Introduction
Diabetic retinopathy (DR) is a major complication of
diabetes in the working-age population, and causes serious loss of
vision or even blindness (1–3). At
present, hyperglycemia is the main cause of the development of this
disease, which can cause pathological metabolism and biochemical
changes that damage retinal cells (4). During the development of DR, retinal
endothelial cell (REC) dysfunction is a crucial initiator of
multifactorial pathology, which depends on metabolic abnormalities
and inflammation (5–9). There is evidence that high glucose
(HG)-induced aseptic inflammation is associated with the
development and progression of REC injury (10–13).
Retinal pigment epithelial (RPE) cells are important component of
the external blood-retinal barrier that selectively regulates the
flow of molecules into and out of the retina (14). In the development of DR, retinal
micro-vascular dysfunction involves the loss of endothelial cells
and pericytes, capillary occlusion and vascular barrier
destruction, and endothelial cell hypertrophy and degeneration,
leading to capillary perfusion and hypoxia (15,16).
MicroRNAs (miRNAs/miRs) are a group of endogenous,
single stranded, small non-coding RNAs (21–25 nucleotides in
length), which can post-transcriptionally regulate target gene
expression by complementarily binding to their 3′-untranslated
region (3′-UTR) (17–20). It has been demonstrated that miRNAs
are involved in many cellular biological processes in normal
physiology and pathogenesis, such as differentiation, cell growth,
apoptosis and inflammation (21).
Previously, studies have reported that miR-217 participated in the
regulation of a variety of biological processes, such as cell
growth, apoptosis, differentiation and metastasis in various types
of cells (22,23). In addition, miR-217 was determined
to be abnormally expressed in a variety of tumor cells, including
pancreatic adenocarcinoma, osteosarcoma, cervical carcinoma and
lung cancer (24–27). miR-217 inhibition has been found to
upregulate the hypoxia inducible factor (HIF)-1α/vascular
endothelial growth factor pathway to promote angiogenesis and
ameliorate inflammation of diabetic foot ulcer rats (28). Sun et al (29) reported that miR-217 inhibition can
protectively antagonize HG-induced podocyte damage and insulin
resistance by restoring the defective autophagy pathway via
targeting phosphatase and tensin homolog, indicating that miR-217
was a promising therapeutic target for diabetic nephropathy. Shao
et al (30) suggested that
miR-217 promotes inflammation and fibrosis in HG-cultured rat
glomerular mesangial cells via the Sirtuin 1 (Sirt1)/HIF-1α
signaling pathway. Additionally, miR-217 has been reported to be
related to the development of proteinuria in type 2 diabetes
patients; serum miR-217 may be involved in the development of
diabetic kidney disease through promoting chronic inflammation,
renal fibrosis and angiogenesis (31). These results indicated that miR-217
plays an important role in diabetes and its complications; however,
the role of miR-217 in HG-induced retinal epithelial cell damage
remains unclear.
Therefore, in the present study, we aimed to
investigate the role of miR-217 in HG-induced retinal epithelial
cell damage and its molecular mechanisms to determine the role of
miR-217 in diabetic retinopathy.
Materials and methods
Cell culture and HG treatment
The RPE cell line ARPE-19 was acquired from American
Type Culture Collection (ATCC; cat. no. ATCC® CRL-2302)
and cultured in Dulbecco's Modified Eagle's medium (DMEM; Gibco;
Thermo Fisher Scientific, Inc.) supplemented with 10% fetal bovine
serum (Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin/streptomycin (Beyotime Institute of Biotechnology) at
37°C in a humidified incubator with 5% CO2.
For HG treatment, ARPE-19 cells were treated with 50
mM D-glucose (Beyotime Institute of Biotechnology) at 37°C for 24
h. Cells cultured in DMEM without glucose served as the control.
The cultures were conducted in triplicate.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) assay
Total RNA was extracted from cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) as per the manufacturer's protocol. RNA concentration was
measured using a NanoDrop™ 2000 spectrophotometer (NanoDrop
Technologies; Thermo Fisher Scientific, Inc.). RT was conducted
with 1 µg total RNA via a PrimeScript reverse transcription reagent
kit (Takara Biotechnology Co., Ltd.) according to the
manufacturer's protocols. RT conditions were as follows: 42°C for
60 min and 75°C for 5 min. Then, qPCR was performed using the Fast
SYBR™ Green Master Mix (Thermo Fisher Scientific, Inc.) using the
CFX Connect Real-Time System (Bio-Rad Laboratories, Inc.). The
thermocycling conditions were as follows: Initial denaturation at
95°C for 5 min and 40 cycles of denaturation at 95°C for 10 sec,
annealing at 60°C for 10 sec, and extension at 72°C for 30 sec. U6
for miRNA and GAPDH for mRNA were used as internal controls. The
primer sequences for qPCR were as follows: U6, forward
5′-GCTTCGGCAGCACATATACTAAAAT-3′; reverse
5′-CGCTTCACGAATTTGCGTGTCAT-3′; GAPDH, forward
5′-CTTTGGTATCGTGGAAGGACTC-3′; miR-217, forward
5′-TACTGCATCAGGAACTGACTGGA-3′; reverse 5′-GTGCAGGGTCCGAGGT-3′;
SIRT1, forward 5′-AATCCAGTCATTAAAGGTCTACAA-3′; reverse
5′-TAGGACCATTACTGCCAGAGG-3′; reverse 5′-GTAGAGGCAGGGATGATGTTCT-3′.
The 2−ΔΔCq method (32)
was used to quantify the relative expression of genes.
Dual-luciferase reporter assay
Bioinformatics software (TargetScan 7.2, http://www.targetscan.org/vert_72/) was used to
predict target gene of miR-217. The results revealed the binding
sites between the 3′-UTR of SIRT1 and miR-217. To verify whether
miR-217 could target SIRT1, we performed a dual-luciferase reporter
assay. The wild type (WT-SIRT1) and mutant (MUT-SIRT1) 3′-UTR of
SIRT1 were respectively cloned into a pmiR-RB-Report™ dual
luciferase reporter gene plasmid vector (Guangzhou RiboBio Co.,
Ltd.) according to the manufacturer's instructions. To point-mutate
the miR-217 binding domain on the 3′UTR of SIRT1, a QuikChange
Site-Directed Mutagenesis kit (Stratagene; Agilent Technologies,
Inc.) was performed following the manufacturer's instructions.
Then, 50 nM miR-217 mimic (agomir; 5′-UACUGCAUCAGGAACUGAUUGGA-3′;
Shanghai GenePharma Co., Ltd.) or 50 nM mimic control
(5′-UUUGUACUACACAAAAGUACUG-3′; Shanghai GenePharma Co., Ltd.), and
the WT or MUT 3′-UTR of SIRT1 were co-transfected into ARPE-19
cells using Lipofectamine® 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.) for 48 h. Subsequently, relative
luciferase activity was detected using a Dual Luciferase Assay
System (Promega Corporation) with a microplate reader (Molecular
Devices, LLC). Renilla luciferase was used for the
normalization of firefly luciferase activity. Each experiment was
performed three times.
Western blotting assay
Total protein from ARPE-19 cells was extracted using
radioimmunoprecipitation assay buffer (Beyotime Institute of
Biotechnology). We used Bicinchoninic Acid Protein Assay kit
(Thermo Fisher Scientific, Inc.) to measure protein concentration.
A total of 20 µg of protein from each sample was subjected to 12%
SDS-PAGE electrophoresis and transferred to polyvinylidene
difluoride membranes (Pierce; Thermo Fisher Scientific, Inc.).
Then, the membrane was blocked with 5% skim milk at room
temperature for 1.5 h. After washing with PBST three times, the
membrane was probed with primary antibodies against SIRT1 (1:1,000;
cat. no. 9475; Cell Signaling Technology, Inc.), phosphorylated
(p)-NF-κB p65 (p-p65; 1:1,000; cat. no. 3033; Cell Signaling
Technology, Inc.), p65 (1:1,000; cat. no. 8242; Cell Signaling
Technology, Inc.), or β-actin (45 kDa; 1:1,000; cat. no. 4970; Cell
Signaling Technology, Inc.), at 4°C overnight. The next day, the
membrane was washed three times with PBST buffer and then incubated
with HRP-conjugated secondary antibody (1:2,000; cat. no. 7074;
Cell Signaling Technology, Inc.) at room temperature for 2 h. The
residue antibody solution was completely washed off with PBST, and
protein band was visualized by enhanced chemiluminescence method
(EMD Millipore). β-actin was served as loading control for
normalization using AlphaView 3.4.0 software (ProteinSimple).
Cell transfection
ARPE-19 cells were plated into 6-well plates and
incubated for 24 h before transfection. Cells were transfected with
100 nM inhibitor control (5′-GCCUCCGGCUUCGCACCUCU-3′; Shanghai
GenePharma Co., Ltd., Shanghai, China), 100 nM miR-217 inhibitor
(antagomir; 5′-UACUGCAUCAGGAACUGAUUGGA-3′; Shanghai GenePharma Co.,
Ltd.), 0.2 µM control-small interfering (si)RNA (cat. no. sc-36869;
Santa Cruz Biotechnology, Inc.), 0.2 µM SIRT1-siRNA (cat. no.
sc-40986; Santa Cruz Biotechnology, Inc.), or 100 nM miR-217
inhibitor + 0.2 µM SIRT1-siRNA using Lipofectamine 2000 reagent for
24 h. Then, we performed RT-qPCR assay to detect the transfection
efficiency.
MTT assay
ARPE-19 cells were first transfected with inhibitor
control, miR-217 inhibitor, or miR-217 inhibitor + SIRT1-siRNA for
6 h, and then the cells were treated with 50 mM D-glucose at 37°C
for 24 h, and cell viability was determined by using MTT assay. In
brief, 5 mg/ml MTT (Sigma-Aldrich; Merck KGaA) was added to cell
culture medium and incubated for 4 h at 37°C. Subsequently,
dimethyl sulfoxide was added to each well to dissolve the formazan
crystals. The absorbance was measured at 490 nm using a Synergy™ 2
Multi-function Microplate Reader (Bio-Tek Instruments). Experiments
were repeated at least three times.
Flow cytometry assay
We used specific a Annexin V-fluorescein
isothiocyanate (FITC) Apoptosis Detection kit I (BD Bioscience) to
detect cell apoptosis according to the manufacturer's introduction.
In brief, cells were washed twice with 1X PBS. Then, cells
(1×106) were collected, centrifuged (1,000 × g, 5 min,
4°C), and re-suspended in 100 µl of FITC-binding buffer.
Subsequently, the buffer was added with ~5 µl ready-to-use annexin
V-FITC (BD Bioscience) and 5 µl propidium iodide (PI). In the dark,
cells were incubated for 30 min at room temperature. Annexin V-FITC
and PI fluorescence was assessed by BD FACSCalibur flow cytometer
(BD Technologies) with FlowJo software (version 7.6.1; FlowJo LLC).
The apoptotic rate (early apoptotic rate + late apoptotic rate) was
calculated and presented.
ELISA
After treatment, cell supernatant was collected
through centrifugation (500 × g, 5 min, 4°C). The levels of (IL)-1β
(cat. no. PT305; Beyotime Institute of Biotechnology), tumor
necrosis factor α (TNF-α; cat. no. PT518; Beyotime Institute of
Biotechnology), and IL-6 (cat. no. PT330; Beyotime Institute of
Biotechnology) in the supernatant (100 µl) of ARPE-19 cells were
detected using sandwich ELISA kits from according to the
manufacturer's protocol of each kit.
Statistical analysis
All experiments were carried out three times. Data
were displayed from three independent experiments in triplicate. We
conducted analyses with a Student's t-test or one-way ANOVA
followed by Tukey's test for biostatistical analysis. Analyses were
performed using SPSS 19.0 statistical software (IMB Corp.). The
data were presented as the mean ± SD, and P<0.05 was considered
to indicate a statistically significant difference.
Results
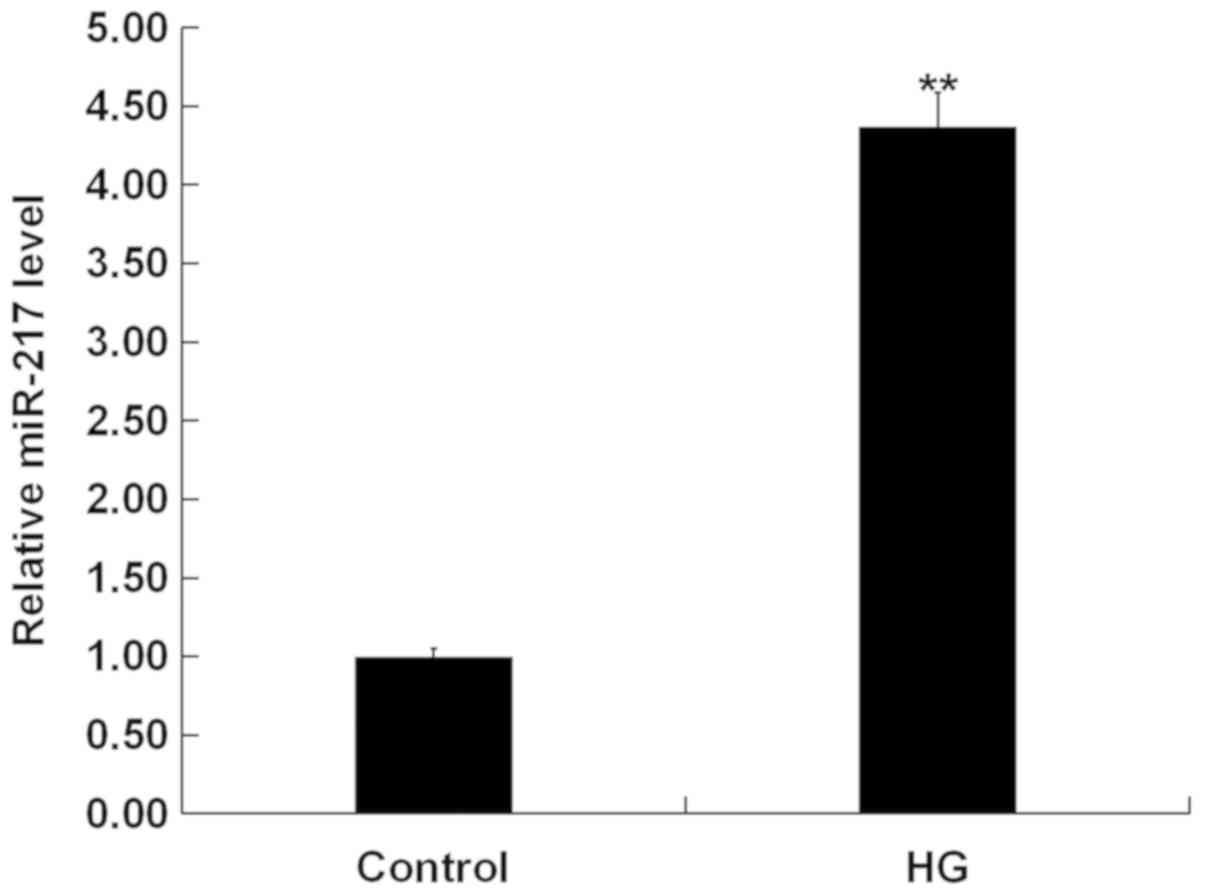
Expression of miR-217 in HG-induced
ARPE-19 cells
To explore the role of miR-271 in HG-induced ARPE-19
cells, we firstly performed RT-qPCR assay to detect the expression
of miR-217 in HG-induced ARPE-19 cells. RT-qPCR assay indicated
that compared with the control group, the expression of miR-217 was
significantly increased in the HG treatment group (Fig. 1).
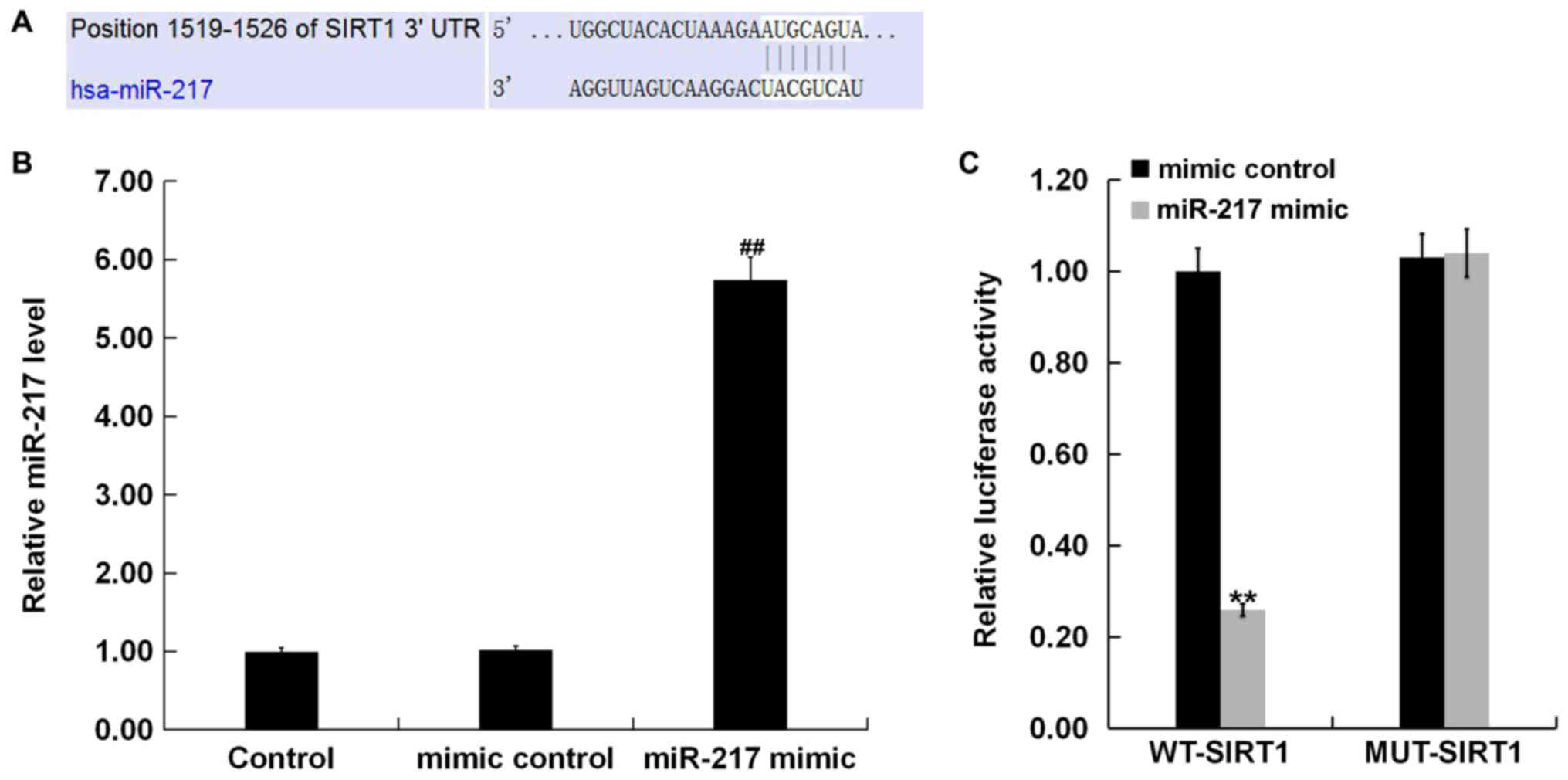
SIRT1 is the direct target gene of
miR-217
To further investigate the molecular mechanism of
miR-217 in HG-induced ARPE-19 cells, we performed TargetScan to
predict the target gene of miR-217. The results showed the binding
sites between 3′-UTR of SIRT1 and miR-217 (Fig. 2A). Next, to verify the binding
sites between miR-217 and SIRT1, we performed a dual-luciferase
reporter assay. First, we confirmed that miR-217 mimic
significantly enhanced the level of miR-217 in ARPE-19 cells
(Fig. 2B). The dual-luciferase
reporter assay showed that miR-217 mimic inhibited the luciferase
activity of ARPE-19 cells co-transfected with the SIRT1-WT and
miR-217 mimic. However, no significant change was observed in the
luciferase activity of ARPE-19 cells co-transfected with the
SIRT1-MUT and miR-217 mimic (Fig.
2C). Taken together, SIRT1 was suggested to be a direct target
gene of miR-217.
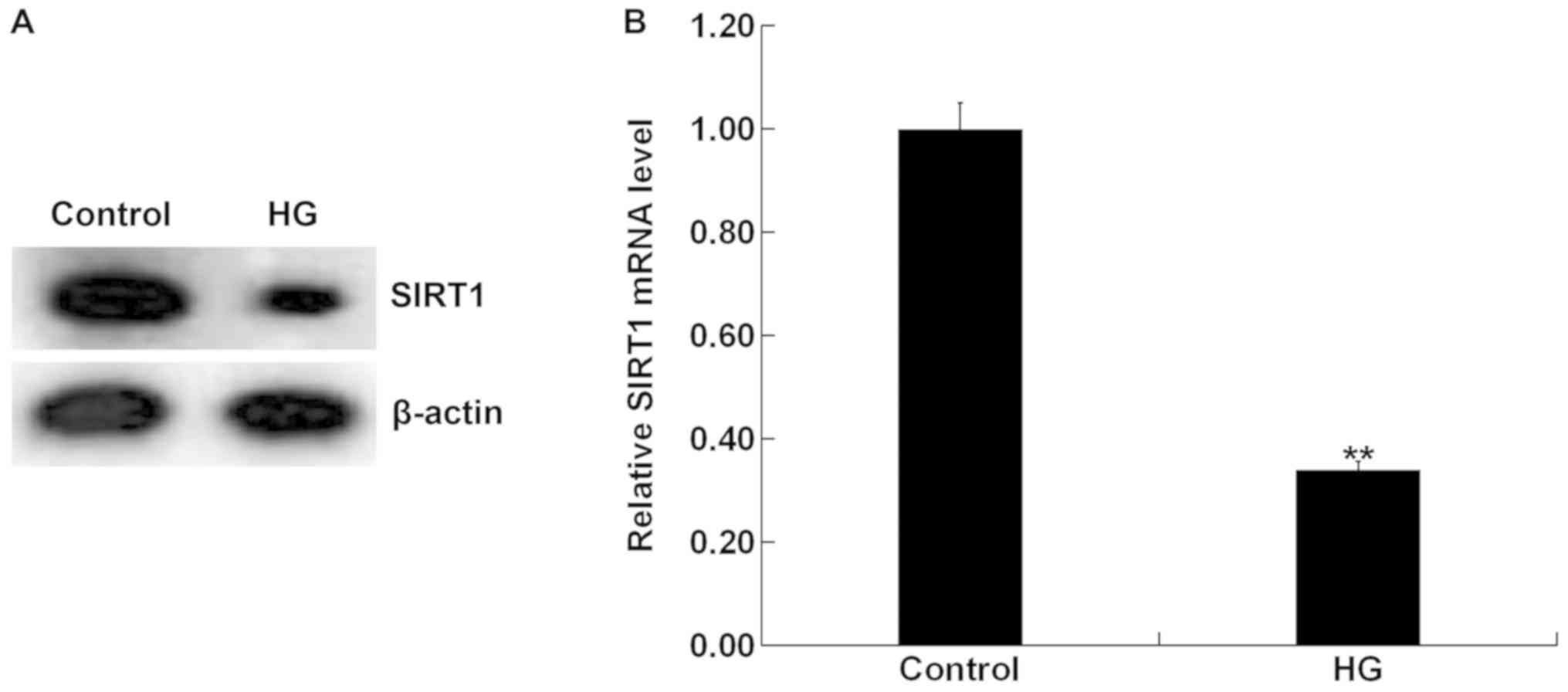
Expression of SIRT1 in HG-induced
ARPE-19 cells
To detect the expression of SIRT1 in HG-induced
ARPE-19 cells, we performed western blotting and RT-qPCR. Western
blotting indicated that the expression of SIRT1 was notably
decreased at the protein level in the HG induced-ARPE-19 cells
(Fig. 3A). RT-qPCR revealed that
compared with the control group, SIRT1 expression was significantly
decreased at mRNA level in the HG-induced ARPE-19 cells (Fig. 3B).
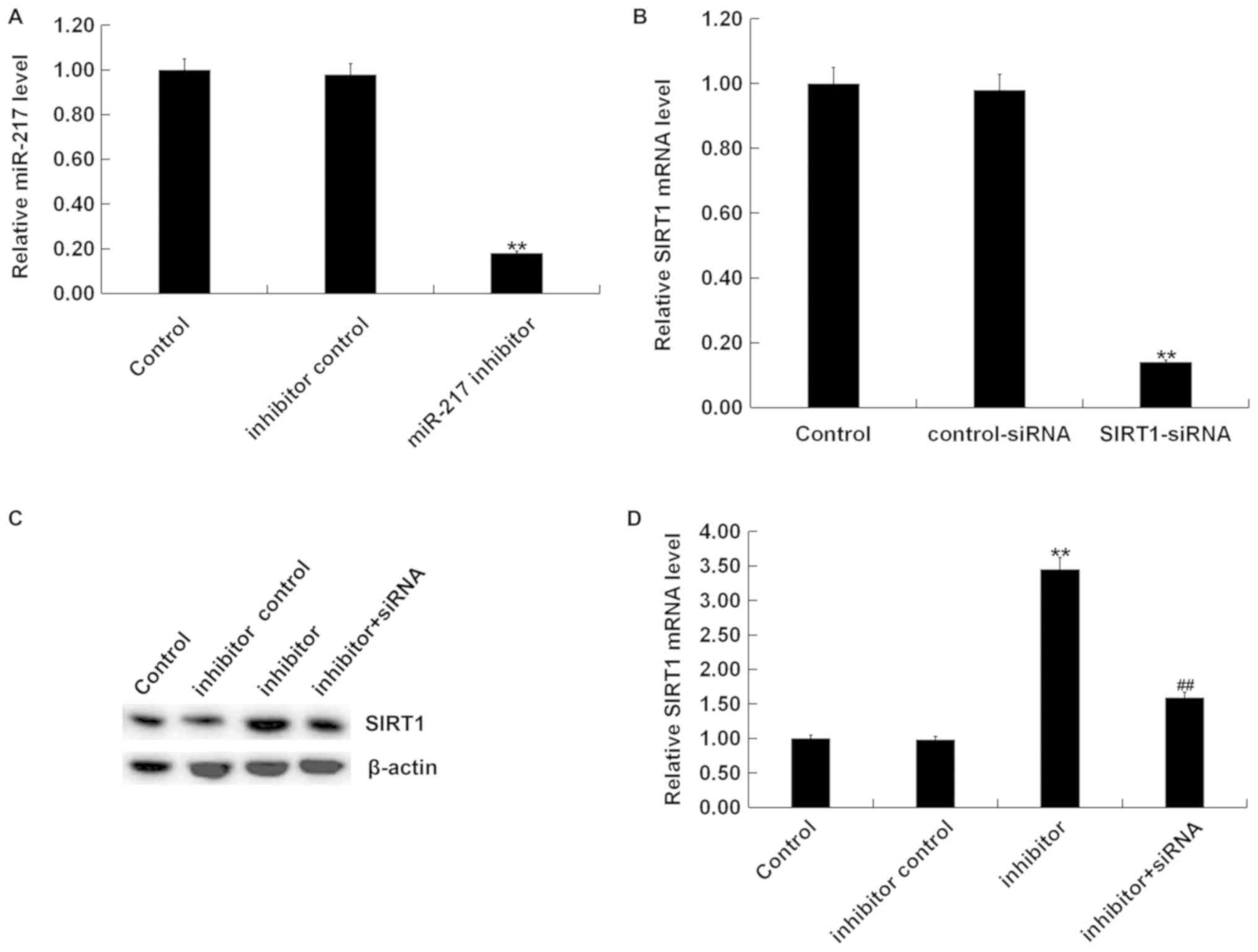
miR-217 negatively regulates SIRT1
expression in ARPE-19 cells
ARPE-19 cells were transfected with inhibitor
control, miR-217 inhibitor, control-siRNA, SIRT1-siRNA, or miR-217
inhibitor + SIRT1-siRNA using Lipofectamine 2000 for 24 h. We
performed RT-qPCR to detect the transfection efficiency. RT-qPCR
assay indicated that miR-217 inhibitor significantly reduced the
expression of miR-217 in ARPE-19 cells compared with the control
(Fig. 4A). SIRT1-siRNA
significantly decreased SIRT1 mRNA expression in ARPE-19 cells
compared with the control (Fig.
4B). miR-217 inhibitor significantly increased the expression
of SIRT1 in ARPE-19 cells, but this increase was abolished by
SIRT1-siRNA (Fig. 4C and D). These
results indicated that SIRT1 was negatively regulated by miR-217 in
ARPE-19 cells.
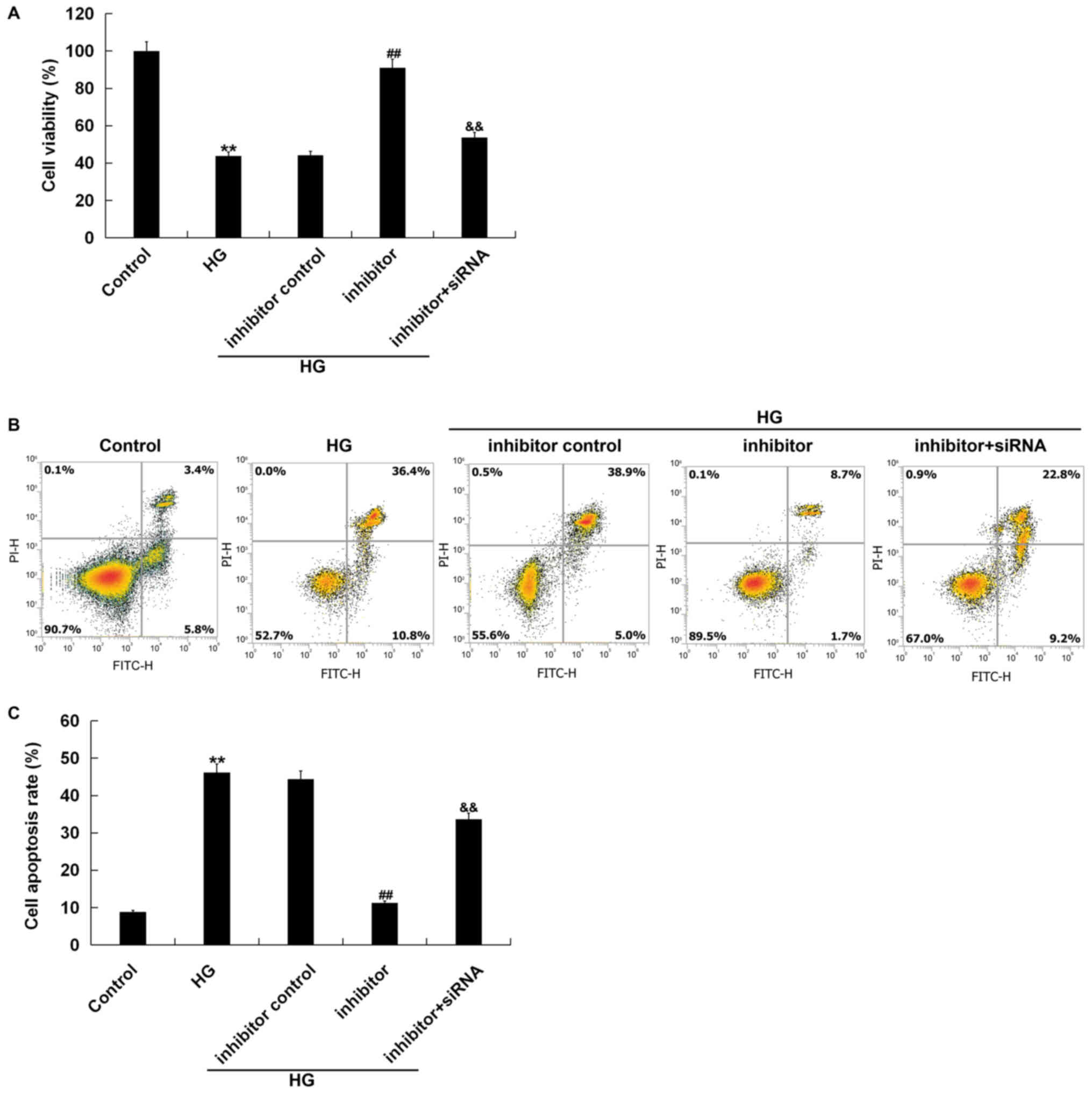
Effects of miR-217 inhibitor on the
viability and apoptosis of ARPE-19 cells treated with HG
To explore the effects of miR-217 on HG treated
ARPE-19 cells, we performed an MTT assay and flow cytometry. The
MTT assay indicated that compared with the control group, the cell
viability was significantly reduced in the HG group; compared with
the HG group, miR-217 inhibitor significantly increased the
viability of ARPE-19 cells, which was significantly reversed by the
silencing of the SIRT1 gene (Fig.
5A). Flow cytometry assay revealed that compared with the
control group, cell apoptosis was significantly increased in the HG
group; compared with the HG group, miR-217 inhibitor significantly
decreased the apoptosis of ARPE-19 cells, which was significantly
reversed by the silencing of the SIRT1 gene (Fig. 5B and C). Taken together, miR-217
inhibitor could enhance cell viability and reduce cell apoptosis in
HG-treated ARPE-19 cells.
Effects of miR-217 inhibitor on the
expression of inflammatory cytokines in HG-induced ARPE-19
cells
In order to determine the expression of inflammatory
cytokines, we performed ELISA to detect the expression of IL-1β,
TNF-α and IL-6. ELISA showed that compared with the control group,
the expression of IL-1β, TNF-α and IL-6 was significantly increased
in the HG group. Compared with the HG group, miR-217 inhibitor
decreased the expression of inflammatory factors in ARPE-19 cells.
This reduction was significantly reversed by the silencing of the
SIRT1 gene (Fig. 6).
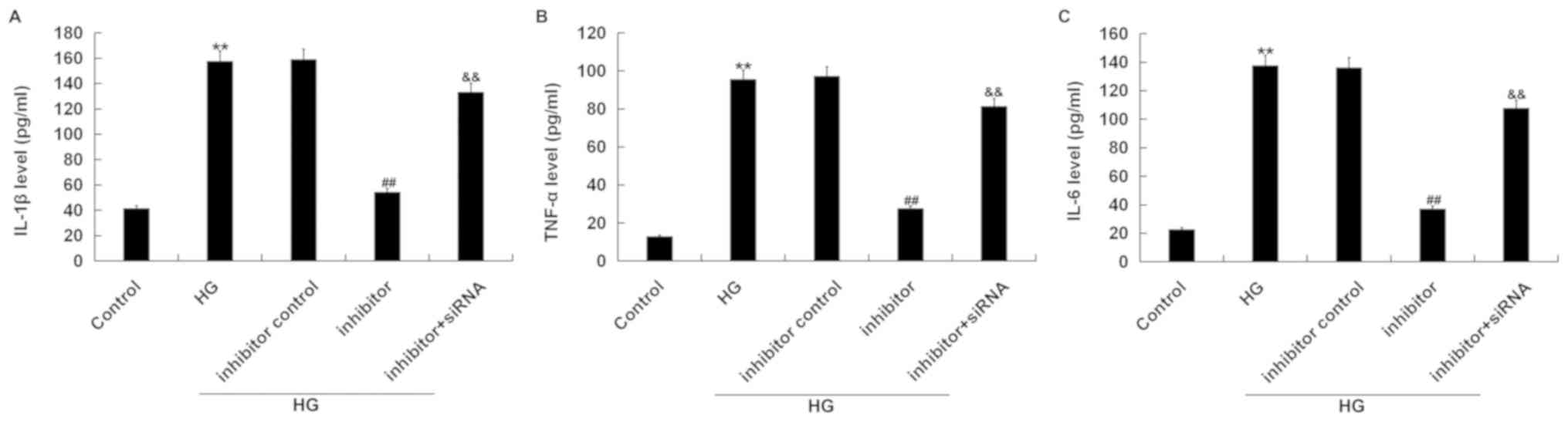 | Figure 6.miR-217 inhibitor decreases the
expression of IL-1β, TNF-α and IL-6 in HG induced ARPE-19 cells.
ARPE-19 cells were transfected with inhibitor control, miR-217
inhibitor, or miR-217 inhibitor + SIRT1-siRNA for 6 h, then the
cells were treated with 50 mM D-glucose for 24 h. Then, ELISA assay
was used to measure the expression of (A) IL-1β, (B) TNF-α and (C)
IL-6. Experiments were repeated three times. The data were shown as
the mean ± SD. **P<0.01 vs. Control; ##P<0.01 vs.
HG; &&P<0.01 vs. inhibitor. HG, high glucose;
IL, interleukin; siRNA, small interfering RNA; SIRT1, Sirtuin 1;
TNF-α, tumor necrosis factor-α. |
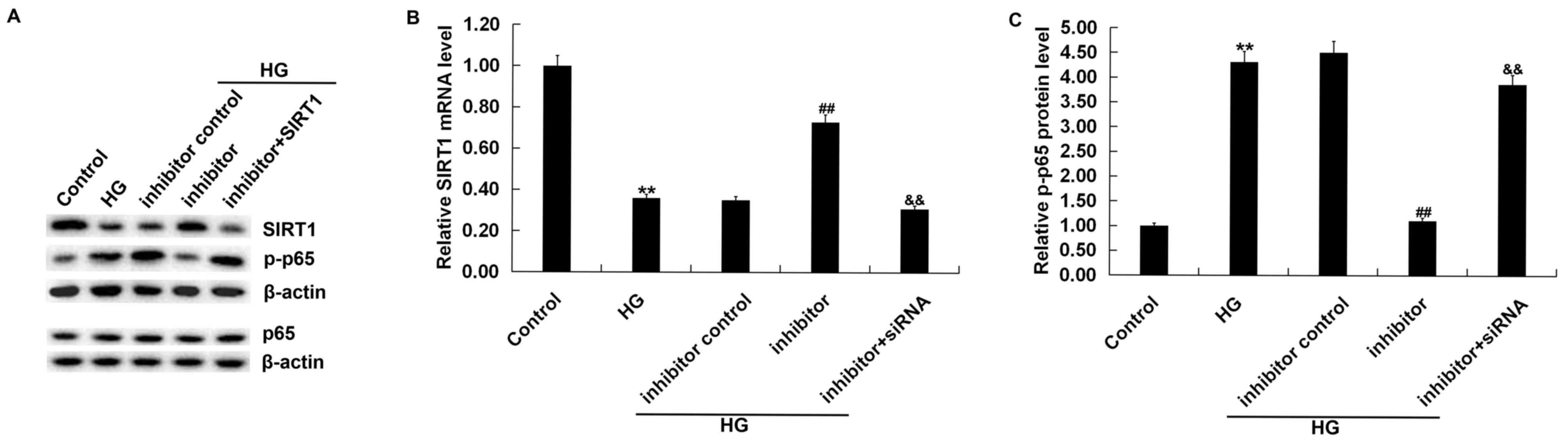
Effects of miR-217 on the expression
of SIRT1 and p-p65 in HG-induced ARPE-19 cells
In order to explore the specific mechanism of
miR-217 in HG-induced ARPE-19 cells, we performed western blotting
to detect the expression of SIRT1 and p-p65. The results showed
that compared with the control group, the protein expression of
SIRT1 was decreased and p-p65 expression was significantly
increased in the HG group. Compared with the HG group, miR-217
inhibitor increased the protein expression of SIRT1 (Fig. 7A) and significantly decreased p-p65
expression (Fig. 7A and C) in
ARPE-19 cells; this change was significantly reversed by the
silencing of the SIRT1 gene. Moreover, the data indicated that
HG-induced downregulation of SIRT1 mRNA in ARPE-19 cells was
significantly reversed by miR-217 inhibitor, and this upregulation
was significantly abolished by SIRT1-siRNA (Fig. 7B).
Discussion
Diabetic retinopathy is a major complication of
diabetes, and retinal pigment epithelial cell damage is involved in
the development and progression of diabetic retinopathy (1,10–13).
It has been reported that chronic hyperglycemia can induce retinal
vascular endothelial cell death (4). Leal et al (33) researched that high glucose was
found to reduce the rat retinal endothelial cells viability that
were exposed to 30 mM glucose (HG) for 7 days (long-term exposure).
Fan et al (34) reported
that rat retinal capillary endothelial cells stimulated with 30 mM
glucose for 48 and 72 h exhibited significantly reduced cell
viability. In this study, we found that the expression of miR-217
was significantly increased in the HG-induced ARPE-19 cells.
At present, several studies have demonstrated that
miRNAs, as bio-markers and pharmacological targets, play an
important role in ocular diseases, including diabetic retinopathy
(35–38). For example, it has been revealed
that miR-9, miR-23a, miR-27a, miR-34a, miR-146a, and miR-155
represent potential bio-markers, and novel pharmacological targets
for age related macular degeneration (36). miR-383 was highly expressed in
HG-induced ARPE-19 cells, and overexpression of miR-383 inhibited
cell viability and promoted apoptosis and reactive oxygen species
formation in ARPE-19 cells; thus, targeting miR-383 may have
therapeutic potential in the treatment of diabetic retinopathy
(39). miR-145 decreased
HG-induced oxidative stress and inflammation in retinal endothelial
cells by modulating the Toll-like receptor 4/nuclear factor-κB
signaling (40). miR-217, an
important miRNA, serves as a tumor suppressor or an oncogene that
depended on the cancer type. For example, Zhao et al
(41) showed that miR-217
regulated either V-Ki-Ras2 KRAS or SIRT1 in pancreatic cancer. It
has been reported that miRNA-217 acts as a anti-oncogene and is
related to the drug-resistance of lung cancer (27). Yin et al (26) revealed that miR-217 was
downregulated in clinical cervical cancer tissues. A recent study
indicated that miR-217 was upregulated in the hearts of CHF
patients (42). However, the
expression and mechanism of miR-217 in HG-induced retinal
epithelial cells remains unclear. In this study, we investigated
the role of miR-217 in HG-induced ARPE-19 cells.
To explore the specific mechanism underlying the
role of miR-217 in HG-induced ARPE-19 cells, we researched the
target gene of miR-217. TargetScan and a dual-luciferase reporter
gene assay suggested that SIRT1 was the direct target gene of
miR-217. SIRT1 is a cellular histone deacetylase that is highly
expressed in many cancers (43).
Lu et al (44) reported
that SIRT1 was the target gene of miR-138. Sun et al
(45) indicated that SIRT1 was the
target gene of miR-29b and miR-29b reversibly modulated the
expression of SIRT1. Borji et al (46) showed that SIRT1 was target gene of
miR-23b and miR-23b mimic suppressed the expression of SIRT1. A
recent study indicated that SIRT1 regulates apoptosis, inflammation
and oxidative stress, alleviating diabetic retinopathy (47). Improving SIRT1 inhibition is a
potentially effective strategy for treating diabetic retinopathy
(48). In our study, we found that
the expression of SIRT1 was downregulated at the mRNA and protein
levels in HG-induced ARPE-19 cells. Furthermore, we reported that
miR-217 negatively regulated SIRT1 expression in ARPE-19 cells. The
data suggested that miR-217 might serve a role in HG-induced
ARPE-19 cells by regulating the expression of SIRT1.
Additionally, we investigated the effects of miR-217
on HG-treated ARPE-19 cell viability, apoptosis, and inflammatory
factors release. ARPE-19 cells were first transfected with
inhibitor control, miR-217 inhibitor, or miR-217 inhibitor +
SIRT1-siRNA for 6 h, and then the cells were treated with 50 mM
D-glucose for 24 h. Our data demonstrated that miR-217 inhibitor
promoted cell viability and suppressed cell apoptosis. However,
only ARPE-19 cell viability at 24 h was investigated after HG
treatment, which poses as a limitation of our study. Thus, the cell
viability at different time points (such as 24, 48 and 72 h) after
HG treatment should be determined to verify our findings. In
addition, miR-217 inhibitor could significantly reduce the
expression of inflammatory factors, such as IL-1β, TNF-α and IL-6
in ARPE-19 cells. Studies have confirmed that the NF-κB signaling
pathway was activated during diabetic retinopathy (49–51).
Thus, in this study, we explored the whether miR-217 could affect
the activation of NF-κB signaling pathway through measuring the
protein levels of p-p65 (52). The
results indicated that miR-217 inhibitor inhibited the expression
of p-p65, suggesting suppression of the NF-κB signaling pathway. Of
note, the effects of miR-217 inhibitor on HG-treated ARPE-19 cells
were significantly reversed by SIRT1 silencing. These results
indicated that miR-217 regulated the expression of inflammatory
cytokines and NF-κB activity in HG-treated ARPE-19 cells by
regulating the expression of SIRT1.
In conclusion, miR-217 inhibitor protected against
retinal epithelial cell damage caused by HG by targeting SIRT1,
thereby playing a protective role in diabetic retinopathy. However,
this study was only a preliminary study of the role of miR-217 in
diabetic retinopathy in vitro. There were some limitations
of the present study, for example, an in vivo study into the
role of miR-217 in diabetic retinopathy was not performed. In
addition, the expression of miR-217 in patients with diabetic
retinopathy was not determined. Moreover, the relationship between
miR-217 expression and the clinical features of patients with
diabetic retinopathy was not explored. We will further investigate
these issues in the future.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The analyzed datasets generated during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
HX designed the current study, collected and
analyzed the data, performed statistical analysis, and prepared the
manuscript. ZL contributed to data collection and data
interpretation, and searched the literature. Both authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Stewart MW: Treatment of diabetic
retinopathy: Recent advances and unresolved challenges. World J
Diabetes. 7:333–341. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Klein BE: Overview of epidemiologic
studies of diabetic retinopathy. Ophthalmic Epidemiol. 14:179–183.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yau JW, Rogers SL, Kawasaki R, Lamoureux
EL, Kowalski JW, Bek T, Chen SJ, Dekker JM, Fletcher A, Grauslund
J, et al: Global prevalence and major risk factors of diabetic
retinopathy. Diabetes Care. 35:556–564. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang CF, Yuan JR, Qin D, Gu JF, Zhao BJ,
Zhang L, Zhao D, Chen J, Hou XF, Yang N, et al: Protection of
tauroursodeoxycholic acid on high glucose-induced human retinal
microvascular endothelial cells dysfunction and
streptozotocin-induced diabetic retinopathy rats. J Ethnopharmacol.
185:162–170. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Siasos G, Gouliopoulos N, Moschos MM,
Oikonomou E, Kollia C, Konsola T, Athanasiou D, Siasou G, Mourouzis
K, Zisimos K, et al: Role of endothelial dysfunction and arterial
stiffness in the development of diabetic retinopathy. Diabetes
Care. 38:e9–e10. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Frank RN: Diabetic retinopathy. N Engl J
Med. 350:48–58. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mizutani M, Kern TS and Lorenzi M:
Accelerated death of retinal microvascular cells in human and
experimental diabetic retinopathy. J Clin Invest. 97:2883–2890.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Schroder K, Zhou R and Tschopp J: The
NLRP3 inflammasome: A sensor for metabolic danger? Science.
327:296–300. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Roy S, Kern TS, Song B and Stuebe C:
Mechanistic insights into pathological changes in the diabetic
retina: Implications for targeting diabetic retinopathy. Am J
Pathol. 187:9–19. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Curtis TM, Gardiner TA and Stitt AW:
Microvascular lesions of diabetic retinopathy: Clues towards
understanding pathogenesis? Eye (Lond). 23:1496–1508. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Spijkerman AM, Gall MA, Tarnow L, Twisk
JW, Lauritzen E, Lund-Andersen H, Emeis J, Parving HH and Stehouwer
CD: Endothelial dysfunction and low-grade inflammation and the
progression of retinopathy in type 2 diabetes. Diabet Med.
24:969–976. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shin ES, Sorenson CM and Sheibani N:
Diabetes and retinal vascular dysfunction. J Ophthalmic Vis Res.
9:362–373. 2014.PubMed/NCBI
|
|
13
|
Toma L, Stancu CS, Botez GM, Sima AV and
Simionescu M: Irreversibly glycated LDL induce oxidative and
inflammatory state in human endothelial cells; added effect of high
glucose. Biochem Biophys Res Commun. 390:877–882. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cunha-Vaz J, Bernardes R and Lobo C:
Blood-retinal barrier. Eur J Ophthalmol. 21 (Suppl 6):S3–S9. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lorenzi M and Gerhardinger C: Early
cellular and molecular changes induced by diabetes in the retina.
Diabetologia. 44:791–804. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang P, Zhang Z and Kador PF: Polyol
effects on growth factors and MAPK signaling in rat retinal
capillary cells. J Ocul Pharmacol Ther. 30:4–11. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Poy MN, Eliasson L, Krutzfeldt J, Kuwajima
S, Ma X, Macdonald PE, Pfeffer S, Tuschl T, Rajewsky N, Rorsman P
and Stoffel M: A pancreatic islet-specifc microRNA regulates
insulin secretion. Nature. 432:226–230. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang Q, Liu N, Yang X, Tu L and Zhang X:
Small RNA-mediated responses to low- and high-temperature stresses
in cotton. Sci Rep. 6:355582016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lim LP, Lau NC, Garrett-Engele P, Grimson
A, Schelter JM, Castle J, Bartel DP, Linsley PS and Johnson JM:
Microarray analysis shows that some microRNAs downregulate large
numbers of target mRNAs. Nature. 433:769–773. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mendell J and Olson E: MicroRNAs in stress
signaling and human disease. Cell. 148:1172–1187. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wei R, Deng Z and Su J: miR-217 targeting
Wnt5a in osteosarcoma functions as a potential tumor suppressor.
Biomed. Pharmacother. 72:158–164. 2015. View Article : Google Scholar
|
|
23
|
Yin H, Liang X, Jogasuria A, Davidson NO
and You M: miR-217 regulates ethanolinduced hepatic inflammation by
disrupting sirtuin 1-lipin-1 signaling. Am J Pathol. 185:1286–1296.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Azam AT, Bahador R, Hesarikia H, Shakeri M
and Yeganeh A: Downregulation of microRNA-217 and microRNA-646 acts
as potential predictor biomarkers in progression, metastasis, and
unfavorable prognosis of human osteosarcoma. Tumour Biol.
37:5769–5773. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Popov A, Szabo A and Mandys V: Small
nucleolar RNA U91 is a new internal control for accurate microRNAs
quantifcation in pancreatic cancer. BMC Cancer. 15:7742015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yin Z and Ren W: MicroRNA-217 acts as a
tumor suppressor and correlates with the chemoresistance of
cervical carcinoma to cisplatin. OncoTargets and Terapy.
12:759–771. 2019. View Article : Google Scholar
|
|
27
|
Guo J, Feng Z, Huang Z, Wang H and Lu W:
MicroRNA-217 functions as a tumour suppressor gene and correlates
with cell resistance to cisplatin in lung cancer. Mol Cells.
37:664–671. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lin CJ, Lan YM, Ou MQ, Ji LQ and Lin SD:
Expression of miR-217 and HIF-1α/VEGF pathway in patients with
diabetic foot ulcer and its effect on angiogenesis of diabetic foot
ulcer rats. J Endocrinol Invest. May 11–2019.doi:
10.1007/s40618-019-01053-2 (Epub ahead of print). View Article : Google Scholar
|
|
29
|
Sun J, Li ZP, Zhang RQ and Zhang HM:
Repression of miR-217 protects against high glucose-induced
podocyte injury and insulin resistance by restoring PTEN-mediated
autophagy pathway. Biochem Biophys Res Commun. 483:318–324. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shao Y, Lv C, Wu C, Zhou Y and Wang Q:
Mir-217 promotes inflammation and fibrosis in high glucose cultured
rat glomerular mesangial cells via Sirt1/HIF-1α signaling pathway.
Diabetes Metab Res Rev. 32:534–543. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shao Y, Ren H, Lv C, Ma X, Wu C and Wang
Q: Changes of serum Mir-217 and the correlation with the severity
in type 2 diabetes patients with different stages of diabetic
kidney disease. Endocrine. 55:130–138. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Leal EC, Aveleira CA, Castilho AF, Serra
AM, Baptista FI, Hosoya K, Forrester JV and Ambrósio AF: High
glucose and oxidative/nitrosative stress conditions induce
apoptosis in retinal endothelial cells by a caspase-independent
pathway. Exp Eye Res. 88:983–991. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Fan Y, Qiao Y, Huang J and Tang M:
Protective effects of panax notoginseng saponins against high
glucose-induced oxidativeinjury in rat retinal capillary
endothelial cells. Evid Based Complement Alternat Med.
2016:53263822016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Rassi DM, De Paiva CS, Dias LC, Módulo CM,
Adriano L, Fantucci MZ and Rocha EM: Review: MicroRNAS in ocular
surface and dry eye diseases. Ocul Surf. 15:660–669. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Romano GL, Platania CBM, Drago F, Salomone
S, Ragusa M, Barbagallo C, Di Pietro C, Purrello M, Reibaldi M,
Avitabile T, et al: Retinal and circulating miRNAs in age-related
macular degeneration: An in vivo animal and human study. Front
Pharmacol. 8:1682017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ma J, Wang J, Liu Y, Wang C, Duan D, Lu N,
Wang K, Zhang L, Gu K, Chen S, et al: Comparisons of serum miRNA
expression profiles in patients with diabetic retinopathy and type
2 diabetes mellitus. Clinics (Sao Paulo). 72:111–115. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gong Q and Su G: Roles of miRNAs and long
noncoding RNAs in the progression of diabetic retinopathy. Biosci
Rep. 37:BSR201711572017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jiang Y, Sang Y and Qiu Q: microRNA-383
mediates high glucose-induced oxidative stress and apoptosis in
retinal pigment epithelial cells by repressing peroxiredoxin 3. Am
J Transl Res. 9:2374–2383. 2017.PubMed/NCBI
|
|
40
|
Ying H and Yan Y: MicroRNA-145 attenuates
high glucose-induced oxidative stress and inflammation in retinal
endothelial cells through regulating TLR4/NF-κB signaling. Life
Sci. 207:212–218. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhao WG, Yu SN, Lu ZH, Ma YH, Gu YM and
Chen J: The miR-217 microRNA functions as a potential tumor
suppressor in pancreatic ductal adenocarcinoma by targeting KRAS.
Carcinogenesis. 31:1726–1733. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Li H, Fan J, Yin Z, Wang F, Chen C and
Wang DW: Identification of cardiac-related circulating microRNA
profile in human chronic heart failure. Oncotarget. 7:33–45.
2016.PubMed/NCBI
|
|
43
|
Lian B, Yang D, Liu Y, Shi G, Li J, Yan X,
Jin K, Liu X, Zhao J, Shang W and Zhang R: miR-128 Targets the
SIRT1/ROS/DR5 pathway to sensitize colorectal cancer to
TRAIL-induced apoptosis. Cell Physiol Biochem. 49:2151–2162. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lu Y, Tan L and Wang X: Circular
HDAC9/microRNA-138/Sirtuin-1 pathway mediates synaptic and amyloid
precursor protein processing deficits in Alzheimer's disease.
Neurosci Bull. Mar 18–2019.doi: 10.1007/s12264-019-00361-0 (Epub
ahead of print).
|
|
45
|
Sun QR, Zhang X and Fang K: Phenotype of
vascular smooth muscle cells (VSMCs) is regulated by miR-29b by
targeting Sirtuin 1. Med Sci Monit. 24:6599–6607Yue. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Borji M, Nourbakhsh M, Shafiee SM, Owji
AA, Abdolvahabi Z, Hesari Z, Ilbeigi D, Seiri P and Yousefi Z:
Down-regulation of SIRT1 expression by mir-23b contributes to lipid
accumulation in HepG2 Cells. Biochem Genet. 57:507–521. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Karbasforooshan H and Karimi G: The role
of SIRT1 in diabetic retinopathy. Biomed Pharmacother. 97:190–194.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Mishra M, Duraisamy AJ and Kowluru RA:
Sirt1: A guardian of the development of diabetic retinopathy.
Diabetes. 67:745–754. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wang W, Zhang Y, Jin W, Xing Y and Yang A:
Catechin weakens diabetic retinopathy by inhibiting the expression
of NF-κB signaling pathway-mediated inflammatory factors. Ann Clin
Lab Sci. 48:594–600. 2018.PubMed/NCBI
|
|
50
|
Yin Y, Chen F, Wang W, Wang H and Zhang X:
Resolvin D1 inhibits inflammatory response in STZ-induced diabetic
retinopathy rats: Possible involvement of NLRP3 inflammasome and
NF-κB signaling pathway. Mol Vis. 23:242–250. 2017.PubMed/NCBI
|
|
51
|
Kim SJ, Yoo WS, Choi M, Chung I, Yoo JM
and Choi W: Increased O-GlcNAcylation of NF-κB enhances retinal
ganglion cell death in streptozotocin-induced diabetic retinopathy.
Curr Eye Res. 41:249–257. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Tey SK, Tse EYT, Mao X, Ko FCF, Wong AST,
Lo RC, Ng IO and Yam JWP: Nuclear Met promotes hepatocellular
carcinoma tumorigenesis and metastasis by upregulation of TAK1 and
activation of NF-κB pathway. Cancer Lett. 411:150–161. 2017.
View Article : Google Scholar : PubMed/NCBI
|