Introduction
Acute liver failure (ALF) is a serious disorder or
decompensation caused by various factors, including liver
synthesis, detoxification, excretion and biotransformation, and the
mortality rate of ALF caused by hepatitis B virus (HBV) is more
than 50% in China (1).
Enterogenous endotoxemia plays an important role in the development
and prognosis of ALF (2). When the
intestinal flora is disturbed during ALF, intestinal mucosal damage
and intestinal bacterial translocation occur. Gram-negative
bacteria or their products can be transferred from the intestinal
lumen to the circulatory system, causing an inflammatory response
in the liver and inducing the necrosis of hepatocytes (3).
As a novel type of necrosis, ferroptosis was
described by Dixon et al (4) in 2012. Ferroptosis is characterized
by intracellular iron ion accumulation, elevated lipid
peroxidation, mitochondria and mitochondrial membrane density.
Oxidative stress is the key pathogenic link with ALF and could
promote ALF (5). Ferroptosis is
also characterized by the accumulation of reactive oxygen species
(ROS) under high oxidative stress. Therefore, if ferroptosis can be
inhibited, ALF may be effectively alleviated.
Glycyrrhizin (GLY) is the main extract from the
glycyrrhiza root and is an important compound (6). Its molecular formula is
C42H62O16 (7). It has been reported that GLY has
anti-oxidative, anti-inflammatory, anti-virus activities, as well
as anti-fibrotic activity in the liver, and that GLY can inhibit
tumor growth and enhance immunity (8–11).
GLY is a natural antioxidant that has a protective effect on the
liver and is widely used in the treatment of chronic hepatitis
(12).
However, whether GLY can alleviate ferroptosis in
ALF requires further study. In the present study, tumor necrosis
factor-α (TNF-α), lipopolysaccharide (LPS) and D-galactosamine
(D-GalN) were used to stimulate L02 hepatocytes and mice in order
to construct a cellular and mouse model of ALF. GLY was
administered to these ALF models to evaluate the degree of
ferroptosis, and the corresponding pathway changes, in
hepatocytes.
Materials and methods
Reagents
The normal human liver cell line (L02) was purchased
from The Cell Collection Center of Wuhan University. The Cell
Counting Kit-8 (CCK-8) assay was purchased from Dojindo Chemical
Technology Co., Ltd. GLY was purchased from Selleck Chemicals. The
lactate dehydrogenase (LDH), malondialdehyde (MDA), glutathione
(GSH) and ROS kits were purchased from Beyotime Institute of
Biotechnology. The primary antibodies against glutathione
peroxidase 4 (GPX4, cat. no. sc-166570) was purchased from Santa
Cruz Biotechnology, Inc. GAPDH (cat. no. 10494-1-AP) were purchased
from ProteinTech Group, Inc. The primary antibodies against high
mobility group box 1 (HMGB1, cat. no. 3935), nuclear factor
E2-related factor 2 (Nrf2, cat. no. 12721) and homooxygenase-1
(HO-1, cat. no. 43966) were purchased from Cell Signaling
Technology, Inc. The goat anti-rabbit IRDye fluorescent secondary
antibody IRDye800CW (cat. no. 926-32211) was purchased from LI-COR
Biosciences. Cy3 (cat. no. BA1031 and BA1032) and FITC (cat. no.
BA1101 and BA1105) fluorescently labeled rabbit anti-goat secondary
antibodies were purchased from Wuhan Boster Biological Technology,
Ltd. The iron ion detection kit was purchased from Abcam. D-GalN,
LPS and TNF-α were purchased from Sigma-Aldrich; Merck KGaA.
Cell culture and drug
intervention
L02 cells were cultured in DMEM (Gibco; Thermo
Fisher Scientific, Inc.) containing 10% fetal bovine Serum (FBS,
Gibco; Thermo Fisher Scientific, Inc.) The cells were cultured at
37°C in an incubator with 5% CO2. When cells reached a
confluency of 70–80%, cells were digested with trypsin and seeded
into 6-(1.5×106 cells/1.5 ml DMEM per plate) or 96-well
(5×103 cells/100 µl DMEM per plate) plates. The cells
were divided into five groups as follows: The normal group, model
group, GLY 0.5 mM intervention group, GLY 1 mM intervention group
and GLY 2 mM intervention group. The GLY intervention groups were
stimulated with the corresponding concentration of GLY for 2 h at
37°C in advance of further treatment. After 2 h, TNF-α (100 ng/ml)
and D-GalN (44 µg/ml) was added to the cells in the model group and
GLY intervention groups for 24 h. The cells and supernatant were
collected for subsequent experimental testing.
Animal model preparation
In total, 40 male specific- pathogen-free C57BL/6
mice (Hubei Animal Experimental Center) 6–8 weeks old and weighing
20–25 g were housed in the animal experiment center in Renmin
Hospital of Wuhan University with 12 h light/dark cycle,
temperature of 25±2°C and relative humidity of 50±15%. All mice
were provided with food and water ad libitum. The mice were
randomly divided into 5 groups: The normal group, model group, 15
mg/kg GLY group, 30 mg/kg GLY group and 60 mg/kg GLY group. Except
for the normal group, the other four groups of mice were injected
intraperitoneally with D-GalN (400 mg/kg) and LPS (100 µg/kg) to
induce the ALF model. According to a previous study on GLY gavage
doses (13), three doses of GLY
(15, 30 and 60 mg/kg/day) intervention groups were used. A total of
24 mice were divided into three groups. Mice received gavage with
different doses of GLY for 3 days before induction of the ALF
model. The ALF model group received the same amount of isotonic
saline. After 24 h, the liver tissues were removed. The serum was
collected to determine the levels of alanine aminotransferase
(ALT), aspartate aminotransferase (AST) and total bilirubin (TBIL)
by using a fully automatic biochemical analyzer (ADVIA 2400,
Siemens AG). The present study was approved by the Institutional
Animal Care and Use Committee of Renmin Hospital of Wuhan
University.
CCK-8 detection of cell viability
L02 cells were uniformly seeded in a 96-well plate
at a density of 5×103 cells/100 µl in DMEM with
different concentrations of GLY (0, 0.5, 1 and 2 mM). According to
the manufacturer's protocol, 10 µl CCK-8 reagent was added to each
well after 24 h. After mixing, the optical density (OD) for each
well was measured using a microplate reader. Cell viability was
calculated according to the following formula: Cell viability=(OD
value of experimental group-OD value of blank control group)/(OD
value of normal group-OD value of blank control) ×100.
LDH, MDA, Fe2+, ROS and GSH
levels in the L02 cell line and liver tissues
The LDH, MDA, Fe2+, ROS and GSH kits were
performed according to the manufacturers' protocols. The values for
the levels of LDH, MDA, Fe2+, ROS and GSH in the cells,
cell supernatant and liver tissues were measured using a microplate
reader at the absorption wavelengths of 490, 532, 593, 490 and 412
nm.
Western blotting and
immunofluorescence detection of protein expression
The cells or liver tissues were homogenized by radio
immunoprecipitation assay (RIPA) lysis buffer (Beyotime Institute
of Biotechnology) on ice to extract total protein. Protein
concentration was determined by bicinchoninic acid (BCA) protein
assay reagent assay kit (Beyotime Institute of Biotechnology).
Protein lysates (30 µg) were subjected to 12% SDS-PAGE. The
proteins were transferred to PVDF membranes (EMD Millipore). After
blocking with 20% non-fat milk at room temperature for 1 h, the
membranes were incubated with primary antibodies against GPX4
(1:1,000), Nrf2 (1:1,000), HO-1 (1:1,000), HMGB1 (1:1,000) and
GAPDH (1:1,000) overnight at 4°C. The IRDye800CW secondary antibody
(1:10,000) was incubated at 37°C for 1 h. The protein bands were
visualized using the Odyssey Infrared Imaging System (version 3.0,
LI-COR Biosciences).
For immunofluorescence, circular slides were placed
in 24-well plates and were seeded with 1×104 L02
cells/400 µl DMEM per plate. After treatments, the slides were
fixed with 4% paraformaldehyde at 37°C for 30 min, permeabilized
with 0.2% Triton X-100 (Beyotime Institute of Biotechnology) at
37°C for 20 min, blocked with 5% bovine serum albumin (BSA, Beijing
Solarbio Science & Technology Co., Ltd.) at 37°C for 30 min and
incubated with primary antibodies against HMGB1 (1:100) and GPX4
(1:100) at 4°C overnight. Slides were then incubated with Cy3 and
FITC-labeled fluorescent secondary antibodies (1:100) at 37°C for 1
h in dark room. Then the sections were stained with 5 µg/ml DAPI
(Beyotime Institute of Biotechnology) at 37°C for 5 min in a dark
room. The slides were observed under a fluorescence microscope at
×200 magnification.
Hematoxylin and eosin (H&E)
staining and immunofluorescence detection of liver tissue
Mouse liver tissues were fixed in 4%
paraformaldehyde solution at 37°C for 24 h and embedded in
paraffin, then sectioned at 5 µm thickness. The sections were
stained with Harris hematoxylin (Beyotime Institute of
Biotechnology) at 37°C for 5 min and eosin (95% ethanol
preparation, Beyotime Institute of Biotechnology) at 37°C for 15
sec. After staining, the liver tissues were observed under a light
microscope at ×200 magnification. For the immunofluorescence
detection of liver tissues, paraffin sections were dewaxed and
hydrated, and immersed in citrate buffer for antigen retrieval.
Sections were blocked with 5% BSA at 37°C for 30 min, and incubated
with GPX4 (1:200) and HMGB1 (1:200) antibodies at 4°C overnight.
Slices were then incubated with the Cy3 and FITC-labeled secondary
antibody at 37°C for 1 h in a dark room. DAPI (5 µg/ml) was used to
stain nuclei at 37°C for 5 min in dark room. Finally, the slices
were imaged under an inverted fluorescence microscope at ×200
magnification.
Statistical analysis
All data are presented as the mean ± SEM. Data were
analyzed using SPSS 17.0 statistical software (SPSS, Inc.). All
experiments were repeated three times. The differences among three
or more groups were analyzed using one-way ANOVA followed by
Bonferroni's post hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
GLY enhances cell viability in
TNF-α/D-GalN-induced L02 cells
The destruction of the cell membrane structure
induced by ferroptosis causes the release of LDH in the cytoplasm
into the culture medium. Therefore, the degree of liver damage can
be determined by detecting the levels of LDH in the supernatant of
cultured cells. As shown in Fig. 1A
and B, when compared with the normal group, the cell viability
in the L02 model group induced by TNF-α/D-GalN was significantly
reduced, while the LDH level was significantly increased
(P<0.01). Treatment with 1 or 2 mM GLY increased cell viability
and decreased the level of LDH compared with the model group
(P<0.01).
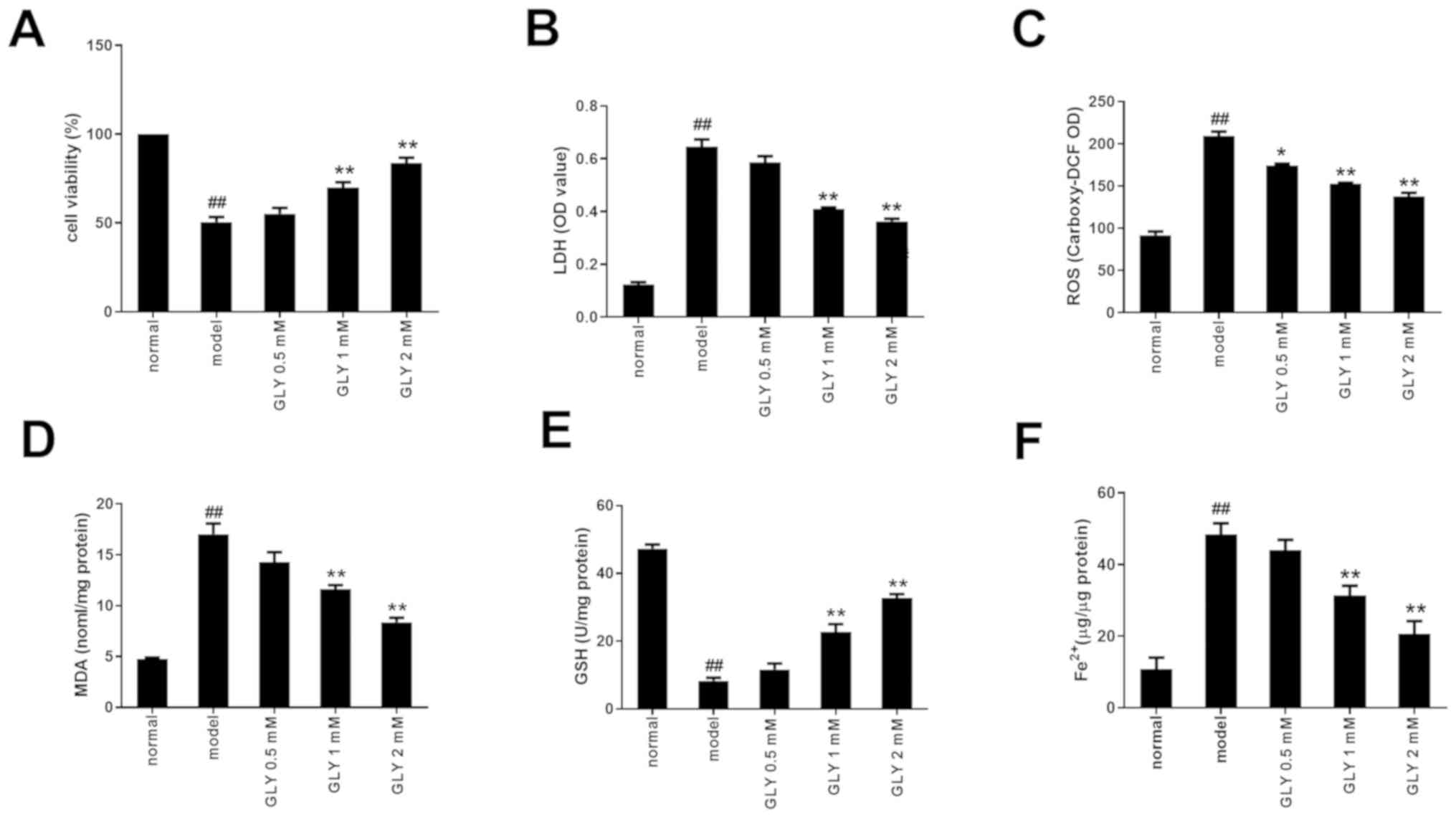 | Figure 1.GLY decreased cell injury in
TNF-α/D-GalN induced L02 cells. (A) GLY increased cell viability in
TNF-α/D-GalN-induced L02 cells. GLY decreased the levels of (B)
LDH, (C) ROS and (D) MDA, and (E) increased the level of GSH in
TNF-α/D-GalN-induced L02 cells. (F) GLY decreased the level of
Fe2+ in TNF-α/D-GalN induced L02 cells. n=3.
##P<0.01 vs. normal; **P<0.01, *P<0.05 vs.
model. TNF-α, tumour necrosis factor-α; D-GalN, D-galactosamine;
GLY, glycyrrhizin; LDH, lactate dehydrogenase; ROS, reactive oxygen
species; MDA, malondialdehyde; GSH, glutathione; OD, optical
density; carboxy-DCF,
6-carboxy-2′,7′-dichlorodihydrofluorescein. |
GLY enhances the anti-oxidative
capacity in TNF-α/D- GalN-induced L02 cells
ROS is the general term for a class of highly
reactive compounds that are composed of oxygen, including oxygen
free radicals and non-radical oxygenated products (14). ROS accumulate following adverse
external stimuli and can exceed the scavenging ability of the
peroxidase system, leading to oxidative stress, triggering lipid
peroxidation, damage to the cell membrane and finally inducing
ferroptosis (15). MDA is a
product of lipid peroxidation. It has strong biological toxicity
and its production can aggravate biofilm damage. Therefore, it is
an index for membrane lipid peroxidation, which indirectly reflects
the degree of damage to a cell (16). In order to validate the effect of
GLY on the antioxidant capacity of a TNF-α/D-GalN-induced L02 cell
injury model, ROS and MDA levels were detected. As shown in
Fig. 1C and D, when compared with
the normal group, ROS and MDA levels in the model group were
significantly increased (P<0.01). Treatment with GLY reduced the
levels of ROS, MDA and LDH.
Effect of GLY on GSH and
Fe2+ levels in TNF-α/D-GalN- induced L02 cells
GSH is an important non-enzymatic antioxidant. GSH
plays an important role in restoring liver function through
removing lipid peroxides, including O2−,
H2O2 and LOOH (17). Therefore, the level of GSH is an
important factor in determining antioxidant capacity. GSH and
Fe2+ are key regulators of ROS in the process of
ferroptosis (18). In the process
of ferroptosis, Fe2+ can convert lipid peroxide into
ROS, and GPX4 converts lipid peroxide into the corresponding
alcohol with the help of GSH (19). As shown in Fig. 1E and F, when compared with the
normal group, the level of GSH was significantly decreased
(P<0.01) and the Fe2+ level in the model group was
significantly increased (P<0.01). After treatment with 1 or 2 mM
GLY, the levels of GSH were significantly increased (P<0.01) and
the levels of Fe2+ were significantly decreased
(P<0.01) compared with the model group. Therefore, it can be
concluded that GLY downregulates the levels of ROS by promoting GSH
and inhibiting Fe2+, thereby alleviating
ferroptosis.
GLY increases GPX4 expression and
promotes the Nrf2/HO-1/HMGB1 pathway in TNF-α/D-GalN-induced L02
cells
GPX4 competitively inhibits the production of ROS;
when the activity of GPX4 is inhibited, the effect of ROS on
ferroptosis is more subtle (20).
As shown in Fig. 2A-C, when
compared with the normal group, the protein levels of GPX4, Nrf2
and HO-1 in the model group were significantly reduced (P<0.01),
while the protein level of HMGB1 in the model group was increased
(P<0.01). After treatment with GLY, the levels of GPX4, Nrf2 and
HO-1 were significantly increased, while the protein level of HMGB1
in the model group was significantly decreased. When compared with
the normal group, the level of GPX4 in the model group was markedly
reduced (Fig. 2D), while the level
of HMGB1 in the model group was markedly increased (Fig. 2E), as determined by
immunofluorescence analysis. After treatment with 1 or 2 mM GLY,
the level of GPX4 was increased, while the level of HMGB1 in model
group was decreased.
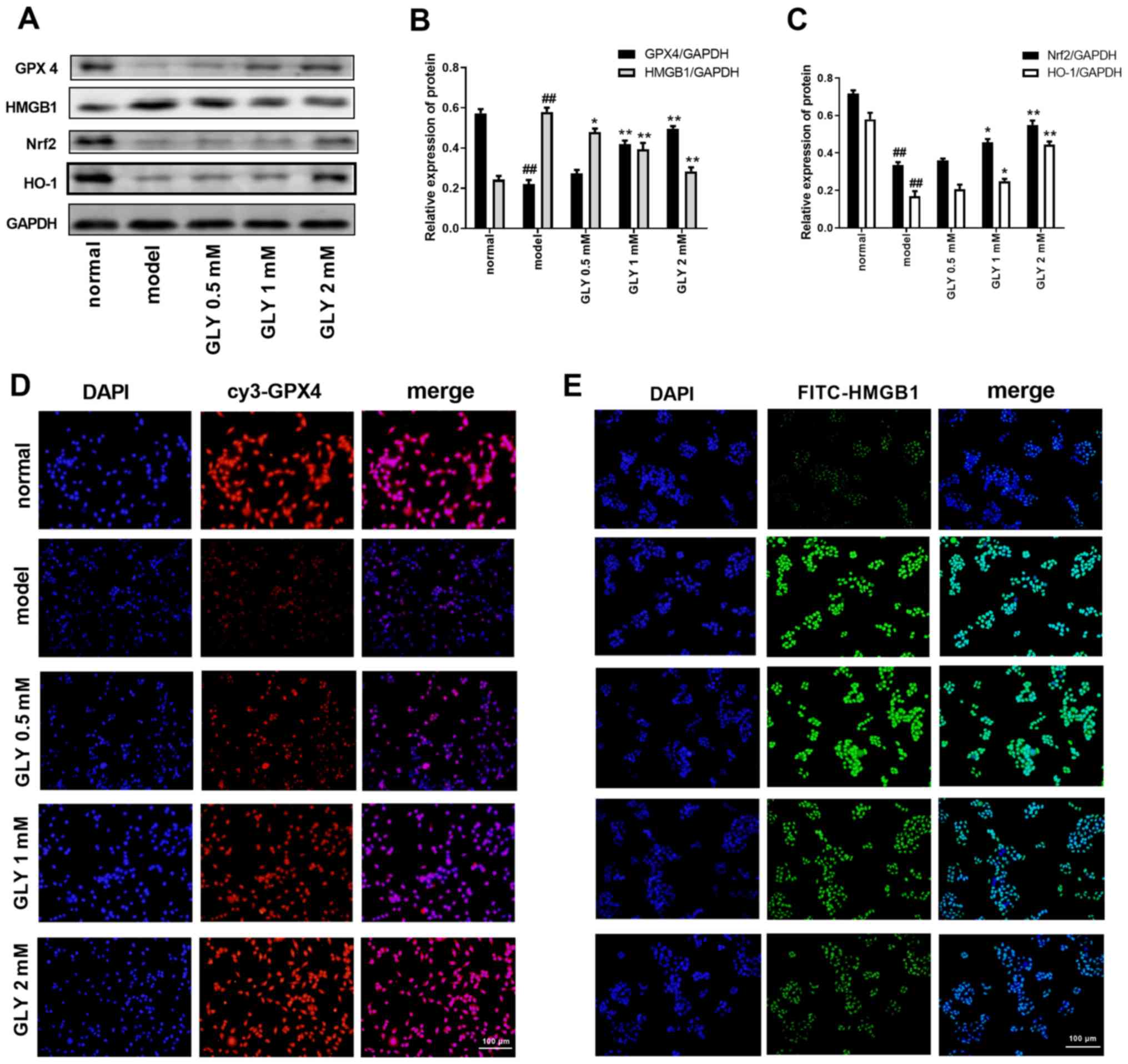 | Figure 2.GLY increased GPX4 expression and
promotes the Nrf2/HO-1/HMGB1 pathway in TNF-α/D-GalN induced L02
cells. (A) Protein levels of GPX4, HMGB1, Nrf2 and HO-1 were
determined by western blotting. Quantification of (B) GPX4 and
HMGB1, and (C) Nrf2 and HO-1. (D) GPX4 and (E) HMGB1 levels were
detected by immunofluorescence in L02 cells. n=3.
##P<0.01 vs. normal; **P<0.01, *P<0.05 vs.
model. TNF-α, tumour necrosis factor-α; D-GalN, D-galactosamine;
GLY, glycyrrhizin; GPX4, glutathione peroxidase 4; Nrf2, nuclear
factor erythroid 2-related factor 2; HO-1, heme oxygenase-1; HMGB1,
high mobility group protein B1. |
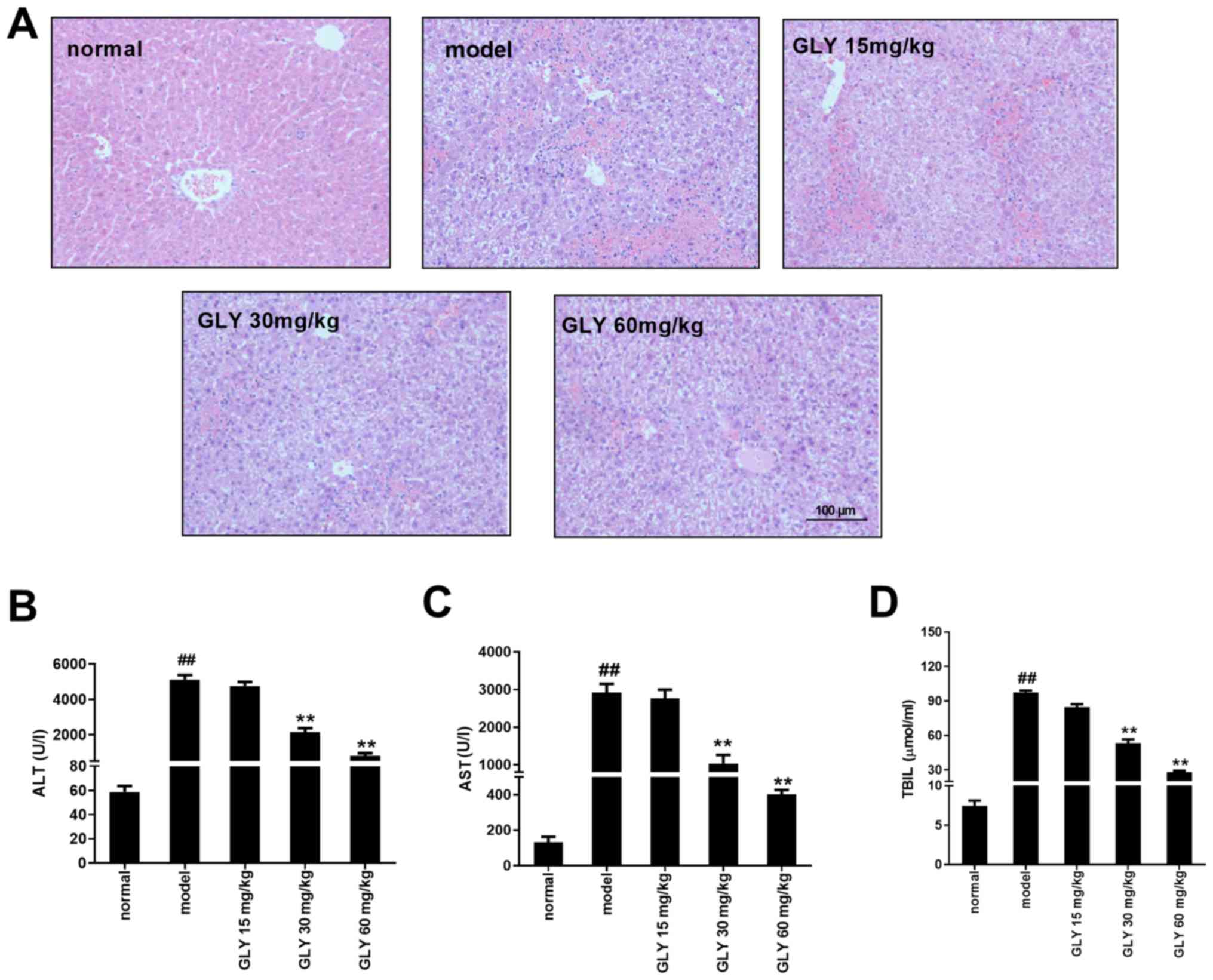
GLY reduces pathological changes in
the liver and improves liver function in ALF mice
As shown by the H&E staining in Fig. 3A, the liver tissue in the normal
group was clear and intact, with the liver plates neatly arranged.
There were no signs of degeneration, necrosis or inflammatory cell
infiltration. In the ALF model group, the structure of the liver
tissue was disordered and the hepatocytes were necrotic.
Inflammatory cell infiltration was also observed. The degree of
liver cell damage in the GLY treatment groups was reduced compared
with the model group. Inflammatory cell infiltration was also
reduced. The degree of liver damage was lower following the higher
doses of GLY. As shown in Fig.
3B-D the serum levels of ALT, AST and TBIL in the model group
were significantly higher than those in the control group
(P<0.01). Following treatment with GLY, the serum levels of ALT,
AST and TBIL were significantly lower than those in the ALF model
group; however, they were still higher than the normal group.
GLY enhances the anti-oxidative
capacity of LPS/D-GalN-induced ALF mouse liver
As shown in Fig.
4A-C, when compared with the normal group, the levels of LDH,
ROS and MDA in the model group were increased. After treatment with
GLY, the levels of LDH, ROS and MDA were decreased.
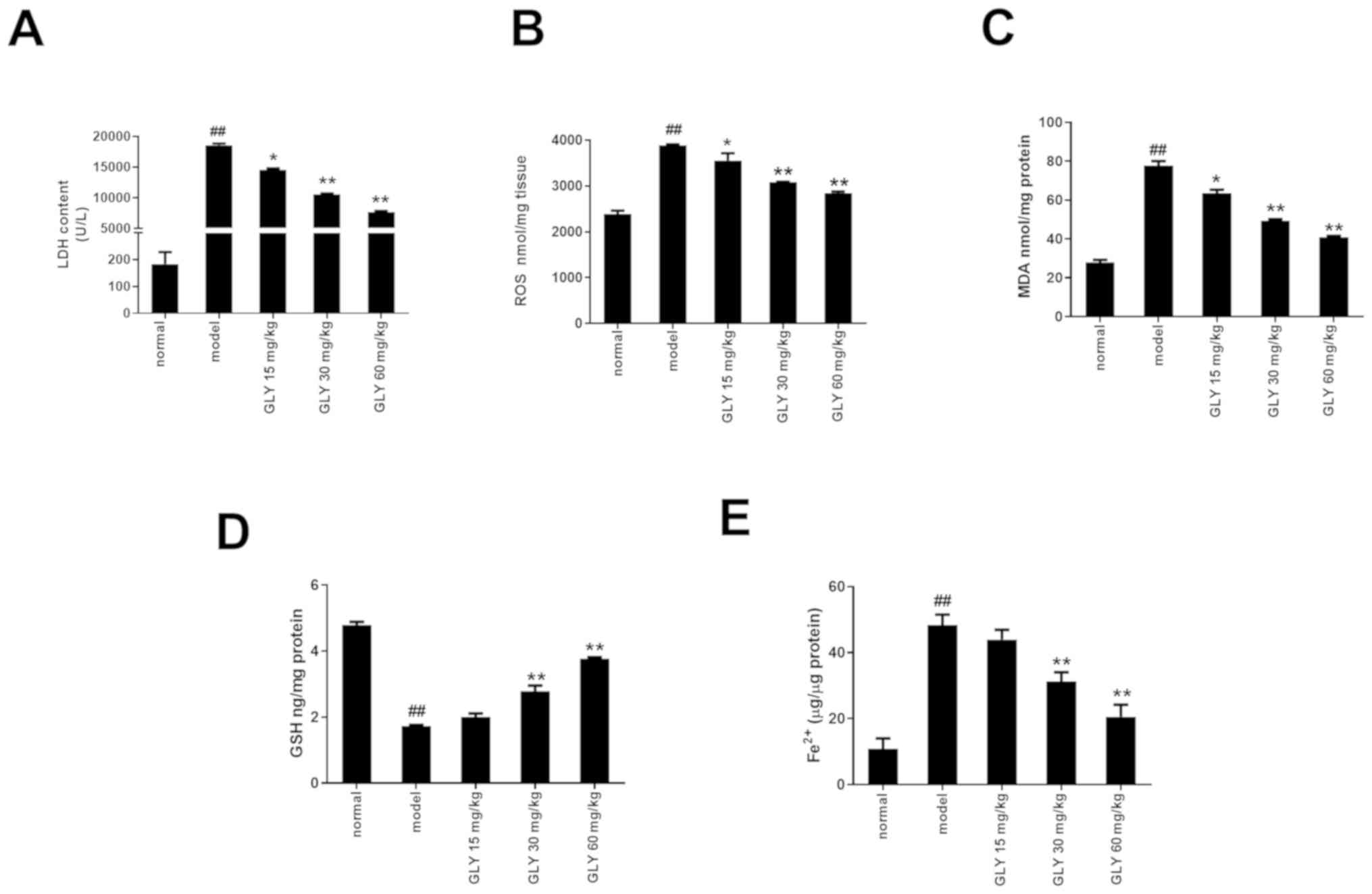 | Figure 4.GLY decreased oxidative damage in
LPS/D-GalN-induced ALF mice. GLY decreased the levels of (A) LDH,
(B) ROS and (C) MDA, and increased the level of (D) GSH in
LPS/D-GalN-induced ALF mice. (E) GLY decreased the level of
Fe2+ in LPS/D-GalN-induced ALF mice. n=3.
##P<0.01 vs. normal; **P<0.01, *P<0.05 vs.
model. ALF, acute liver failure; LPS, lipopolysaccharide; D-GalN,
D-galactosamine; GLY, glycyrrhizin; LDH, lactate dehydrogenase;
ROS, reactive oxygen species; MDA, malondialdehyde; GSH,
glutathione. |
GLY increases the level of GSH and
decreases the level of Fe2+ in LPS/D-GalN-induced ALF
mouse liver
As shown in Fig. 4D and
E, when compared with the normal group, the level of GSH was
significantly decreased (P<0.01), while the level of
Fe2+ in the model group was significantly increased
(P<0.01). After treatment with GLY, the level of GSH level was
increased (P<0.01), while the level of Fe2+
significantly decreased (P<0.01).
GLY increases the expression of GPX4
and promotes the Nrf2/HO-1/HMGB1 pathway in LPS/D-GalN-induced ALF
mouse liver
As shown in Fig.
5A-C, when compared with the normal group, the expression
levels of GPX4, Nrf2 and HO-1 in the model group were significantly
decreased (P<0.01), while the expression of HMGB1 in the model
group was significantly increased (P<0.01). After treatment with
GLY, the levels of GPX4, Nrf2 and HO-1 were increased (P<0.01),
while the level of HMGB1 was reduced. When compared with the normal
group, the protein level of GPX4 in the model group was reduced,
while the level of HMGB1 was increased, as determined by
immunofluorescence (Fig. 5D).
After treatment with GLY, the level of GPX4 was increased and the
level of HMGB1 was decreased.
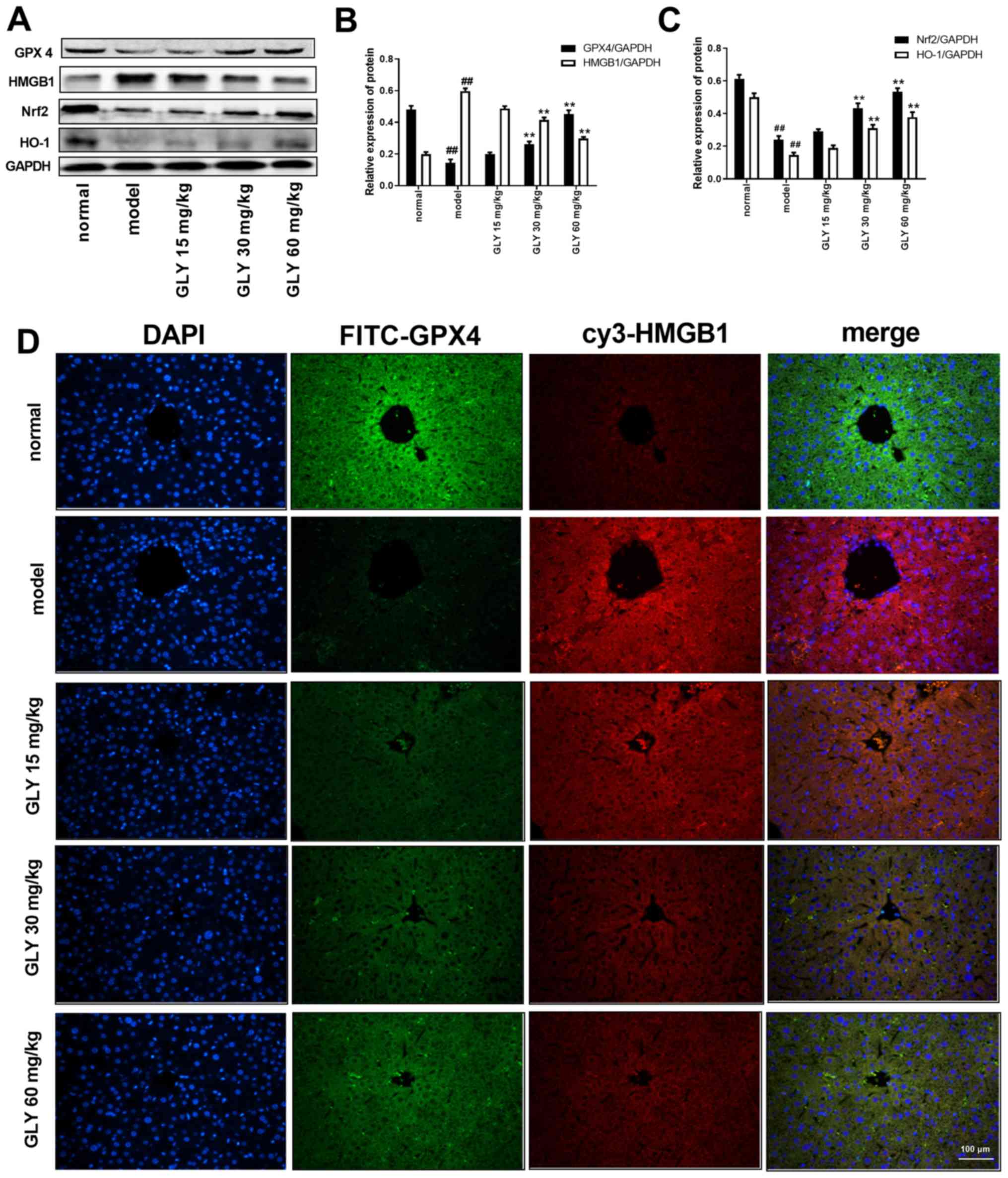 | Figure 5.GLY increases the expression of GPX4
and promotes the Nrf2/HO-1/HMGB1 pathway in LPS/D-GalN-induced ALF
mice. (A) Protein levels of GPX4, HMGB1, Nrf2 and HO-1 were
determined by western blotting. Quantification of (B) GPX4 and
HMGB1, and (C) Nrf2 and HO-1. (D) Protein levels of GPX4 and HMGB1
were determined by immunofluorescence in LPS/D-GalN-induced ALF
mice. n=3. ##P<0.01 vs. normal; **P<0.01 vs.
model. LPS, lipopolysaccharide; D-GalN, D-galactosamine; ALF, acute
liver failure; GLY, glycyrrhizin; GPX4, glutathione peroxidase 4;
Nrf2, nuclear factor erythroid 2-related factor 2; HO-1, heme
oxygenase-1; HMGB1, high mobility group protein B1. |
Discussion
LPS is present in the outer membrane of
gram-negative bacteria. LPS consists of an endotoxin-like portion
and a core sugar component, which consists of ~10 monosaccharides
(21). In the pathogenesis of
liver failure, enterogenous endotoxin can cause inflammatory cells
to secrete TNF-α, further damaging the liver cells; this is known
as the ‘two-hit’ theory (3). Liver
injury induced through the ‘two-hit’ mechanism is more serious than
that caused by LPS alone. LPS or TNF-α alone cause a lower specific
damage to hepatocytes (21).
Previous studies have shown that D-GalN can be used as a sensitizer
in LPS-induced liver injury; specifically, uridine triphosphate is
depleted by the galactose pathway, which inhibits protein
synthesis, resulting in ROS-mediated liver damage (22–24).
Therefore, LPS can be used in combination with D-GalN to induce ALF
in animal models (25,26) and TNF-α combined with D-GalN can be
used to create an L02 cell injury model (27); these models have been widely used
to explore and develop new liver protection agents for the
treatment of ALF.
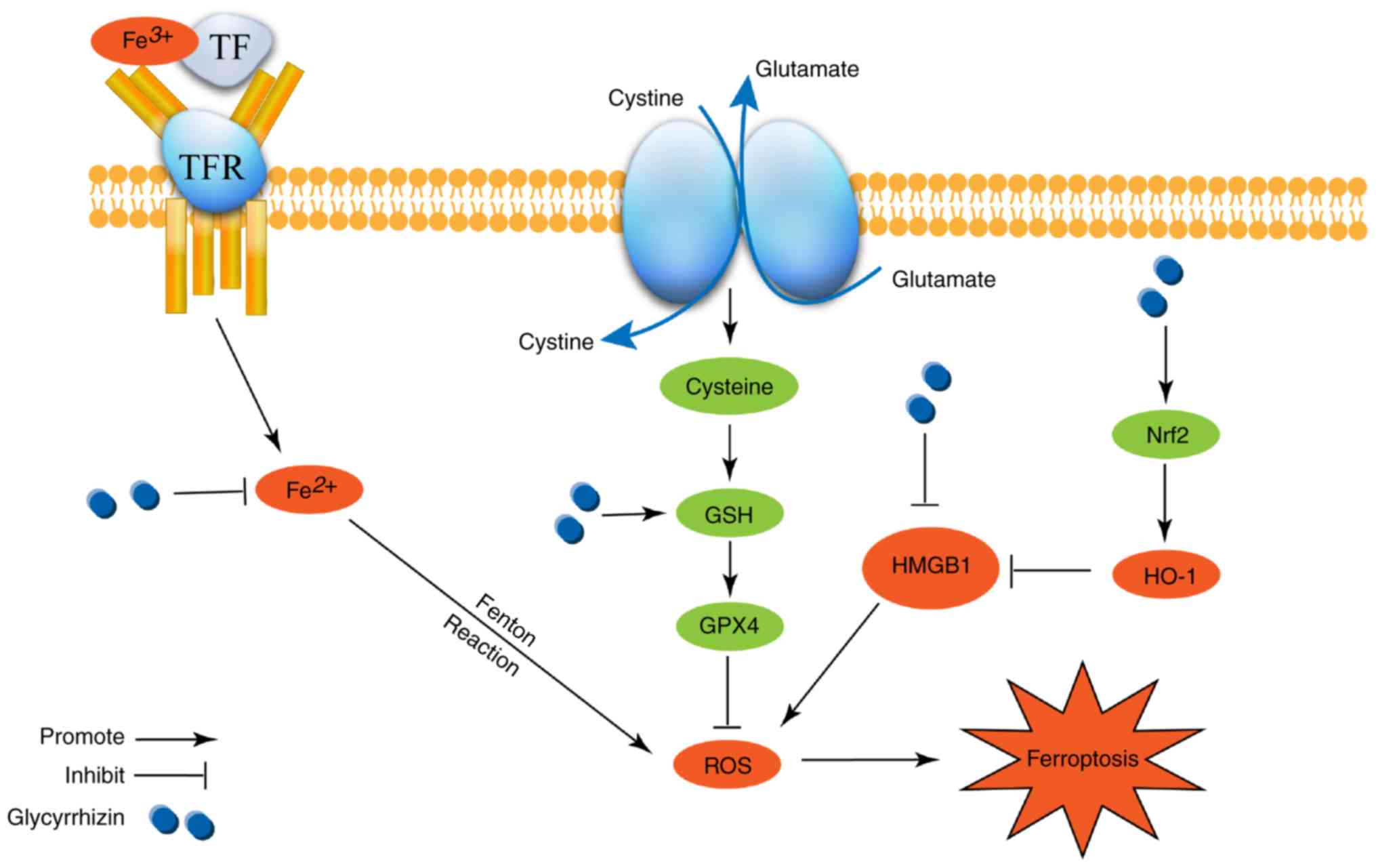
Ferroptosis is a type of cell death that is distinct
from apoptosis, necrosis, pyroptosis and autophagy. As is shown in
Fig. 6, ferroptosis is
predominantly characterized by iron homeostasis and ROS. Therefore,
Dixon et al (4) named this
mechanism of cell death ferroptosis. The morphological changes that
occur are due to decreased mitochondrial volume, increased lipid
bilayer membrane density and the decrease or disappearance of
mitochondria (28). Research into
the mechanism of ferroptosis predominantly focuses on the imbalance
of iron homeostasis and the decreased activity of GPX4 (29). Circulating iron in the body mainly
binds to transferrin and exists as Fe3+. Fe3+
enters cells through the membrane protein transferrin receptor 1
(30). Fe3+ is reduced
to Fe2+ by iron oxidoreductase six-transmembrane
epithelial antigen of the prostate 3 (31). In an unbalanced state, the level of
Fe2+ increases inside cells, which is an important
factor in the process of ferroptosis. The key process of
ferroptosis is the Fenton reaction, in which Fe2+
converts lipid peroxide into ROS. While GPX4 converts lipid
peroxide into the corresponding alcohol with the help of GSH
(19). The ROS produced by this
process could cause oxidatively damage and cell death. At the same
time, GPX4 competitively inhibits the production of ROS, and when
its activity is inhibited, ferroptosis occurs (20). Therefore, the excessive production
of ROS is important to induce ferroptosis. Moreover, the detection
of ferroptosis has no specific indicators (32). The changes to the levels of GPX4,
Fe2+, GSH and ROS could be combined to define
ferroptosis.
Research into ferroptosis in liver disease is
predominantly focused on the regulation of oxidative stress,
focusing on hepatocellular carcinoma and drug-induced liver injury.
It has been reported that sorafenib can induce ferroptosis in tumor
cells through the p62-kelch-like ECH-associated protein 1-NRF2
pathway to achieve anti-tumor efficacy (33). Acetaminophen can aggravate
oxidative stress and induce ferroptosis (34). In addition, survival is increased
in vitamin E-fed GPX4-deficient mouse models by inhibiting lipid
oxidation (35). Oxidative stress
caused by ROS and other free radicals/oxidants plays an important
role in the pathogenesis of atherosclerosis, diabetes, ischemic
stroke and related central nervous system diseases (36,37).
When the body is damaged by oxidation, various anti-oxidant
molecules and enzymes can be produced to remove the ROS generated,
and protect the body from ROS damage (38). GSH is an important non-enzymatic
antioxidant in the body. GSH content is an important biomarker for
measuring antioxidant capacity, while LDH can be used as a cell
damage marker (37). GSH is an
important cofactor for GPX4. When the synthesis of GSH synthesis is
blocked, the activity of GPX4 is decreased, as is the cellular
antioxidant capacity, which leads to the accumulation of ROS and
promotes ferroptosis (39). Nrf2
is also an important endogenous antioxidant mechanism. By
activating Nrf2, the expression of downstream signaling molecules,
such as HO-1, can be induced to exert an anti-oxidative stress
response (40). Previous studies
have shown that the upregulation of the Nrf2/HO-1 signaling
pathway, and inhibition of HMGB1 expression, has an
anti-inflammatory effect, attenuating ischemia-reperfusion injury
and providing cell protection (41–43).
HMGB1 is a potent inflammatory factor that plays a key role in the
initiation and maintenance of inflammatory cascade responses
(44,45). Large amounts of hepatocyte necrosis
during liver failure can lead to the release of HMGB1 from
hepatocytes, as HMGB1 has a similar effect to endotoxin, HMGB1 is
also involved in the pathogenesis of ALF (46). Furthermore, the exposure of normal
human bronchial epithelial cells to HMGB1 resulted in an increase
in the levels of ROS (47).
Therefore, if oxidative stress can be prevented through the
Nrf2/HO-1/HMGB1 pathway to inhibit ferroptosis, ALF can be
alleviated.
GLY acts as an anti-inflammatory, anti-oxidant and
immune-regulator and can protect hepatocytes. As a chronic
hepatitis drug, GLY has been used in clinical practice for many
years with good therapeutic effects (12,48).
Previous studies have shown that GLY could exert a good effect on
alcoholic liver disease (49),
non-alcoholic fatty liver disease (50), intrahepatic cholestasis (51), metabolic syndrome-induced liver
damage (52), acute liver injury
(53), liver fibrosis (11) and hepatocellular carcinoma
(54).
GLY is a verified selective inhibitor of HMGB1
(55). GLY directly binds to HMG
boxes, blocking HMGB1 release into the extracellular space. The
specific mechanism involves the interaction of GLY with two shallow
concave surfaces formed by the two arms of the HMG boxes (55,56).
Our previous study showed that HMGB1 plays an important role in the
development of ALF (57). High
expression of HMGB1 is present in patients with liver failure
caused by chronic hepatitis B virus (HBV) infection. HMGB1 is
involved in the inflammatory response during liver failure by
inhibiting the activity of T regulatory cells in chronically
infected HBV (57). However, the
relationship between HMGB1 and ferroptosis in ALF is still unclear.
GLY has been rarely reported in the study of ALF and ferroptosis in
ALF. In the present study, when L02 cell and mouse models of ALF
were induced using TNF-α/D-GalN or LPS/D-GalN, respectively, the
levels of GSH GPX4, and cell viability were decreased in the cell
and mouse models of ALF compared with the normal group,
Fe2+ and ROS levels were increased. The changes in these
molecules indicated that the ferroptosis/ALF model had been
successfully induced. Moreover, the levels of MDA and LDH were
increased, and the Nrf2/HO-1/HMGB1 pathway was inhibited,
supporting the hypothesis of oxidative stress injury in
ferroptosis. The severity of the histopathologic changes in the
liver of the model group mice was notable. Serum ALT, AST and TBIL
levels were increased. However, after treatment with GLY, the
degree of liver damage was reduced, the level of HMGB1 was also
reduced, and the levels of GPX4, Nrf2 and HO-1 were increased. The
levels of LDH, Fe2+, MDA and ROS were decreased by GLY
treatment. The level of GSH was increased by GLY treatment, as was
cell viability. The results of the present study are consistent
with those of a previous study that found GLY could ameliorate
obesity by activating the Nrf2/HO-1 pathway (58). After treatment with GLY, the
severity of the histopathologic changes observed in the liver was
reduced. Serum ALT, AST and TBIL levels were decreased by GLY.
Therefore, the present study indicated that GLY could increase
anti-oxidative capacity, GSH and GPX4 levels, and active the
Nrf2/HO-1/HMGB1 pathway, inhibiting ferroptosis in an
LPS/D-GalN-induced ALF model.
In conclusion, GLY reduced the level of ferroptosis
during ALF, and the mechanism may depend on the inhibition of the
oxidative stress pathway. However, the pharmacological effects of
GLY are diverse. From the present study, it was found that GLY
plays a role in multiple aspects of ferroptosis in ALF, including
the regulation of the level of Fe2+, the GSH/GPX4
pathway and the Nrf2/HO-1/HMGB1 pathway (Fig. 6). These three functions of GLY
would inhibit ROS accumulation, inhibiting ferroptosis and
alleviating ALF. In the future, the specific mechanisms of GLY in
the process of ferroptosis and liver failure should be addressed.
The present study provides a scientific basis for the treatment of
ALF and the inhibition of ferroptosis.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Natural
Science Foundation of China (grant no. 81870413).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
ZG and YW conceived and designed the experiments.
YW, QC, CS and FJ performed the experiments. YW analyzed the data.
QC and CS contributed reagents/materials/analysis tools. YW wrote
the paper. ZG edited the article. All authors read and approved the
final version of the manuscript and agree to take responsibility
for the published article.
Ethics approval and consent to
participate
The present study was approved by The Institutional
Animal Care and Use Committee of Renmin Hospital of Wuhan
University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bernal W, Auzinger G, Dhawan A and Wendon
J: Acute liver failure. Lancet. 376:190–201. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liu Y, Wang Y, Chen Q, Jiao F, Wang L and
Gong Z: HDAC2 inhibitor CAY10683 reduces intestinal epithelial cell
apoptosis by inhibiting mitochondrial apoptosis pathway in acute
liver failure. Histol Histopathol. 29:181202019.
|
|
3
|
Wang Y, Chen H, Chen Q, Jiao FZ, Zhang WB
and Gong ZJ: The protective mechanism of CAY10683 on intestinal
mucosal barrier in acute liver failure through LPS/TLR4/MyD88
pathway. Mediators Inflamm. 13:78596012018.
|
|
4
|
Dixon SJ, Lemberg KM, Lamprecht MR, Skouta
R, Zaitsev EM, Gleason CE, Patel DN, Bauer AJ, Cantley AM, Yang WS,
et al: Ferroptosis: An iron-dependent form of nonapoptotic cell
death. Cell. 149:1060–1072. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wei L, Ren F, Zhang X, Wen T, Shi H, Zheng
S, Zhang J, Chen Y, Han Y and Duan Z: Oxidative stress promotes
D-GalN/LPS-induced acute hepatotoxicity by increasing glycogen
synthase kinase 3β activity. Inflamm Res. 63:485–494. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mishra D, Jain N, Rajoriya V and Jain AK:
Glycyrrhizin conjugated chitosan nanoparticles for
hepatocyte-targeted delivery of lamivudine. J Pharm Pharmacol.
66:1082–1093. 2014.PubMed/NCBI
|
|
7
|
Nazari S, Rameshrad M and Hosseinzadeh H:
Toxicological effects of glycyrrhiza glabra (Licorice): A review.
Phytother Res. 31:1635–1650. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Michaelis M, Geiler J, Naczk P, Sithisarn
P, Leutz A, Doerr HW and Cinatl J Jr: Glycyrrhizin exerts
antioxidative effects in H5N1 influenza A virus-infected cells and
inhibits virus replication and pro-inflammatory gene expression.
PLoS One. 6:e197052011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ojha S, Javed H, Azimullah S, Abul Khair
SB and Haque ME: Glycyrrhizic acid attenuates neuroinflammation and
oxidative stress in rotenone model of parkinson's disease. Neurotox
Res. 29:275–287. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang XR, Hao HG and Chu L: Glycyrrhizin
inhibits LPS-induced inflammatory mediator production in
endometrial epithelial cells. Microb Pathog. 109:110–113. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhou Y, Tong X, Ren S, Wang X, Chen J, Mu
Y, Sun M, Chen G, Zhang H and Liu P: Synergistic anti-liver
fibrosis actions of total astragalus saponins and glycyrrhizic acid
via TGF-β1/Smads signaling pathway modulation. J Ethnopharmacol.
190:83–90. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lin CC and Wang PH: Intravenous
glycyrrhizin improved serum transaminases rapidly in a chronic
hepatitis B patient with acute exacerbation. J Formosan Med Assoc.
114:188–189. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kong ZH, Chen X, Hua HP, Liang L and Liu
LJ: The oral pretreatment of glycyrrhizin prevents surgery-induced
cognitive impairment in aged mice by reducing neuroinflammation and
alzheimer's-related pathology via HMGB1 inhibition. J Mol Neurosci.
63:385–395. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mittler R, Vanderauwera S, Suzuki N,
Miller G, Tognetti VB, Vandepoele K, Gollery M, Shulaev V and Van
Breusegem F: ROS signaling: The new wave? Trends Plant Sci.
16:300–309. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xie Y, Hou W, Song X, Yu Y, Huang J, Sun
X, Kang R and Tang D: Ferroptosis: Process and function. Cell Death
Differ. 23:369–379. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Barrera G, Pizzimenti S, Daga M, Dianzani
C, Arcaro A, Cetrangolo GP, Giordano G, Cucci MA, Graf M and
Gentile F: Lipid peroxidation-derived aldehydes, 4-hydroxynonenal
and malondialdehyde in aging-related disorders. Antioxidants
(Basel). 30:82018.
|
|
17
|
Levine WG: Glutathione and hepatic
mixed-function oxidase activity. Drug Metab Rev. 14:909–930. 1983.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Du J, Wang T, Li Y, Zhou Y, Wang X, Yu X,
Ren X, An Y, Wu Y, Sun W, et al: DHA inhibits proliferation and
induces ferroptosis of leukemia cells through autophagy dependent
degradation of ferritin. Free Radic Biol Med. 131:356–369. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Proneth B and Conrad M: Ferroptosis and
necroinflammation, a yet poorly explored link. Cell Death Differ.
26:14–24. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Latunde-Dada GO: Ferroptosis: Role of
lipid peroxidation, iron and ferritinophagy. Biochimica Biophysica
Acta Gen Subj. 1861:1893–1900. 2017. View Article : Google Scholar
|
|
21
|
Hamesch K, Borkham-Kamphorst E, Strnad P
and Weiskirchen R: Lipopolysaccharide-induced inflammatory liver
injury in mice. Lab Anim. 49:37–46. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Maes M, Vinken M and Jaeschke H:
Experimental models of hepatotoxicity related to acute liver
failure. Toxicol Appl Pharmacol. 290:86–97. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhuo T, Zhou S, Zhang W, Lambertucci C and
Volpini R: Synthesis and ability of new ligands for G
protein-coupled receptors 17 (GPR17). Med Sci Monit. 23:953–959.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Song J, Lu C, Zhao W and Shao X: Melatonin
attenuates TNF-α-mediated hepatocytes damage via inhibiting
mitochondrial stress and activating the Akt-Sirt3 signaling
pathway. J Cell Physiol. 234:20969–20979. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shang Y, Liu Y, Du L, Wang Y, Cheng X,
Xiao W, Wang X, Jin H, Yang X, Liu S and Chen Q: Targeted
expression of uncoupling protein 2 to mouse liver increases the
susceptibility to lipopolysaccharide/galactosamine-induced acute
liver injury. Hepatology. 50:1204–1216. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chai FN, Zhang J, Xiang HM, Xu HS, Li YF,
Ma WY, Li XG and Ye XL: Protective effect of coptisine from rhizoma
coptidis on LPS/D-GalN-induced acute liver failure in mice through
up-regulating expression of miR-122. Biomed Pharmacother.
98:180–190. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yan X, Jiang Z, Bi L, Yang Y and Chen W:
Salvianolic acid A attenuates TNF-α- and D-GalN-induced ER
stress-mediated and mitochondrial-dependent apoptosis by modulating
Bax/Bcl-2 ratio and calcium release in hepatocyte LO2 cells. Naunyn
Schmiedebergs. Arch Pharmacol. 388:817–830. 2015. View Article : Google Scholar
|
|
28
|
Yu H, Guo P, Xie X, Wang Y and Chen G:
Ferroptosis, a new form of cell death, and its relationships with
tumourous diseases. J Cell Mol Med. 21:648–657. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yoshida M, Minagawa S, Araya J, Sakamoto
T, Hara H, Tsubouchi K, Hosaka Y, Ichikawa A, Saito N, Kadota T, et
al: Involvement of cigarette smoke-induced epithelial cell
ferroptosis in COPD pathogenesis. Nat Commun. 10:31452019.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Anderson GJ and Frazer DM: Current
understanding of iron homeostasis. Am J Clin Nutr. 106 (Suppl
6):1559S–1566S. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gauss GH, Kleven MD, Sendamarai AK,
Fleming MD and Lawrence CM: The crystal structure of
six-transmembrane epithelial antigen of the prostate 4 (Steap4), a
ferri/cuprireductase, suggests a novel interdomain flavin-binding
site. J Biol Chem. 288:20668–20682. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen D, Eyupoglu IY and Savaskan N:
Ferroptosis and cell death analysis by flow cytometry. Methods Mol
Biol. 1601:71–77. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sun X, Ou Z, Chen R, Niu X, Chen D, Kang R
and Tang D: Activation of the p62-Keap1-NRF2 pathway protects
against ferroptosis in hepatocellular carcinoma cells. Hepatology.
63:173–184. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hu J, Kholmukhamedov A, Lindsey CC, Beeson
CC, Jaeschke H and Lemasters JJ: Translocation of iron from
lysosomes to mitochondria during acetaminophen-induced
hepatocellular injury: Protection by starch-desferal and
minocycline. Free Radic Biol Med. 97:418–426. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Carlson BA, Tobe R, Yefremova E, Tsuji PA,
Hoffmann VJ, Schweizer U, Gladyshev VN, Hatfield DL and Conrad M:
Glutathione peroxidase 4 and vitamin E cooperatively prevent
hepatocellular degeneration. Redox Biol. 9:22–31. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ritchie RH, Drummond GR, Sobey CG, De
Silva TM and Kemp-Harper BK: The opposing roles of NO and oxidative
stress in cardiovascular disease. Pharmacol Res. 116:57–69. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Niemann B, Rohrbach S, Miller MR, Newby
DE, Fuster V and Kovacic JC: Oxidative stress and cardiovascular
risk: Obesity, diabetes, smoking, and pollution part 3 of a 3-part
series. J Am Coll Cardiol. 70:230–251. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Henchcliffe C and Beal MF: Mitochondrial
biology and oxidative stress in parkinson disease pathogenesis.
Nature Clin Pract Neurol. 4:600–609. 2008. View Article : Google Scholar
|
|
39
|
Maiorino M, Conrad M and Ursini F: GPx4,
Lipid peroxidation, and cell death: Discoveries, rediscoveries, and
open issues. Antioxid Redox Signal. 29:61–74. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ma Q: Role of nrf2 in oxidative stress and
toxicity. Annu Rev Pharmacol Toxicol. 53:401–426. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chang CY, Kao TK, Chen WY, Ou YC, Li JR,
Liao SL, Raung SL and Chen CJ: Tetramethylpyrazine inhibits
neutrophil activation following permanent cerebral ischemia in
rats. Biochem Biophys Res Commun. 463:421–427. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kim SR, Ha YM, Kim YM, Park SW, Kim HJ,
Chung HT and Chang KC: Ascorbic acid reduces HMGB1 secretion in
lipopolysaccharide-activated RAW 264.7 cells and improves survival
rate in septic mice by activation of Nrf2/HO-1 signals. Biochem
Pharmacol. 95:279–289. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Liu C, Zhu C, Wang G, Xu R and Zhu Y:
Higenamine regulates Nrf2-HO-1-Hmgb1 axis and attenuates intestinal
ischemia-reperfusion injury in mice. Inflammation Res. 64:395–403.
2015. View Article : Google Scholar
|
|
44
|
Andersson U and Tracey KJ: HMGB1 is a
therapeutic target for sterile inflammation and infection. Ann Rev
Immunol. 29:139–162. 2011. View Article : Google Scholar
|
|
45
|
Yu M, Wang HC, Ding AH, Golenbock DT, Latz
E, Czura CJ, Fenton MJ, Tracey KJ and Yang H: HMGB1 signals through
toll-like receptor (TLR) 4 and TLR2. Shock. 26:174–179. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yang RK, Zou XP, Tenhunen J and Tonnessen
TI: HMGB1 and extracellular histones significantly contribute to
systemic inflammation and multiple organ failure in acute liver
failure. Mediators Inflamm. 2017:59280782017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lv YH, Li YL, Zhang DD, Zhang AB, Guo WH
and Zhu SF: HMGB1-induced asthmatic airway inflammation through
GRP75-mediated enhancement of ER-mitochondrial Ca2+
transfer and ROS increased. J Cell Biochem. 119:4205–4215. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Carreno V: Review article: Management of
chronic hepatitis C in patients with contraindications to
anti-viral therapy. Aliment Pharmacol Ther. 39:148–162. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Milosevic N, Milanovic M, Turkulov V,
Medic-Stojanoska M, Abenavoli L and Milic N: May patients with
alcohol liver disease benefit from herbal medicines? Rev Recent
Clin Trials. 11:227–237. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Sun X, Duan X, Wang C, Liu Z, Sun P, Huo
X, Ma X, Sun H, Liu K and Meng Q: Protective effects of
glycyrrhizic acid against non-alcoholic fatty liver disease in
mice. Eur J Pharmacol. 806:75–82. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wang H, Fang ZZ, Meng R, Cao YF, Tanaka N,
Krausz KW and Gonzalez FJ: Glycyrrhizin and glycyrrhetinic acid
inhibits alpha-naphthyl isothiocyanate-induced liver injury and
bile acid cycle disruption. Toxicology. 386:133–142. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Sil R, Ray D and Chakraborti AS:
Glycyrrhizin ameliorates metabolic syndrome-induced liver damage in
experimental rat model. Mol Cell Biochem. 409:177–189. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Yan T, Wang H, Zhao M, Yagai T, Chai Y,
Krausz KW, Xie C, Cheng X, Zhang J, Che Y, et al: Glycyrrhizin
protects against acetaminophen-induced acute liver injury via
alleviating tumor necrosis factor α-mediated apoptosis. Drug Metab
Dispos. 44:720–731. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Zhang X, Yang H, Yue S, He G, Qu S, Zhang
Z, Ma B, Ding R, Peng W, Zhang H, et al: The mTOR inhibition in
concurrence with ERK1/2 activation is involved in excessive
autophagy induced by glycyrrhizin in hepatocellular carcinoma.
Cancer Med. 6:1941–1951. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Mollica L, De Marchis F, Spitaleri A,
Dallacosta C, Pennacchini D, Zamai M, Agresti A, Trisciuoglio L,
Musco G and Bianchi ME: Glycyrrhizin binds to high-mobility group
box 1 protein and inhibits its cytokine activities. Chem Biol.
14:431–441. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Li YJ, Wang L, Zhang B, Gao F and Yang CM:
Glycyrrhizin, an HMGB1 inhibitor, exhibits neuroprotective effects
in rats after lithium-pilocarpine-induced status epilepticus. J
Pharm Pharmacol. 71:390–399. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Wang LW, Chen H and Gong ZJ: High mobility
group box-1 protein inhibits regulatory T cell immune activity in
liver failure in patients with chronic hepatitis B. Hepatobiliary
Pancreat Dis Int. 9:499–507. 2010.PubMed/NCBI
|
|
58
|
Abo El-Magd NF, El-Mesery M, El-Karef A
and El-Shishtawy MM: Glycyrrhizin ameliorates high fat diet-induced
obesity in rats by activating NrF2 pathway. Life Sci. 193:159–170.
2018. View Article : Google Scholar : PubMed/NCBI
|