In the 1960s, two independent groups discovered
several soluble factors when measuring mouse lymphoid leukemia cell
growth (1,2) and these soluble factors were named
‘colony-stimulating factors’ (CSFs). After isolation and
purification, four CSFs were identified, including macrophage CSF
(M-CSF, CSF1) (3),
granulocyte-macrophage CSF (GM-CSF, CSF2) (4), granulocyte CSF (G-CSF, CSF3)
(5) and multipotential CSF (also
known as interleukin-3) (6). All
these factors are essential stimulators of blood cell development
and play a crucial role in hematopoietic stem cell proliferation
and differentiation at different stages (7).
The biological effects of G-CSF are mediated by the
specific G-CSF receptor (G-CSFR) (8). G-CSF is a critical regulator of
neutrophil production and activity. It promotes proliferation and
differentiation of the neutrophil lineage, and enhances the
transition of immature metamyelocytes into mature neutrophils.
G-CSF not only prolongs the survival of neutrophils and their
precursors, but also promotes the functions of mature neutrophils,
such as superoxide production, phagocytosis and pathogen killing
(9).
G-CSF can act as a mobilizer of hematopoietic
progenitor stem cells in blood donors or cancer patients (10). Therefore, recombinant human G-CSF
(rhG-CSF) is commonly used to prevent and treat febrile neutropenia
and mucositis after chemotherapy and radiotherapy for cancer
patients (11). However, recent
studies have found that G-CSF plays a crucial role in
tumorigenesis. G-CSF promotes tumor growth, metastasis and
chemotherapy resistance (12),
inhibits tumor cell apoptosis, induces angiogenesis (13,14),
participates in cancer-associated thrombosis (15,16),
and is associated with a poor clinical prognosis (17).
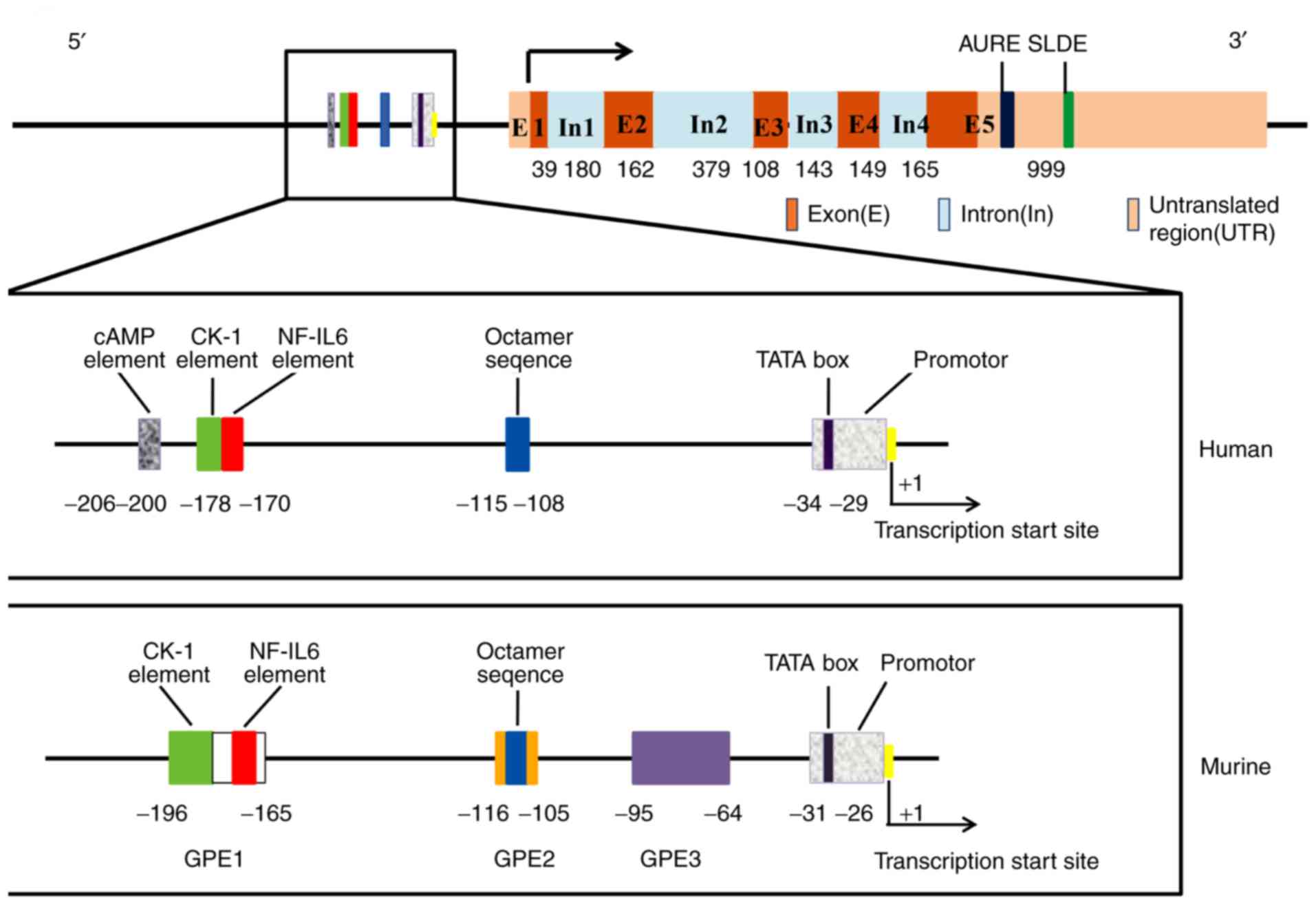
It has been acknowledged that there are two
different G-CSF mRNA isoforms in humans: G-CSFa and G-CSFb.
Compared with G-CSFa, G-CSFb lacks 9 base pairs (GTGAGTGAG) in the
second exon (21). G-CSFa and
G-CSFb mRNAs encode polypeptides that consist of 207 and 204 amino
acids, respectively. After cleavage of the 30-amino acid signal
peptide, mature proteins containing 177 and 174 amino acids are
secreted. Arakawa et al (33), found that the activity of the
174-amino acid form is 50-fold higher than that of the 177-amino
acid form. The secreted form of the protein was found to be
O-glycosylated and to have a molecular weight of 19,600 Da
(34). One O-linked glycosyl group
at Thr 133 in G-CSF isolated from human blood protects the molecule
from aggregation (35).
The G-CSF protein contains five cysteines and two
pairs of disulfide bonds are formed between residues Cys36 and
Cys42 and residues Cys74 and Cys64. The disulfide bonds play an
important role in maintaining the biological functions of G-CSF.
Within the G-CSF protein, 104 of the 175 residues form a total of
four α-helix bundles that are designated helix A (residues 11–39),
B (71–91), C (100–123) and D (143–172) (36). A study of the three-dimensional
crystal structure of recombinant interferon (IFN)-β suggested that
the receptor binding region of G-CSF is located on the loop
connecting helix A and B and on the outer surface of helix D
(37).
Under physiological conditions, the G-CSF
concentration in plasma is almost undetectable, but when an
infection occurs, the G-CSF concentration is significantly
increased. The number of neutrophils is dependent on the G-CSF
concentration, especially during the infection process or
chemotherapy use (38). G-CSF can
be secreted by numerous cells, including monocytes, macrophages,
endothelial cells, epithelial cells and fibroblasts, when they are
stimulated by inflammatory mediators such as LPS (39), IL-17 (40), TNF-α and IFN-β (41). Moreover, some malignant cells, such
as triple-negative breast cancer (17), lung carcinoma (42,43),
bladder cancer (44) and squamous
cell carcinoma (45), can
constitutively express and secrete G-CSF.
G-CSF expression in breast cancer is under the
control of various signaling pathways. It has been reported that
carbonic anhydrase IX (CAIX) stimulates G-CSF production by
activating NF-κB signaling in hypoxic conditions (46). Extracellular signal-regulated
kinase (ERK) 2 is responsible for the transcriptional regulation of
G-CSF and ERK2 knockdown by short hairpin RNA significantly
inhibits the expression of tumor-derived G-CSF (47). H-Ras upregulates G-CSF expression
and promotes breast epithelial MCF10A cell invasiveness (48). Protease-activated receptor (PAR) 2
stimulates G-CSF expression in breast cancer and PAR2 gene
knockdown or PAR2 antagonist use can reduce G-CSF secretion
(49). Carcinoembryonic
antigen-related cell adhesion molecule (CEACAM) 1 expression in
breast cancer MCF-7 cells inhibits G-CSF secretion by M1
macrophages (50). In addition,
G-CSF is the main downstream mediator of the mammalian target of
rapamycin (mTOR) pathway during the induction of myeloid-derived
suppressor cell (MDSC) formation in breast cancer and Welte et
al (51), suggested that the
regulation of G-CSF by mTOR may occur at the transcriptional level.
In other diseases, some factors have been shown to regulate G-CSF
expression, all of which are shown in Table I.
The G-CSFR gene located on chromosome 1p35-34.3 is a
member of the class I cytokine receptor superfamily (52). G-CSFR is a single transmembrane
protein consisting of 813 amino acid residues, which is composed of
extracellular, transmembrane and intracellular regions. Its
extracellular region includes immunoglobulin-like (Ig-like) domains
and cytokine receptor homology (CRH) domains, as well as three
fibronectin type III domains. The intracellular region of G-CSFR
protein includes two motifs called box 1 and box 2, and cytoplasmic
region of G-CSFR contains four conserved tyrosine residues which
function as docking sites for the phosphorylation of multiple
SH2-containing signaling proteins (53). G-CSF binds to the extracellular
Ig-like and CRH domains of G-CSFR, which triggers receptor
homodimerization (54) and
activates Janus tyrosine kinases (JAKs), leading to a
cross-phosphorylation. Activated JAKs proteins can phosphorylate
G-CSFR by binding to its Box 1 and 2 domains and generate potential
docking sites of signal transducer and activator of transcription
(STAT) protein in cytoplasm. The inactive STAT protein binds to the
phosphorylated G-CSFR through its SH2 domain and phosphorylates it
under the cooperation with JAKs (55). Activated STATs then form a
homodimer/heterodimer and translocate into the nucleus to activate
the transcription of target genes, which promote the proliferation
and metastasis of cancer cells (56). Although the G-CSF-induced
JAK2/STAT3 pathway has been well-established (57), previous studies also show that
G-CSF can activate other downstream signaling pathways, including
mitogen-activated protein kinase (MAPK)/ERK and
phosphatidylinositol 3 kinase/protein kinase B (AKT) (54,58).
Some studies have reported that serum G-CSF levels
are significantly higher in breast cancer patients compared with
healthy controls (48,59–63).
Lawicki et al (60),
demonstrated that the plasma levels of G-CSF and M-CSF were
significantly increased in 54 breast cancer patients compared with
in control group patients. The authors of the present review were
surprised to learn that, after surgical resection, the level of
G-CSF decreased significantly, but the level of M-CSF increased,
suggesting that measuring G-CSF may be useful in the diagnosis of
breast cancer. Compared with 20 healthy controls, the mean level of
serum G-CSF in 20 breast cancer patients was significantly
increased (48). In a total of 190
samples, plasma G-CSF levels were significantly increased in
samples from 110 patients with ductal breast cancer and 40 patients
with benign breast cancer compared with samples from untreated
healthy patients. Moreover, the serum levels of G-CSF were
significantly increased in patients with clinical stage III and IV
tumors compared with in healthy controls or patients with benign
breast cancer (61). In addition,
in a total of 196 samples, serum G-CSF levels were increased in
patients with advanced breast cancer compared with in patients with
early-stage cancer and the highest G-CSF levels were observed in
patients with N3 tumors (62). By
analyzing the wound healing fluid of breast cancer surgery
patients, it was found that G-CSF, together with IL-6 and monocyte
chemotactic protein (MCP)-1/CCL2, was more abundant in invasive and
high-grade breast cancer than in situ breast cancer (63).
Cancer-related thrombosis is the second leading
cause of death and is usually associated with poor prognosis in
cancer patients. NET formation is crucial for thrombosis formation
in tumor-bearing mice. 4T1 cell-derived exosomes induce NET
formation in neutrophils from G-CSF-treated mice, which can promote
thrombus formation in tumor-free neutrophilic mice. The results
suggested that tumor-derived exosomes and neutrophils play a
synergistic role in the formation of cancer-associated thrombosis
(70). Demers et al
(15), discovered that
cancer-associated G-CSF exacerbates the innate immune response of
the host which leads to thrombosis. NET formation induces a
pro-thrombotic state which may result in the consumption of
platelets, clotting factors and microthrombosis in rhG-CSF-treated
4T1 mice. IL-1β modulates the expression of G-CSF and the levels of
G-CSF and IL-1β are elevated in 4T1 mice which exhibit a
NET-dependent prothrombotic state. Blocking IL-1R reduces the G-CSF
level, NET formation and abolishes the pre-thrombotic state in 4T1
tumor-bearing mice (16).
rhG-CSF was shown to promote the proliferation of
MCF-7 and SKBR-3 breast cancer cells, but it had little effect on
normal breast epithelial cells. Chronic exposure to low doses of
rhG-CSF (0.125 µg) promotes tumorigenesis in estrogen
receptor-positive breast cancer by promoting the proliferation of
normal and precancerous tissues in MMTV-erbB2 mice (71). Waight et al (72), showed that tumor-derived G-CSF can
directly promote tumor growth and G-CSF knockdown slows tumor
growth in mouse breast tumor models. G-CSF, in combination with
other proinflammatory cytokines such as GM-CSF, IL-8 and MCP-1 that
are secreted by highly aggressive tumor cells, induces an
epithelial mesenchymal transition/stemness-like invasive phenotype
in nonaggressive breast cancer cells (73). Higher G-CSF expression increases
the invasiveness of breast and lung cancer cells, and ERK2
inhibition is necessary to reduce the expression of TNF-α-induced
G-CSF in aggressive cancer cells (47). These results indicate that G-CSF is
a critical factor that promotes breast tumorigenesis and specific
ERK2 inhibitors may be used to treat G-CSF-producing tumors.
G-CSF-induced invasiveness in breast epithelial
MCF10A cells is closely related to H-Ras oncogene upregulation.
Stable expression of G-CSF induced by H-Ras upregulates matrix
metalloproteinase (MMP)-2 expression by activating Rac 1 and
promotes MCF10A cell migration/invasion. MMP-2-mediated degradation
of extracellular matrix components is a key step in the development
of invasiveness. Overexpression of G-CSF in MCF10A cells also
activates other signaling pathways, including MKK3/6, p38 MAPK,
ERK1/2 and AKT (48). The results
of a serological in vivo analysis of breast cancer patients
were also consistent with observations made in vitro,
suggesting that G-CSF may be used as a serum indicator in the
treatment of breast cancer (48).
G-CSF induces ErbB2 expression in breast cancer cell
lines. The present review was surprised that the binding of both
trastuzumab and G-CSF inhibits tumor colony formation and
simultaneously induces apoptosis in these cells. This inhibition is
more pronounced after pretreatment with G-CSF. A total of five of
the nine breast cancer patients showed an increase in their
Herceptest scores, which were used to detect ErbB2 expression after
G-CSF administration (74). The
ErbB2 (HER2) proto-oncogene encodes a tyrosine kinase receptor that
is overexpressed in 15–20% of human breast cancer cases with
aggressive clinical behavior (75).
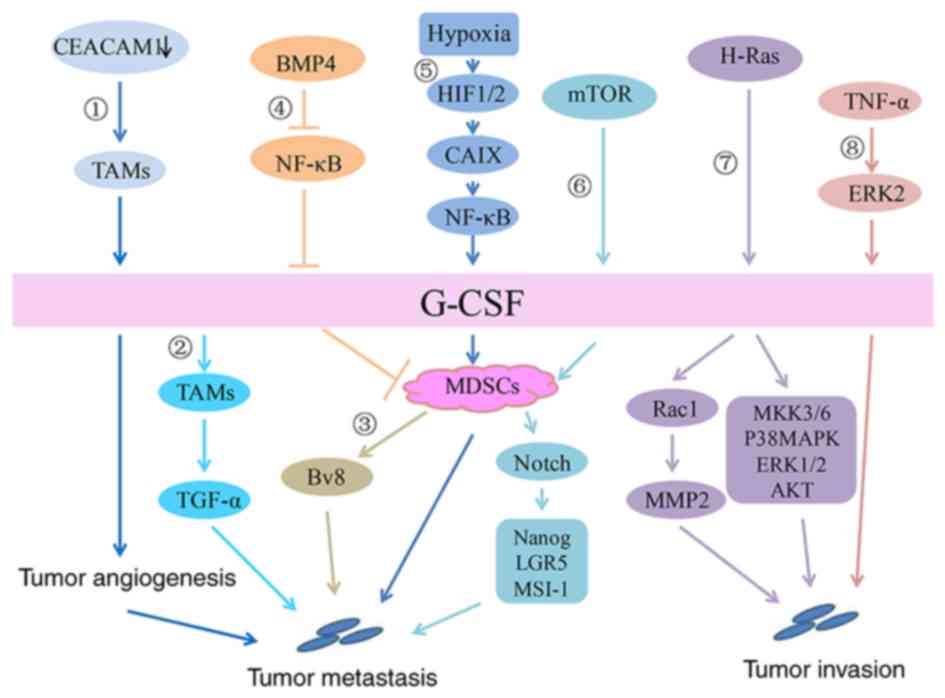
Studies have found that G-CSF promotes MDSC
accumulation in breast cancer via the mTOR signaling pathway.
Reverse-phase protein array analysis in mammary tumor models
revealed that MDSC accumulation is accompanied by increased
AKT-mTOR signaling pathway activity and induces G-CSF expression in
cancer cells. Surprisingly, the expression of G-CSF in
tumor-initiating cells (TICs) is high and MDSCs facilitate the
expression of stemness-related genes in cancer cells, including
Nanog, LGR5 and MSI-1. Moreover, MDSCs stimulate improved TIC
performance via the Notch signaling pathway and TICs promote G-CSF
enhancement and thus increase MDSC accumulation, which therefore
establishes a feed-forward loop between TICs and MDSCs. In
addition, mice treated with the mTOR inhibitor rapamycin showed
significant tumor growth delay. These data demonstrate that the
mTOR/G-CSF/MDSC signaling pathway regulates the malignant
progression of breast tumors (51). Tumor-secreted G-CSF can increase
the number of Ly6G+Ly6C+ granulocytes, which are a subset of
CD11b+Gr1+ cells, in organ-specific transfer sites and further
promote the production of the proangiogenic factor Bv8 protein to
enhance breast tumor metastasis. Anti-G-CSF treatment can
significantly reduce lung metastasis in mammary carcinoma models
(66).
The hypoxic tumor microenvironment is conducive to
driving metastatic niche development (84). Breast tumor cell exposure to a
hypoxic microenvironment results in the activation of
hypoxia-inducible factor (HIF)-1/2-mediated transcriptional
programs that mediate adaptive responses in cells. Chafe et
al (46), first revealed the
relationship between the CAIX-NF-κB-G-CSF cell signaling axis and
breast cancer lung metastasis. In the absence of oxygen, the
expression of CAIX in breast cancer is significantly upregulated
due to HIF-1 activation. NF-κB activation in the microenvironment
is critical for the expression of CAIX, which is required for the
G-CSF-driven mobilization of granulocytic MDSCs to the breast
cancer-derived lung metastatic niche. Constitutive NF-κB activation
can normalize the secretion of G-CSF, even if CAIX is completely
consumed. Mobilized G-CSF-dependent granulocytic MDSCs can enhance
the growth and proliferation of disseminated tumor cells to promote
the formation of lung metastasis via immunosuppression (46).
Tumor-associated macrophages (TAMs) play an
extremely important role in the tumor microenvironment. Macrophages
in the peripheral circulation are recruited to the tumor area as a
result of the action of chemokines and cytokines, such as MCP-1,
M-CSF, CCL8 and vascular epithelial growth factor (VEGF), which are
secreted by tumor cells or the tumor stroma. M-CSF (CSF-1) is a
major contributor to TAM infiltration and promotes tumor growth
(85). Blockade of the
M-CSF/CSF-1R signaling pathway suppresses tumor growth in mammary
carcinoma models (86) and
targeting TAMs with a CSF-1R antibody is a viable strategy for
cancer therapy (87).
CEACAM1 is a cell adhesion molecule that is
downregulated in numerous cancers that originate from the
epithelium (90). CEACAM1 plays a
role in inhibiting inflammation, partly by inhibiting G-CSF
production by myeloid cells. The lack of CEACAM1 expression in
breast tumors promoted the secretion of high levels of G-CSF by
TAMs, which in turn promoted tumor angiogenesis and initial tumor
establishment. It has been suggested that G-CSF plays an important
role in tumor promotion induced by CEACAM1 downregulation (50). Generally, as shown in Fig. 2, G-CSF plays a crucial role in
breast cancer malignant progression.
G-CSF stimulates the proliferation and survival of
hematopoietic stem progenitor cells and their differentiation into
neutrophils by acting on their specific receptor G-CSFR. Under
physiological conditions, G-CSF affects the mobilization of
hematopoietic stem cells, progenitor cells and mature cells,
especially neutrophils, to the blood circulation. When the body is
infected, the serum G-CSF level is significantly increased to
promote neutrophil mobilization to the peripheral circulation.
Therefore, rhG-CSF can be used to treat neutropenia induced by
chemotherapy and radiation therapy. As an adjunct to cancer
therapy, G-CSF induces ErbB2 proto-oncogene expression in breast
cancer patients, making it an effective drug for improving the
sensitivity of breast cancer patients to trastuzumab (91). Currently, an increasing number of
studies have found that tumors with high G-CSF expression show
significant proliferative and metastatic properties and lead to
poor prognosis (17,47,48,67).
Therefore, the safety of G-CSF as an adjunct to cancer treatment
should be addressed.
Some basic studies have shown that G-CSF is a
promoter of tumor growth, which plays a role in immunosuppression
by increasing tumor angiogenesis and mobilizing MDSCs (13,14,46,51,72).
Kim et al (92), confirmed
that G-CSF treatment in mice with precise focused radiation
promoted tumor growth by stimulating angiogenesis in tumor-bearing
mice and reduced the antitumor effect of radiotherapy.
Coincidentally, in cervical cancer patients treated with
platinum-based chemotherapy drugs, G-CSF expression in tumors is an
indicator of poor prognosis in patients. Secreted G-CSF not only
has an antiapoptotic effect but also promotes the formation of
tumors by mobilizing MDSCs to inhibit T cell activity and Bv8
secretion (12). In some clinical
case reports, the use of safe therapeutic doses of G-CSF may cause
unpredictable side effects such as bone pain, local skin reactions
at the injection site and even spleen rupture or infarction
(93–96). This evidence also raised concerns
for clinical work. Effective methods are needed to evaluate the
G-CSF usage window.
The JAK/STAT signal transduction pathway has been
shown to be an important downstream pathway for G-CSF regulation in
cancer models such as colorectal cancer and is inseparable from
cancer proliferation and migration (57,97–99).
Studying the relationship between the G-CSF-JAK/STAT signaling
pathway and breast cancer can provide new insights for targeted
breast cancer therapy and its prognostic strategies.
The authors of the present study would like to thank
Professor Daya Luo (Department of Biochemistry and Molecular
Biology, School of Basic Medical Sciences, Nanchang University),
who made valuable suggestions regarding this manuscript.
The present review was supported by grants from the
National Natural Science Foundation of China (grant no. 81760509)
and the Natural Science Foundation of Jiangxi Province of China
(grant no. 20181BAB205043).
Not applicable.
LL and YL wrote the manuscript, prepared the figures
and repeatedly revised the paper. CZ collected the articles
regarding G-CSF expression. XX and XY reviewed drafts of the paper.
All authors read and approved the final manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Bradley TR and Metcalf D: The growth of
mouse bone marrow cells in vitro. Aust J Exp Biol Med Sci.
44:287–299. 1966. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ichikawa Y, Pluznik DH and Sachs L: In
vitro control of the development of macrophage and granulocyte
colonies. Proc Natl Acad Sci USA. 56:488–495. 1966. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Stanley ER and Heard PM: Factors
regulating macrophage production and growth. Purification and some
properties of the colony stimulating factor from medium conditioned
by mouse L cells. J Biol Chem. 252:4305–4312. 1977.PubMed/NCBI
|
|
4
|
Burgess AW, Camakaris J and Metcalf D:
Purification and properties of colony-stimulating factor from mouse
lung-conditioned medium. J Biol Chem. 252:1998–2003.
1977.PubMed/NCBI
|
|
5
|
Nicola NA, Metcalf D, Matsumoto M and
Johnson GR: Purification of a factor inducing differentiation in
murine myelomonocytic leukemia cells. Identification as granulocyte
colony-stimulating factor. J Biol Chem. 258:9017–9023.
1983.PubMed/NCBI
|
|
6
|
Ihle JN, Keller J, Henderson L, Klein F
and Palaszynski E: Procedures for the purification of interleukin 3
to homogeneity. J Immunol. 129:2431–2436. 1982.PubMed/NCBI
|
|
7
|
Metcalf D: The colony stimulating factors.
Discovery, development, and clinical applications. Cancer.
65:2185–2195. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fukunaga R, Ishizaka-Ikeda E and Nagata S:
Purification and characterization of the receptor for murine
granulocyte colony-stimulating factor. J Biol Chem.
265:14008–14015. 1990.PubMed/NCBI
|
|
9
|
Demetri GD and Griffin JD: Granulocyte
colony-stimulating factor and its receptor. Blood. 78:2791–2808.
1991. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bendall LJ and Bradstock KF: G-CSF: From
granulopoietic stimulant to bone marrow stem cell mobilizing agent.
Cytokine Growth Factor Rev. 25:355–367. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mitchell S, Li X, Woods M, Garcia J,
Hebard-Massey K, Barron R and Samuel M: Comparative effectiveness
of granulocyte colony-stimulating factors to prevent febrile
neutropenia and related complications in cancer patients in
clinical practice: A systematic review. J Oncol Pharm Pract.
22:702–716. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kawano M, Mabuchi S, Matsumoto Y, Sasano
T, Takahashi R, Kuroda H, Kozasa K, Hashimoto K, Isobe A, Sawada K,
et al: The significance of G-CSF expression and myeloid-derived
suppressor cells in the chemoresistance of uterine cervical cancer.
Sci Rep. 5:182172015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Okazaki T, Ebihara S, Asada M, Kanda A,
Sasaki H and Yamaya M: Granulocyte colony-stimulating factor
promotes tumor angiogenesis via increasing circulating endothelial
progenitor cells and Gr1+CD11b+ cells in cancer animal models. Int
Immunol. 18:1–9. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shojaei F, Wu X, Qu X, Kowanetz M, Yu L,
Tan M, Meng YG and Ferrara N: G-CSF-initiated myeloid cell
mobilization and angiogenesis mediate tumor refractoriness to
anti-VEGF therapy in mouse models. Proc Natl Acad Sci USA.
106:6742–6747. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Demers M, Krause DS, Schatzberg D,
Martinod K, Voorhees JR, Fuchs TA, Scadden DT and Wagner DD:
Cancers predispose neutrophils to release extracellular DNA traps
that contribute to cancer-associated thrombosis. Proc Natl Acad Sci
USA. 109:13076–13081. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gomes T, Várady CBS, Lourenço AL, Mizurini
DM, Rondon AMR, Leal AC, Gonçalves BS, Bou-Habib DC, Medei E and
Monteiro RQ: IL-1β blockade attenuates thrombosis in a neutrophil
extracellular trap-dependent breast cancer model. Front Immunol.
10:20882019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hollmén M, Karaman S, Schwager S, Lisibach
A, Christiansen AJ, Maksimow M, Varga Z, Jalkanen S and Detmar M:
G-CSF regulates macrophage phenotype and associates with poor
overall survival in human triple-negative breast cancer.
OncoImmunology. 5:e11151772015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kanda N, Fukushige S, Murotsu T, Yoshida
MC, Tsuchiya M, Asano S, Kaziro Y and Nagata S: Human gene coding
for granulocyte-colony stimulating factor is assigned to the
q21-q22 region of chromosome 17. Somat Cell Mol Genet. 13:679–684.
1987. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tsuchiya M, Kaziro Y and Nagata S: The
chromosomal gene structure for murine granulocyte
colony-stimulating factor. Eur J Biochem. 165:7–12. 1987.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shannon MF, Pell LM, Lenardo MJ, Kuczek
ES, Occhiodoro FS, Dunn SM and Vadas MA: A novel tumor necrosis
factor-responsive transcription factor which recognizes a
regulatory element in hemopoietic growth factor genes. Mol Cell
Biol. 10:2950–2959. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nagata S, Tsuchiya M, Asano S, Yamamoto O,
Hirata Y, Kubota N, Oheda M, Nomura H and Yamazaki T: The
chromosomal gene structure and two mRNAs for human granulocyte
colony-stimulating factor. EMBO J. 5:575–581. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
He RL, Zhou J, Hanson CZ, Chen J, Cheng N
and Ye RD: Serum amyloid A induces G-CSF expression and
neutrophilia via Toll-like receptor 2. Blood. 113:429–437. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Himes SR, Coles LS, Katsikeros R, Lang RK
and Shannon MF: HTLV-1 tax activation of the GM-CSF and G-CSF
promoters requires the interaction of NF-kB with other
transcription factor families. Oncogene. 8:3189–3197.
1993.PubMed/NCBI
|
|
24
|
Nishizawa M and Nagata S: Regulatory
elements responsible for inducible expression of the granulocyte
colony-stimulating factor gene in macrophages. Mol Cell Biol.
10:2002–2011. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nagata S, Tsuchiya M, Asano S, Kaziro Y,
Yamazaki T, Yamamoto O, Hirata Y, Kubota N, Oheda M, Nomura H, et
al: Molecular cloning and expression of cDNA for human granulocyte
colony-stimulating factor. Nature. 319:415–418. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Akira S, Isshiki H, Sugita T, Tanabe O,
Kinoshita S, Nishio Y, Nakajima T, Hirano T and Kishimoto T: A
nuclear factor for IL-6 expression (NF-IL6) is a member of a C/EBP
family. EMBO J. 9:1897–1906. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mitchell PJ and Tjian R: Transcriptional
regulation in mammalian cells by sequence-specific DNA binding
proteins. Science. 245:371–378. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shannon MF, Coles LS, Fielke RK, Goodall
GJ, Lagnado CA and Vadas MA: Three essential promoter elements
mediate tumour necrosis factor and interleukin-1 activation of the
granulocyte-colony stimulating factor gene. Growth Factors.
7:181–193. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hareng L, Meergans T, von Aulock S, Volk
HD and Hartung T: Cyclic AMP increases endogenous granulocyte
colony-stimulating factor formation in monocytes and THP-1
macrophages despite attenuated TNF-alpha formation. Eur J Immunol.
33:2287–2296. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nishizawa M, Tsuchiya M, Watanabe-Fukunaga
R and Nagata S: Multiple elements in the promoter of granulocyte
colony-stimulating factor gene regulate its constitutive expression
in human carcinoma cells. J Biol Chem. 265:5897–5902.
1990.PubMed/NCBI
|
|
31
|
Asano M, Nishizawa M and Nagata S: Three
individual regulatory elements of the promoter positively activate
the transcription of the murine gene encoding granulocyte
colony-stimulating factor. Gene. 107:241–246. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Brown CY, Lagnado CA and Goodall GJ: A
cytokine mRNA-destabilizing element that is structurally and
functionally distinct from A+U-rich elements. Proc Natl Acad Sci
USA. 93:13721–13725. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Arakawa T, Horan TP, Leong K, Prestrelski
SJ, Narhi LO and Hu S: Structure and activity of granulocyte
colony-stimulating factor derived from CHO cells containing cDNA
coding for alternatively spliced sequences. Arch Biochem Biophys.
316:285–289. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Souza LM, Boone TC, Gabrilove J, Lai PH,
Zsebo KM, Murdock DC, Chazin VR, Bruszewski J, Lu H, Chen KK, et
al: Recombinant human granulocyte colony-stimulating factor:
Effects on normal and leukemic myeloid cells. Science. 232:61–65.
1986. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kubota N, Orita T, Hattori K, Oh-eda M,
Ochi N and Yamazaki T: Structural characterization of natural and
recombinant human granulocyte colony-stimulating factors. J
Biochem. 107:486–492. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hill CP, Osslund TD and Eisenberg D: The
structure of granulocyte-colony-stimulating factor and its
relationship to other growth factors. Proc Natl Acad Sci USA.
90:5167–5171. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Senda T, Shimazu T, Matsuda S, Kawano G,
Shimizu H, Nakamura KT and Mitsui Y: Three-dimensional crystal
structure of recombinant murine interferon-beta. EMBO J.
11:3193–3201. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cheers C, Haigh AM, Kelso A, Metcalf D,
Stanley ER and Young AM: Production of colony-stimulating factors
(CSFs) during infection: Separate determinations of macrophage-,
granulocyte-, granulocyte-macrophage-, and multi-CSFs. Infect
Immun. 56:247–251. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Vellenga E, Rambaldi A, Ernst TJ,
Ostapovicz D and Griffin JD: Independent regulation of M-CSF and
G-CSF gene expression in human monocytes. Blood. 71:1529–1532.
1988. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Jones CE and Chan K: Interleukin-17
stimulates the expression of interleukin-8, growth-related
oncogene-alpha, and granulocyte-colony-stimulating factor by human
airway epithelial cells. Am J Respir Cell Mol Biol. 26:748–753.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Sano E, Ohashi K, Sato Y, Kashiwagi M,
Joguchi A and Naruse N: A possible role of autogenous IFN-beta for
cytokine productions in human fibroblasts. J Cell Biochem.
100:1459–1476. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Jardin F, Vasse M, Debled M, Dominique S,
Courville P, Callonnec F, Buchonnet G, Thiberville L and Tilly H:
Intense paraneoplastic neutrophilic leukemoid reaction related to a
G-CSF-secreting lung sarcoma. Am J Hematol. 80:243–245. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Uemura Y, Kobayashi M, Nakata H, Kubota T,
Saito T, Bandobashi K and Taguchi H: Role of protein kinase C in
expression of granulocyte-colony stimulating factor and granulocyte
macrophage-colony stimulating factor in lung cancer cells. Int J
Mol Med. 16:873–881. 2005.PubMed/NCBI
|
|
44
|
Tachibana M, Miyakawa A, Uchida A, Murai
M, Eguchi K, Nakamura K, Kubo A and Hata JI: Granulocyte
colony-stimulating factor receptor expression on human transitional
cell carcinoma of the bladder. Br J Cancer. 75:1489–1496. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Nomura H, Imazeki I, Oheda M, Kubota N,
Tamura M, Ono M, Ueyama Y and Asano S: Purification and
characterization of human granulocyte colony-stimulating factor
(G-CSF). EMBO J. 5:871–876. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chafe SC, Lou Y, Sceneay J, Vallejo M,
Hamilton MJ, McDonald PC, Bennewith KL, Möller A and Dedhar S:
Carbonic anhydrase IX promotes myeloid-derived suppressor cell
mobilization and establishment of a metastatic niche by stimulating
G-CSF production. Cancer Res. 75:996–1008. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lee CH, Lin SH, Chang SF, Chang PY, Yang
ZP and Lu SC: Extracellular signal-regulated kinase 2 mediates the
expression of granulocyte colony-stimulating factor in invasive
cancer cells. Oncol Rep. 30:419–424. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Park S, Kim ES, Noh DY, Hwang KT and Moon
A: H-Ras-specific upregulation of granulocyte colony-stimulating
factor promotes human breast cell invasion via matrix
metalloproteinase-2. Cytokine. 55:126–133. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Carvalho É, Hugo de Almeida V, Rondon AMR,
Possik PA, Viola JPB and Monteiro RQ: Protease-activated receptor 2
(PAR2) upregulates granulocyte colony stimulating factor (G-CSF)
expression in breast cancer cells. Biochem Biophys Res Commun.
504:270–276. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Samineni S, Zhang Z and Shively JE:
Carcinoembryonic antigen-related cell adhesion molecule 1
negatively regulates granulocyte colony-stimulating factor
production by breast tumor-associated macrophages that mediate
tumor angiogenesis. Int J Cancer. 133:394–407. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Welte T, Kim IS, Tian L, Gao X, Wang H, Li
J, Holdman XB, Herschkowitz JI, Pond A, Xie G, et al: Oncogenic
mTOR signalling recruits myeloid-derived suppressor cells to
promote tumour initiation. Nat Cell Biol. 18:632–644. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Cosman D: The hematopoietin receptor
superfamily. Cytokine. 5:95–106. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Touw IP and van de Geijn GJ: Granulocyte
colony-stimulating factor and its receptor in normal myeloid cell
development, leukemia and related blood cell disorders. Front
Biosci. 12:800–815. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
54
|
Avalos BR: Molecular analysis of the
granulocyte colony-stimulating factor receptor. Blood. 88:761–777.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Avalos BR, Parker JM, Ware DA, Hunter MG,
Sibert KA and Druker BJ: Dissociation of the Jak kinase pathway
from G-CSF receptor signaling in neutrophils. Exp Hematol.
25:160–168. 1997.PubMed/NCBI
|
|
56
|
Pencik J, Pham HT, Schmoellerl J, Javaheri
T, Schlederer M, Culig Z, Merkel O, Moriggl R, Grebien F and Kenner
L: JAK-STAT signaling in cancer: From cytokines to non-coding
genome. Cytokine. 87:26–36. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Fan Z, Li Y, Zhao Q, Fan L, Tan B, Zuo J,
Hua K and Ji Q: Highly expressed granulocyte colony-stimulating
factor (G-CSF) and granulocyte colony-stimulating factor receptor
(G-CSFR) in human gastric cancer leads to poor survival. Med Sci
Monit. 24:1701–1711. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Dwivedi P and Greis KD: Granulocyte
colony-stimulating factor receptor signaling in severe congenital
neutropenia, chronic neutrophilic leukemia, and related
malignancies. Exp Hematol. 46:9–20. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Fukui Y, Kawashima M, Kawaguchi K,
Takeuchi M, Hirata M, Kataoka TR, Sakurai T, Kataoka M, Kanao S,
Nakamoto Y, et al: Granulocyte-colony-stimulating factor-producing
metaplastic carcinoma of the breast with significant elevation of
serum interleukin-17 and vascular endothelial growth factor levels.
Int Cancer Conf J. 7:107–113. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Lawicki S, Czygier M, Omyła J and
Szmitkowski M: The plasma levels of granulocyte-colony stimulating
factor (G-CSF) and macrophage-colony stimulating factor (M-CSF) in
breast cancer patients. Pol Arch Med Wewn. 116:749–755. 2006.(In
Polish). PubMed/NCBI
|
|
61
|
Ławicki S, Będkowska GE, Wojtukiewicz M
and Szmitkowski M: Hematopoietic cytokines as tumor markers in
breast malignancies. A multivariate analysis with ROC curve in
breast cancer patients. Adv Med Sci. 58:207–215. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Bordbar E, Malekzadeh M, Ardekani MT,
Doroudchi M and Ghaderi A: Serum levels of G-CSF and IL-7 in
Iranian breast cancer patients. Asian Pac J Cancer Prev.
13:5307–5312. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Agresti R, Triulzi T, Sasso M, Ghirelli C,
Aiello P, Rybinska I, Campiglio M, Sfondrini L, Tagliabue E and
Bianchi F: Wound healing fluid reflects the inflammatory nature and
aggressiveness of breast tumors. Cells. 8:82019. View Article : Google Scholar
|
|
64
|
Wojtukiewicz MZ, Sierko E, Skalij P,
Kamińska M, Zimnoch L, Brekken RA and Thorpe PE: Granulocyte-colony
stimulating factor receptor, tissue factor, and VEGF-R bound VEGF
in human breast cancer in loco. Adv Clin Exp Med. 25:505–511. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Cao Y, Slaney CY, Bidwell BN, Parker BS,
Johnstone CN, Rautela J, Eckhardt BL and Anderson RL: BMP4 inhibits
breast cancer metastasis by blocking myeloid-derived suppressor
cell activity. Cancer Res. 74:5091–5102. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Kowanetz M, Wu X, Lee J, Tan M, Hagenbeek
T, Qu X, Yu L, Ross J, Korsisaari N, Cao T, et al:
Granulocyte-colony stimulating factor promotes lung metastasis
through mobilization of Ly6G+Ly6C+ granulocytes. Proc Natl Acad Sci
USA. 107:21248–21255. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Guo L, Chen G, Zhang W, Zhou L, Xiao T, Di
X, Wang Y, Feng L and Zhang K: A high-risk luminal A dominant
breast cancer subtype with increased mobility. Breast Cancer Res
Treat. 175:459–472. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Park J, Wysocki RW, Amoozgar Z, Maiorino
L, Fein MR, Jorns J, Schott AF, Kinugasa-Katayama Y, Lee Y, Won NH,
et al: Cancer cells induce metastasis-supporting neutrophil
extracellular DNA traps. Sci Transl Med. 8:361ra1382016. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Demers M and Wagner DD: Neutrophil
extracellular traps: A new link to cancer-associated thrombosis and
potential implications for tumor progression. OncoImmunology.
2:e229462013. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Leal AC, Mizurini DM, Gomes T, Rochael NC,
Saraiva EM, Dias MS, Werneck CC, Sielski MS, Vicente CP and
Monteiro RQ: tumor-derived exosomes induce the formation of
neutrophil extracellular traps: Implications for the establishment
of cancer-associated thrombosis. Sci Rep. 7:64382017. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Zhao CL, Zhang GP, Xiao ZZ, Ma ZK, Lei CP,
Song SY, Feng YY, Zhao YC and Feng XS: Recombinant human
granulocyte colony-stimulating factor promotes preinvasive and
invasive estrogen receptor-positive tumor development in MMTV-erbB2
mice. J Breast Cancer. 18:126–133. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Waight JD, Hu Q, Miller A, Liu S and
Abrams SI: Tumor-derived G-CSF facilitates neoplastic growth
through a granulocytic myeloid-derived suppressor cell-dependent
mechanism. PLoS One. 6:e276902011. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Espinoza-Sánchez NA, Vadillo E, Balandrán
JC, Monroy-García A, Pelayo R and Fuentes-Pananá EM: Evidence of
lateral transmission of aggressive features between different types
of breast cancer cells. Int J Oncol. 51:1482–1496. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Cavalloni G, Sarotto I, Pignochino Y,
Gammaitoni L, Migliardi G, Sgro L, Piacibello W, Risio M, Aglietta
M and Leone F: Granulocyte-colony stimulating factor upregulates
ErbB2 expression on breast cancer cell lines and converts primary
resistance to trastuzumab. Anticancer Drugs. 19:689–696. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Pondé N, Brandão M, El-Hachem G, Werbrouck
E and Piccart M: Treatment of advanced HER2-positive breast cancer:
2018 and beyond. Cancer Treat Rev. 67:10–20. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Motz GT and Coukos G: Deciphering and
reversing tumor immune suppression. Immunity. 39:61–73. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Almand B, Clark JI, Nikitina E, van Beynen
J, English NR, Knight SC, Carbone DP and Gabrilovich DI: Increased
production of immature myeloid cells in cancer patients: A
mechanism of immunosuppression in cancer. J Immunol. 166:678–689.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Quail DF and Joyce JA: Microenvironmental
regulation of tumor progression and metastasis. Nat Med.
19:1423–1437. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Aliper AM, Frieden-Korovkina VP, Buzdin A,
Roumiantsev SA and Zhavoronkov A: A role for G-CSF and GM-CSF in
nonmyeloid cancers. Cancer Med. 3:737–746. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Pickup MW, Owens P, Gorska AE, Chytil A,
Ye F, Shi C, Weaver VM, Kalluri R, Moses HL and Novitskiy SV:
Development of aggressive pancreatic ductal adenocarcinomas depends
on granulocyte colony stimulating factor secretion in carcinoma
cells. Cancer Immunol Res. 5:718–729. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Diaz-Montero CM, Salem ML, Nishimura MI,
Garrett-Mayer E, Cole DJ and Montero AJ: Increased circulating
myeloid-derived suppressor cells correlate with clinical cancer
stage, metastatic tumor burden, and doxorubicin-cyclophosphamide
chemotherapy. Cancer Immunol Immunother. 58:49–59. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Markowitz J, Wesolowski R, Papenfuss T,
Brooks TR and Carson WE III: Myeloid-derived suppressor cells in
breast cancer. Breast Cancer Res Treat. 140:13–21. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Sceneay J, Chow MT, Chen A, Halse HM, Wong
CS, Andrews DM, Sloan EK, Parker BS, Bowtell DD, Smyth MJ, et al:
Primary tumor hypoxia recruits CD11b+/Ly6Cmed/Ly6G+ immune
suppressor cells and compromises NK cell cytotoxicity in the
premetastatic niche. Cancer Res. 72:3906–3911. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Lin EY, Nguyen AV, Russell RG and Pollard
JW: Colony-stimulating factor 1 promotes progression of mammary
tumors to malignancy. J Exp Med. 193:727–740. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Aharinejad S, Paulus P, Sioud M, Hofmann
M, Zins K, Schäfer R, Stanley ER and Abraham D: Colony-stimulating
factor-1 blockade by antisense oligonucleotides and small
interfering RNAs suppresses growth of human mammary tumor
xenografts in mice. Cancer Res. 64:5378–5384. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Ries CH, Cannarile MA, Hoves S, Benz J,
Wartha K, Runza V, Rey-Giraud F, Pradel LP, Feuerhake F, Klaman I,
et al: Targeting tumor-associated macrophages with anti-CSF-1R
antibody reveals a strategy for cancer therapy. Cancer Cell.
25:846–859. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Mantovani A and Sica A: Macrophages,
innate immunity and cancer: Balance, tolerance, and diversity. Curr
Opin Immunol. 22:231–237. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Swierczak A, Cook AD, Lenzo JC, Restall
CM, Doherty JP, Anderson RL and Hamilton JA: The promotion of
breast cancer metastasis caused by inhibition of CSF-1R/CSF-1
signaling is blocked by targeting the G-CSF receptor. Cancer
Immunol Res. 2:765–776. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Huang J, Simpson JF, Glackin C, Riethorf
L, Wagener C and Shively JE: Expression of biliary glycoprotein
(CD66a) in normal and malignant breast epithelial cells. Anticancer
Res. 18((5A)): 3203–3212. 1998.PubMed/NCBI
|
|
91
|
Meixner A, Zenz R, Schonthaler HB, Kenner
L, Scheuch H, Penninger JM and Wagner EF: Epidermal JunB represses
G-CSF transcription and affects haematopoiesis and bone formation.
Nat Cell Biol. 10:1003–1011. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Kim JS, Son Y, Bae MJ, Lee M, Lee CG, Jo
WS, Kim SD and Yang K: Administration of granulocyte
colony-stimulating factor with radiotherapy promotes tumor growth
by stimulating vascularization in tumor-bearing mice. Oncol Rep.
34:147–154. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Alshamrani MA, Al-Foheidi M and Abdulrahim
AH: Granulocyte Colony Stimulating Factor (G-CSF) Induced splenic
infarction in breast cancer patient treated with dose-dense
chemotherapy regimen. Case Rep Oncol Med.
2019:81749862019.PubMed/NCBI
|
|
94
|
Kinjo Y, Kurita T, Ueda T, Kagami S,
Matsuura Y and Yoshino K: Acute arteritis after G-CSF
administration. Int Cancer Conf J. 8:77–80. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Lu X, Wu Y, Wang H and Xia L:
G-CSF-induced severe thrombocytopenia in a healthy donor: A rare
case report. Medicine (Baltimore). 98:e147862019. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Kim YG, Kim SR, Hwang SH, Jung JY, Kim HA
and Suh CH: Mesenteric vasculitis after G-CSF administration in a
severe neutropenic patient with systemic lupus erythematosus.
Lupus. 25:1381–1384. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Li W, Zhang X, Chen Y, Xie Y, Liu J, Feng
Q, Wang Y, Yuan W and Ma J: G-CSF is a key modulator of MDSC and
could be a potential therapeutic target in colitis-associated
colorectal cancers. Protein Cell. 7:130–140. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Kumar J, Fraser FW, Riley C, Ahmed N,
McCulloch DR and Ward AC: Granulocyte colony-stimulating factor
receptor signalling via Janus kinase 2/signal transducer and
activator of transcription 3 in ovarian cancer. Br J Cancer.
110:133–145. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Agarwal S, Lakoma A, Chen Z, Hicks J,
Metelitsa LS, Kim ES and Shohet JM: G-CSF Promotes Neuroblastoma
Tumorigenicity and Metastasis via STAT3-Dependent Cancer Stem Cell
Activation. Cancer Res. 75:2566–2579. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Nakata H, Uemura Y, Kobayashi M, Harada R
and Taguchi H: Cyclooxygenase-2 inhibitor NS-398 suppresses cell
growth and constitutive production of granulocyte-colony
stimulating factor and granulocyte macrophage-colony stimulating
factor in lung cancer cells. Cancer Sci. 94:173–180. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Cui YH, Suh Y, Lee HJ, Yoo KC, Uddin N,
Jeong YJ, Lee JS, Hwang SG, Nam SY, Kim MJ, et al: Radiation
promotes invasiveness of non-small-cell lung cancer cells through
granulocyte-colony-stimulating factor. Oncogene. 34:5372–5382.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Ramakrishna C and Cantin EM: IFNγ inhibits
G-CSF induced neutrophil expansion and invasion of the CNS to
prevent viral encephalitis. PLoS Pathog. 14:e10068222018.
View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Chang SF, Li HC, Huang YP, Tasi WJ, Chou
YY and Lu SC: SB203580 increases G-CSF production via a stem-loop
destabilizing element in the 3′ untranslated region in macrophages
independently of its effect on p38 MAPK activity. J Biomed Sci.
23:32016. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Fujimoto A, Akifusa S, Hirofuji T and
Yamashita Y: Involvement of suppressor of cytokine signaling-1 in
globular adiponectin-induced granulocyte colony-stimulating factor
in RAW 264 cell. Mol Immunol. 48:2052–2058. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Kamio N, Akifusa S, Yamaguchi N and
Yamashita Y: Induction of granulocyte colony-stimulating factor by
globular adiponectin via the MEK-ERK pathway. Mol Cell Endocrinol.
292:20–25. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Zhang L, Yang M, Wang Q, Liu M, Liang Q,
Zhang H and Xiao X: HSF1 regulates expression of G-CSF through the
binding element for NF-IL6/CCAAT enhancer binding protein beta. Mol
Cell Biochem. 352:11–17. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Aoki Y, Hirano D, Kodama H, Nishi Y and
Nakamura M: Stimulation of G-CSF gene expression in the macrophage
cell line by contact with extracellular matrix proteins and a pre-B
leukaemia cell line. Cytokine. 10:596–602. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Chou YY and Lu SC: Inhibition by rapamycin
of the lipoteichoic acid-induced granulocyte-colony stimulating
factor expression in mouse macrophages. Arch Biochem Biophys.
508:110–119. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Sallerfors B and Olofsson T:
Granulocyte-macrophage colony-stimulating factor (GM-CSF) and
granulocyte colony-stimulating factor (G-CSF) secretion by adherent
monocytes measured by quantitative immunoassays. Eur J Haematol.
49:199–207. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Tajuddin T, Ryan EJ, Norris S, Hegarty JE
and O'Farrelly C: Interferon-α suppressed granulocyte colony
stimulating factor production is reversed by CL097, a TLR7/8
agonist. J Gastroenterol Hepatol. 25:1883–1890. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Ichinose Y, Hara N, Ohta M, Aso H, Chikama
H, Kawasaki M, Kubota I, Shimizu T and Yagawa K: Recombinant
granulocyte colony-stimulating factor and lipopolysaccharide
maintain the phenotype of and superoxide anion generation by
neutrophils. Infect Immun. 58:1647–1652. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Lindemann A, Riedel D, Oster W,
Ziegler-Heitbrock HW, Mertelsmann R and Herrmann F:
Granulocyte-macrophage colony-stimulating factor induces cytokine
secretion by human polymorphonuclear leukocytes. J Clin Invest.
83:1308–1312. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Lu L, Srour EF, Warren DJ, Walker D,
Graham CD, Walker EB, Jansen J and Broxmeyer HE: Enhancement of
release of granulocyte- and granulocyte-macrophage
colony-stimulating factors from phytohemagglutinin-stimulated
sorted subsets of human T lymphocytes by recombinant human tumor
necrosis factor-alpha. Synergism with recombinant human IFN-gamma.
J Immunol. 141:201–207. 1988.PubMed/NCBI
|
|
114
|
Lennard Richard ML, Brandon D, Lou N, Sato
S, Caldwell T, Nowling TK, Gilkeson G and Zhang XK: Acetylation
impacts Fli-1-driven regulation of granulocyte colony stimulating
factor. Eur J Immunol. 46:2322–2332. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Rajavashisth TB, Andalibi A, Territo MC,
Berliner JA, Navab M, Fogelman AM and Lusis AJ: Induction of
endothelial cell expression of granulocyte and macrophage
colony-stimulating factors by modified low-density lipoproteins.
Nature. 344:254–257. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Seelentag WK, Mermod JJ, Montesano R and
Vassalli P: Additive effects of interleukin 1 and tumour necrosis
factor-alpha on the accumulation of the three granulocyte and
macrophage colony-stimulating factor mRNAs in human endothelial
cells. EMBO J. 6:2261–2265. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Saba S, Soong G, Greenberg S and Prince A:
Bacterial stimulation of epithelial G-CSF and GM-CSF expression
promotes PMN survival in CF airways. Am J Respir Cell Mol Biol.
27:561–567. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Suzukawa M, Koketsu R, Baba S, Igarashi S,
Nagase H, Yamaguchi M, Matsutani N, Kawamura M, Shoji S, Hebisawa
A, et al: Leptin enhances ICAM-1 expression, induces migration and
cytokine synthesis, and prolongs survival of human airway
epithelial cells. Am J Physiol Lung Cell Mol Physiol.
309:L801–L811. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Numasaki M, Takahashi H, Tomioka Y and
Sasaki H: Regulatory roles of IL-17 and IL-17F in G-CSF production
by lung microvascular endothelial cells stimulated with IL-1beta
and/or TNF-alpha. Immunol Lett. 95:97–104. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Witowski J, Ksiazek K, Warnecke C, Kuźlan
M, Korybalska K, Tayama H, Wiśniewska-Elnur J, Pawlaczyk K,
Trómińska J, Breborowicz A, et al: Role of mesothelial cell-derived
granulocyte colony-stimulating factor in interleukin-17-induced
neutrophil accumulation in the peritoneum. Kidney Int. 71:514–525.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Demetri GD, Zenzie BW, Rheinwald JG and
Griffin JD: Expression of colony-stimulating factor genes by normal
human mesothelial cells and human malignant mesothelioma cells
lines in vitro. Blood. 74:940–946. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Carr MJ, Li Y, Rezakhanlou AM and Ghahary
A: Keratinocyte-releasable factors stimulate the expression of
granulocyte colony-stimulating factor in human dermal fibroblasts.
J Cell Biochem. 118:308–317. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Ramachandran R, Morice AH and Compton SJ:
Proteinase-activated receptor2 agonists upregulate granulocyte
colony-stimulating factor, IL-8, and VCAM-1 expression in human
bronchial fibroblasts. Am J Respir Cell Mol Biol. 35:133–141. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Koeffler HP, Gasson J, Ranyard J, Souza L,
Shepard M and Munker R: Recombinant human TNF alpha stimulates
production of granulocyte colony-stimulating factor. Blood.
70:55–59. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Seelentag W, Mermod JJ and Vassalli P:
Interleukin 1 and tumor necrosis factor-alpha additively increase
the levels of granulocyte-macrophage and granulocyte
colony-stimulating factor (CSF) mRNA in human fibroblasts. Eur J
Immunol. 19:209–212. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Zgheib A, Lamy S and Annabi B:
Epigallocatechin gallate targeting of membrane type 1 matrix
metalloproteinase-mediated Src and Janus kinase/signal transducers
and activators of transcription 3 signaling inhibits transcription
of colony-stimulating factors 2 and 3 in mesenchymal stromal cells.
J Biol Chem. 288:13378–13386. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Fibbe WE, van Damme J, Billiau A, Goselink
HM, Voogt PJ, van Eeden G, Ralph P, Altrock BW and Falkenburg JH:
Interleukin 1 induces human marrow stromal cells in long-term
culture to produce granulocyte colony-stimulating factor and
macrophage colony-stimulating factor. Blood. 71:430–435. 1988.
View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Tesio M, Oser GM, Baccelli I, Blanco-Bose
W, Wu H, Göthert JR, Kogan SC and Trumpp A: Pten loss in the bone
marrow leads to G-CSF-mediated HSC mobilization. J Exp Med.
210:2337–2349. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Grace MB, Singh VK, Rhee JG, Jackson WE
III, Kao TC and Whitnall MH: 5-AED enhances survival of irradiated
mice in a G-CSF-dependent manner, stimulates innate immune cell
function, reduces radiation-induced DNA damage and induces genes
that modulate cell cycle progression and apoptosis. J Radiat Res
(Tokyo). 53:840–853. 2012. View Article : Google Scholar
|
|
130
|
Kimura A, Kinjyo I, Matsumura Y, Mori H,
Mashima R, Harada M, Chien KR, Yasukawa H and Yoshimura A: SOCS3 is
a physiological negative regulator for granulopoiesis and
granulocyte colony-stimulating factor receptor signaling. J Biol
Chem. 279:6905–6910. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Smith A, Witte E, McGee D, Knott J, Narang
K and Racicot K: Cortisol inhibits CSF2 and CSF3 via DNA
methylation and inhibits invasion in first-trimester trophoblast
cells. Am J Reprod Immunol. 78:e127412017. View Article : Google Scholar
|
|
132
|
Ordelheide AM, Gommer N, Böhm A, Hermann
C, Thielker I, Machicao F, Fritsche A, Stefan N, Häring HU and
Staiger H: Granulocyte colony-stimulating factor (G-CSF): A
saturated fatty acid-induced myokine with insulin-desensitizing
properties in humans. Mol Metab. 5:305–316. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Hudock KM, Liu Y, Mei J, Marino RC, Hale
JE, Dai N and Worthen GS: Delayed resolution of lung inflammation
in Il-1rn-/- mice reflects elevated IL-17A/granulocyte
colony-stimulating factor expression. Am J Respir Cell Mol Biol.
47:436–444. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Soria-Castro I, Krzyzanowska A, Pelaéz ML,
Regadera J, Ferrer G, Montoliu L, Rodríguez-Ramos R, Fernández M
and Alemany S: Cot/tpl2 (MAP3K8) mediates myeloperoxidase activity
and hypernociception following peripheral inflammation. J Biol
Chem. 285:33805–33815. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Janelle MF, Doucet A, Bouchard D,
Bourbonnais Y and Tremblay GM: Increased local levels of
granulocyte colony-stimulating factor are associated with the
beneficial effect of pre-elafin (SKALP/trappin-2/WAP3) in
experimental emphysema. Biol Chem. 387:903–909. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Bohannon JK, Luan L, Hernandez A, Afzal A,
Guo Y, Patil NK, Fensterheim B and Sherwood ER: Role of G-CSF in
monophosphoryl lipid A-mediated augmentation of neutrophil
functions after burn injury. J Leukoc Biol. 99:629–640. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Ellis GS, Carlson DE, Hester L, He JR,
Bagby GJ, Singh IS and Hasday JD: G-CSF, but not corticosterone,
mediates circulating neutrophilia induced by febrile-range
hyperthermia. J Appl Physiol. 98:1799–1804. 2005. View Article : Google Scholar : PubMed/NCBI
|