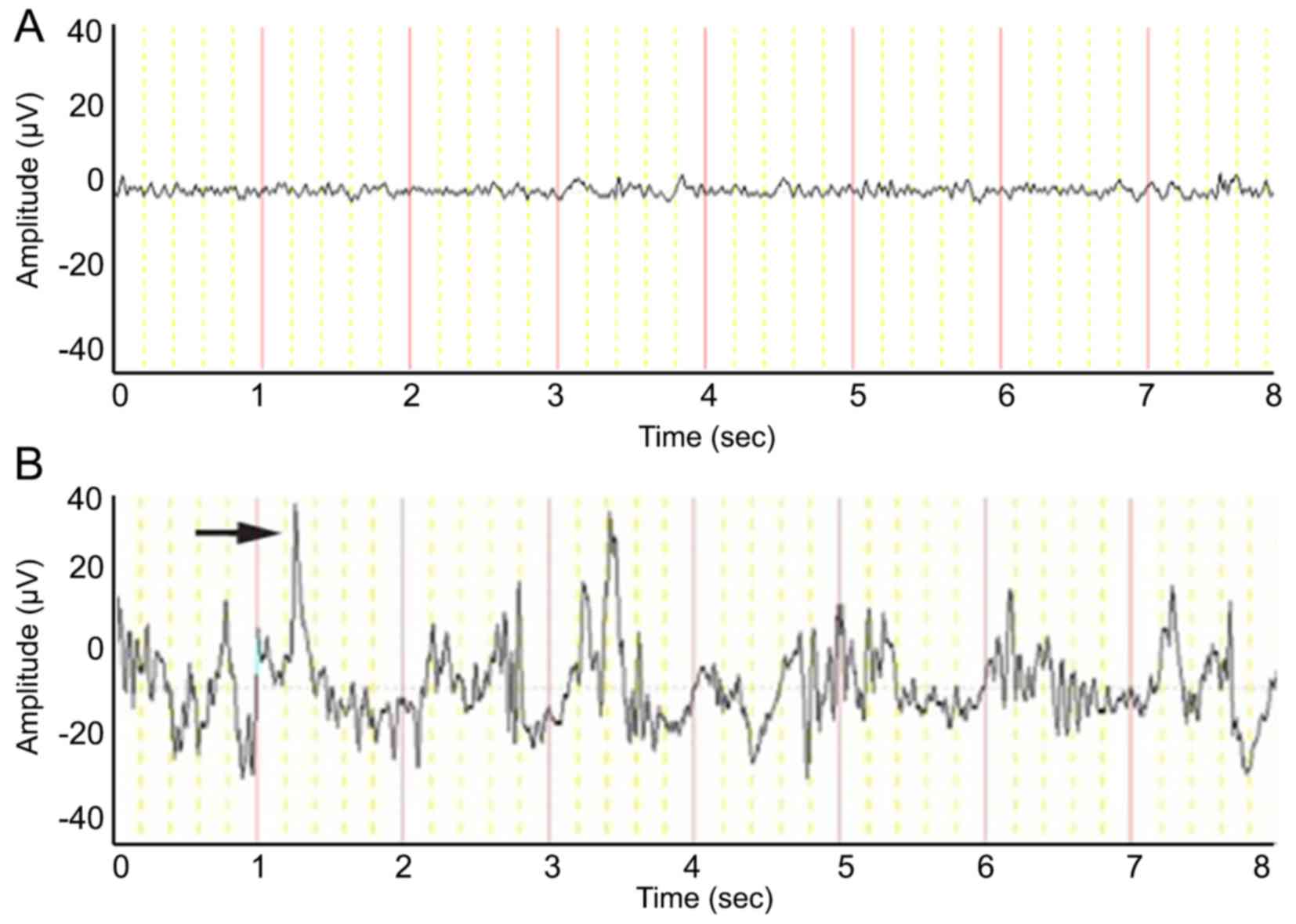
EEG recordings
In the control group, EEG recorded waves of 8–10 Hz
and 20–120 µV voltages, followed by low β waves and a single δ
wave. In sharp contrast, the 28 rats with status epilepticus
exhibited great spike-wave discharges (maximum of 28 Hz and 200
µV). Some spike-wave discharges presented individually, whereas
others appeared in a short string (Fig. 1).
P2X7R expression in the rat model
Immunohistochemistry
P2X7R immunoreactivity was found to be high in the
cortex, hippocampus, thalamus and amygdala of rats with spontaneous
recurrent seizures, but exhibited a significant reduction in the
BBG-treated groups 9 [50 mg/kg (V), 100 mg/kg (VI) and 200 mg/kg
(VII); Fig. 2], demonstrating a
negative correlation with BBG dose. Astroglial activation occurred
in the hippocampus of rats with spontaneous recurrent seizures, but
this phenomenon was prevented by BBG. Concomitantly, GLU expression
was upregulated in rats with spontaneous recurrent seizures
compared with the control group, but was also reduced in the
BBG-treated groups [50 mg/kg (V), 100 mg/kg (VI) and 200 mg/kg
(VII); Fig. 2]. However, there was
no significant difference in P2X7R expression between the control
group and rats with status epilepticus (I and III, respectively;
Fig. 2).
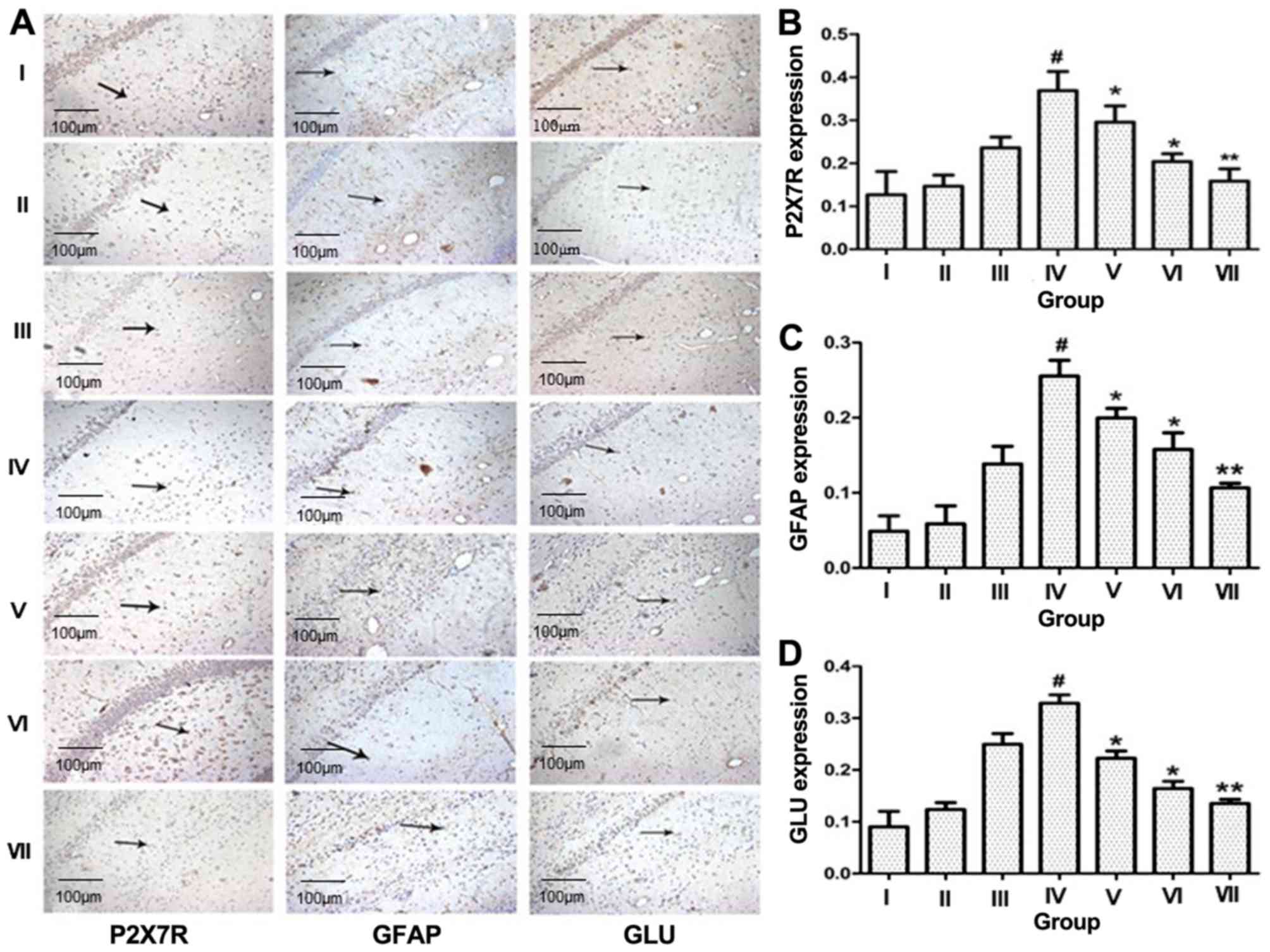 | Figure 2.Increased presence of P2X7R, GFAP and
GLU in the hippocampal CA3 region of temporal lobe epilepsy rats.
(A) Immunohistochemical staining for P2X7R, GFAP and GLU in the CA3
region of the rat hippocampus (×200). Intensity of staining for (B)
P2X7R, (C) GFAP and (D) GLU in the CA3 region of the rat
hippocampus. One-way analysis of variance was conducted to compare
differences among groups. I, control group; II, no seizure group;
III, status epilepticus group; IV, spontaneous recurrent seizures
group; V, BBG-treated group (50 mg/kg); VI, BBG-treated group (100
mg/kg); VII, BBG-treated group (200 mg/kg). #P<0.01
vs. I; *P<0.05 and **P<0.01 vs. IV. GLU, glutamate; GFAP,
glial fibrillary acidic protein; P2X7R, P2X7 receptor; BBG,
brilliant blue G. |
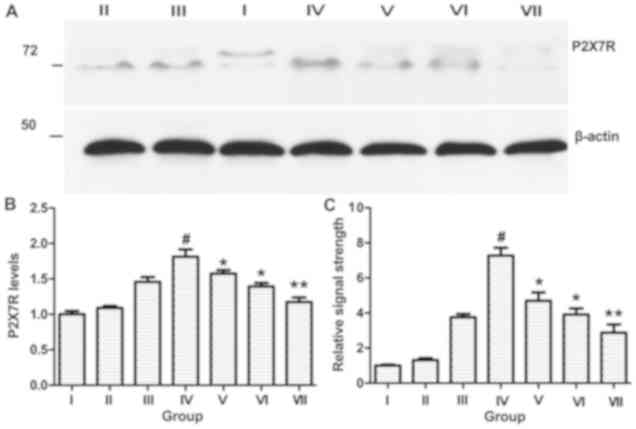
Western blotting and RT-qPCR
Western blotting and RT-qPCR analyses demonstrated
that P2X7R expression was increased after spontaneous recurrent
seizures within the ipsilateral CA3 subfield of the hippocampus.
However, there was no difference between the control group and rats
with status epilepticus (Fig. 3).
In contrast to rats with spontaneous recurrent seizures, P2X7R
expression was significantly reduced in the BBG-treated groups
(P<0.05).
P2X7R expression in patients with
TLE
Compared with the control group, immunostaining for
P2X7R, GFAP and GLU revealed significant upregulation in the
temporal lobe of patients with TLE (P<0.05; Fig. 4).
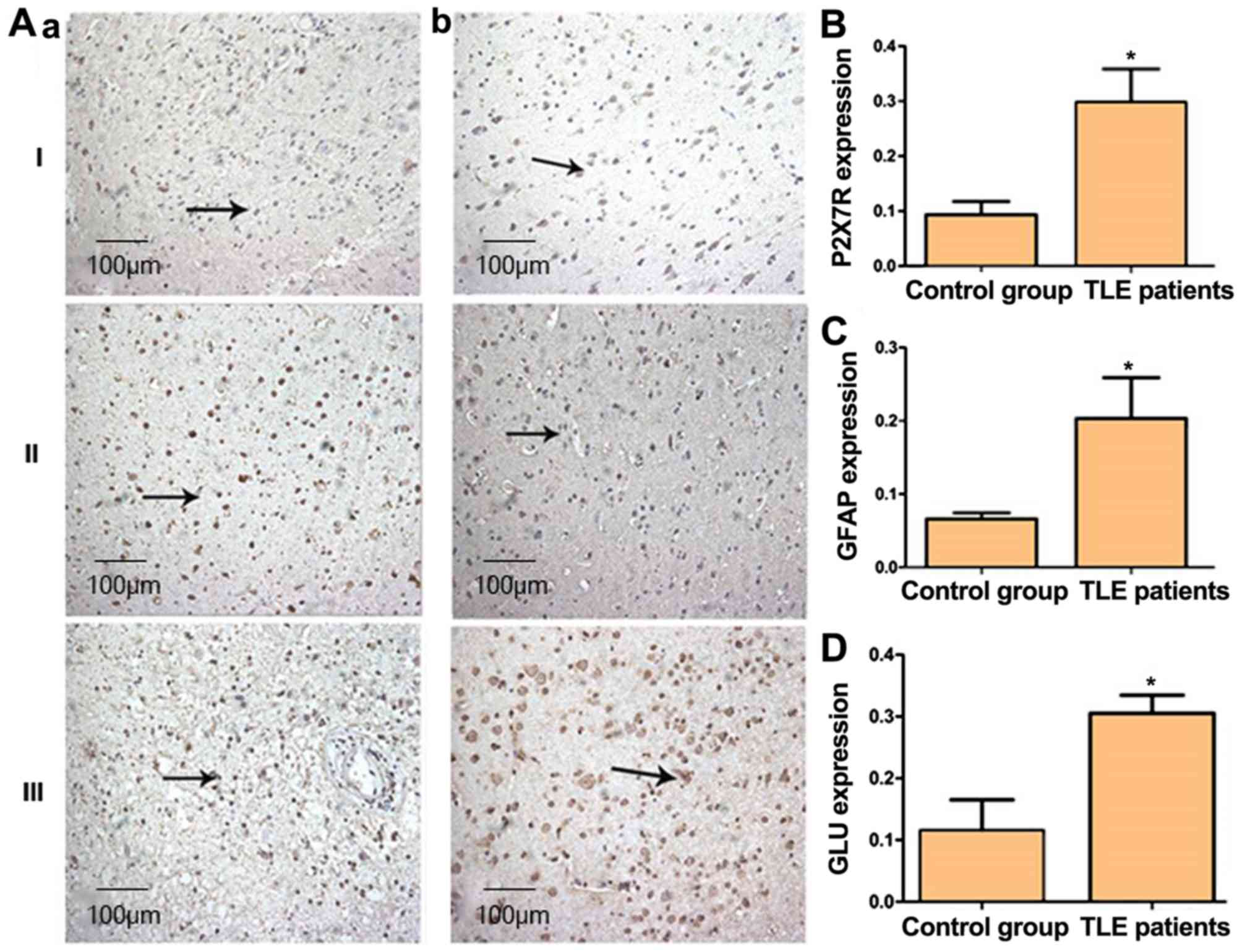 | Figure 4.Increased presence of P2X7R, GFAP and
GLU in the temporal lobe of TLE patients. (A) Immunohistochemical
staining for P2X7R, GFAP and GLU in human brain tissue
(magnification, ×200). Positive staining is indicated by arrows.
(aI-aIII) are control cases and (bI-bIII) are patients with TLE.
Intensity of staining for (B) P2X7R, (C) GFAP and (D) GLU in human
brain tissue. I, P2X7R; II, GFAP; III, GLU. *P<0.05. GLU,
glutamate; GFAP, glial fibrillary acidic protein; P2X7R, P2X7
receptor; TLE, temporal lobe epilepsy. |
Discussion
To the best of our knowledge, the present study was
the first to demonstrate that P2X7R expression is elevated in
patients with intractable TLE. This investigation confirmed changes
in ATP-gated P2X7R expression in the brain tissue of experimental
rats and humans suffering from refractory epilepsy. Furthermore,
the expression of GFAP, an astrocytic protein and GLU, the main
excitatory neurotransmitter was measured. The results of the
present study demonstrated that the levels of these two proteins
were increased in the hippocampus of rats in a chronic phase of
spontaneous recurrent seizures as well as in the temporal lobe of
patients with TLE. Of note, inhibition of P2X7R using BBG, a P2X7R
antagonist, effectively reduced the levels of the two proteins in
the rat hippocampus. These results suggest that P2X7R may
contribute to the pathogenic mechanism of spontaneous seizures and
may represent a novel drug target for seizure prevention.
In humans, epilepsy is a neurological emergency
associated with various mechanisms, such as proliferation and
repeated activation of astrocytes (16), cerebral cortical developmental
disorders (17), immunological
dysfunction (18) and disturbance
of the glutamatergic system (19).
A previous study revealed that TLE is a common type of refractory
epilepsy that does not appear to respond to initiation of
anticonvulsant therapy (20). In
the present study, a pilocarpine-induced seizure model was
selected, which highly resembles the morphological and synaptic
characteristics of TLE in humans. Rats injected with pilocarpine
endure a period of status epilepticus followed by a chronic
indefinite period of spontaneous recurrent seizures (21).
P2X7R, a purine receptor, is located in multiple
parts of the central nervous system, including the hippocampus,
cerebellum, thalamus, striatum and nerve terminals, and performs
numerous physiological functions, such as adjustment of
neurotransmitter release, stimulation of cytokines and chemotactic
factors, as well as induction of cell injury and apoptosis. BBG, a
new type of biological stain that can cross the blood-brain
barrier, has been demonstrated to block P2X7R with no obvious
biological toxicity and has been widely applied in recent
P2X7R-related studies (22,23).
In addition to the discovery of the P2X7R expression trend in the
seizure samples, another main finding was that BBG reduced the
levels of both GFAP and GLU in the CA3 region of rat hippocampus in
a dose-dependent manner. In addition, BBG-treated rats experienced
milder seizures compared with rats that were not treated. The
expression of P2X7R was also examined in patients with TLE.
Immunohistochemical examination demonstrated that P2X7R expression
was increased in patients with TLE compared with in the control
group, which was consistent with the results of the present study's
animal research, indicating that P2X7R plays a key role in the
pathogenesis of intractable epilepsy. Due to ethical restrictions,
normal brain tissue samples could not be obtained; therefore,
tissue samples from patients with head trauma were used. In
addition, although brain tissue samples were only obtained from
patients with refractory epilepsy, the results of the present study
may provide some insight into epilepsy in general.
The analysis of GFAP and GLU expression suggested
that the possible mechanism through which P2X7R contributes to
intractable epilepsy may be associated with preventing enhancement
of GLU released by astrocytes. Astrocytes play a central role in
maintaining homeostasis of the neuronal microenvironment by means
of transporting transmitters (24,25).
Studies have found that the process of epilepsy is often
accompanied by obvious increases in astrocyte number, as well as
changes in morphological and biochemical indicators (26,27).
In TLE patients, there was a further decline in the number of
principal cells in all hippocampal sub-areas analyzed, which was
associated with an increase in GFAP immunoreactivity (28,29).
As a vital excitatory neurotransmitter, GLU has been a research
focus with regard to central nervous system diseases, such as
epilepsy (30). Excessive
accumulation of extracellular GLU may lead to excitotoxicity of the
neurons through different N-methyl-D-aspartate receptors and
eventually trigger a cascade of intracellular signals that result
in neuronal death (31). Earlier
evidence has revealed that modulation of GLU by astrocytes plays an
important role in the homeostasis of extracellular GLU (32,33),
thus participating in the molecular pathogenesis of epilepsy
(34,35). The present study demonstrated that
P2X7R antagonists may be a potential putative therapy independently
or in combination with GLU receptor antagonists.
Further research is required to determine whether
BBG affects P2X7R and how this signaling mediates astrogliosis.
Moreover, further studies are needed to investigate the effects of
P2X7R modulation on TLE.
In conclusion, increased levels of P2X7R were
observed in patients and experimental animals with intractable TLE.
Therefore, P2X7R may be involved in the pathogenesis of TLE and its
suppression may provide a new treatment to promote remission in
epilepsy.
Acknowledgements
Not applicable.
Funding
The present study was funded by the Natural Science
Foundation of Hubei Province in China, (grant no.
02.02.040458).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
XD designed the study and drafted the manuscript; PS
performed the animal experiments, and was a major contributor in
writing the manuscript. JH and XL performed the western blot and
PCR assay. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All procedures adhered to the conduct of research
involving human subjects established by the National Institutes of
Health of China and the Committee on Human Research of Huazhong
University of Science and Technology. The present study was
approved by the Ethics Committee of Huazhong University of Science
and Technology. All subjects provided written informed consent to
participate in this study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Fisher RS, van Emde Boas W, Blume W, Elger
C, Genton P, Lee P and Engel J Jr: Epileptic seizures and epilepsy:
Definitions proposed by the International League Against Epilepsy
(ILAE) and the International Bureau for Epilepsy (IBE). Epilepsia.
46:470–472. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Guangming Z, Wenjing Z, Jiuluan L, Zhaohui
S, Bingqing Z, Gaoxiang S and Huancong Z: Long-term therapeutic
effects of corticoamygdalohippocampectomy for bilateral mesial
temporal lobe epilepsy. Surg Neurol Int. 4:1472013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kandratavicius L, Ruggiero RN, Hallak JE,
Garcia-Cairasco N and Leite JP: Pathophysiology of mood disorders
in temporal lobe epilepsy. Rev Bras Psiquiatr. 34 (Suppl
2):S233–S245. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Braakman HM, Vaessen MJ, Jansen JF,
Debeij-van Hall MH, de Louw A, Hofman PA, Vles JS, Aldenkamp AP and
Backes WH: Frontal lobe connectivity and cognitive impairment in
pediatric frontal lobe epilepsy. Epilepsia. 54:446–454. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Harden TK, Boyer JL and Nicholas RA:
P2-purinergic receptors: Subtype-associated signaling responses and
structure. Annu Rev Pharmacol Toxicol. 35:541–579. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang F, Pracheil T, Thornton J and Liu Z:
Adenosine triphosphate (ATP) is a candidate signaling molecule in
the mitochondria-to-nucleus retrograde response pathway. Genes
(Basel). 4:86–100. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dal Ben D, Buccioni M, Lambertucci C,
Marucci G, Thomas A and Volpini R: Purinergic P2X receptors:
Structural models and analysis of ligand-target interaction. Eur J
Med Chem. 89:561–580. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Booth JW, Tam FW and Unwin RJ: P2
purinoceptors: Renal pathophysiology and therapeutic potential.
Clin Nephrol. 78:154–163. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Baines A, Parkinson K, Sim JA, Bragg L,
Thompson CR and North RA: Functional properties of five
Dictyostelium discoideum P2X receptors. J Biol Chem.
288:20992–21000. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hopfner M, Lemmer K, Jansen A, Hanski C,
Riecken EO, Gavish M, Mann B, Buhr H, Glassmeier G and Scherübl H:
Expression of functional P2-purinergic receptors in primary
cultures of human colorectal carcinoma cells. Biochem Biophys Res
Commun. 251:811–817. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Grygorowicz T, Sulejczak D and Struzynska
L: Expression of purinergic P2X7 receptor in rat brain during the
symptomatic phase of experimental autoimmune encephalomyelitis and
after recovery of neurological deficits. Acta Neurobiol Exp (Wars).
71:65–73. 2011.PubMed/NCBI
|
|
12
|
Norenberg W, Schunk J, Fischer W, Sobottka
H, Riedel T, Oliveira JF, Franke H and Illes P:
Electrophysiological classification of P2X7 receptors in rat
cultured neocortical astroglia. Br J Pharmacol. 160:1941–1952.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Eleftheriadis T, Pissas G, Karioti A,
Antoniadi G, Golfinopoulos S, Liakopoulos V, Mamara A, Speletas M,
Koukoulis G and Stefanidis I: Uric acid induces caspase-1
activation, IL-1β secretion and P2X7 receptor dependent
proliferation in primary human lymphocytes. Hippokratia.
17:141–145. 2013.PubMed/NCBI
|
|
14
|
Skaper SD, Debetto P and Giusti P: The
P2X7 purinergic receptor: From physiology to neurological
disorders. FASEB J. 24:337–345. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Héja L: Astrocytic target mechanisms in
epilepsy. Curr Med Chem. 21:755–763. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kuchukhidze G, Koppelstaetter F,
Unterberger I, Dobesberger J, Walser G, Höfler J, Zamarian L,
Haberlandt E, Rostasy K, Ortler M, et al: Midbrain-hindbrain
malformations in patients with malformations of cortical
development and epilepsy: A series of 220 patients. Epilepsy Res.
106:181–190. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lorigados-Pedre L, Morales-Chacon L,
Pavón-Fuentes N, Serrano-Sánchez T, Robinson-Agramonte MA,
García-Navarro ME and Bender-del Busto JE: Immunological disorders
in epileptic patients are associated to the epileptogenic focus
localization. Rev Neurol. 39:101–104. 2004.(In Spanish). PubMed/NCBI
|
|
19
|
Peng WF, Ding J, Mao LY, Li X, Liang L,
Chen CZ, Cheng WZ, Fan W and Wang X: Increased ratio of
glutamate/glutamine to creatine in the right hippocampus
contributes to depressive symptoms in patients with epilepsy.
Epilepsy Behav. 29:144–149. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Menon R, Radhakrishnan A and Radhakrishnan
K: Status epilepticus. J Assoc Physicians India 61 (8 Suppl).
S58–S63. 2013.
|
|
21
|
Grabenstatter HL, Del AY, Carlsen J, Wempe
MF, White AM, Cogswell M, Russek SJ and Brooks-Kayal AR: The effect
of STAT3 inhibition on status epilepticus and subsequent
spontaneous seizures in the pilocarpine model of acquired epilepsy.
Neurobiol Dis. 62:73–85. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sakamoto K, Inukai M, Mori A and Nakahara
T: Brilliant Blue G protects against photoreceptor injury in a
murine endotoxin-induced uveitis model. Exp Eye Res. 177:45–49.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bartlett R, Sluyter V, Watson D, Sluyter R
and Yerbury JJ: P2X7 antagonism using Brilliant Blue G reduces body
weight loss and prolongs survival in female SOD1G93A
amyotrophic lateral sclerosis mice. PeerJ. 5:e30642017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hussmann KL, Samuel MA, Kim KS, Diamond MS
and Fredericksen BL: Differential replication of pathogenic and
nonpathogenic strains of West Nile virus within astrocytes. J
Virol. 87:2814–2822. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Benediktsson AM, Marrs GS, Tu JC, Worley
PF, Rothstein JD, Bergles DE and Dailey ME: Neuronal activity
regulates glutamate transporter dynamics in developing astrocytes.
Glia. 60:175–188. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kozela E, Juknat A and Vogel Z: Modulation
of astrocyte activity by cannabidiol, a nonpsychoactive
cannabinoid. Int J Mol Sci. 18(pii): E16692017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shen HY, Sun H, Hanthorn MM, Zhi Z, Lan
JQ, Poulsen DJ, Wang RK and Boison D: Overexpression of adenosine
kinase in cortical astrocytes and focal neocortical epilepsy in
mice. J Neurosurg. 120:628–638. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Peixoto-Santos JE, Velasco TR,
Galvis-Alonso OY, Araujo D, Kandratavicius L, Assirati JA, Carlotti
CG, Scandiuzzi RC, Santos AC and Leite JP: Temporal lobe epilepsy
patients with severe hippocampal neuron loss but normal hippocampal
volume: Extracellular matrix molecules are important for the
maintenance of hippocampal volume. Epilepsia. 56:1562–1570. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Proper EA, Oestreicher AB, Jansen GH,
Veelen CW, van Rijen PC, Gispen WH and de Graan PN:
Immunohistochemical characterization of mossy fibre sprouting in
the hippocampus of patients with pharmaco-resistant temporal lobe
epilepsy. Brain 123 (Pt 1). 19–30. 2000.
|
|
30
|
Swamy AH, Patel NL, Gadad PC, Koti BC,
Patel UM, Thippeswamy AH and Manjula DV: Neuroprotective activity
of pongamia pinnata in monosodium glutamate-induced neurotoxicity
in rats. Indian J Pharm Sci. 75:657–663. 2013.PubMed/NCBI
|
|
31
|
Zhu S and Paoletti P: Allosteric
modulators of NMDA receptors: Multiple sites and mechanisms. Curr
Opin Pharmacol. 20:14–23. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jiang S, Wang YQ, Xu CF, Li YN, Guo R and
Li L: Involvement of connexin43 in the infrasonic noise-induced
glutamate release by cultured astrocytes. Neurochem Res.
39:833–842. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Morales I and Rodriguez M: Self-induced
accumulation of glutamate in striatal astrocytes and basal ganglia
excitotoxicity. Glia. 60:1481–1494. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sheean RK, Lau CL, Shin YS, O'Shea RD and
Beart PM: Links between L-glutamate transporters, Na+/K+-ATPase and
cytoskeleton in astrocytes: Evidence following inhibition with
rottlerin. Neuroscience. 254:335–346. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lee ES, Sidoryk M, Jiang H, Yin Z and
Aschner M: Estrogen and tamoxifen reverse manganese-induced
glutamate transporter impairment in astrocytes. J Neurochem.
110:530–544. 2009. View Article : Google Scholar : PubMed/NCBI
|