Introduction
According to a 2017 United Nations report on the
aging of the world population (1),
the number of people aged ≥60 years worldwide will more than double
by 2050, reaching ~2.1 billion (2–4).
Aging is an independent risk factor for the incidence and
prevalence of cardiovascular diseases, such as hypertension and
atherosclerosis, which can lead to heart failure (5). Changes in the aging heart are mainly
associated with structural and functional abnormalities of the
cardiac muscle cells, including increased left ventricular wall
thickness, cardiac fibrosis and impaired diastolic function
(6,7). However, the molecular mechanisms
underlying the decline in cardiac function during aging remain
unknown.
Renin-angiotensin system (RAAS) activation is
closely associated with the aging process and the etiology of
cardiac diseases. Mineralocorticoid receptors (MR) are an important
component of RAAS. The activated MR signal induced by a high salt
load leads to fibrosis and cardiac dysfunction, independent of
renal effects and elevated blood pressure (3). MR antagonists decrease morbidity and
mortality in patients with heart failure; however, hyperkalemia,
gynecomastia and erectile dysfunction limit their widespread use
(8). In addition,
cardiomyocyte-specific MR inactivation improves infarct healing,
and prevents progressive adverse cardiac remodeling and contractile
dysfunction (9). Identifying the
cardiac-specific features of MR signaling may provide a basis for
improving cardiovascular conditions without compromising renal
function.
Local myocardial RAAS activation is involved in
age-associated cardiac remodeling. However, to the best of our
knowledge, the dynamic expression of MR during the aging process
has not been characterized. The aim of the present study was to
investigate changes in MR expression in the aging heart, and to
identify the relationship between these changes and oxidative
stress and mitochondrial abnormalities.
Materials and methods
Experimental reagents
Rabbit primary polyclonal MR antibody (cat. no.
21854-1-AP) was obtained from ProteinTech Group, Inc. Mouse
monoclonal antibodies against GAPDH (cat. no. sc-59540), p53 (cat.
no. sc-71819), p16 (cat. no. sc-377412), p21 (cat. no. sc-817),
peroxisome proliferator-activated receptor γ coactivator-1α
(PGC-1α; cat. no. sc-518025), transforming growth factor-β1
(TGF-β1; cat. no. sc-130348), gp91-phox (cat. no. sc-130543),
p47-phox (cat. no. sc-17845), p67-phox (cat. no. sc-374510),
superoxide dismutase (SOD)-1 (cat. no. sc-101523) and SOD-2 (cat.
no. sc-133134) were obtained from Santa Cruz Biotechnology, Inc. A
horseradish peroxidase-conjugated goat anti-rabbit secondary
antibody (cat. no. BA1056) and goat anti-mouse antibody (cat. no.
BA1075) were obtained from Wuhan Boster Biological Technology, Ltd.
Eplerenone (cat. no. HY-B0251) was purchased from MedChemExpress.
All other chemicals and reagents were purchased from Sigma-Aldrich
(Merck KGaA), unless otherwise specified.
Animals
All animals were fed normal chow and housed at 22°C
and ~40-70% humidity under a 12-h light/dark cycle with free access
to adequate food and water. Animal health and behavior were
monitored once a week. All experimental procedures were approved by
the Animal Research Ethics Board at Huazhong University of Science
and Technology, and conformed to the Update of the Guide for the
Care and Use of Laboratory Animals published by the National
Research Council (US) Committee. All animal studies also complied
with the ARRIVE guidelines (10)
and the AVMA euthanasia guidelines (11).
In total, 16 male Sprague-Dawley rats (Hubei
Provincial Animal Research Center, Hubei Provincial Center for
Disease Control and Prevention) were divided into two groups (n=8
in each group): Young (age, 3 months; weight, 250–350 g) and old
(age, 24 months; weight, 650–750 g). At the designated age of 3 or
24 months, rats were intraperitoneally anesthetized with 3%
pentobarbital sodium (35–40 mg/kg body weight; Nembutal; Sumitomo
Dainippon Pharma Co., Ltd.) and were then euthanized by cervical
dislocation following the collection of blood samples. Animals
suffering chronic diseases, such as tumors, respiratory failure and
heart failure, as characterized by weight loss, a change in vital
signs such as heart rate, breathing or hypothermia, and abnormal
behavior, were immediately euthanized by cervical dislocation
following anesthesia with 3% pentobarbital sodium (~35–40 mg/kg
body weight) to relieve or end suffering in advance. The intestinal
tumor of the rat (age, 18 months; weight, 550 g) in the old group
weighed 6.2 g and the maximum tumor burden was ~1% (tumor
weight/body weight). This animal displayed reduced consumption of
food and water, and a subcutaneous mass in the abdomen. Cachexia
index was ~2.9% of body weight, calculated according to the
equation: [(Initial body weight-final body mass + mass of the tumor
+ control weight gain)/(initial body mass + mass gain control
group)] ×100 (12,13). Rats in the old group (age, 18
months old) showing signs of distress were sacrificed and their
tissues were not used for subsequent experiments. Subsequently
their hearts were excised, washed with saline, cleaned and weighed.
Large vessels and connective tissues were removed, and the left
ventricular cardiac tissues were stored at −80°C until subsequent
experiments.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from the left ventricle
using TRIzol® reagent (cat. no. 15596026; Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. First-strand cDNA was synthesized with 1 µg RNA using a
cDNA Synthesis kit (cat. no. K1622; Thermo Fisher Scientific, Inc.)
as previously described (14). The
primers were designed using Primer 5.0 (Premier Biosoft
International) and confirmed by BLAST
(blast.ncbi.nlm.nih.gov/Blast.cgi) searches against the GenBank
database (www.ncbi.nlm.nih.gov/genbank). The sequences of
primers were as follows: Nr3c2, forward
5′-AGTGGGGTCCATCGGCAG-3′, reverse 5′-CCTCTGTCTTAGGGAAAGGAACG-3′;
and β-actin, forward 5′-AGTGTGACGTTGACATCCGT-3′ and reverse
5′-GACTCATCGTACTCCTGCTT-3′. A SYBR Green-based master mix (cat no.
A25742; Thermo Fisher Scientific, Inc.) was used to amplify the
cDNA using an ABI Prism 7700 sequence detection system (Thermo
Fisher Scientific, Inc.). Thermocycling conditions were as follows:
40 cycles of 95°C for 15 sec and 60°C for 1 min. The results were
analyzed using the 2−∆∆Cq method (14).
Western blot analysis
The heart tissue homogenate was centrifuged at
14,000 × g for 15 min at 4°C and resolved in RIPA lysis buffer
(Beyotime Institute of Biotechnology) containing a protease
inhibitor cocktail. Protein concentration was determined using the
bicinchoninic acid assay kit (Beyotime Biotechnology, Wuhan).
Loading volume of each sample was calculated in order to load 80 µg
protein per lane. After protein quantification, appropriate amount
of 6× sample loading buffer was added, and samples were boiled at
95°C for 5 min. The suspension was separated by 10% SDS-PAGE and
transferred onto a PVDF membrane. The membranes were blocked using
5% BSA for 1.5 h at room temperature. The membranes were incubated
overnight at 4°C with primary antibodies against MR (ProteinTech
Group, Inc.), and p16, p21, p53, PGC-1α, SOD-1, TGF-β1, gp91-phox,
p47-phox, p67-phox and SOD-2 (Santa Cruz Biotechnology, Inc.), at a
final dilution of 1:1,000. The membranes were then incubated for 2
h at room temperature (27°C) with a horseradish
peroxidase-conjugated secondary antibody (cat. no. BA1056; Wuhan
Boster Biological Technology, Ltd.), diluted to 1:10,000. Following
exposure using a Luminescent Imaging system (Tanon Science and
Technology Co., Ltd.), blots were washed and re-incubated with
monoclonal GAPDH antibody as a control. Protein expression levels
were normalized to the corresponding GAPDH levels, as determined
using Gel-Pro Analyzer 4 (Media Cybernetics, Inc.).
Hemodynamic measurements and
echocardiography
Body weight was recorded during the experiments.
Blood pressure measurements were obtained at room temperature using
a photoelectric tail-cuff system (PowerLab; ADInstruments). The
cardiac function of rats was analyzed using a VIVO 2100
echocardiography machine (VisualSonics, Inc.); for analysis of
cardiac function, sedation of the rats was induced with 4% inhaled
isoflurane (cat. no. R510-22; RWD Life Science Co., Ltd.) and
anesthesia was subsequently maintained with 2% isoflurane for ~8
min (0.2-0.3 l/min). The rats exhibited a steady heart rate
(350–400 beats/min) and breathing, as previously described
(15).
Wheat germ agglutinin (WGA)
fluorescence staining
Fresh-frozen heart sections (8 µm) were washed with
phosphate-buffered saline three times and incubated with
FITC-conjugated WGA (Beyotime Institute of Biotechnology) diluted
to 50 µg/ml for 1 h in the dark. After washing out the dye, images
were obtained using a fluorescence microscope (magnification,
×200). All steps were performed in the dark.
Electron microscopy
Following previously published protocols (16), fresh left ventricle specimens were
fixed in 2.5% glutaraldehyde for 2 h, post-fixed in 1% osmium
tetroxide for 2 h at 4°C, dehydrated, embedded in epoxy resin at
30°C for 4 h and sectioned (30–40 nm). Finally, the sections were
double stained with 0.5% uranyl acetate and 1% lead citrate for 10
min at room temperature. A morphometric analysis was performed to
determine mitochondrial damage using a Tecnai G 12 transmission
electron microscope (FEI; Thermo Fisher Scientific, Inc.), as
previously described (17).
Picrosirius red staining
Heart fibrosis was evaluated by Picrosirius Red
staining. Fresh left ventricular heart tissues were fixed with 4%
paraformaldehyde for 24 h at room temperature, were dehydrated by
an increasing alcohol series and treated with xylene for 1 h at
room temperature. Specimens were embedded in paraffin at 57°C and
cut into 4–5-µm-thick sections. Paraffin-embedded left ventricle
sections (4–5 µm) were dewaxed and stained using a Picrosirius Red
Staining kit (Nanjing Jiancheng Bioengineering Institute) according
to the manufacturer's protocol. Images were obtained using a Nikon
inverted light microscope (Nikon Corporation).
Dihydroethidium (DHE) histological
staining
Fresh-frozen heart specimens were incubated with DHE
diluted to 1:1,000 (cat. no. S0063; Beyotime Institute of
Biotechnology), as previously described (18). Images were obtained using a
fluorescence microscope (magnification, ×200).
Immunohistochemical staining
Fresh left ventricular heart tissues were fixed with
4% paraformaldehyde for 24 h at room temperature, dehydrated by an
increasing alcohol series, and treated with xylen for 1 h.
Specimens were embedded with paraffin at 57°C and cut into
4–5-µ-thick sections. Endogenous peroxidase activity was inhibited
by 0.3% H2O2 solution for 10 min at room
temperature and samples were blocked using 5% BSA for 20 min both
at room temperature. Sections were incubated with an appropriate MR
antibody (1:100 dilution; cat. no. 21854-1-AP; ProteinTech Group,
Inc.) at 4°C overnight. Following application of the biotinylated
secondary antibody at a dilution of 1:100 at room temperature for 1
h (cat. no. BA1003; Wuhan Boster Biological Technology, Ltd.),
samples were developed with DAB substrates. Images were captured
using a Nikon light inverted microscope (Nikon Corporation) and
analyzed using Image-Pro Plus 6 (Media Cybernetics, Inc.).
H9C2 cell culture and treatment
H9C2 rat cardiomyocytes (American Type Culture
Collection) were cultured in DMEM supplemented with 10% BSA (Gibco;
Thermo Fisher Scientific, Inc.), 100 IE/ml penicillin and 100 µg/ml
streptomycin at 37°C in an atmosphere of 5% CO2/95% air.
At 70–80% confluence, H9C2 cells were starved with 5% BSA for 6 h
and then pre-treated with eplerenone (1 µM; cat. no. HY-B0251) at
37°C 1 h before H2O2 (200 µM) was added for 2
h at 37°C. The cells were then harvested for subsequent
experimentation.
Statistical analysis
SPSS (version 19.0; IBM Corp.) was used to analyze
data. All data are presented as the means ± standard error of the
mean. Data from in vivo experiments conforming to the normal
distribution and homogeneity of variances were analyzed by
two-sample t-tests. One way ANOVA was used to evaluate differences
among the four in vitro groups and the LSD t-test was used
for pairwise comparisons between the groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
Effects of aging on cardiac morphology
and hemodynamics
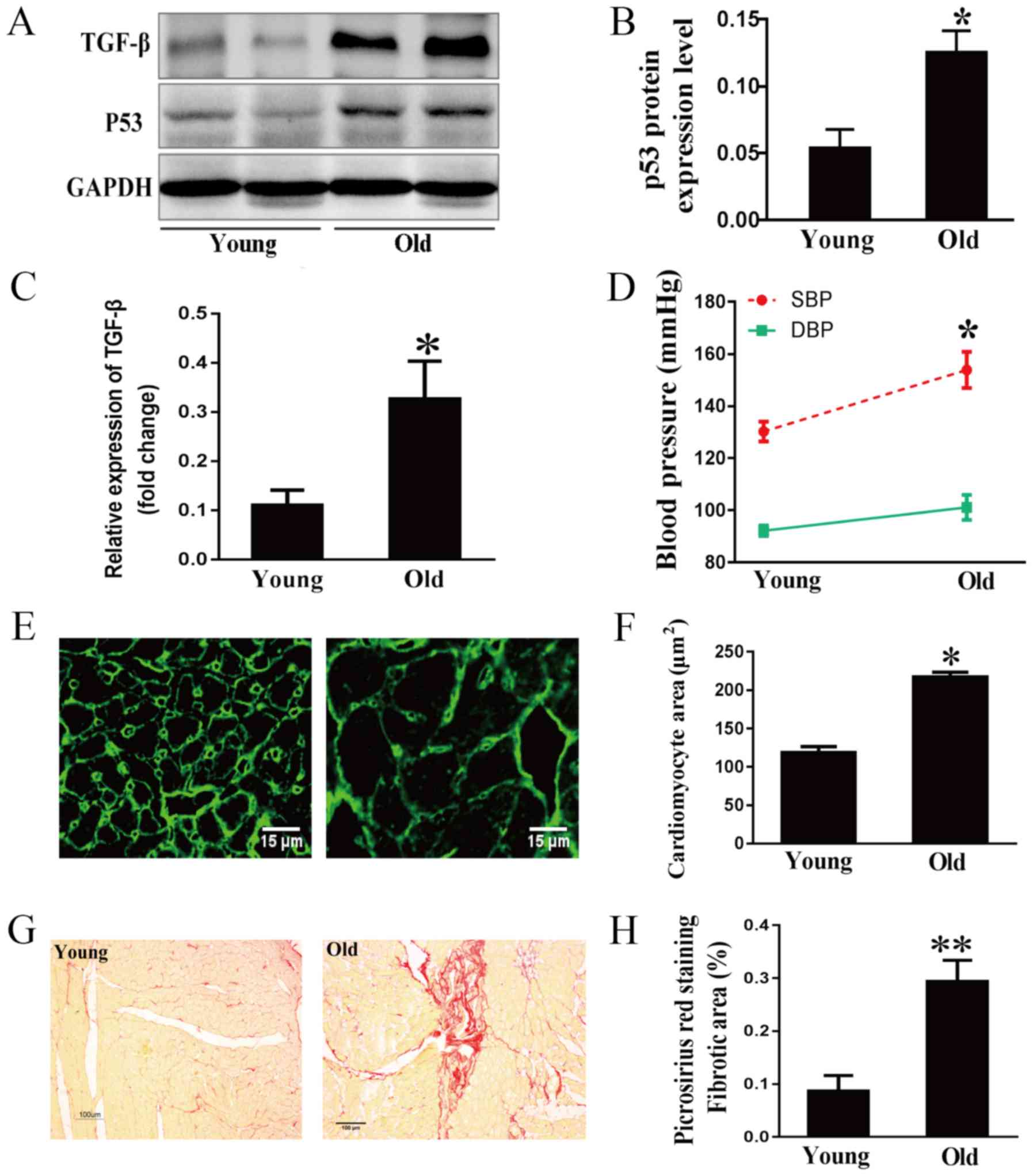
The tumor suppressor p53, an aging marker, and
TGF-β, a marker of extracellular matrix synthesis and degradation,
were examined in rat hearts by western blot analysis (19,20).
As expected, age-related increases in the expression levels of p53
and TGF-β were observed (Fig.
1A-C). Hemodynamic results indicated that the systolic blood
pressure of old rats was much higher than that of young rats,
whereas the difference in diastolic blood pressure between the two
groups was not significant (Fig.
1D). This indicated that the isolated systolic hypertension of
older rats was similar to that of humans (21). To clarify the development of the
aging heart, morphological characteristics were examined. WGA
staining demonstrated that the area of cardiomyocytes was much
greater in the old group compared with in the young group (Fig. 1E and F). Cardiac fibrosis in aging
rats was evaluated by Picrosirius Red staining. As shown in
Fig. 1G and H, collagen
accumulation in the myocardial interstitial and perivascular
regions was markedly increased in the 24-month-old group compared
with in the 3-month-old group.
Aging-related alterations in cardiac
function
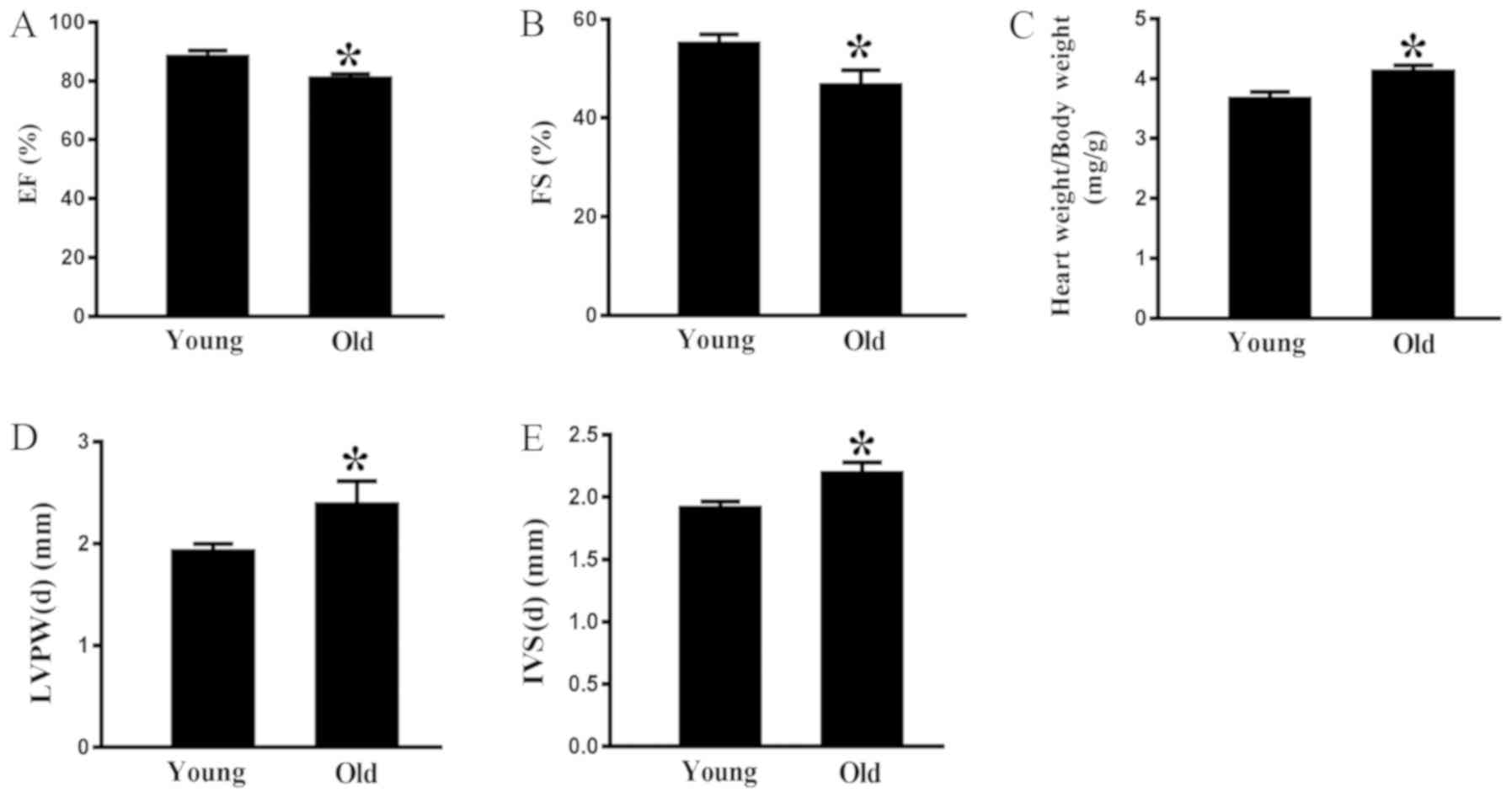
Echocardiography indicated that there was a
significant age-associated decrease in ejection fraction (Fig. 2A) and fractional shortening (FS)
(Fig. 2B) in the old rats compared
with in the young animals. Conversely, the heart weight/body weight
ratio was greater in the rats aged 24 months compared with in the
rats aged 3 months (Fig. 2C). As
shown in Fig. 2D and E, the
interventricular septum at end-diastole (IVS-d) and the left
interventricular posterior wall at end-diastole (LVPW-d) were
thicker in 24-month-old rats compared with in 3-month-old rats.
These data indicated that aging may impair cardiac diastolic
function.
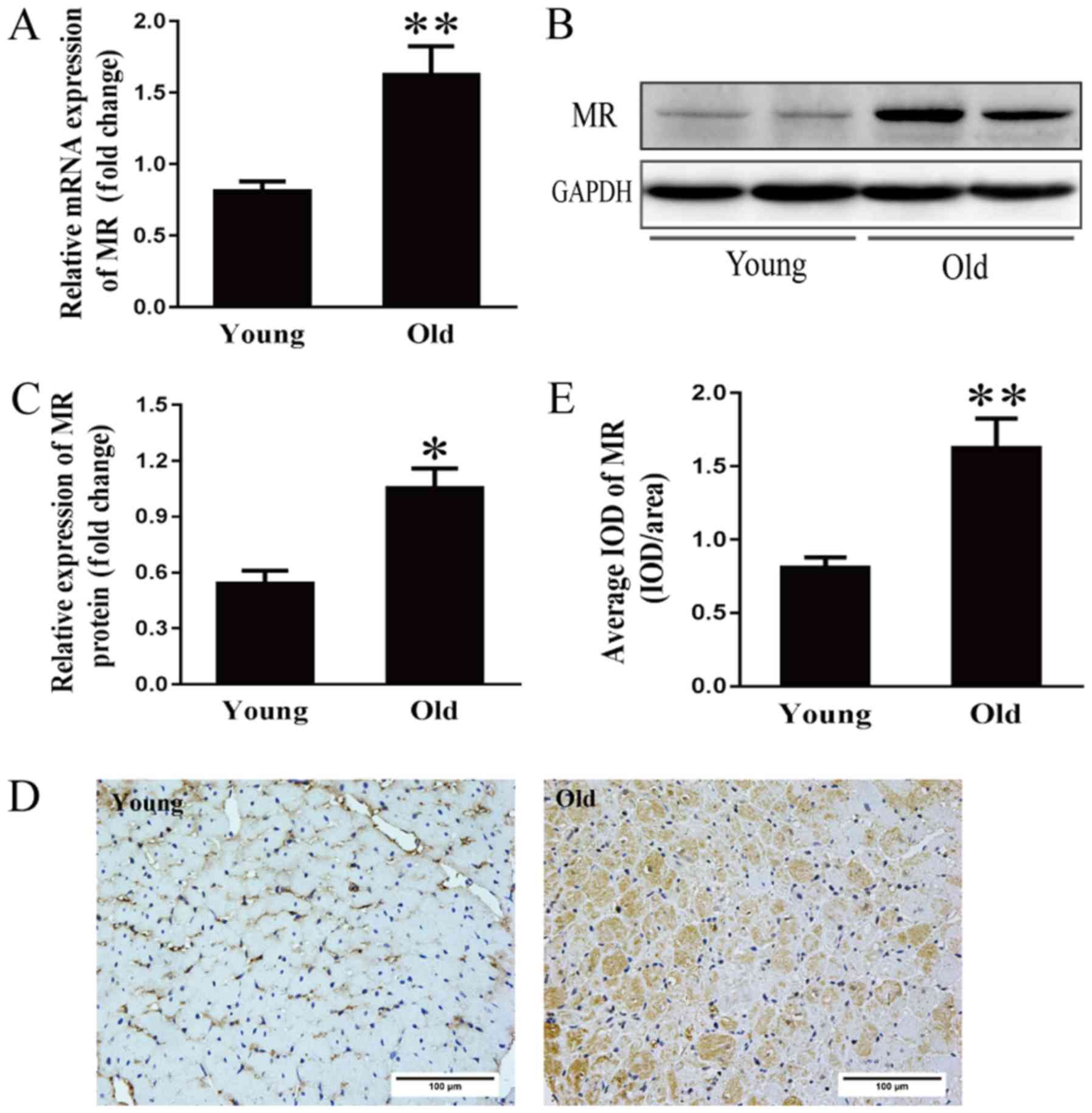
Increased MR expression in the aging
heart
Activation of the renin-angiotensin-aldosterone
system (RAAS) is considered an important cause of hypertension and
cardiac diseases in the aging population (22). However, the mechanism underlying
RAAS activation at the onset and development of cardiomyopathy
during the aging process remains to be elucidated. Therefore, MR
mRNA expression was examined. As shown in Fig. 3A, the mRNA expression of MR was
two-fold higher in the older group compared with in the younger
group. Additionally, MR protein expression in the older hearts was
approximately double that in the younger hearts (Fig. 3B and C). As determined by
immunohistochemical analysis, the average optical density of MR in
the older hearts was more than double that in the younger hearts
(Fig. 3D and E).
Effects of aging on mitochondrial
structure and cardiac function
It has previously been reported that the aging
process is closely related to obvious alterations in mitochondrial
structure (23). In this study,
cardiac ultrastructure was examined by electron microscopy. As
shown in Fig. 4A, myocardial
mitochondria were enlarged, swollen and lacked cristae in
24-month-old rats. PGC-1α drives all aspects of mitochondrial
biogenesis and cardiac function, including mitochondrial number,
mitochondrial respiration and ROS levels (24–27).
Mitochondrial dysfunction induces a profound p53-dependent decrease
in PGC-1α, resulting in impaired oxidative phosphorylation and ATP
generation (25). As expected,
PGC-1α expression was lower in the older group compared with in the
younger group (Fig. 4B and C).
These data suggested that aging may aggravate the impaired
mitochondrial ultrastructure and function.
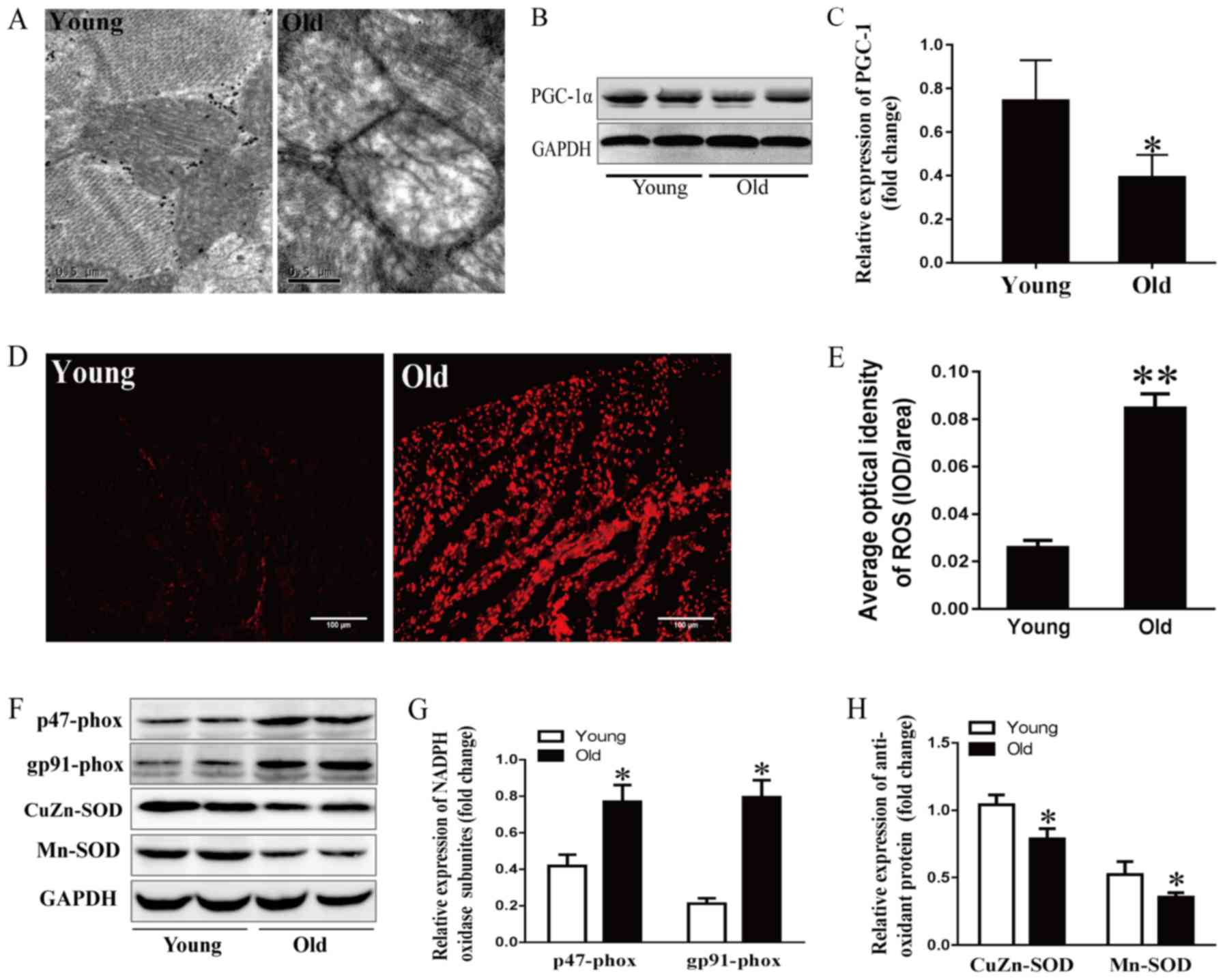 | Figure 4.Effects of aging on mitochondrial
stress and the redox state. (A) Mitochondrial ultrastructure of
myocytes detected by electron microscopy; scale bar, 0.5 µm. (B)
Representative immunoblots and (C) semi-quantitation of PGC-1α. (D)
ROS levels detected by DHE fluorescence staining and (E)
corresponding semi-quantification; scale bar, 50 µm. (F)
Representative immunoblots of p47-phox, gp91-phox, CuZn- and
Mn-SOD. (G) Semi-quantification of p47-phox and gp91-phox. (H)
Semi-quantification of CuZn-SOD and Mn-SOD. Data are presented as
the means ± standard error of the mean from each group; n=8 per
group. *P<0.05, **P<0.01 vs. young group. DHE,
dihydroethidium; NADPH, nicotinamide adenine dinucleotide
phosphate; PGC-1α, peroxisome proliferator-activated receptor γ
coactivator-1α; ROS, reactive oxygen species; SOD, superoxide
dismutase. |
Effects of aging on oxidative stress
and the redox state
DHE fluorescence staining was performed to examine
ROS production in the aging hearts. As shown in Fig. 4D and E, DHE fluorescence intensity
was stronger in 24-month-old rats compared with in the younger
rats, indicating that more ROS was generated and accumulated in the
aging left ventricular myocytes. The tissue levels of ROS are
regulated by ROS generation and antioxidant enzymes. The present
study detected subunits of nicotinamide adenine dinucleotide
phosphate (NADPH) oxidase, including p47-phox and gp91-phox, and
the antioxidant enzyme SOD. As shown in Fig. 4F-H, p47-phox and gp91-phox levels
were higher in the 24-month-old group compared with the younger
group, whereas SOD-1- and SOD-2-levels were lower in the older
group.
MR antagonism suppresses
H2O2-induced cardiac aging and mitochondrial
dysfunction in vitro
To establish the relationship between MR and cardiac
mitochondrial abnormalities, H9C2 cells were treated with
H2O2 to mimic cardiomyocyte aging.
H2O2 treatment increased the expression of
the aging markers p16 and p21 (Fig.
5A). Additionally, H2O2 administration
decreased the expression of p53-dependent PGC-1α and stimulated
excess production of ROS (Fig. 5).
Eplerenone, an MR antagonist, effectively blocked these effects,
illustrating that MR blockage partially alleviated aging-induced
mitochondrial abnormalities. Furthermore, cardiomyocytes induced by
H2O2 demonstrated higher expression levels of
NADPH oxidase subunits and lower antioxidant contents, suggesting
that aging exacerbated the imbalanced redox state. Eplerenone
treatment markedly reversed the upregulation of p47-phox, p67-phox
and gp91-phox, and increased the levels of SOD-1 and SOD-2. These
data suggested that MR antagonism partially protected against aging
via mitochondria dysfunction and oxidative stress.
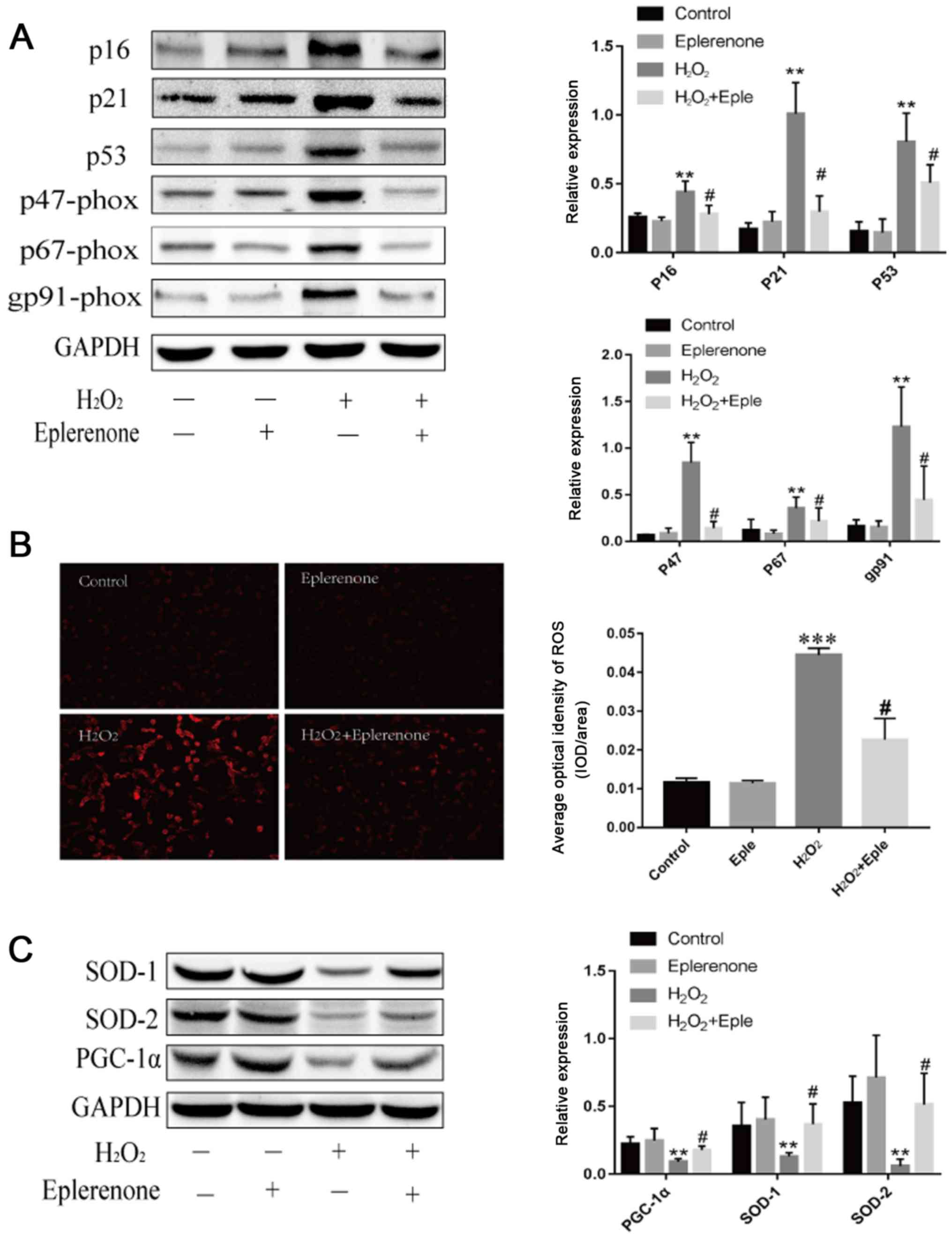 | Figure 5.MR antagonism suppresses
H2O2-induced cardiac aging and mitochondrial
dysfunction. (A) Immunoblotting and semi-quantification of p16,
p21, p53 and NADP subunits, p47-phox, p67-phox and gp91-phox
expression in H9C2 cells. Protein expression levels were normalized
to GAPDH. (B) ROS levels detected by DHE staining in H9C2 cells
following different treatments (magnification, ×100; scale bar, 100
µm). (C) SOD-1, SOD-2 and PGC-1α protein expression levels detected
by western blotting. Data are representative of three experiments,
n=3. **P<0.01 and ***P<0.001 vs. control group;
#P<0.05 vs. H2O2 group. DHE,
dihydroethidium; MR, mineralocorticoid receptors; PGC-1α,
peroxisome proliferator-activated receptor γ coactivator-1α; ROS,
reactive oxygen species; SOD, superoxide dismutase; Eple,
eplerenone. |
Discussion
The results of the present study indicated that the
hearts of aging rats exhibited altered RAAS components,
characterized by enhanced MR expression. The altered expression of
MR was accompanied by cardiac dysfunction, increased fibrosis,
mitochondrial damage and an imbalanced redox state in aging
hearts.
Cardiac aging manifests as increased left
ventricular thickness, enlarged chamber size, increased fibrosis
and diastolic dysfunction (7,28,29).
In the present study, aging induced cardiac remodeling, including
myocyte hypertrophy, interstitial fibrosis and subsequent diastolic
dysfunction, as characterized by decreased FS and increased IVS-d
and LVPW-d. These data suggested that the aging rat model was
sufficient to investigate the potential mechanisms underlying the
role of MR in age-related cardiac dysfunction.
The activation of local RAAS components has been
demonstrated in various tissues. Intrarenal RAAS activation is
associated with the renal aging process (30,31).
Enhanced vascular RAAS activation is associated with arterial aging
(32) and age-related cardiac
remodeling involves myocyte RAAS activation (33). MR signaling is a terminal effector
in RAAS, and exerts more cardiac actions than aldosterone, due to
the absence of 11β hydroxysteroid dehydrogenase type 2 (HSD2) in
the heart, making cortisol (or corticosterone in rodents) the
predominant ligand for MR (34).
Previous research has indicated that the conditional
cardiac-specific overexpression of human MR in mice generates a
high rate of arrhythmia-related sudden death (35). Cardiomyocyte-specific MR deficiency
improves infarct healing, and prevents progressive adverse cardiac
remodeling (9) and
deoxycorticosterone/salt-induced tissue inflammation and remodeling
by blocking the recruitment of macrophages to the myocardium
(3). Loss of
cardiomyocytes-derived MR protects the transaortic constriction
model from cardiac dilatation and failure; however, MR knockout
does not prevent the development of cardiac remodeling and an
inflammatory response after pressure overload (4). In the present study, the protein and
mRNA expression levels of MR in the aging model of fibrosis were
approximately two-fold higher than those in younger rats,
indicating that the increased expression of MR in myocytes of aging
hearts may exacerbate cardiac remodeling and cardiac dysfunction.
MR expression in adult rat hearts (12-month-old) rose moderately
(data not shown), suggesting that cardiomyocyte MR may serve a
universal role during the whole aging process. However, whether MR
accumulation in hearts is elevated in healthy older humans and its
potential clinical implications remain to be investigated.
The p53 tumor suppressor, a cellular stress sensor,
is an aging marker, as evidenced by the early aging phenotypes of
p53-mutant mice (19). PGC-1α is a
master regulator of mitochondrial biogenesis and renewal, and
interacts with p53 by post-transcriptional regulation (24,36).
Aging is characterized by reduced renewal of mitochondria and an
impaired ultrastructure (37,38),
and this is mainly ascribed to decreased PGC-1α activity. The
p53-induced PGC-1α mitochondrial/metabolic axis integrates many
factors involved in the aging process in the heart (25). The present study indicated that
24-month-old rats exhibited damaged mitochondrial architecture and
reduced PGC-1α. It also indicated that the increased MR level in
aging hearts may be associated with impaired mitochondrial
structure and decreased mitochondrial biogenesis renewal via
p53-induced PGC-1α downregulation.
Cellular ROS sources are mainly composed of
mitochondrial and extramitochondrial sources, e.g., membrane NADPH
oxidase. The mitochondrial respiratory chain, particularly complex
I and complex III, is the major cellular source of ROS (39). NADPH oxidases are multi-protein
enzyme complexes that generate superoxide by the catalysis of
electron transfer from NADPH to molecular oxygen (40). Myocytes are rich in mitochondria,
in order to accommodate their high demand for ATP, and mitochondria
are the major targets of free radicals (41,42).
Therefore, mitochondria are vulnerable to ROS and are particularly
susceptible to oxidative stress damage (43). In the present study, mitochondrial
damage and increased cellular ROS accumulation were observed. These
changes may be associated with alterations in MR expression.
The tissue ROS balance is precisely regulated by the
ROS generation system and antioxidant enzyme system. NADPH oxidase
catalyzes electron passage to molecular oxygen, whereas SOD
converts superoxide into H2O2 and then
catalyzes H2O2 into H2O and
O2, in sequence (44).
In the present study, the aging heart exhibited increased p47-phox
and gp91-phox expression from NADPH oxidase, and reduced SOD
expression. These data suggested that the age-associated MR
increase may be accompanied by increased ROS generation and
decreased antioxidant enzymes in the aging heart.
Further in vitro analyses demonstrated that
cardiomyocyte aging induced by H2O2 markedly
upregulated the expression of the aging markers p16 and p21, and
induced an imbalanced redox state. In addition, an imbalanced redox
state, including increased p47-phox, p67-phox and gp91-phox
expression, and reduced SOD1 and SOD2 expression, further increased
ROS accumulation. Eplerenone partially antagonized aging-induced
cardiomyocyte damage and improved the imbalanced redox state. These
results suggested that these protective effects were partly
mediated by MR activation.
The increase in MR activation may be a key
pathogenic factor that links aging-related mitochondrial
dysfunction to cardiac aging in rats. In addition, the results of
the present study indicated that the increase in age-associated MR
expression may lead to mitochondrial dysfunction, increased
oxidative stress and cardiac remodeling via an imbalance in the
p53/PGC-1α axis. Further studies are required to clarify the
interaction between MR and age-related mitochondrial dysfunction,
and to clarify whether the antagonism of MR in hearts can delay the
progression of cardiac aging and prolong lifespan.
In conclusion, the results of the present study
suggested that an increase in MR may serve an important role in
age-related cardiac dysfunction by decreasing mitochondrial renewal
mediated by the p53/PGC-1α metabolic axis, impairing mitochondrial
ultrastructure and inducing accumulation of ROS. Further
investigation of MR in the pathophysiology of cardiac aging may
facilitate the understanding of the mechanism underlying the
protective effects of MR antagonists in aging-induced cardiac
dysfunction, and it may be a suitable therapeutic target for
delaying age-related cardiac dysfunction and increasing
longevity.
Acknowledgements
Not applicable.
Funding
The present study was partly supported by Hu Bei
Health and Family Planning Commission (grant no. WJ2015MB006).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
DH designed the experiments, performed the research
and wrote the manuscript. RD and YY analyzed data. YZ, ZC, YT and
MF performed the experiments. LT conceived and designed the study,
and gave final approval of the version to be published. XX analyzed
and interpreted the data, and revised the manuscript. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the Animal
Research Ethics Board at Huazhong University of Science and
Technology and conformed to the Guide for the Care and Use of
Laboratory Animals published by the National Institutes of Health.
The present study was approved by the ethics committee of Tongji
Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
MR
|
mineralocorticoid receptor
|
|
SOD
|
superoxide dismutase
|
|
PGC-1α
|
peroxisome proliferator-activated
receptor γ coactivator-1α
|
|
ROS
|
reactive oxygen species
|
References
|
1
|
United Nations, Department of Economic,
Social Affairs and Population Division: World Population Ageing
2017 - Highlights (ST/ESA/SER.A/397). United Nations, New York, NY,
2017.
|
|
2
|
Wyss-Coray T: Ageing, neurodegeneration
and brain rejuvenation. Nature. 539:180–186. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rickard AJ, Morgan J, Bienvenu LA,
Fletcher EK, Cranston GA, Shen JZ, Reichelt ME, Delbridge LM and
Young MJ: Cardiomyocyte mineralocorticoid receptors are essential
for deoxycorticosterone/salt-mediated inflammation and cardiac
fibrosis. Hypertension. 60:1443–1450. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lother A, Berger S, Gilsbach R, Rösner S,
Ecke A, Barreto F, Bauersachs J, Schütz G and Hein L: Ablation of
mineralocorticoid receptors in myocytes but not in fibroblasts
preserves cardiac function. Hypertension. 57:746–754. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
López-Otin C, Blasco MA, Partridge L,
Serrano M and Kroemer G: The hallmarks of aging. Cell.
153:1194–1217. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Biernacka A and Frangogiannis NG: Aging
and cardiac fibrosis. Aging Dis. 2:158–173. 2011.PubMed/NCBI
|
|
7
|
Boyle AJ, Shih H, Hwang J, Ye J, Lee B,
Zhang Y, Kwon D, Jun K, Zheng D, Sievers R, et al: Cardiomyopathy
of aging in the mammalian heart is characterized by myocardial
hypertrophy, fibrosis and a predisposition towards cardiomyocyte
apoptosis and autophagy. Exp Gerontol. 46:549–559. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pitt B, Zannad F, Remme WJ, Cody R,
Castaigne A, Perez A, Palensky J and Wittes J: The effect of
spironolactone on morbidity and mortality in patients with severe
heart failure. randomized aldactone evaluation study investigators.
N Engl J Med. 341:709–717. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fraccarollo D, Berger S, Galuppo P, Kneitz
S, Hein L, Schütz G, Frantz S, Ertl G and Bauersachs J: Deletion of
cardiomyocyte mineralocorticoid receptor ameliorates adverse
remodeling after myocardial infarction. Circulation. 123:400–408.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kilkenny C, Browne W, Cuthill IC, Emerson
M and Altman DG; NC3Rs Reporting Guidelines Working Group, : Animal
research: Reporting in vivo experiments: The ARRIVE guidelines. Br
J Pharmacol. 160:1577–1579. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
AVMA Guidelines for the Euthanasia of
Animals: 2013 Edition. American Veterinary Medical Association.
(Schaumburg, IL). 2013.
|
|
12
|
Padilha CS, Borges FH, Costa Mendes da
Silva LE, Frajacomo FTT, Jordao AA, Duarte JA, Cecchini R, Guarnier
FA and Deminice R: Resistance exercise attenuates skeletal muscle
oxidative stress, systemic pro-inflammatory state, and cachexia in
Walker-256 tumor-bearing rats. Appl Physiol Nutr Metab. 42:916–923.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Guarnier FA, Cecchini AL, Suzukawa AA,
Maragno AL, Simão AN, Gomes MD and Cecchini R: Time course of
skeletal muscle loss and oxidative stress in rats with Walker 256
solid tumor. Muscle Nerve. 42:950–958. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lai J, Chen F, Chen J, Ruan G, He M, Chen
C, Tang J and Wang D: Overexpression of decorin promoted
angiogenesis in diabetic cardiomyopathy via IGF1R-AKT-VEGF
signaling. Sci Rep. 7:444732017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mascorro JA and Bozzola JJ: Processing
biological tissues for ultrastructural study. Methods Mol Biol.
369:19–34. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Feng W, Xu X, Zhao G, Zhao J, Dong R, Ma
B, Zhang Y, Long G, Wang DW and Tu L: Increased Age-related cardiac
dysfunction in bradykinin B2 receptor-deficient mice. J Gerontol A
Biol Sci Med Sci. 71:178–187. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Takimoto E, Champion HC, Li M, Ren S,
Rodriguez ER, Tavazzi B, Lazzarino G, Paolocci N, Gabrielson KL,
Wang Y and Kass DA: Oxidant stress from nitric oxide synthase-3
uncoupling stimulates cardiac pathologic remodeling from chronic
pressure load. J Clin Invest. 115:1221–1231. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tyner SD, Venkatachalam S, Choi J, Jones
S, Ghebranious N, Igelmann H, Lu X, Soron G, Cooper B, Brayton C,
et al: p53 mutant mice that display early ageing-associated
phenotypes. Nature. 415:45–53. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen T, Li J, Liu J, Li N, Wang S, Liu H,
Zeng M, Zhang Y and Bu P: Activation of SIRT3 by resveratrol
ameliorates cardiac fibrosis and improves cardiac function via the
TGF-β/Smad3 pathway. Am J Physiol Heart Circ Physiol.
308:H424–H434. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Susic D, Varagic J and Frohlich ED:
Isolated systolic hypertension in elderly WKY is reversed with
L-arginine and ACE inhibition. Hypertension. 38:1422–1426. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Neves MF, Cunha AR, Cunha MR, Gismondi RA
and Oigman W: The role of renin-angiotensin-aldosterone system and
its new components in arterial stiffness and vascular aging. High
Blood Press Cardiovasc Prev. 25:137–145. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cheng Z, Ito S, Nishio N, Thanasegaran S,
Fang H and Isobe K: Characteristics of cardiac aging in C57BL/6
mice. Exp Gerontol. 48:341–348. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sahin E, Colla S, Liesa M, Moslehi J,
Müller FL, Guo M, Cooper M, Kotton D, Fabian AJ, Walkey C, et al:
Telomere dysfunction induces metabolic and mitochondrial
compromise. Nature. 470:359–365. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Moslehi J, DePinho RA and Sahin E:
Telomeres and mitochondria in the aging heart. Circ Res.
110:1226–1237. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Leone TC and Kelly DP: Transcriptional
control of cardiac fuel metabolism and mitochondrial function. Cold
Spring Harb Symp Quant Bio. 76:175–182. 2011. View Article : Google Scholar
|
|
27
|
Lehman JJ, Barger PM, Kovacs A, Saffitz
JE, Medeiros DM and Kelly DP: Peroxisome proliferator-activated
receptor gamma coactivator-1 promotes cardiac mitochondrial
biogenesis. J Clin Invest. 106:847–856. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bonda TA, Szynaka B, Sokolowska M,
Dziemidowicz M, Winnicka MM, Chyczewski L and Kamiński KA:
Remodeling of the intercalated disc related to aging in the mouse
heart. J Cardiol. 68:261–268. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Burks TN, Marx R, Powell L, Rucker J,
Bedja D, Heacock E, Smith BJ, Foster DB, Kass D, O'Rourke B, et al:
Combined effects of aging and inflammation on renin-angiotensin
system mediate mitochondrial dysfunction and phenotypic changes in
cardiomyopathies. Oncotarget. 6:11979–11993. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yoon HE and Choi BS: The renin-angiotensin
system and aging in the kidney. Korean J Intern Med. 29:291–295.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Uneda K, Wakui H, Maeda A, Azushima K,
Kobayashi R, Haku S, Ohki K, Haruhara K, Kinguchi S, Matsuda M, et
al: Angiotensin II Type 1 receptor-associated protein regulates
kidney aging and lifespan independent of angiotensin. J Am Heart
Assoc. 6(pii): e0061202017.PubMed/NCBI
|
|
32
|
Yoon HE, Kim EN, Kim MY, Lim JH, Jang IA,
Ban TH, Shin SJ, Park CW, Chang YS and Choi BS: Age-associated
changes in the vascular renin-angiotensin system in mice. Oxid Med
Cell Longev. 2016:67310932016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang M, Zhang J, Walker SJ, Dworakowski R,
Lakatta EG and Shah AM: Involvement of NADPH oxidase in
age-associated cardiac remodeling. J Mol Cell Cardiol. 48:765–772.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Young MJ and Rickard AJ: Mineralocorticoid
receptors in the heart: Lessons from cell-selective transgenic
animals. J Endocrinol. 224:R1–R13. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ouvrard-Pascaud A, Sainte-Marie Y, Bénitah
JP, Perrier R, Soukaseum C, Nguyen Dinh Cat A, Royer A, Le Quang K,
Charpentier F, Demolombe S, et al: Conditional mineralocorticoid
receptor expression in the heart leads to life-threatening
arrhythmias. Circulation. 111:3025–3033. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sen N, Satija YK and Das S: PGC-1α, a key
modulator of p53, promotes cell survival upon metabolic stress. Mol
Cell. 44:621–634. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Viña J, Gomez-Cabrera MC, Borras C, Froio
T, Sanchis-Gomar F, Martinez-Bello VE and Pallardo FV:
Mitochondrial biogenesis in exercise and in ageing. Adv Drug Deliv
Rev. 61:1369–1374. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
López-Lluch G, Irusta PM, Navas P and de
Cabo R: Mitochondrial biogenesis and healthy aging. Exp Gerontol.
43:813–819. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sahin E and Depinho RA: Linking functional
decline of telomeres, mitochondria and stem cells during ageing.
Nature. 464:520–528. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Bedard K and Krause KH: The NOX family of
ROS-generating NADPH oxidases: Physiology and pathophysiology.
Physiol Rev. 87:245–313. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Tsutsui H, Kinugawa S and Matsushima S:
Mitochondrial oxidative stress and dysfunction in myocardial
remodelling. Cardiovasc Res. 81:449–456. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Di Lisa F, Kaludercic N, Carpi A, Menabó R
and Giorgio M: Mitochondria and vascular pathology. Pharmacol Rep.
61:123–130. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Gomez-Cabrera MC, Sanchis-Gomar F,
Garcia-Valles R, Pareja-Galeano H, Gambini J, Borras C and Viña J:
Mitochondria as sources and targets of damage in cellular aging.
Clin Chem Lab Med. 50:1287–1295. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Rajamohan SB, Raghuraman G, Prabhakar NR
and Kumar GK: NADPH oxidase-derived H(2)O(2) contributes to
angiotensin II-induced aldosterone synthesis in human and rat
adrenal cortical cells. Antioxid Redox Signal. 17:445–459. 2012.
View Article : Google Scholar : PubMed/NCBI
|