Introduction
Endothelial cell aging is a major risk factor for
cardiovascular disease (CVD) development (1–3).
Numerous studies have shown that hydrogen peroxide
(H2O2)-induced vascular endothelial
dysfunction is partially responsible for the development of aging
(4–6). H2O2-induced
endothelial dysfunction reflects a loss of the balance between pro-
and anti-oxidant, pro- and anti-inflammatory, and pro- and
anti-thrombotic signals, all of which contribute to increased
release of proinflammatory cytokines, including plasminogen
activator inhibitor-1 (PAI-1), and a decline in antioxidants
including endothelial nitric oxide synthase (eNOS) and nitric oxide
(NO) (7–11). Thus, the prevention of endothelial
dysfunction is essential to treat endothelial aging and CVD.
Ginseng, the root of Panax ginseng CA Meyer,
is one of the most popular herbs in traditional Asian medicine. A
growing body of evidence suggests that ginsenoside Rb1 (Rb1), a
major component of ginsenosides extracted from ginseng, has various
biological activities including antioxidative stress relief,
anti-obesity and anti-inflammation (12–14).
One of our previous studies also suggested that Rb1 at the
concentration of 10–40 µM inhibits free fatty acid-induced
inflammation partially through the blockade of nuclear factor
(NF)-κB phosphorylation in 3T3-L1 adipocytes (15). Additionally, another study by our
group demonstrated that Rb1 at the concentration of 20 µM
attenuates human umbilical vein endothelial cell (HUVEC) senescence
by improving the redox status (16). However, the range of effective
concentrations and further modulated mechanisms of Rb1 in the
endothelium are not fully elucidated.
AMP-activated protein kinase (AMPK) is a
heterotrimeric member of an evolutionarily conserved protein kinase
family that is sensitive to changes in oxygen tension and ATP
consumption (17). Accumulating
evidence has revealed that AMPK participates in the regulation of
lipid metabolism, inflammation and angiogenesis in various animal
models and cell types (18–22).
Ido et al (23) reported
that AMPK protects endothelial cells from the adverse effects of
sustained hyperglycemia. Nagata et al (22) demonstrated that endothelial AMPK
signaling plays a critical role in blood vessel recruitment to
tissues responding to ischemic stress. In addition, studies have
shown that AMPK exerts its beneficial role through multiple
signaling pathways, including activation of eNOS and production of
NO (24,25). However, it is still unclear whether
endothelial senescence, eNOS activation and NO synthesis in HUVECs
in response to Rb1 are related to the activation of AMPK.
Complementing our previous studies, the present
study was undertaken to investigate the protective effects of Rb1
against H2O2-induced HUVEC dysfunction
mediated by AMPK and the underlying mechanisms.
Materials and methods
Cell culture and treatments
Primary HUVECs were isolated from different six
neonatal umbilical cords as previously described (26). Briefly, HUVECs at passages 2–4 were
maintained in M199 medium (Invitrogen, Thermo Fisher Scientific,
Inc.) supplemented with 20% fetal bovine serum (Hyclone, GE
Healthcare Life Sciences) and 60 µg/ml endothelial cell growth
supplement (BD Biosciences) at 37°C in a 5% CO2
incubator and then exposed to the desired treatment in triplicate.
The isolation procedure for HUVECs was approved by the Research
Committee at the Third Affiliated Hospital of Sun Yat-sen
University (approval nos. 2010-2–48 and 2018-02-057-01). The donors
were negative for human immunodeficiency virus and hepatitis B
virus and provided written informed consent to donate the umbilical
cords.
To induce senescence, isolated HUVECs were treated
with 60 µM H2O2 (Sigma-Aldrich, Merck KGaA)
for 1 h and then cultured for another 24 h at 37°C. Rb1 (16071307,
Chengdu Pufei De Biotech Co., Ltd.) used in the present study was
extracted from Panax ginseng by HPLC according to the
manufacturer's instructions and the purity of Rb1 used in the
present study was 98.85%. To evaluate the effect of Rb1 on
senescence, the cells were pretreated with 10 or 20 µM Rb1 for 30
min prior to H2O2 treatment. To measure the
effect of the SIRT1 inhibitor nicotinamide (NAM; Sigma-Aldrich,
Merck KGaA) and AMPK inhibitor compound C (Sigma-Aldrich, Merck
KGaA), the cells were incubated with 20 mM NAM (27) or 8 µM compound C for 30 min as
reported previously (18,28,29)
and then treated with or without Rb1 at concentrations of 10 or 20
µM for 30 min before H2O2 treatment. At the
end of each experiment, the cultured supernatants and monolayered
cells were harvested for analyses.
Measurement of NO production
NO production was evaluated by measuring the
accumulation of nitrites. The Griess method (30) was used to detect NO using a NO
assay kit (Beyotime Institute of Biotechnology), following the
manufacturer's instructions. Briefly, after the cells were cultured
and treated as described above, 50 µl culture supernatant was
incubated with 50 µl Greiss reagent I and 50 µl Greiss reagent II
in a 96-well microplate at room temperature for 30 min. The optical
density was measured with a Victor microplate reader (PerkinElmer,
Inc.) at 540 nm. Nitrite concentrations in the medium were
calculated according to a standard curve.
Senescence-associated β-galactosidase
activity assay
Senescence was detected using a
senescence-associated β-galactosidase (SA-β-gal)-positive approach
according to a published protocol (31). After HUVECs were washed twice with
prechilled PBS, the cells were fixed with 2% formaldehyde plus 1%
glutaraldehyde for 5–10 min at room temperature. The cells were
then washed twice with prechilled PBS for 3 min and stained with a
staining solution [40 mmol/l citric acid/sodium phosphate buffer, 5
mmol/l potassium ferrocyanide
(K4[Fe(CN)6]3H2O), 5 mmol/l
potassium ferricyanide (K3[Fe(CN)6]), 150
mmol/l sodium chloride, 2 mmol/l magnesium chloride and 1 mg/ml
X-gal] overnight at 37°C without CO2. Senescent cells
were identified as blue-stained cells under a TS100 inverted
microscope (Nikon Corporation) at ×100 magnification. At least 400
cells were examined to determine the percentage of
SA-β-gal-positive cells in each group.
NAD+/NADH assay
The cellular NADP+/NADPH ratio was
determined using a NAD+/NADH Quantification Kit
(Beyotime Institute of Biotechnology), according to the
manufacturer's instructions. In brief, HUVECs
(1×106/well) were seeded in six-well plates and exposed
to the experimental conditions. To measure the
NADP+/NADPH ratio, the cells were harvested, lysed with
200 µl NAD+/NADH buffer and gently pipetted to promote
cell lysis. Then, 50–100 µl of the samples were collected and
incubated for 30 min at 60°C. Then, 20 µl of the reacted samples
were added to a 96-well plate and analyzed at 450 nm as the
reference wavelength in the Victor microplate reader. The
NAD+/NADH ratio was calculated according to a standard
curve and normalized to the cell number, following the
manufacturer's protocol.
Measurement of intracellular ATP
Determination of intracellular ATP in the indicated
groups of HUVECs was performed by a bioluminescence assay (ATP
Assay kit; Beyotime Institute of Biotechnology), according to the
manufacturer's instructions. Briefly, the cells were washed twice
with prechilled PBS and then lysed in lysis buffer on ice. Then,
the samples were harvested and centrifuged at 12,000 × g for 5 min
at 4°C and the supernatants were collected for subsequent analysis.
After the reaction solutions containing luciferase and luciferin
were added and background luminescence was measured, the ATP
standard solution and samples were added and luminescence was
measured. Then, the background luminescence was subtracted and the
standard curve was constructed. The ATP concentrations were
calculated from the standard curve and normalized to total protein
content.
Western blot analysis
HUVECs were grown in six-well plates. After the
indicated treatments, the cells were washed twice with prechilled
PBS and then lysed in radioimmunoprecipitation buffer with a
protease inhibitor cocktail, phenylmethylsulfonyl fluoride and
sodium orthovanadate (Santa Cruz Biotechnology, Inc.). The protein
concentration was measured by the Bradford method. Proteins (30 µg)
in 30 µl reducing sample buffer were boiled for 5 min at 100°C and
then resolved by SDS-PAGE (8 or 12% gels) for 2 h at 100 V. The
proteins were transferred onto a polyvinylidene difluoride membrane
for 90 min at 100 V. After transfer, the membrane was incubated in
25 ml blocking buffer [1X Tris-buffered saline (TBS) and 0.1%
Tween-20 with 5% non-fat dry milk] for 1 h at room temperature. The
primary antibodies were rabbit polyclonal anti-eNOS (1:500; Cell
Signaling Technology, Inc.), Rabbit polyclonal anti-PAI-1 (1:500;
Cell Signaling Technology, Inc.), Rabbit polyclonal anti-AMPK
(1:2,000; Cell Signaling Technology, Inc.), rabbit polyclonal
anti-phosphorylated AMPK (Thr172; 1:1,000; Cell Signaling
Technology, Inc.), rabbit polyclonal anti-SIRT1 (1:1,000; Cell
Signaling Technology, Inc.), and rabbit polyclonal anti-GAPDH
(control, 1:5,000; Cell Signaling Technology, Inc.). The membrane
was incubated with the primary antibody in 10 ml primary antibody
dilution buffer with gentle agitation overnight at 4°C. After
washing three times for 10 min each with 15 ml of 10X TBS/0.1%
TBST, the membrane was incubated with a horseradish
peroxidase-conjugated secondary antibody (1:3,000; cat. no. 7074;
Cell Signaling Technology, Inc.) in 10 ml blocking buffer with
gentle agitation for 1 h at room temperature, followed by 3 washes
for 10 min each. Membranes were developed using the enhanced
chemiluminescence detection method (EMD Millipore). The signals
were quantified using Quantity One software (version 4.6.9; Bio-Rad
Laboratories, Inc.).
Statistical analysis
Results were obtained from at least three
independent experiments (n≥3) if not otherwise stated. Data are
expressed as the mean ± standard deviation (n≥3). Statistical
significance was calculated by one-way or two-way analysis of
variance with Bonferroni's post-hoc test using GraphPad Prism
software (version 5.0; GraphPad Software, Inc.). P<0.05 was
considered to indicate a statistically significant difference.
Results
Effects of H2O2
and Rb1 treatment in HUVECs
A number of studies have reported that
H2O2 is a major inducer of cell senescence
and oxidative stress in HUVECs and other different cell lines
(32–35), which plays a central role in cell
dysfunction. H2O2 was used at a concentration
of 60 µM in the present study based on our previous studies
(16,36,37)
in which 60 µM H2O2 was reported to induce
cell senescence and oxidative stress sufficiently.
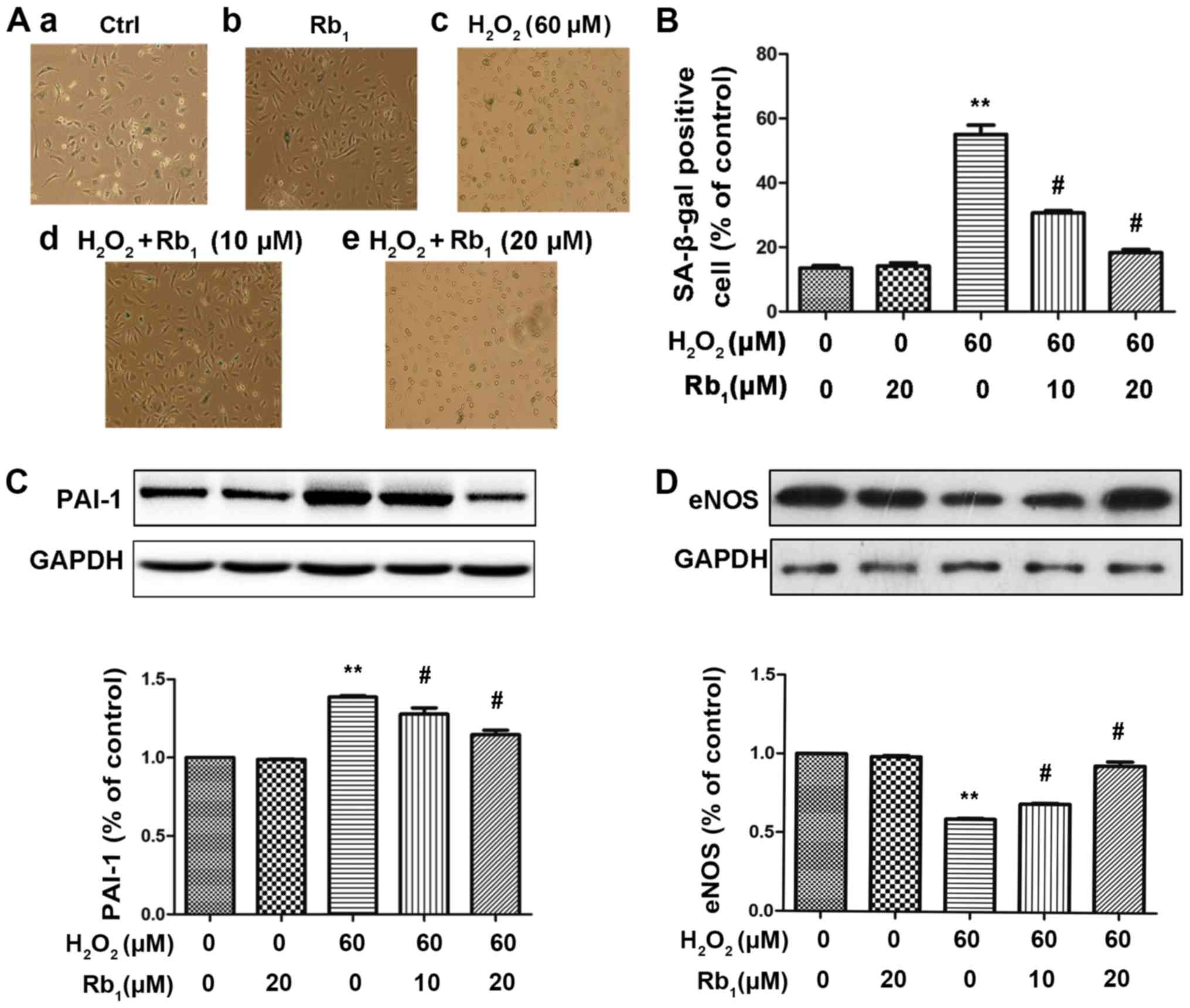
Senescence-associated β-galactosidase (SA-β-gal) staining was used
to determine the degree of cell senescence. Aging cells stained
blue indicated the senescent phenotype of HUVECs. In line with our
previous studies (16,36,37),
it was demonstrated that 60 µM H2O2 increased
the SA-β-gal+ cell number (blue staining). In addition,
the H2O2 treated group had more cells with a
clear outline, enlarged cell body and difficulty in adhering to the
bottom of the culture plate, all characteristics of stressed
senescent cells (38,39). By contrast, pretreatment with Rb1
decreased the SA-β-gal+ cell number and attenuated cell
senescence induced by H2O2 in a
dose-dependent manner (Fig. 1A and
B). PAI-1 and eNOS expression were then measured after the same
treatment. The results demonstrated that 60 µM
H2O2 increased PAI-1 expression (Fig. 1C) and decreased eNOS expression
(Fig. 1D) in HUVECs, which were
restored by Rb1 in a dose-dependent manner.
Effects of H2O2
and Rb1 on SIRT1 expression, AMPK phosphorylation and the
NAD+/NADH ratio
A number of studies have shown that SIRT1 exhibits
anti-inflammatory (40–42) and antioxidant effects (43–45)
in the endothelium. The effect of Rb1 on SIRT1 and phosphorylation
of the catalytic subunit of AMPK (Thr172) in the presence or
absence of H2O2 was examined. The results
demonstrated that 60 µM H2O2 inhibited SIRT1
expression (Fig. 2A and B) as well
as phosphorylation of AMPK (Fig. 2A
and C) and that treatment with Rb1 at 10 and 20 µM restored
Sirt-1 expression (Fig. 2A and B)
and AMPK phosphorylation (Fig. 2A and
C) in a dose-dependent manner.
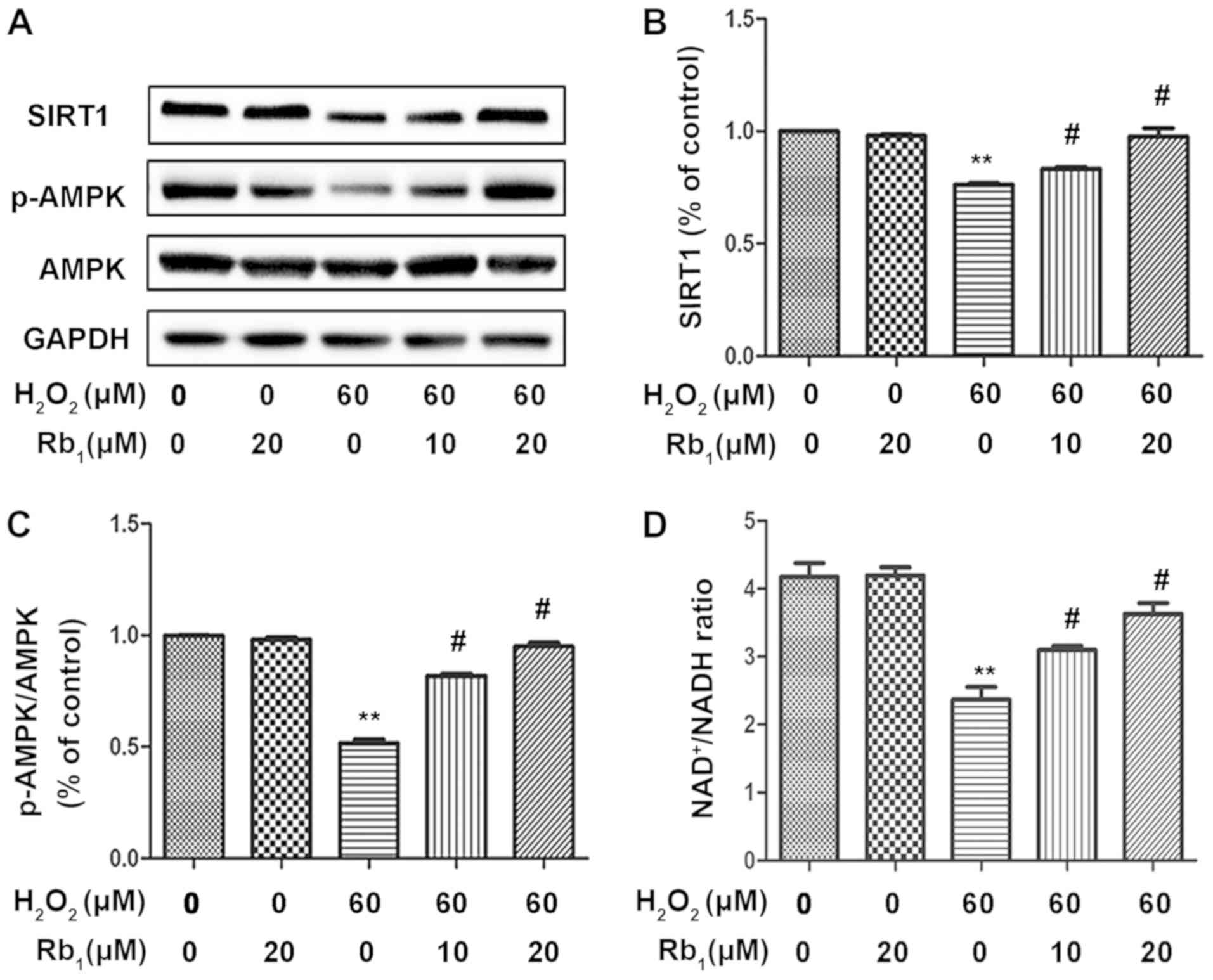 | Figure 2.Rb1 restores the
H2O2-induced reduction of SIRT1, promotes
phosphorylation of AMPK and increases the NAD+/NADH
ratio. The HUVECs were pretreated with 10–20 µM Rb1, treated with
or without 60 µM H2O2 for 1 h and then
cultured for 24 h. (A) The levels of SIRT1, phosphorylated AMPK,
AMPK and GAPDH were determined by western blot analysis. (B)
Quantitative analysis of SIRT1 expression ratio and SIRT1
expression levels. (C) Quantitative analysis of phosphorylated AMPK
ratio and phosphorylated AMPK expression levels. (D)
NAD+/NADH ratio measured using a colorimetric assay.
**P<0.01 vs. control group; #P<0.05 vs. 60 µM
H2O2 group. Rb1, ginsenoside Rb1;
H2O2, hydrogen peroxide; SIRT1, Sirtuin-1;
AMPK, AMP-activated protein kinase; HUVEC, human umbilical vein
endothelial cell; p, phosphorylated. |
The intracellular NAD+/NADH ratio in the
presence or absence of Rb1 was further examined and it was
identified that treatment with 60 µM H2O2
reduced the NAD+/NADH ratio, which was restored with Rb1
pretreatment (Fig. 2D). The data
confirmed that H2O2-induced decrease in the
NAD+/NADH ratio were associated with reduced SIRT1
expression and it was demonstrated for the first time, to the best
of the authors' knowledge, that treatment with Rb1 at 10 and 20 µM
Rb1 prevented the reductions in SIRT1 and the NAD+/NADH
ratio, and inhibited phosphorylated AMPK, in HUVECs exposed to
H2O2.
Involvement of AMPK in Rb1-mediated
inhibition of the H2O2-induced oxidative
response
Next, it was determined whether AMPK was involved in
the inhibitory effects of Rb1 on the
H2O2-induced oxidative response. HUVECs were
treated with 10 or 20 µM Rb1 for 24 h in the presence or absence of
compound. C, a specific inhibitor of AMPK. As shown in Fig. 3A, compound C clearly downregulated
the phosphorylation of AMPK at 6, 8 and 10 µM in a dose-dependent
manner. According to the results, 8 µM compound C was chosen to
inhibit the phosphorylation of AMPK in the following experiment.
Pretreatment of HUVECs with compound C significantly abolished the
inhibitory effects of Rb1 on H2O2-induced
PAI-1 expression (Fig. 3C). Rb1
also significantly restored eNOS expression (Fig. 3D) and NO production (Fig. 3E) and this beneficial effect was
markedly reversed by pretreatment of HUVECs with compound C
(Fig. 3). The results suggest that
AMPK activation is essential for the inhibitory effect of Rb1 on
H2O2-induced senescence and the oxidative
response in HUVECs.
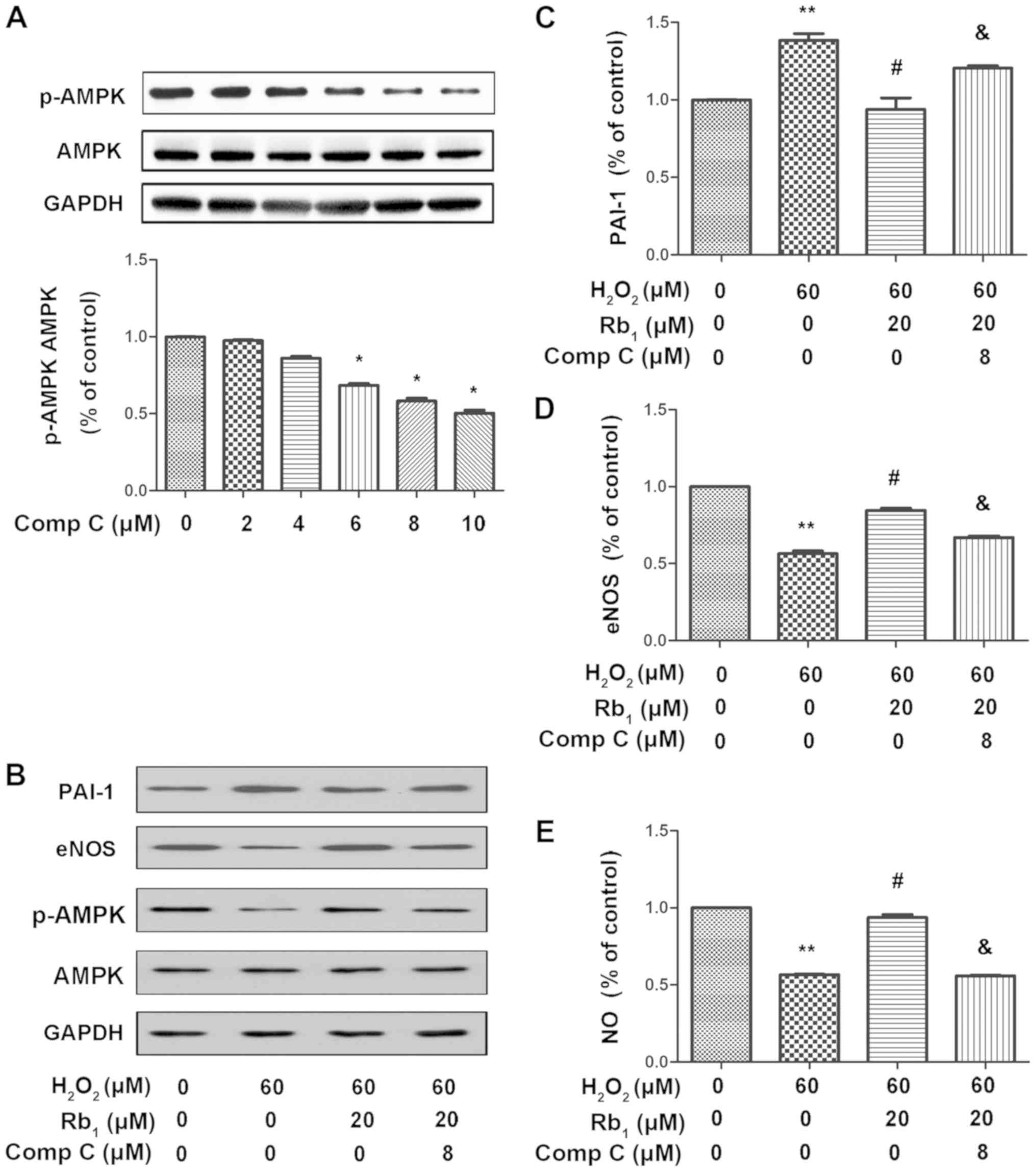 | Figure 3.Compound C suppresses the beneficial
effects of Rb1 on H2O2-induced antioxidant
and anti-inflammatory functions. The HUVECs were pretreated with
the indicated concentrations of compound C for 30 min and then
pretreated with 20 µM Rb1 for 30 min, followed by treatment with or
without 60 µM H2O2 for 1 h and then culture
for 24 h. (A) Phosphorylation of AMPK in HUVECs pretreated with
compound C (0, 2, 4, 6, 8 and 10 µM) for 30 min detected by western
blot analysis and semi-quantified by densitometry. Total AMPK and
GAPDH were also determined. (B) The protein levels of PAI-1, eNOS,
phosphorylated AMPK, AMPK and GAPDH were determined by western
blot. (C) Quantitative analysis of PAI-1 expression ratio and PAI-1
expression levels. (D) Quantitative analysis of eNOS expression
ratio and eNOS expression levels. (E) NO production measured by the
Griess reaction and expressed as the percentage of the control.
Results represent three independent experiments. *P<0.05 and
**P<0.01 vs. control group; #P<0.05 vs. 60 µM
H2O2 group; &P<0.05 vs. 60
µM H2O2+Rb1 group. Rb1, ginsenoside Rb1;
H2O2, hydrogen peroxide; HUVEC, human
umbilical vein endothelial cell; AMPK, AMP-activated protein
kinase; PAI-1, plasminogen activator inhibitor-1; eNOS, endothelial
nitric oxide synthase; p, phosphorylated; NO, nitric oxide; Comp C,
compound C. |
Involvement of SIRT1 in Rb1-mediated
inhibition of the H2O2-induced oxidative
response
To further understand the role of SIRT1 in the AMPK
pathway, it was investigated whether inactivation of SIRT1 by
nicotinamide affected H2O2-induced cell
injury. As Guo et al (27)
reported that 20 mM NAM decreases SIRT1 gene expression
significantly, HUVECs were treated with H2O2
in the presence or absence of 20 mM NAM, followed by observation of
the effects on H2O2-induced senescence and
the expression of antioxidant genes. As shown in Fig. 4, NAM reduced the phosphorylation of
AMPK (Fig. 4A and B), indicating
that AMPK is the downstream protein modulated by SIRT1.
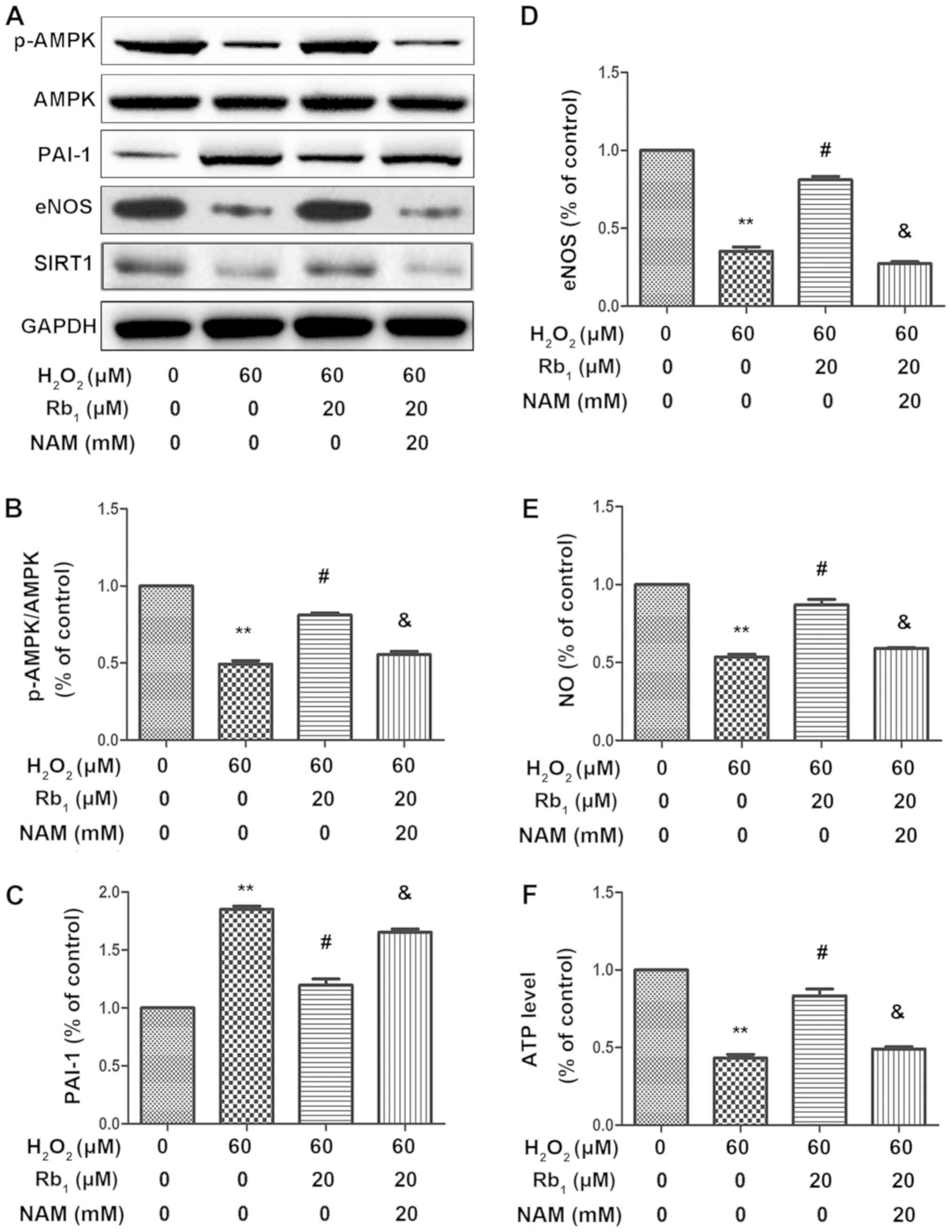 | Figure 4.NAM blocks the protective effects of
Rb1 against H2O2-induced senescence in
HUVECs. (A) The protein levels of phosphorylated AMPK, AMPK, PAI-1,
eNOS, SIRT1 and GAPDH were determined by western blot. (B)
Quantitative analysis of phosphorylated AMPK expression ratio and
phosphorylated AMPK levels. Total AMPK and GAPDH were also
determined. (C) Quantitative analysis of PAI-1 expression ratio and
PAI-1 expression levels. (D) Quantitative analysis of eNOS
expression ratio and eNOS expression levels. (E) NO production
measured by the Griess reaction and expressed as the percentage of
the control. (F) The amount of ATP in the cells was measured using
an ATP determination kit. Results represent three independent
experiments. **P<0.01, vs. control group. #P<0.05
vs. 60 µM H2O2 group.
&P<0.05, vs. 60 µM H2O2+Rb1
group. NAM, nicotinamide; Rb1, ginsenoside Rb1;
H2O2, hydrogen peroxide; HUVEC, human
umbilical vein endothelial cell; AMPK, AMP-activated protein
kinase; PAI-1, plasminogen activator inhibitor-1; eNOS, endothelial
nitric oxide synthase; SIRT1, Sirtuin-1; p, phosphorylated; NO,
nitric oxide. |
As AMPK is one of the most important proteins in
modulating the cellular energy metabolism and ATP is a major
downstream product of mitochondrial energy coordinators, the
effects of NAM with or without Rb1 on the ATP level were next
investigated. Rb1 protected HUVECs from
H2O2-induced PAI-1 expression (Fig. 4C) and rescued the downregulation of
eNOS expression (Fig. 4D) and NO
production (Fig. 4E). As shown in
Fig. 4F, treatment with Rb1
increased the ATP level under H2O2-induced
oxidative stress. However, the promotional effect on the ATP level
by Rb1 was not observed in the NAM-treated group (Fig. 4F). The beneficial effects of Rb1 on
H2O2-induced injuries were reversed by
treatment with NAM (Fig. 4). These
in vitro data confirmed that inhibition of SIRT1 by NAM
blocks Rb1 to increase phosphorylation of AMPK, ATP production,
eNOS expression and NO production, which collectively indicate that
Rb1 plays a protective role against
H2O2-induced injury of HUVEC via the
SIRT1/AMPK pathway.
Discussion
Our previous studies (16,36,37)
demonstrated that endothelial senescence and dysfunction in HUVEC
were characterized by enhanced H2O2-induced
β-galactosidase activity and impaired anti-oxidant capacity. This
was associated with an increased protein expression of PAI-1,
decreased NO production and eNOS expression in HUVEC. The present
study identified that Ginsenoside Rb1 protects against cell
senescence and dysfunction through activation of Sirt1. However,
the involved molecular mechanisms have yet to be elucidated. In
line with our previous studies (16,36,37),
the present study demonstrated that Rb1 significantly increased NO
content, eNOS expression and Sirt1 expression in
H2O2-induced HUVEC senescence. In addition,
it was identified for the first time, to the best of the authors'
knowledge, that these changes were associated with upregulated
phosphorylation of AMPK. Compound C, an inhibitor of AMPK, was used
to clarify the relation of AMPK and Sirt1. It was identified that
treatment with compound C markedly attenuated the protective
effects of Rb1 in HUVECs. Furthermore, the role of SIRT1/AMPK
pathway was further confirmed by administration of nicotinamide,
which enhanced H2O2-induced senescence and
dysfunction and decreased the protective effect of Rb1 on HUVEC.
Collectively, the results provide novel mechanisms for Rb1
protected H2O2-induced HUVEC senescence
involving SIRT1/AMPK pathway.
A number of studies have reported that Rb1, a main
constituent of the root of P. ginseng, has various
pharmacological effects that include mitigating endothelial
inflammation and obesity in vitro and in vivo. Zhou
et al (46) reported that
Rb1 protects HUVECs from TNF-α-induced oxidative stress and
inflammation by inhibiting NF-κB, JNK and p38 signaling pathways.
Yuan et al (47) reported
that Rb1 protects pulmonary microvascular endothelial cells from
lipopolysaccharide-induced expression of inflammatory cytokines.
Our previous studies demonstrated that treatment with Rb1 at the
concentration of 20 µM significantly reduces
H2O2-induced cell senescence and inhibits
PAI-1 expression (37) partly
through stimulation of SIRT1 (36). The present study demonstrated that
Rb1 at the concentration of 10–20 µM played a beneficial role on
H2O2-induced endothelial dysfunction, which
complements our previous research. However, the molecular
mechanisms need further elucidation.
Previous studies have suggested a critical role of
AMPK in stabilizing endothelial functions through regulation of
eNOS signaling to suppress inflammation and oxidative stress
(48–51). In addition, there are some studies
on the effects of AMPK activators on the expression of eNOS,
suggesting that AMPK acts as an eNOS activator (52–54).
Thus far, there has been a lack of studies from the present authors
concerning the role of AMPK in the regulation of
H2O2-induced oxidative stress and
inflammatory responses by Rb1 in HUVECs. However, the present study
explored the use of compound C to inhibit AMPK and determined
whether Rb1 upregulated eNOS expression and NO production through
AMPK. As a result, it was identified that AMPK activity was
enhanced by Rb1. The present study not only confirmed that the
expression and activity of eNOS suppressed by
H2O2 stimulation was enhanced by Rb1
treatment, which is in accordance with previous findings, but also
identified that the beneficial effects of Rb1 were abolished by
compound C, indicating a novel role of AMPK in regulating
Rb1-dependent eNOS activity and NO synthesis. However, the
mechanism of Rb1 in AMPK-dependent eNOS activation and NO
production in HUVECs requires further investigation.
SIRT1 is highly sensitive to cellular redox states
and considered to have a cardioprotective effect (55,56)
that maintains endothelial functions by counteracting the effects
of reactive oxygen species as a NAD+-dependent class III
histone deacetylase (57). SIRT1
has been reported to regulate the functions of several important
transcription factors with anti-inflammatory effects (58–60).
It antagonizes H2O2-induced premature
senescence via negative modulation of p53 by deacetylation of
Lys-373, Lys-382 and Lys-320 in the human endothelium (61). Ota et al (62,63)
demonstrated that the overexpression of SIRT1 in the endothelium
reverses H2O2-induced premature cellular
senescence through an eNOS-dependent signaling pathway. The present
study also confirmed that inhibition of SIRT1 by nicotinamide
caused a stress-induced increase in PAI-1 with a concomitant
decrease in eNOS expression and NO production, which are consistent
with Cacicedo et al (64).
Furthermore, the present study demonstrated that Rb1 induced AMPK
phosphorylation, which was abolished by nicotinamide. Previous
studies have reported the functional connections between the two
master regulators SIRT1 and AMPK: Gao et al (65) reported that activated SIRT1
functionally interacts with AMPKα and upregulates its
phosphorylation in aortic endothelial and smooth muscle cells of
Klotho knockout mice and other studies have shown that AMPK can
also function as a SIRT1 activator by increasing the
NAD+/NADH ratio (66,67).
These studies suggest that AMPK may crosstalk with SIRT1 to
modulate downstream targets. The present study demonstrated that
inhibition of SIRT1 by nicotinamide functionally suppressed the
phosphorylation of AMPK, which in turn abolished the protective
effect against Rb1 on H2O2-induced
endothelial dysfunction. However, in vivo experiments are
required to confirm the anti-aging and antioxidant effects of Rb1
demonstrated in vitro. Further research is also required to
elucidate the complex molecular mechanisms underlying the
involvement of AMPK and SIRT1 in anti-aging and antioxidant
processes.
In conclusion, Rb1 improved
H2O2-induced HUVEC senescence and dysfunction
through the SIRT1/AMPK signaling pathway. The results of the
present study suggested that the associated mechanisms may be
related to decreased PAI-1 expression and upregulated eNOS
expression and NO production. The present study provides evidence
to support the novel role of AMPK in the beneficial effects of Rb1
on HUVEC senescence, which can be explored further in animal models
and clinical studies.
Acknowledgements
The authors would like to thank Mr Mitchell Arico
for editing the English text of a draft of this manuscript.
Funding
This study was supported by the grants from the
National Natural Science Foundation of China (grant nos. 81370447;
81300707); Science and Technology Planning Project of Guangdong
Province (grant no. 2016A050502014). The funders had no role in
study design, data collection and analysis, decision to publish, or
preparation of the manuscript.
Availability of data and materials
The datasets used and/or analyzed during the
present study are available from the corresponding author on
reasonable request.
Authors' contributions
ZZ, JZ and XQ designed the study. ZZ, MW, CC, DL
and LW conducted the research. ZZ and MW analyzed the data and
wrote the manuscript. ZZ and MW revised the discussion section of
the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The isolation procedure for HUVECs was approved by
the Research Committee at the Third Affiliated Hospital of Sun
Yat-sen University. The donors provided written informed consent to
donate the umbilical cords.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Quyyumi AA: Endothelial function in health
and disease: New insights into the genesis of cardiovascular
disease. Am J Med. 105:32S–39S. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Minamino T, Miyauchi H, Yoshida T, Ishida
Y, Yoshida H and Komuro I: Endothelial cell senescence in human
atherosclerosis: Role of telomere in endothelial dysfunction.
Circulation. 105:1541–1544. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Barton M: Obesity and aging: Determinants
of endothelial cell dysfunction and atherosclerosis. Pflugers Arch.
460:825–837. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ota H, Eto M, Kano MR, Kahyo T, Setou M,
Ogawa S, Iijima K, Akishita M and Ouchi Y: Induction of endothelial
nitric oxide synthase, SIRT1, and catalase by statins inhibits
endothelial senescence through the Akt pathway. Arterioscler Thromb
Vasc Biol. 30:2205–2211. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ruan Y, Wu S, Zhang L, Chen G and Lai W:
Retarding the senescence of human vascular endothelial cells
induced by hydrogen peroxide: Effects of 17beta-estradiol (E2)
mediated mitochondria protection. Biogerontology. 15:367–375. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cai H: Hydrogen peroxide regulation of
endothelial function: Origins, mechanisms, and consequences.
Cardiovasc Res. 68:26–36. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Han X, Tai H, Wang X, Wang Z, Zhou J, Wei
X, Ding Y, Gong H, Mo C, Zhang J, et al: AMPK activation protects
cells from oxidative stress-induced senescence via autophagic flux
restoration and intracellular NAD(+) elevation. Aging Cell.
15:416–427. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Conti V, Corbi G, Simeon V, Russomanno G,
Manzo V, Ferrara N and Filippelli A: Aging-related changes in
oxidative stress response of human endothelial cells. Aging Clin
Exp Res. 27:547–553. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
El Assar M, Angulo J and Rodriguez-Manas
L: Oxidative stress and vascular inflammation in aging. Free Radic
Biol Med. 65:380–401. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sindler AL, Delp MD, Reyes R, Wu G and
Muller-Delp JM: Effects of ageing and exercise training on eNOS
uncoupling in skeletal muscle resistance arterioles. J Physiol.
587:3885–3897. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kang LS, Reyes RA and Muller-Delp JM:
Aging impairs flow-induced dilation in coronary arterioles: Role of
NO and H(2)O(2). Am J Physiol Heart Circ Physiol. 297:H1087–H1095.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xiong Y, Shen L, Liu KJ, Tso P, Xiong Y,
Wang G, Woods SC and Liu M: Antiobesity and antihyperglycemic
effects of ginsenoside Rb1 in rats. Diabetes. 59:2505–2512. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen S, Li X, Wang Y, Mu P, Chen C, Huang
P and Liu D: Ginsenoside Rb1 attenuates intestinal
ischemia/reperfusion-induced inflammation and oxidative stress via
activation of the PI3K/Akt/Nrf2 signaling pathway. Mol Med Rep.
19:3633–3641. 2019.PubMed/NCBI
|
|
14
|
Sun Q, Meng QT, Jiang Y, Liu HM, Lei SQ,
Su WT, Duan WN, Wu Y and Xia ZY and Xia ZY: Protective effect of
ginsenoside Rb1 against intestinal ischemia-reperfusion induced
acute renal injury in mice. PLoS One. 8:e808592013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang M, Chen Y, Xiong Z, Yu S, Zhou B,
Ling Y, Zheng Z, Shi G, Wu Y and Qian X: Ginsenoside Rb1 inhibits
free fatty acidsinduced oxidative stress and inflammation in 3T3-L1
adipocytes. Mol Med Rep. 16:9165–9172. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu DH, Chen YM, Liu Y, Hao BS, Zhou B, Wu
L, Wang M, Chen L, Wu WK and Qian XX: Rb1 protects endothelial
cells from hydrogen peroxide-induced cell senescence by modulating
redox status. Biol Pharm Bull. 34:1072–1077. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Herzig S and Shaw RJ: AMPK: Guardian of
metabolism and mitochondrial homeostasis. Nat Rev Mol Cell Biol.
19:121–135. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang M, Wang S, Pan Z, Ou T, Ma J, Liu H,
Li R, Yang P, Han W, Guan S, et al: AMPK/NF-KB signaling pathway
regulated by ghrelin participates in the regulation of HUVEC and
THP1 Inflammation. Mol Cell Biochem. 437:45–53. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Thornton CC, Al-Rashed F, Calay D, Birdsey
GM, Bauer A, Mylroie H, Morley BJ, Randi AM, Haskard DO, Boyle JJ
and Mason JC: Methotrexate-mediated activation of an
AMPK-CREB-dependent pathway: A novel mechanism for vascular
protection in chronic systemic inflammation. Ann Rheum Dis.
75:439–448. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Martinez-Martin N, Blas-Garcia A, Morales
JM, Marti-Cabrera M, Monleon D and Apostolova N: Metabolomics of
the effect of AMPK activation by AICAR on human umbilical vein
endothelial cells. Int J Mol Med. 29:88–94. 2012.PubMed/NCBI
|
|
21
|
Huang NL, Chiang SH, Hsueh CH, Liang YJ,
Chen YJ and Lai LP: Metformin inhibits TNF-alpha-induced IkappaB
kinase phosphorylation, IkappaB-alpha degradation and IL-6
production in endothelial cells through PI3K-dependent AMPK
phosphorylation. Int J Cardiol. 134:169–175. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nagata D, Mogi M and Walsh K:
AMP-activated protein kinase (AMPK) signaling in endothelial cells
is essential for angiogenesis in response to hypoxic stress. J Biol
Chem. 278:31000–31006. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ido Y, Carling D and Ruderman N:
Hyperglycemia-induced apoptosis in human umbilical vein endothelial
cells: Inhibition by the AMP-activated protein kinase activation.
Diabetes. 51:159–167. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhou H, Fu B, Xu B, Mi X, Li G, Ma C, Xie
J, Li J and Wang Z: Rosmarinic acid alleviates the endothelial
dysfunction induced by hydrogen peroxide in rat aortic rings via
activation of AMPK. Oxid Med Cell Longev. 2017:70919042017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zaborska KE, Edwards G, Austin C and
Wareing M: The role of O-GlcNAcylation in perivascular adipose
tissue dysfunction of offspring of high-fat diet-fed rats. J Vasc
Res. 54:79–91. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Baudin B, Bruneel A, Bosselut N and
Vaubourdolle M: A protocol for isolation and culture of human
umbilical vein endothelial cells. Nat Protoc. 2:481–485. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Guo Y, Xing L, Qian C, Ding Z and Jin B:
Involvement of flavonoids from the leaves of carya cathayensis
sarg. In sirtuin 1 expression in HUVEC senescence. Evid Based
Complement Alternat Med. 2018:82465602018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Esfahanian N, Shakiba Y, Nikbin B, Soraya
H, Maleki-Dizaji N, Ghazi-Khansari M and Garjani A: Effect of
metformin on the proliferation, migration, and MMP-2 and −9
expression of human umbilical vein endothelial cells. Mol Med Rep.
5:1068–1074. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zheng S, Li W, Xu M, Bai X, Zhou Z, Han J,
Shyy JY and Wang X: Calcitonin gene-related peptide promotes
angiogenesis via AMP-activated protein kinase. Am J Physiol Cell
Physiol. 299:C1485–C1492. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Green LC, Wagner DA, Glogowski J, Skipper
PL, Wishnok JS and Tannenbaum SR: Analysis of nitrate, nitrite, and
[15N]nitrate in biological fluids. Anal Biochem. 126:131–138. 1982.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Debacq-Chainiaux F, Erusalimsky JD,
Campisi J and Toussaint O: Protocols to detect
senescence-associated beta-galactosidase (SA-betagal) activity, a
biomarker of senescent cells in culture and in vivo. Nat Protoc.
4:1798–1806. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Luo W, Wang Y, Yang H, Dai C, Hong H, Li
J, Liu Z, Guo Z, Chen X, He P, et al: Heme oxygenase-1 ameliorates
oxidative stress-induced endothelial senescence via regulating
endothelial nitric oxide synthase activation and coupling. Aging
(Albany NY). 10:1722–1744. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yan J, Wang J, Huang H, Huang Y, Mi T,
Zhang C and Zhang L: Fibroblast growth factor 21 delayed
endothelial replicative senescence and protected cells from
H2O2-induced premature senescence through
SIRT1. Am J Transl Res. 9:4492–4501. 2017.PubMed/NCBI
|
|
34
|
Lin XL, Liu Y, Liu M, Hu H, Pan Y, Fan XJ,
Hu XM and Zou WW: Inhibition of hydrogen peroxide-induced human
umbilical vein endothelial cells aging by allicin depends on
sirtuin1 activation. Med Sci Monit. 23:563–570. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hahn HJ, Kim KB, An IS, Ahn KJ and Han HJ:
Protective effects of rosmarinic acid against hydrogen
peroxide-induced cellular senescence and the inflammatory response
in normal human dermal fibroblasts. Mol Med Rep. 16:9763–9769.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Song Z, Liu Y, Hao B, Yu S, Zhang H, Liu
D, Zhou B, Wu L, Wang M, Xiong Z, et al: Ginsenoside Rb1 prevents
H2O2-induced HUVEC senescence by stimulating sirtuin-1 pathway.
PLoS One. 9:e1126992014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu DH, Chen YM, Liu Y, Hao BS, Zhou B, Wu
L, Wang M, Chen L, Wu WK and Qian XX: Ginsenoside Rb1 reverses
H2O2-induced senescence in human umbilical endothelial cells:
Involvement of eNOS pathway. J Cardiovasc Pharmacol. 59:222–230.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Erusalimsky JD and Kurz DJ: Cellular
senescence in vivo: Its relevance in ageing and cardiovascular
disease. Exp Gerontol. 40:634–642. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Shan HY, Bai XJ and Chen XM: Apoptosis is
involved in the senescence of endothelial cells induced by
angiotensin II. Cell Biol Int. 32:264–270. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li CX, Gao JG, Wan XY, Chen Y, Xu CF, Feng
ZM, Zeng H, Lin YM, Ma H, Xu P, et al: Allyl isothiocyanate
ameliorates lipid accumulation and inflammation in nonalcoholic
fatty liver disease via the Sirt1/AMPK and NF-KB signaling
pathways. World J Gastroenterol. 25:5120–5133. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang T, Gu J, Wu PF, Wang F, Xiong Z, Yang
YJ, Wu WN, Dong LD and Chen JG: Protection by tetrahydroxystilbene
glucoside against cerebral ischemia: Involvement of JNK, SIRT1, and
NF-kappaB pathways and inhibition of intracellular ROS/RNS
generation. Free Radic Biol Med. 47:229–240. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lee JH, Song MY, Song EK, Kim EK, Moon WS,
Han MK, Park JW, Kwon KB and Park BH: Overexpression of SIRT1
protects pancreatic beta-cells against cytokine toxicity by
suppressing the nuclear factor-kappaB signaling pathway. Diabetes.
58:344–351. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Huang K, Li R and Wei W: Sirt1 activation
prevents anti-Thy 1.1 mesangial proliferative glomerulonephritis in
the rat through the Nrf2/ARE pathway. Eur J Pharmacol. 832:138–144.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Huang K, Chen C, Hao J, Huang J, Wang S,
Liu P and Huang H: Polydatin promotes Nrf2-ARE anti-oxidative
pathway through activating Sirt1 to resist AGEs-induced
upregulation of fibronetin and transforming growth factor-β1 in rat
glomerular messangial cells. Mol Cell Endocrinol. 399:178–189.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chen AF, Chen DD, Daiber A, Faraci FM, Li
H, Rembold CM and Laher I: Free radical biology of the
cardiovascular system. Clin Sci (Lond). 123:73–91. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhou P, Lu S, Luo Y, Wang S, Yang K, Zhai
Y, Sun G and Sun X: Attenuation of TNF-α-induced inflammatory
injury in endothelial cells by ginsenoside Rb1 via inhibiting
NF-KB, JNK and p38 signaling pathways. Front Pharmacol. 8:4642017.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yuan Q, Jiang YW, Ma TT, Fang QH and Pan
L: Attenuating effect of Ginsenoside Rb1 on LPS-induced lung injury
in rats. J Inflamm (Lond). 11:402014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Hwang HJ, Jung TW, Kim JW, Kim JA, Lee YB,
Hong SH, Roh E, Choi KM, Baik SH and Yoo HJ: Protectin DX prevents
H2O2-mediated oxidative stress in vascular
endothelial cells via an AMPK-dependent mechanism. Cell Signal.
53:14–21. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wang L, Han J, Shan P, You S, Chen X, Jin
Y, Wang J, Huang W, Wang Y and Liang G: MD2 blockage protects
obesity-induced vascular remodeling via activating AMPK/Nrf2.
obesity (Silver Spring). 25:1532–1539. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Lu Q, Mundy M, Chambers E, Lange T, Newton
J, Borgas D, Yao H, Choudhary G, Basak R, Oldham M and Rounds S:
Alda-1 protects against acrolein-induced acute lung injury and
endothelial barrier dysfunction. Am J Respir Cell Mol Biol.
57:662–673. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kroller-Schon S, Jansen T, Hauptmann F,
Schüler A, Heeren T, Hausding M, Oelze M, Viollet B, Keaney JF Jr,
Wenzel P, et al: α1AMP-activated protein kinase mediates vascular
protective effects of exercise. Arterioscler Thromb Vasc Biol.
32:1632–1641. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Morrow VA, Foufelle F, Connell JM, Petrie
JR, Gould GW and Salt IP: Direct activation of AMP-activated
protein kinase stimulates nitric-oxide synthesis in human aortic
endothelial cells. J Biol Chem. 278:31629–31639. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Gaskin FS, Kamada K, Yusof M and Korthuis
RJ: 5′-AMP-activated protein kinase activation prevents
postischemic leukocyte-endothelial cell adhesive interactions. Am J
Physiol Heart Circ Physiol. 292:H326–H332. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Han F, Zhang S, Hou N, Wang D and Sun X:
Irisin improves endothelial function in obese mice through the
AMPK-eNOS pathway. Am J Physiol Heart Circ Physiol.
309:H1501–H1508. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Winnik S, Auwerx J, Sinclair DA and Matter
CM: Protective effects of sirtuins in cardiovascular diseases: From
bench to bedside. Eur Heart J. 36:3404–3412. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Stein S and Matter CM: Protective roles of
SIRT1 in atherosclerosis. Cell Cycle. 10:640–647. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Imai S, Armstrong CM, Kaeberlein M and
Guarente L: Transcriptional silencing and longevity protein Sir2 is
an NAD-dependent histone deacetylase. Nature. 403:795–800. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Yuan Q, Zhang D, Liu C, Zhang C and Yuan
D: Chikusetsusaponin V inhibits LPS-activated inflammatory
responses via SIRT1/NF-KB signaling pathway in RAW264.7 cells.
Inflammation. 41:2149–2159. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Qian Y, Chen C, Ma L, Wang Z, Wang LF, Zuo
L, Yang Y, Huang X, Jiang M, Wang X, et al: CD38 deficiency
promotes inflammatory response through activating
Sirt1/NF-κB-mediated inhibition of TLR2 expression in macrophages.
Mediators Inflamm. 2018:87369492018. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Li M, Hong W, Hao C, Li L, Wu D, Shen A,
Lu J, Zheng Y, Li P and Xu Y: SIRT1 antagonizes liver fibrosis by
blocking hepatic stellate cell activation in mice. FASEB J.
32:500–511. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Kao CL, Chen LK, Chang YL, Yung MC, Hsu
CC, Chen YC, Lo WL, Chen SJ, Ku HH and Hwang SJ: Resveratrol
protects human endothelium from H(2)O(2)-induced oxidative stress
and senescence via SirT1 activation. J Atheroscler Thromb.
17:970–979. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Ota H, Akishita M, Eto M, Iijima K, Kaneki
M and Ouchi Y: Sirt1 modulates premature senescence-like phenotype
in human endothelial cells. J Mol Cell Cardiol. 43:571–579. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Ota H, Eto M, Kano MR, Ogawa S, Iijima K,
Akishita M and Ouchi Y: Cilostazol inhibits oxidative
stress-induced premature senescence via upregulation of Sirt1 in
human endothelial cells. Arterioscler Thromb Vasc Biol.
28:1634–1639. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Cacicedo JM, Gauthier MS, Lebrasseur NK,
Jasuja R, Ruderman NB and Ido Y: Acute exercise activates AMPK and
eNOS in the mouse aorta. Am J Physiol Heart Circ Physiol.
301:H1255–H1265. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Gao D, Zuo Z, Tian J, Ali Q, Lin Y, Lei H
and Sun Z: Activation of SIRT1 attenuates klotho deficiency-induced
arterial stiffness and hypertension by enhancing AMP-activated
protein kinase activity. Hypertension. 68:1191–1199. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Canto C, Gerhart-Hines Z, Feige JN,
Lagouge M, Noriega L, Milne JC, Elliott PJ, Puigserver P and Auwerx
J: AMPK regulates energy expenditure by modulating NAD+ metabolism
and SIRT1 activity. Nature. 458:1056–1060. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Chau MD, Gao J, Yang Q, Wu Z and Gromada
J: Fibroblast growth factor 21 regulates energy metabolism by
activating the AMPK-SIRT1-PGC-1alpha pathway. Proc Natl Acad Sci
USA. 107:12553–12558. 2010. View Article : Google Scholar : PubMed/NCBI
|