Introduction
Genetic information is stored in linear DNA
molecules - chromosomes in eukaryotic cells (1). McClintock and Möller in 1930 found
that complete chromosomes and their fragments behave differently in
the cells (2,3). The different properties of
chromosomes could be due to the presence of special nucleotide
sequences at the chromosomal ends, which are called telomeres
(4). Telomeres consist of
repeating nucleotide sequences and a set of special proteins that
interact with DNA to form a nucleoprotein complex (5). When these repetitive sequences were
used for in situ hybridization, the following conclusions
were made: Human heterochromatin is composed of some
heterochromatic pieces, (e.g., that of chromosome) that appear to
have more heterogeneous composition than others, while the highly
repetitive human DNA fractions are located primarily at the
centromeric and telomeric regions. Moreover, more slowly
re-associating repetitious sequences are distributed along the
chromatid, with a slight bias towards telomeric regions (6). During each cell division, the 5′ end
of the newly synthesized strand shortens due to the deletion of
terminal RNA primer, leading to incomplete replication of linear
DNA molecules. Olovnikov (1971) and Watson (1972) independently
formulated the ‘problem of insufficient replication’ in the 1970s
(7,8).
Aging is caused by degenerative diseases, which
generates an irreversible accumulation of mutations, inability of
cell division, increased susceptibility to morbidity, and
ultimately death. Human cells can undergo a limited number of cell
divisions and eventually reach an indivisible state called
replicative senescence, as telomeres get shorter in each cell
division. The link between aging and cancer is a critical aspect of
modern research. Unlimited replicative potential, or immortality,
is considered the main factor distinguishing cancer cells from
normal cells. Numerous studies have shown that tumour cell
immortality is associated with the activation of the enzyme
telomerase that is responsible for telomere replication and
compensates for the telomere shortening that occurs during each
cell division. Since the development of methods able to determine
telomerase activity, many human tissue samples have been examined
for the presence of telomerase activity. Telomerase activity has
been recorded in more than 85% of malignant tumours, whereas it was
absent in normal tissue cells (9).
It is claimed that telomere shortening may be the molecular clock
that triggers aging. Studies attempted to verify this hypothesis,
by transfecting two telomerase-negative normal human cell types,
retinal epithelial cells and foreskin fibroblasts, with vectors
encoding the catalytic subunit of human telomerase (hTERT). Unlike
the control clones that were telomerase-negative and exhibited
telomere shortening and aging, telomerase-positive clones had
elongated telomeres and exhibited reduced staining for
β-galactosidase, which is an aging biomarker. Importantly,
telomerase-positive clones exceeded their normal life expectancy by
at least 20 doublings, making it possible to establish a link
between telomere shortening and cellular aging in vitro
(10,11). Moreover, apart from the inhibition
of telomerase activity, telomere shortening may be a result of loss
of telomere protection and stability. This is caused by the
accumulation of unrepaired DNA damage during the cell cycle via
destabilization of telomeric protein complexes such as Shelterin
complex components i.e. telomeric repeat binding factor 1 and 2
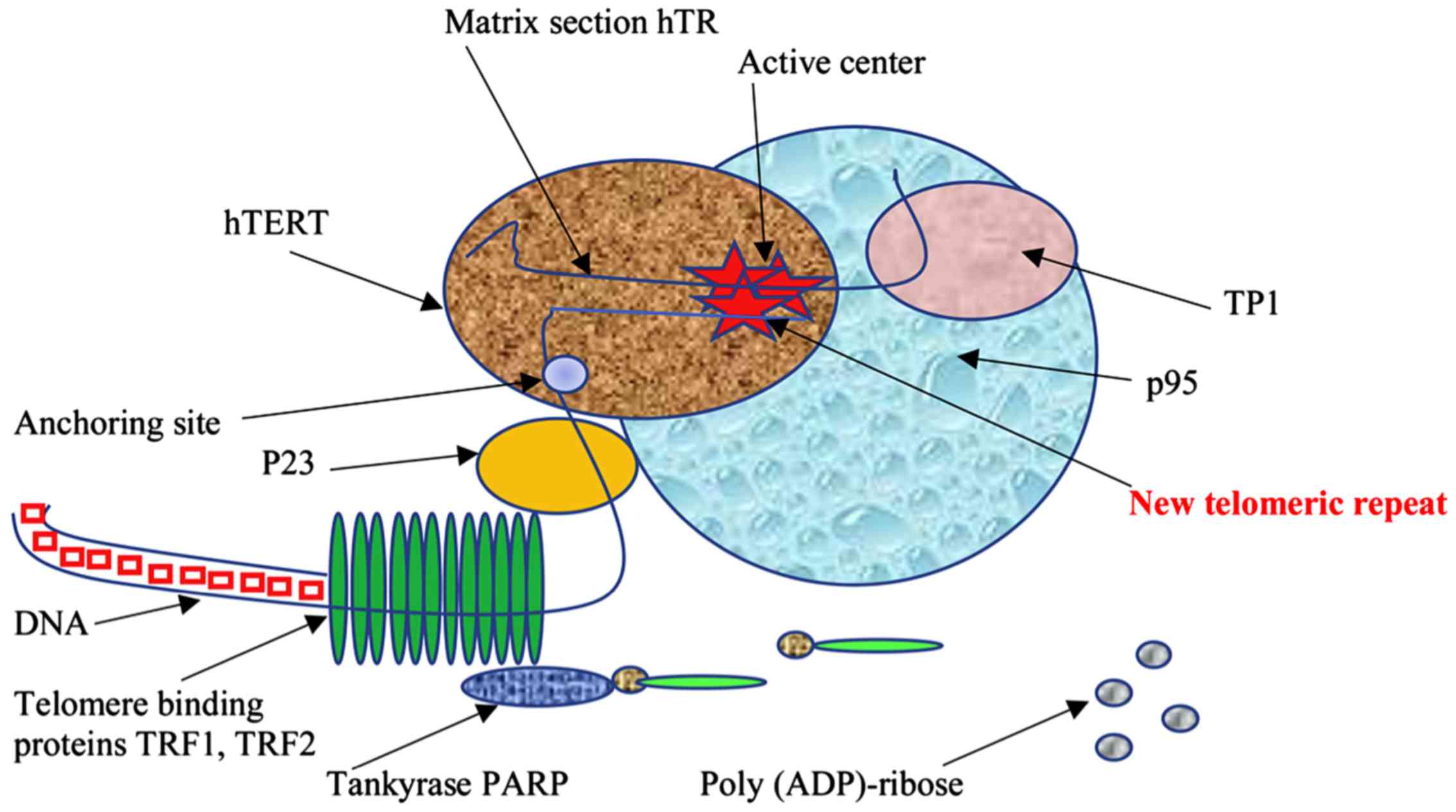
(TRF1 and TRF2) (Fig. 1) (12–15).
Telomeres and telomerase: The generation of
aging theory
In a historical perspective, Hayflick and Olovnikov
greatly contributed to the field of telomeres and telomerase, with
the work of Hayflick in 1961, triggering many scientists to try to
explain how cells count their divisions (16). He showed that human cells divide
only a limited number of times in culture, while at that time the
public opinion was dominated by the conviction that T cells are
immortal in the hands of a skilled researcher. The article received
a tremendous public response and the term ‘Hayflick limit’ was
created and included in the dictionaries.
At first, the telomere functions were unknown, and
the sequence of their nucleotides was not known either. In 1958,
the DNA polymerase enzyme responsible for DNA replication was
discovered by Lehman et al (17) and Bessman et al (18). DNA polymerase binds to a primer, a
short RNA fragment sitting on a DNA strand, that is synthesized by
another enzyme in order to initiate replication of telomeres and is
subsequently removed. During replication, DNA polymerase moves from
the 5′-end to the 3′-end, and as a result, it cannot replicate the
entire DNA molecule (there should be a non-copied fragment on one
of the ends to which it is attached), the so-called end replication
problem. Olovnikov (1971) and Watson (1972) first noticed this in
their independent observations (7,8).
Later, Olovnikov (1973) published the ‘theory of marginotomy’ that
can be considered a response to the work of Hayflick (1961),
explaining how the division counter works (19). In this work it was predicted that
in the case of the matrix synthesis of a linear polynucleotide
(chromosome replication), the copy should be shorter leading to the
limitation of the number of cell divisions observed. Also, the
existence of a special enzyme that extends the ends of chromosomes
in immortal cells was discussed.
The studies of Greider and Blackburn led the
discovery of telomerase in 1984, suggesting that the sequences at
the ends of chromosomes may have some special replication mechanism
(20). Moreover, Szostak and
Blackburn studied the molecular mechanisms of the theory of Möller
and McClintock, according to which, telomeres are required for
chromosome stabilization (2,3).
Importantly in 1988, Harley combined the idea of Olovnikov with the
studies of Greider and Blackburn, leading to a breakthrough in the
field (21) i.e., observing a
shortening of telomeres in human cells overtime (22).
Telomerase activity and aging
Telomerase is an enzyme responsible for the
replication of the telomeric regions of chromosomal DNA. It is
known that shortening of telomeric regions of DNA to critically
short length can serve as a signal for the replicative senescence
of somatic cells and destabilization of their chromosomes (23). Telomerase activity combined with
the telomere length (TEL) reflects the cell's proliferative
potential (24). Telomerase
activity is mainly present in stem, sex, and tumor cells but there
is also evidence of detectable activity in intestinal mucous cells,
in lymphocytes of peripheral blood (PB), and thymus (25). In addition, the catalytic subunit
of telomerase (hTERT), which is an RNA-dependent DNA polymerase
that synthesizes telomeric repeats, as well as the mRNA of this
subunit, is also expressed in these cells at a relatively low, but
detectable level.
During life, telomeres of PB lymphocytes gradually
shorten due to cell division, accumulating with age in vivo
(25). It has been shown that in
many diseases, including immune-associated diseases (rheumatoid
arthritis, atopic dermatitis, and bronchial asthma), PB lymphocytes
are characterized by shortened telomeres and increased
proliferation compared with those in healthy lymphocytes. Numerous
studies have shown that telomere shortening may contribute to
osteoarthritis and osteoporosis as an epigenetic factor and it is
further supported that TEL measurement of chondrocytes and PB cells
may be an appropriate marker for the evaluation of the progression
of these diseases (26–29). Moreover, other factors can
influence the rate of telomere shortening in lymphocytes, such as
the presence of chronic stress (30) and drug abuse (31). It has been shown that telomerase
expression in lymphocytes is strictly controlled during their
development, differentiation, and activation (30), and because of stimulation, ‘adult’
lymphocytes become capable of expressing telomerase at a rather
high level. After any re-stimulation, telomerase expression may
also increase but its level does not reach the level of response to
the primary stimulus (32). Such
dynamics may reflect the regulatory mechanisms of clonal expansion
of lymphocytes, which is part of the immune response and control
mechanism of their proliferation.
Somatic cells are mostly devoid of telomerase
activity. It is suggested that telomerase activity can be regulated
in human cells by intranuclear transfer of key components. It was
shown in HeLa cervical carcinoma cells that hTR and hTERT, during
most of the cell cycle, accumulate in intranuclear sites separately
from telomeres, whereas during S phase, they are specifically
recruited at telomeres. Therefore, the recruitment of telomerase at
telomeres is dynamic, reaching a peak in the middle of S phase.
Moreover, both hTR and hTERT are associated with nucleoli and Cajal
bodies during S phase, participating in telomerase transfer
(33). Interestingly, human
peritoneal mesothelial cells (HPMCs) appear to have unusually short
telomeres (3.5 kbps) despite the presence of active telomerase.
These telomeres do not shorten due to aging but due to a decrease
in telomerase activity. HPMC aging is associated with mitochondrial
dysfunction, leading to increased production of active oxygen
species and reduced mitochondrial membrane potential, indicating
that premature aging is highly associated with oxidative damage in
non-telomeric regions of the genome (34).
Telomerase activity in autoimmune diseases, such as
rheumatoid arthritis, lupus erythematosus, atypical dermatitis,
psoriasis, multiple sclerosis, and aplastic anaemia is summarized
and presented in Table I.
 | Table I.Telomerase activity in autoimmune
diseases. |
Table I.
Telomerase activity in autoimmune
diseases.
| No. | Disease | Telomerase activity
in autoimmune disease | (Refs.) |
|---|
| 1 | Rheumatoid
arthritis | The telomerase
activity level in synovial infiltrating lymphocytes was
significantly correlated with the intensity of synovial lining
hyperplasia, microvessel proliferation, lymphocyte infiltration,
and percentage of synovial cells positive for proliferating cell
nuclear antigen in rheumatoid synovium | Yudoh et al
(26) |
|
|
| Activated
lymphocytes with high telomerase activity distribute systemically
through the circulation and contribute disease activity in
connective tissue disease. The stimulatory effect of estrogens on
telomerase activity and female predominance in connective tissue
disease may contribute to high telomerase activity in these
pathologies. | Hiyama and Hiyama
(27); Yamanishi et al
(187); Tarhan et al
(188) |
|
|
| Telomerase activity
in lymphoid cells of lymphoid organs is higher than in peripheral
blood. | Hiyama and Hiyama
(27); |
|
|
| Highest hTERT
values were observed in Rheumatoid arthritis group (21.24±28.54).
hTERT values were significantly higher than the healthy control
group (7.62±4.21) (P<0.05). | Tarhan et al
(188); Yudoh et al
(26) |
|
|
| Lack of telomerase
activity leads to excessive loss of T cells, which limits the
possibility of homeostatic control of naive T-lymphocytes. | Fujii et al
(28) |
|
|
| T cells as a result
of repeated stimulations correspond to decreasing peaks of mRNA and
hTERT induction, compared with the first highest peak. | Valensuela and
Effros (141) |
| 2 | Systemic lupus
erythematosus | High telomerase
activity in peripheral blood mononuclear cells. | Katayama and
Kohriyama (29) |
|
|
| Significant
correlation between telomerase activity and disease severity. With
an exacerbation of the disease, telomerase activity in mononuclear
cells of peripheral blood is significantly higher than in the
control group. | Klapper et
al (43); Kurosaka et
al (189) |
|
|
| There was no
evidence of telomere shortening in lymphocytes and no correlation
between telomerase activity and telomere length. | Klapper et
al (43); Kurosaka et
al (189) |
|
|
| The activity of
telomerase in the cells is sufficient to compensate for the
shortening of telomeres, and thus prevent the accelerated aging of
the immune system of patients. | Klapper et
al (43) |
| 3 | Systemic
sclerosis | The most
interesting finding was very low PBC hTERT levels in the Systemic
sclerosis group. If really there is a tendency for lower telomerase
activity in Systemic sclerosis, this may be related with the
telomere shortening reported by Artlett et al. The authors
hypothesized that the chromosomal instability observed in Systemic
sclerosis patients may result from the loss of long stretches of
the telomeric repeats. Telomeric shortening may be expected to
stimulate telomerase activation; however, the reverse may also be
true. In other words, since the expression of telomerase stabilizes
telomere length and allows for continual replication, low
telomerase activity may contribute to telomere shortening. | Tarhan et al
(188); Artlett et al
(58) |
|
|
| T lymphocyte
homeostasis is significantly impaired. | Hug et al
(190); Tarhan et al
(188); Katayama and Kohriyama
(29) |
|
|
| Thymic T cell
production was estimated by measuring TCR excision circles (TRECs)
as a traceable molecular marker in recent thymic emigrants. an
impaired thymic export function and, as a consequence, altered
ability to maintain T cell homeostasis and immune tolerance may
play an important pathogenic role in relapsing remitting Multiplex
Sclerosis. | Hug et al
(190) |
|
|
| T cells as a result
of repeated stimulations correspond to decreasing peaks of mRNA and
hTERT induction, compared with the first highest peak. | Valensuela and
Effros (141) |
| 4 | Atopic dermatitis
and psoriasis | Telomerase activity
in the affected areas of the skin is significantly higher than in
intact skin and is associated with proliferative activity of the
cell. | Jang et al
(191) |
|
|
| Lymphocytes that
infiltrate the epidermis are not responsible for the level of
telomerase, since it does not correlate with the number of these
lymphocytes. | Ogoshi et al
(192); Taylor et al
(193) |
|
|
| The level of
telomerase activity in skin diseases that are not malignant is much
lower than in malignant tumors. | Taylor et al
(193) |
|
|
| A high level of
telomerase activity in peripheral blood mononuclear cells compared
to donors. | Wu et al
(46) |
| 5 |
| The length of
telomeres in blood cells progressively decreases by 216 base pairs
in addition to the age loss of 36 base pairs per year. | Brümmendorf et
al (194); Fogarty et
al (195) |
|
|
| The telomere
reduction and short cell life can be explained by the presence of
mutations associated with this disease in the hTR and hTERT
genes. | Ly (196); Young (197); Yamaguchi et al (198); Marrone et al (199) |
|
|
| Most patients
carrying these mutations are insensitive to immunosuppressive
therapy. | Ly (196); Young (197); Yamaguchi et al (198); Marrone et al (199) |
|
|
| Among patients with
polymorphisms in the TERT gene, the telomere length was within 90%
of the values of healthy donors. | Yamaguchi et
al (198) |
ALT-alternative lengthening of
telomeres
In addition to telomerase activation, there is a
telomere lengthening mechanism, alternative to telomerase
(ALT-alternative lengthening of telomeres). However, this mechanism
in mammalian cells is only present in abnormal situations, such as
in cancer cells, immortalized cell lines, and murine cell knockout
for the telomerase gene, whereas normal human lymphocytes cannot
use ALT to maintain their telomeres. A minority of immortal cell
lines and tumor cells use ALT to counteract telomere erosion due to
cell division (35,36). Observations on the dynamics of the
TEL of immortalized cells suggest that this mechanism is based on
recombination, resulting in individual telomeres that undergo rapid
shortening and rapid elongation (37). Besides, ALT cells can lengthen
their telomeres using telomeric sequences from other chromosomes as
a template for new DNA synthesis (38,39).
Cesare and Reddel (2010) convincingly proved in their research that
tumor cells treated with telomerase inhibitors can activate the ALT
pathway for telomeric maintenance through recombination (40). This can explain the therapeutic
failures and/or a resistance against telomerase inhibition-based
anti-cancer therapy. Therefore, developing new therapeutics
targeting proteins known to be involved in the latter pathway will
be crucial for the targeting of ALT-specific cells (41).
Telomerase is the main mechanism for stabilizing
telomeres in lymphocytes and they use it to reduce telomere
shortening during cell proliferation (42). Thus, in intact lymphocytes in
systemic lupus erythematosus, multiple sclerosis, and atopic
dermatitis, telomerase activity is enhanced (43). Moreover, quantitative fluorescence
in situ hybridization (Q-FISH) on metaphase chromosomes in
the PB lymphocytes showed that the distribution of TEL for
individual chromosomes differs in normal and pathological
conditions. Patients with rheumatoid arthritis had significantly
shorter telomeres of chromosome 4p, an observation that could be
related to the pathology of the disease, while the activity of the
disease is inversely correlated with the length of the telomeres of
the 10p chromosome which carries the genes involved in the
differentiation and proliferation of T-cells (44). The dynamic activity of telomerase
is correlated with the action of an acute stressor and with two
types of reactions to stress-psychological stress and
neuroendocrine (cortisol) responses to the stressor (24). Several studies indicate that in
most of the above situations (immunopathological diseases, stress,
and many others) the telomeres of lymphocytes shorten with age
faster than in healthy individuals (45,46).
Role of telomerase in
mitochondria
It is known that mitochondrial dysfunctions are
found in metabolic disorders, neurodegenerative diseases, and
autoimmune diseases (47,48). Mitochondria are important
organelles for the production of bioenergy, for biosynthesis and
signaling, thus an integral part of cellular adaptation to the
environment. Concomitantly, mitochondria are important mediators of
carcinogenesis, since this process requires flexibility for the
cells to adapt to the environmental changes. Many aspects of
mitochondrial biology support transformation, including
mitochondrial biogenesis and metabolism, fusion dynamics,
regulation of oxidative stress, and cell-signaling. Thus,
understanding the mechanisms of mitochondrial function during
cancer genesis is critical for future cancer treatment (49). Mutations in mitochondrial genes are
common in cancer cells, they do not inactivate mitochondrial energy
metabolism but rather alter the mitochondrial bioenergetic and
biosynthetic state, including succinate dehydrogenase, fumarate
hydratase, and isocitrate dehydrogenase 1, which lead to an
increase in the level of succinate, fumarate or
R-2-hydroxyglutarate. These metabolic changes can inhibit
α-ketoglutarate-dependent dioxygenases, possibly contributing to
oncogenesis (50).
Disruption of mitochondrial activity is directly
related to neurodegenerative diseases, indicative of the importance
of mitochondrial functions in normal cellular physiology. Recent
studies focused on mitochondrial function in reactive oxygen
generation and its participation in the development of
neurodegenerative diseases (51).
As is generally known, the mitochondrial genome is a
highly replicated, specialized genetic system that allows
individual mitochondria to respond to changes in membrane potential
and support oxidative phosphorylation by gene expression (52). Studying this special role, one
should not be surprised that the mitochondrial genome is regulated
and expressed in a unique way. Human mitochondria contain a compact
ring genome of 16,569 bp in length (53). Replication and transcription of
mitochondrial DNA (mtDNA) is initiated from the non-coding region,
loop D, and is regulated by nuclear-encoded proteins that are
post-translationally imported into mitochondria. Mitochondrial RNAs
are transcribed as long polycistronic precursor transcripts from
‘heavy’ and ‘light’ chains, which are processed according to the
‘tRNA punctuation model’, whereby 22 disseminated tRNAs are excised
to release rRNA and mRNA (54).
The released RNAs then undergo maturation, which involves the
polyadenylation of 30 ends of mRNA and rRNA (55). Together, this includes a unique
genetic system that is able to translate genes encoded by
mitochondria into 13 protein subunits of the electron transfer
chain. The subtle features of the mitochondrial transcriptome are
still poorly understood: Regulation of the number of transcripts,
RNA processing and modification sites, and the possible presence of
non-coding RNAs. The recent advent of deep sequencing has provided
a global nuclear transcriptome profile, revealing unanticipated
transcription complexity, which includes widespread
post-transcriptional processing and an abundance of non-coding RNA
(56–58). Mercer et al (2011) used a
similar approach to study mitochondrial transcriptome (59).
Wang et al (60) (2010) investigated the import
functions of some lncRNAs in mitochondria, showing that one of the
RNAs, the RNA component of the telomerase (TERC), is processed in
mitochondria and then exported out of mitochondria. Partial
disruption of mitochondrial function caused the accumulation of
mitochondrial-processed RNA, TERC, in the cytosol. These results
provide a potential link between mitochondrial function and
telomerase activity. The import of RNA into mitochondria is very
important for replication, transcription, and translation of the
mitochondrial genome. PNPase is an important regulator of the
import of nuclear-encoded RNA into the mitochondrial matrix. It has
been shown that the decrease in PNPase impairs mitochondrial RNA
processing. Moreover, the increase in imports of RNase P, 5S rRNA,
and multidrug resistance proteins RNA is directly dependent on the
expression of PNPase. These observations led the identification of
PNPase-imported RNA interactions, suggesting a new role of PNPase
in providing translocation of RNA to mitochondria (60). In addition, Cheng et al
(61) (2018) showed that the RNA
component of mammalian TERC telomerase is imported into
mitochondria, processed into the shorter form of TERC-53, and then
exported back to the cytosol. These data collectively reveal the
existence of a pathway of mitochondrial RNA transfer and provide a
potential mechanism for transferring the functional states of
mitochondria to other cellular compartments.
Several recent reports have studied the transport of
hTERT to mitochondria, which is most likely controlled by a
mitochondrial sequence at the N-terminus of hTERT. It has been
shown that the accumulation of endogenous oxidative stress leads to
the hTERT nuclear export to the cytosol, reducing the nuclear and
total telomerase activity. The addition of antioxidants slows down
this process significantly (62).
Aging is associated with an increase in the number of intracellular
reactive oxygen species (ROS) and a loss of telomerase activity.
Haendeler et al (62)
(2004) investigated whether an age-related increase in the number
of ROS can induce nuclear export of TERT and promote endothelial
cell aging. Haendeler et al found that in parallel with
increased ROS formation, the TERT nuclear protein is exported from
the nucleus to the cytoplasm and Src kinase is activated.
Incubation with N-acetylcysteine reduced the formation of
intracellular ROS and prevented mitochondrial DNA damage, as well
as low doses of atorvastatin that had a similar effect. Therefore,
it has been suggested that antioxidants and statins can delay the
onset of replicative aging by counteracting the increase in ROS
associated with aging of endothelial cells (62–64).
Telomeric stem cell biology
Most human stem cells are characterized by a rather
slow proliferation rate, which was estimated for the hematopoietic
stem cells (HSCs), as one division in 1–2 years (65). During the intervals between
divisions, the cells are in proliferative rest. However, even with
such proliferation rates, it is clear that stem cells have a
proliferative potential that is bigger than the Hayflick limit.
The regulation of TEL and telomerase activity is a
complex and dynamic process that is tightly linked to cell cycle
regulation. HSCs demonstrate telomeric shortening during
replicative aging despite expression of low levels of telomerase
(66). Granulocytes and naive T
cells showed a parallel two-phase decrease in TEL with age, which
most likely reflected the accumulated cell divisions in the HSCs
that are the common precursors of both types of cells. Memory T
cells also first showed a rapid decrease in TEL with age, which
continued for several years, resulting in lymphocytes with shorter
telomeres that granulocytes in older people. These results indicate
a sharp decrease in stem cell turnover in early childhood and
confirm that cell divisions in HSCs and T cells lead to telomere
shortening (67,68). Telomere shortening is most
pronounced during bone marrow resection, as it has been observed
that there is an inverse correlation between the number of
transplanted mononuclear cells and the degree of telomere
shortening. A bone marrow transplant results in a telomere
shortening equivalent to a period of 15 years of life (69).
Interestingly, Scheding et al (70) (2003) attempted to assess whether
repeated stimulation of the hematopoietic system due to regular
blood sampling will negatively affect the TEL of PB leukocytes
(PBLs). It is known that the TEL of PBLs can be used to evaluate
HSCs turnover. A study using fluorescence in situ
hybridisation and flow cytometry (Flow-FISH), showed that when
compared with the corresponding non-donor data, no differences were
observed. In addition, neither age-adjusted donor data for TEL for
granulocytes nor TEL for lymphocytes correlated with either the
total number of years or the total number of whole blood (WB)
and/or PLT donations. Long-term donation of WB does not affect TEL,
as measured by Flow-FISH, which indicates a significant increase in
stem cell turnover (70).
Moreover, lymphoid cells are capable of very intensive clonal
reproduction with antigenic stimulation. Despite the induction of
telomerase, their telomeres are more significantly shortened than
in granulocytes. Accordingly, memory cells have significantly
shorter telomeres than lymphocytes (71).
There are many unresolved issues in telomeric stem
cell biology and the various stem cells of an adult organism vary
greatly regarding the role of telomeres and telomerase in
maintaining proliferation. These stem cells include HSCs and
mesenchymal stem cells (MSCs) from human bone marrow, that are
extensively studied. hMSCs are a type of adult stem cells from the
bone marrow, which can differentiate into bone, cartilage, adipose,
and muscle cells (72). hMSCs show
a total absence of telomerase activity, in contrast to HSC and
other adult stem cells (73).
Ectopic telomerase expression is able to expand their limited
replicative ability in tissue culture, maintaining their functional
characteristics (74), a feature
that can be developed for the application of these cells in tissue
engineering (75).
Importantly, stem and cancer cells share several
characteristics, including factors that regulate self-renewal in
both normal HSCs and leukemic cells (76), leading to the hypothesis of ‘cancer
stem cells’ (77). Cancer stem
cells resemble normal stem cells in the sense that when they
divide, one of the daughter cells differentiates into a certain
type of cell, which eventually ceases to divide, while the other
retains its stem cell properties, being able to divide constantly.
Cancer stem cells, which constitute only a small fraction of the
total population of tumor cells, are the only tumor cells capable
of supporting tumor growth. Therefore, a scientific assumption was
made that a complete cure for cancer may require the development of
treatments targeting cancer stem cells, assuming that this can be
accomplished without destroying the stem cells needed to maintain
tissue regeneration, such as bone marrow and intestines (77). In addition, stem cells may be the
source of some types of cancer, increasing the risk when used to
repair organs, while they can also be of prognostic value as the
proportion of stem cells in a tumor can determine how deadly it
is.
Some therapeutic strategies for cancer treatment are
focused on the destruction of cancer stem cells. However, many
potential uses require stem cells to divide to produce enough cells
to replace damaged tissue, a fact that could possibly contribute to
the accumulation of potentially cancer-causing mutations (77). The origin of stem cells for some
types of cancer means that these stem cells have telomerase that
does not need to be reactivated, although the enzyme activity may
be further increased in the later stages of carcinogenesis
(78). On the contrary, the
limited activity of telomerase, associated with the final lifespan
of adult stem cells, can both contribute to aging and cancer
prevention (79,80).
Forced expression of telomerase
(telomerization of cells)
In many cell types, telomerase activity appears only
after the forced expression of the hTERT gene, a process called
telomerization. The RNA component of telomerase is expressed in
most human cells regardless of telomerase activity; therefore,
after the transfection of human cells with the hTERT gene,
telomerase activity appears in them, and cells gain the ability to
maintain TEL. Such cells are called telomerized. The presence of
telomerase activity does not violate the mechanisms of cell growth
regulation but it contributes to the stability of the genome,
preventing chromosomal rearrangements instead. The sensitivity of
telomerized and initial fibroblasts of adult skin to common
cytostatics i.e., doxorubicin, cisplatin, etoposide, and
dacarbazine was studied. No significant differences in sensitivity
were found. These observations directly support the safekeeping of
normal mechanisms of homeostasis in telomerized cells and
indirectly support the assumption that they can be used in cell
therapy (81).
Saretzki et al (82) (2004) found very similar molecular
events in two unrelated human embryonic stem cell (hESC) lines that
regulate the suppression of telomerase activity, increase oxidative
stress, and decrease DNA repair activity during spontaneous
differentiation. This study characterized resistance to oxidative
stress and restoration of DNA breaks in parallel with the
regulation of telomerase activity and TEL during differentiation of
hESC. Previously, it was reported that murine ESC has a high level
of antioxidant protection and effectively repair DNA chain breakage
but the level of antioxidant protection and restoration of chain
breakdown noticeably decrease during differentiation (82). The data obtained indicated the
deacetylation of histones H3 and H4 in the hTERT promoter region.
The deacetylation of histone H3 in the hTR promoter was suggested
as a candidate mechanism for explaining the suppression of
expression of both genes and the loss of telomerase activity during
differentiation. Other studies have shown that histone modification
is an important means of regulating telomerase gene expression
(83,84). The loss of telomerase activity is
accompanied by a shortening of telomeres, which is especially fast
in the early stages of differentiation. Telomere shortening can be
accelerated due to high ROS levels since telomeres are at least
partially deficient in repairing oxidative DNA damage (85). It was also shown that the
self-renewing potential and longevity of HSCs in vivo are
largely determined by the ROS levels (86).
Lonergan et al (87) (2006) suggested that mitochondrial
characteristics may be an additional and reliable way to test stem
cell competency. The results indicated that, the perinuclear
arrangement of mitochondria in the primate stem cell line,
accompanied by a low ATP/cell content and high oxygen consumption,
may be a true indicator of competence in stem cell differentiation,
and deviations from this profile suggest thaT cells differentiate
or possibly age.
Passos et al (88) (2007) found in their studies that an
increase in mitochondrial density in combination with mitochondrial
dysfunction can accelerate aging. This is confirmed by a negative
correlation between the density of mitochondria and the maximum
life expectancy in interspecific comparisons in mammals.
Strengthening mitochondrial biogenesis and, possibly, impaired
mitochondrial degradation and segregation, if this occurs as an
adaptation to pre-existing mitochondrial dysfunction, it can
significantly increase ROS production and, thus, actively
contribute to aging. In other words, both ATP and ROS production
increase in parallel with the bulk density of mitochondria. In an
aging organism, the mitochondrial mass and ROS production increase,
while the membrane potential decreases due to the activation of the
uncoupling protein (89).
A rather interesting result was obtained after the
introduction of the hTERT gene into the cells of a patient
suffering from Niemann-Pick C disease (NPC). NPC is a recessive
lipidosis that is characterized by excessive accumulation of free
cholesterol and glycosphingolipids in the endosomal lysosomal
system. Walter et al (90)
(2003) confirmed that ectopic expression of hTERT in human cells
leads to increased regulation of small GTPase Rab9 and its p40
effector. Telomerase immortalization of human cells affects the
function of the endosomal lysosomal system and in particular the
efflux of cholesterol from this system, independently of the NPC1
protein. Such changes may be necessary to maintain cells in a
continuous proliferative state (90,91).
In fact, a recent study has provided evidence for a correlation
between cholesterol content and cell cycle progression (92). Inhibition of cholesterol
biosynthesis with low concentrations of lovastatin blocked cell
proliferation. When LDL-cholesterol was added to these cells, cell
cycle progression resumed (92).
In NPC1 disease, cells are unable to metabolize LDL-derived
cholesterol due to its accumulation in the late endosomal lysosomal
system. The expression of hTERT in NPC cells leads to the
correction of the cell phenotype, including the elimination of the
accumulated cholesterol. In particular, in NPC-TERT cells,
cholesterol transport from the endosomal lysosomal system to the
plasma membrane is restored with a concomitant increase in
cholesterol esterification (90).
Transfection of cells with expression vectors
containing hTERT supports TEL and provides normal cells with an
unlimited life span in culture. It is important that hTERT prolongs
the life span of cultured cells well beyond normal aging without
causing a neoplastic transformation (93). Condon et al (93) developed a cell line from
hTERT-infected human myometrial cells (hTERT-HM). Cells expressing
telomerase contained mRNA for α-smooth muscle actin, smoothelin, an
oxytocin receptor, and an estrogen receptor α. Myometrial cells
immortalized with hTERT retained the differentiation markers that
are observed in primary smooth muscle cell (SMC) cultures.
As known, the ectopic expression of hTERT prolongs
the lifespan of certain human cells. McKee et al (94) (2003) introduced hTERT into human
SMCs and found that the resulting cells proliferated well beyond
their normal lifespan and, remarkably, retained the characteristics
of normal control SMCs. Using these neonatal SMCs, they were able
to create reliable human vessels, and then it becomes possible to
create coronary arteries for coronary artery bypass grafting.
Klinger et al (95) (2006)
concluded that this tissue engineering model emphasizes the effect
of donor age on cellular synthetic function, which apparently does
not depend on the increase in hTERT life span.
Chronic stress exposure and telomerase
activity
PB mononuclear cells (PBMCs), are most suitable for
the study of telomerase activity. Maximow (96) (1909), the founder of the stem cell
theory, revealed that PBMCs are stem cells that are common to
various blood elements in embryonic development and during the
fetal life in mammals. It has been shown that there is an
accelerated telomere shortening and reduced telomerase activity in
PBMC in various situations. Recent studies have shown changes in
TEL and telomerase activity due to psychological and social factors
(64). An earlier study showed
that telomere shortening and the decrease in the telomerase
activity is higher in PBMC of women with high stress levels
compared with the control group (24). This study initiated a large number
of similar studies, showing that telomeres are shorter in patients
with mood disorders (97), in
hemodialysis-depended patients (98) and in patients with depressive
disorder (99).
To date, the effect of chronic stress on telomerase
activity in humans has been described in many studies (68,100–102). This effect includes an increase
in the oxidative level, which stimulates hTERT nuclear export to
the cytosol and decreases the nuclear and total telomerase activity
(62,103,104). It has been observed that long
periods of psychological stress can increase vulnerability to the
effects of the virus that induces aging of specific T cells
(32).
According to Lansdorp (105) (2006), telomere shortening can be
due to the influence of psychological stress and social rank,
whereas Adams et al (106)
(2007) contradict the theory that socio-economically deprived
people age faster and, therefore, have shorter telomeres than the
more affluent part of society. Chronic stress can affect human
health through a variety of behavioural aspects and biochemical
pathways (107). The shortening
of telomeres in cells of the immune system occurs through
biochemical reactions (108),
while stress can also affect the occurrence of age-related diseases
(109). It has been shown that
metabolic stress plays an important role in telomere shortening and
that the decrease in the level of ω-3 fatty acids in the blood
correlates with telomeres and may be a marker of aging (110). Moreover, chronic stress and
depression are directly related to high levels of
hydroxy-deoxyguanosine and reduced levels of antioxidant enzymes
(111,112). In addition, oxidative stress
affects telomeres more frequently compared to other areas of
genomic DNA and inhibits telomerase activity in vitro in
various cell types (113,114).
An antioxidant function is associated with various
micronutrients, including vitamin C, E, folic acid, and ω-3 fatty
acids and, therefore, can positively affect TEL (115). Stress leads to having high levels
of pro-inflammatory cytokines, including IL-6 and TNF-α (116,117).
Insulin-like growth factor 1 (IGF-1) exerts a
mitogenic effect implicated in cell proliferation. It was first
reported that IGF-1 is involved in the modulation of telomerase
activity by enhancing the stimulation of telomerase activity caused
by phytohemagglutinin (PHA) in cord blood mononuclear cells, which
have characteristically low levels of telomerase activity and hTERT
expression (118,119).
Fibroblast growth factor 2 (FGF2) has been shown to
induce a dose-dependent increase in both neural precursor cell
proliferation and telomerase activity in primary cortical cultures
(118,120). In contrast, in both neurons and
glial cells in primary cultures of E15 mouse embryo cortex,
down-regulation of telomerase activity and mTERT gene expression
occurred in parallel with cell differentiation (120). In primary endothelial cells,
freshly isolated from intact endothelium, telomerase activity is
observable during logarithmic growth but not when cells enter
quiescence (121). Treatment of
quiescent human umbilical vein endothelial cells (HUVECs) with FGF2
reactivates telomerase activity in a time- and dose-dependent
manner, in contrast to vascular endothelial growth factor-A
(VEGF-A), which has no effect (121). Consistently, FGF2 but not VEGF-A,
up-regulates hTERT expression in parallel with transcription factor
Sp1 (121).
VEGF has angiogenic activity that can contribute to
tumor development by inducing vessel permeability that facilitates
tumor cell proliferation and metastasis (122). Although VEGF-A appears not to
regulate telomerase activity in human umbilical vein endothelial
cells (123), VEGF and
lysophosphatidic acid (LPA), both secreted from ovarian cancer
cells and known to promote cancer cell growth, have been shown to
regulate telomerase activity in non-endothelial cells (118,124). Up-regulation of telomerase
activity by VEGF appears to be receptor-dependent, as unsaturated
vitamin E, tocotrienol, has been shown to down-regulate telomerase
activity through down-regulating the VEGF receptor (125). Vitamin E has also been reported
to down-regulate telomerase activity in ovarian cancer cells
(118,126).
Genetic factors are also related to telomere
shortening. Monoamine oxidase-A and apolipoprotein E (APOE)
polymorphisms were statistically significant for TEL only in people
with mental and depressive disorders but not in healthy
respondents. In addition, sex, the APO-ε2 allele, and the
polymorphism of the monoamine oxidase-A have an indirect effect on
TEL. Thus, mental and depressive disorder is an additional mediator
between monoamine oxidase-A and APOE polymorphisms and TEL
(99). Further, studies revealed
similar patterns in the European population for the APOE-ε4 gene
polymorphism, which is associated with a risk of Alzheimer's
disease. Although telomeres are shortened significantly faster in
female carriers of the APOE-ε4 mutant allele, hormone replacement
therapy in middle-aged women can change the effect of allele
transfer on aging (127).
In order to explain the effect of chronic stress on
telomere shortening, Damjanovic et al (128) (2007) studied immunological
changes in caregivers for relatives with Alzheimer's disease. TEL
was significantly shorter in caregivers than in control subjects
and their immune response was weaker. Moreover, caregivers had
significantly lower T cell proliferation but higher production of
immunoregulatory cytokines (TNF-α and IL-10) than control subjects
in response to in vitro stimulation. These results
demonstrate that chronic stress leads to a reduction in TEL and is
associated with accelerated aging of T-cells.
However, there is some controversy among different
studies on telomere shortening under stress conditions. Effros
(129) (2001) proposed an
interesting model for the study of PB cells in vitro, where
lymphocytes from healthy donors, namely men and women aged 25–55
years, were exposed to various concentrations of cortisol, which is
a stress-related hormone or dimethyl sulfoxide. He showed that the
level of telomerase activity decreases under stress, which explains
the decrease in both the number of immune-competent T cells and the
length of telomeres in these cells under chronic stress, further
accelerating cell aging and deterioration of the immune system
(130,131).
Ornish et al (132) (2008) investigated for three
months the comprehensive lifestyle changes for 30 men with
biopsy-diagnosed low-risk prostate cancer, which led to a
significant increase in telomerase activity by 10%. Increased
telomerase activity was associated with a decrease in the level of
low-density lipoprotein cholesterol (LDL) and a decrease in
psychological stress. Physical activity appears to protect against
metabolic and psychological stress and maintain TEL (133). Moreover, a study on various diets
showed that chronic food restriction (regardless of age), body mass
index, or smoking might be a risk factor for telomere premature
shortening (134). Furthermore,
regular sleep, exercise, and a healthy lifestyle contribute to
normal TEL and telomerase activity, regardless of the level of the
depressive disorder on the Hamilton Depression Rating Scale
(135). These data are consistent
with the results of another study, which showed that the
effectiveness of an antidepressant depends on the initial level of
telomerase activity (136). In
addition, De Punder et al (137) (2018) recently reported that high
levels of chronic stress reduce the ability of immune cells to
induce telomerase activity by 25% compared to moderate or low
levels of chronic stress. Chronic stress is associated with a
decrease in mitogen-induced lymphocyte proliferation and
mitogen-induced IL-2 production (138), which are activators of signalling
pathways that stimulate telomerase activity (139,140). Chronic stress exposure is also
associated with a higher level of oxidative stress, which
ultimately leads to the accumulation of senescent
(CD28−) T-cells (103). This suggests stringent telomerase
regulation in human T cells, which may contribute to telomere
shortening and ultimate replicative potential and loss of control
over certain pathogens (141).
In conclusion, the length of telomeres in PB cells
can be an indicator of human health, stress resistance, and cell
adaptation. Accordingly, changes in TEL and telomerase activity may
reflect the effectiveness of the treatment of age-related diseases.
Most of the recent studies on telomeres and age-related diseases
are based on the fact that increase in telomerase activity
corresponds to either a decrease in telomere shortening (30), or telomere elongation (142).
Effect of TEL on fertility and
pregnancy
Vasilopoulos et al (143) (2019) investigated the potential
relationship between TEL and various factors of female and male
infertility. Most female infertility factors have been associated
with shorter TEL, with the exception of endometriosis, premature
ovarian failure, transparent cell carcinoma, and polycystic ovary
syndrome (PCOS). These types of diseases have been associated with
a longer TEL, which has shown conflicting results in several
studies. On the other hand, male infertility factors were
associated with critically shorter TEL (143).
Fragkiadaki et al (144) (2016) summarized and critically
discussed the evidence linking telomerase activity with pregnancy
complications. Maternal age is a determining factor in
fertilization success and numerous studies have focused on
telomerase activity and its correlation with mammalian
fertilization, as well as subsequent splitting processes and
preimplantation development. It has been also shown that there is a
relationship between telomerase activity and complications during
pregnancy.
Additionally, the effect of psychosocial stress on
pregnancy is an important risk factor for the early onset of common
age-related diseases (145).
Human studies have demonstrated a link between chronic or excessive
psychosocial stress and telomere biology (146). It was shown that stress-related
changes in telomeres might be a possible mechanism linking
psychosocial stress with age-related diseases (147). Indeed, the accelerated shortening
of telomeres reflects stress-induced oxidative cell damage and
accelerated aging (97).
Therefore, Lazarides et al (148) (2019) examined the hypothesis that
the maternal pro-inflammatory state during pregnancy, which is a
balance between TNF-α, the main pro-inflammatory cytokine, and
IL-10, the main anti-inflammatory cytokine, are directly related to
the TEL of the leukocytes (LTL) of the newborn at birth. The higher
average ratio of TNF-α/IL-10 during pregnancy was significantly
associated with shorter TEL of the newborn after adjusting for the
mother's age, body mass index before pregnancy and sex of the
child.
Effect of nutraceutical supplements on
TEL
The Dietary Inflammatory Index (DII) was developed
by Shivappa et al (2014) to measure the inflammatory
potential of diet and it can be used in diverse populations to
predict levels of inflammatory markers, including C-reactive
protein (CRP) and interleukin-6 (149–151). Shivappa et al (152) (2017) examined whether
inflammatory potential of diet, as measured by the Dietary
Inflammatory Index (DII) has an impact on telomere shortening in
the National Health and Nutrition Examination Survey (NHANES).
Validation of the DII with C-reactive protein (CRP) was also
carried out, showing that there was an association between DII and
leukocyte TEL in these NHANES data. The impact of specific fatty
acids on inflammation can be crucial to the effect of dietary fats
on human health, for example polyunsaturated fatty acids, mainly
ω-3 fatty acids, have anti-inflammatory and immunomodulating
properties. Research studies have confirmed that excessive amounts
of ω-6 fatty acids leading to a high ω-6: ω-3 fatty acid ratio can
promote the pathogenesis of chronic diseases (153). Interestingly, a Mediterranean
diet supplemented with two fatty fish meals per week, rich in ω-3
fatty acids, has been shown to have an anti-inflammatory effect
reducing airway inflammation in childhood asthma that is a chronic
disease (154). Therefore, it is
likely that blood polyunsaturated fatty acid levels are involved in
preventing telomere shortening over time (155). In agreement with this, recent
studies revealed that the administration of nutraceutical
supplements to healthy individuals is implicated in TEL maintenance
(156). Participants were
selected from healthy outpatients and divided into an intervention
group that received nutraceutical supplements and a control group.
The length of individual telomeres was estimated in metaphase
leukocytes isolated from PB using Q-FISH analysis. The median
length of the telomeres was significantly increased (P<0.05) in
the intervention group compared with the control group. The
beneficial effect of supplements on the length of short telomeres
was significant (P<0.05) after correction for age and sex,
suggesting that nutraceutical supplements can directly affect the
maintenance of TEL.
Chronographic theory of aging: Neural
chronograph
As mentioned, Olovnikov (1973) described the
‘problem of insufficient replication’ in the 1970s and predicted
that in the case of the matrix synthesis of a linear polynucleotide
(chromosome replication), the copy should be shorter and the
limitation of the number of cell divisions is associated with this
fact (19). This led him to
construct a new theory of aging (160,161). It has been proposed that if
development and aging are programmed, then some ‘watch’ mechanism
that controls aging is required (67,162–165). Biorhythms are often called
clocks, and genes involved in rhythms, usually circadian rhythms,
are referred to as clock genes. But is there a real timekeeping
mechanism in the body that would organize, register, and track
physiological changes according to the age of the body? Olovnikov
bases his theory on the idea of the controlled loss of neurons in
the brain of hypothetical organelles, called chronomeres, which
represent small DNA molecules that were amplified from chromosomal
DNA segments (162,163).
Chronographic theory of development
and aging
A biochronograph to describe the development,
aging, and longevity of a certain species, should consider that
some genes have probably changed their expression in different
species and in different age groups. In C. elegans, for
instance, a relatively small number of genes show significant
changes in transcript levels as they age (<1% of the genome)
(166). Comparison of the results
of the C. elegans shows sets of overlapping genes that are
highly conserved throughout evolution and appear to be genes that
actually control aging and lifespan (167,168). Herndon et al (169) (2002) found convincing evidence
that stochastic as well as genetic factors are important in aging
C. elegans, with extensive variability between cells of the
same type within individuals. Moreover, the timing of gene
expression of adult Drosophila is regulated by independent
mechanisms related to temperature and metabolic rate. Studies of
relative temporary scaling at long-living mutants of C.
elegans showed that the life expectancy is controlled by some
physiological hours different from chronological time (170).
Olovnikov (171)
(2007) claims that the lunar cycle plays a role in aging of living
organisms. He suggested that pineal cells called pinealotcytes
regularly change the endocrine secretory activity, responding to
periodic changes of the gravitational field of the moon. The theory
is based on the idea of a controlled loss by a neuron in the brain
of chronomeres. The regular process of loss of chronomeres in
neurons is controlled by the pineal gland and activated at least
once a lunar month. Hormonal signals can be generated by the pineal
gland and depend on the gravity of the periodic shift of the
mineralized deposits located at the intra-pinealotcyte of neighbour
cellular structures. The calcific large particles that are in
extracellular spaces of a pineal gland can also contribute to the
hormonal secretion.
This pineal secretion activates the hypothalamus,
enhancing the hormonal response, which allows a sharp increase in
the concentration of several hormones, the so-called T-signal. The
T-signal is a ‘hormonal intracerebral explosion’ that happens in
humans every month and is a kind of hormonal combination, which can
enhance transcription in brain cells. Under certain conditions, the
chronomeres in response to a T-signal can lose each neuron
performing temporary functions, and as a result, the neuron changes
the activity. As mentioned above, chronomeres are hypothetical
mini-organelles that are present in the neurons performing
temporary functions, including counting time and participating in
the control of the biological age of an organism. Chronomere is a
perichromosomal copy of the chromosomal original, a rather small
two-chained DNA molecule, which is protected by proteins. Thus,
each chronomere is on a surface of a chromosome and is levelled
along this segment of chromosomal DNA, which serves as the original
for this chronomere. Chronomere DNA contains various regulatory
sequences (171).
Adjusting the sensitivity of endocrine gravity
sensors in different species of vertebrates may be one of the key
factors responsible for the rate of embryonic development in each
species of vertebrate. Gradual suppression of inhibitors is very
important for embryonic brain development (172,173). Changes in functional inhibitory
interactions are necessary for the maturation of the prefrontal
cortex when the cerebral cortex is connected (174). Epigenetic suppression of key
inhibitory loci may play a fundamental role in the initiation of
puberty (175). When the
maturation of the body is complete, clusters of neurons remain in
the brain, which are essentially the endpoint of the time relay,
while the temporary neurons of these clusters contain only terminal
chronomeres, which gradually decrease. The greater the reserves of
such neurons, the longer the lifetime is, considering that all
other things are equal, which could be the reason why life
expectancy is positively correlated with the cephalization index
(176).
Age-related changes in the human brain
as a possible outcome of the chronomere's function
Dedifferentiation of brain cells is used as a means
of development and loss of control and dedifferentiation of neurons
is found in several pathologies (177). However, changes in the brain must
also occur during normal development. Dedifferentiation or
decreased treatment specificity is a robust characteristic of
cognitive aging. Voss et al proved that neural
dedifferentiation was not ubiquitous in different categories of
stimuli, while nervous dedifferentiation was relatively stable
according to the age of the elderly (178,179). Moreover, an age-related decline
in specialization has also been shown for the cognitive function of
the frontal lobes of the hemispheres (180). Dreher et al examined the
relationship between dopamine synthesis in the midbrain and the
prefrontal ligament with reward. It was found that healthy aging
causes functional changes in the reward system, and reveals
age-related changes in the relationship between dopamine synthesis
in the midbrain and prefrontal activity. These results gave insight
into the interaction between dopamine function of the midbrain and
the reward system in young people and the elderly, and also
determined the changes that accompany aging (181). Furthermore, it has been shown
that the contraction of the hippocampus, inferior temporal cortex
and prefrontal white matter are increased with age, while shrinkage
in the hippocampus and cerebellum accelerates (182,183). The weakening of the functional
correlations of individual parts of the brain spreads with age from
the forehead to the back of the head (184).
The decrease in the volume of a brain becomes
noticeable approximately from 30 years of age (185). Total brain volume for mentally
healthy people is steadily declining by 0.22% per year between 20
and 80 years, and this decrease is accelerated in older age
(186). The brain in some
patients has vital deposits of amyloid protein indicative of the
preclinical phase of Alzheimer's disease: Their brain volume is
reduced by 2.5% compared with the brain of healthy volunteers. All
of the listed age-related changes, including a decrease in brain
volume, are considered a result of aging caused by various factors,
among which is the accumulation of damage caused by free radicals.
However, qualitative changes in the brain can be interpreted in a
completely different way, meaning that all these changes in the
brain are not the result of errors, but the product of genetic
programming.
Conclusions and future research
directions
As discussed in this review, telomerase activity is
regulated in response to various biological factors and systems,
including stress hormones, oxidative stress, inflammatory
mediators, and circadian rhythm. Therefore, future studies of
telomerase activity in PB mononuclear cells should include
measurements of biomarkers that participate in these processes,
which will significantly contribute to enhancing our knowledge in
the field. Moreover, it is necessary to further study the role of
telomerase in the defence mechanism of mitochondria against
oxidative stress to gain a broader understanding of the role of
telomerase in human health and the risk of stress-related
diseases.
A decrease in telomerase activity may explain why
telomeres shorten faster under pathological conditions. The study
and characterization of individual differences in the ability of
the telomere biology system to respond to a challenge, in addition
to indicators of basal telomerase activity and TEL, is an
interesting non-standard approach for future studies of telomere
biology and aging.
Acknowledgements
The authors would like to thank Dr Muhammad Amjad
Nawaz from SEC of Nanotechnology, Far Eastern Federal University
(Vladivostok, Russia) for his assistance with the copy-editing of
the manuscript and for correcting the final proofs.
Funding
This study was supported by Spin-Off Toxplus S.A.
and by the Special Research Account of University of Crete (ELKE
nos. 4602, 4920 and 3963).
Availability of data and materials
Not applicable.
Authors' contributions
All the authors contributed in conceiving and
designing the study. MPR, AMZ and KSG searched the literature for
inclusion in the study and wrote the manuscript. MT, ES, and PF
checked and reviewed the manuscript. KN, DT, DAS, and AT gave
advice and critically revised the manuscript. All authors have read
and approved the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
DAS is the Editor-in-Chief for the journal, but had
no personal involvement in the reviewing process, or any influence
in terms of adjudicating on the final decision, for this article.
The other authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
hTERT
|
human telomerase reverse
transcriptase
|
|
TRF1
|
telomeric repeat binding factor 1
|
|
TRF2
|
telomeric repeat binding factor 2
|
|
PB
|
peripheral blood
|
|
HPMCs
|
human peritoneal mesothelial
cells
|
|
ALT
|
alternative lengthening of
telomeres
|
|
Q-FISH
|
quantitative fluorescence in
situ hybridization
|
|
SDH
|
succinate dehydrogenase
|
|
FH
|
fumarate hydratase
|
|
IDH1
|
isocitrate dehydrogenase 1
|
|
lncRNAs
|
long non-coding RNA
|
|
tRNA
|
transfer RNA
|
|
rRNA
|
ribosomal RNA
|
|
mRNA
|
messenger RNA
|
|
TERC
|
telomerase RNA component
|
|
PNPase
|
polynucleotide phosphorylase
|
|
PDL
|
population doublings
|
|
TEL
|
telomere length
|
|
PBLs
|
peripheral blood leukocytes
|
|
Flow-FISH
|
fluorescence in situ
hybridisation and flow cytometry
|
|
WB
|
whole blood
|
|
HSC
|
hematopoietic stem cells
|
|
MSC
|
mesenchymal stem cells
|
|
ATP
|
adenosine triphosphate
|
|
NPC
|
Niemann-Pick C disease
|
|
SMCs
|
smooth muscle cells
|
|
LDL
|
low-density lipoprotein
cholesterol
|
|
ROS
|
reactive oxygen species
|
References
|
1
|
Morgan TH: Random segregation versus
coupling in Mendelian inheritance. Science. 34:636–638. 1911.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
McClintock B: Cytological observations of
deficiencies involving known genes, translocations and an inversion
in Zea mays. Mo Agric Exp Res Stn Res Bull. 163:1–30. 1931.
|
|
3
|
Möller HJ: The remaking of chromosomes.
Collecting Net. 8:182–198. 1938.
|
|
4
|
Blackburn EH and Gall JG: A tandemly
repeated sequence at the termini of the extrachromosomal ribosomal
RNA genes in Tetrahymena. J Mol Biol. 120:33–53. 1978. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
De Lange T, Lundblad V and Blackburn EH:
Telomeres. Cold Spring Harbor Laboratory Press; New York, NY: pp.
21–48. 2006
|
|
6
|
Hsu TC, Arrighi FE and Saunders GF:
Compositional heterogeneity of human heterochromatin. Proc Natl
Acad Sci USA. 69:1464–1466. 1972. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Watson JD: Origin of concatemeric T7 DNA.
Nat N Biol. 239:197–201. 1972. View Article : Google Scholar
|
|
8
|
Olovnikov AM: The principle of marginotomy
in the matrix synthesis of polynucleotides. Dokl Akad Nauk SSSR.
201:1496–1499. 1971.PubMed/NCBI
|
|
9
|
Shay JW and Wright WE: Telomerase activity
in human cancer. Cur Opin Oncol. 8:66–71. 1996. View Article : Google Scholar
|
|
10
|
Bodnar AG, Ouellette M, Frolkis M, Holt
SE, Chiu CP, Morin GB, Harley CB, Shay JW, Lichtsteiner S and
Wright WE: Extension of life-span by introduction of telomerase
into normal human cells. Science. 279:349–352. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Martin M: The biology of aging: 1985–2010
and beyond. FASEB J. 25:3756–3762. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Thanasoula M, Escandell JM, Martinez P,
Badie S, Muñoz P, Blasco MA and Tarsounas M: p53 prevents entry
into mitosis with uncapped telomeres. Curr Biol. 20:521–526. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Thanasoula M, Escandell JM, Suwaki N and
Tarsounas M: ATM/ATR checkpoint activation downregulates CDC25C to
prevent mitotic entry with uncapped telomeres. EMBO J.
31:3398–3410. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tejera AM, Stagno d'Alcontres M,
Thanasoula M, Marion RM, Martinez P, Liao C, Flores JM, Tarsounas M
and Blasco MA: TPP1 is required for TERT recruitment, telomere
elongation during nuclear reprogramming, and normal skin
development in mice. Dev Cell. 18:775–789. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Martínez P, Thanasoula M, Muñoz P, Liao C,
Tejera A, McNees C, Flores JM, Fernández-Capetillo O, Tarsounas M
and Blasco MA: Increased telomere fragility and fusions resulting
from TRF1 deficiency lead to degenerative pathologies and increased
cancer in mice. Genes. 23:2060–2075. 2009. View Article : Google Scholar
|
|
16
|
Hayflick L and Moorhead PS: The serial
cultivation of human diploid cell strains. Exp Cell Res.
25:585–621. 1961. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lehman IR, Bessman MJ, Simms ES and
Kornberg A: Enzymatic synthesis of deoxynucleic acid. I.
Preparation of substances and partial purification of an enzyme
from Escherichia coli. J Biol Chem. 233:163–170. 1958.PubMed/NCBI
|
|
18
|
Bessman MJ, Lehman IR, Simms ES and
Kornberg A: Enzymatic synthesis of deoxynucleic acid. II. General
properties of the reaction J Biol Chem. 233:171–177.
1958.PubMed/NCBI
|
|
19
|
Olovnikov AM: A Theory of marginotomy. The
incomplete copying of template margin in enzymic synthesis of poly
nucleotides and biological significance of the phenomenon. J Theor
Biol. 41:181–190. 1973. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Greider CW and Blackburn EH: The telomere
terminal transferase of tetrahymena is a ribonucleoprotein enzyme
with 2 kinds of primer specificity. Cell. 51:887–898. 1987.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Harley CB, Vaziri H, Counter CM and Allsop
RC: The telomere hypothesis of cellular aging. Exp Gerontol.
27:375–382. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Greider CW and Blackburn EH: Telomeres,
telomerase and cancer. Sci Am. 274:92–97. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chan SRWL and Blackburn EH: Telomeres and
telomerase. Philos Trans R Soc Lond B Biol Sci. 359:109–121. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Epel ES, Blackburn EH, Lin J, Dhabha FS,
Adler NE, Morrow JD and Cawthon RM: Accelerated telomere shortening
in response to life stress. Proc Natl Acad Sci USA.
101:17312–17315. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Osterhage JL and Friedman KL: Chromosome
end maintenance by telomerase. J Biol Chem. 284:16061–16065. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yudoh K, Matsuno H, Nezuka T and Kimura T:
Different mechanisms of synovial hyperplasia in rheumatoid
arthritis and pigmented villonodular synovitis: The role of
telomerase activity in synovial proliferation. Arthritis Rheum.
42:669–677. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hiyama E and Hiyama K: Telomere and
telomerase in stem cells. Br J Cancer. 96:1020–1024. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fujii H, Shao L, Colmegna I, Goronzy JJ
and Weyand CM: Telomerase insufficiency in rheumatoid arthritis.
Proc Natl Acad Sci USA. 106:4360–4365. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Katayama Y and Kohriyama K: Telomerase
activity in peripheral blood mononuclear cells of systemic
connective tissue diseases. J Rheumatol. 28:288–291.
2001.PubMed/NCBI
|
|
30
|
Georgin-Lavialle S, Aouba A, Mouthon L,
Londono-Vallego JA, Lepelletier Y, Gabet AS and Hermine O: The
telomere/telomerase system in autoimmune and systemic
immune-mediated diseases. Autoimmun Rev. 9:646–651. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Vakonaki E, Tsiminikaki K, Plaitis S,
Fragkiadaki P, Tsoukalas D, Katsikantami I, Vaki G, Tzatzarakis MN,
Spandidos DA and Tsatsakis AM: Common mental disorders and
association with telomere length. Biomed Rep. 8:111–116.
2018.PubMed/NCBI
|
|
32
|
Akbar AN and Vukmanovic-Stejic M:
Telomerase in T lymphocytes: Use it and lose it? J Immunol.
178:6689–6694. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tomlinson RL, Ziegler TD, Supakorndej T,
Terns RM and Terns MP: Cell cycle-regulated trafficking of human
telomerase to telomeres. Mol Biol Cell. 17:955–965. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ksiazek K, Passos JF, Olijslagers S,
Saretzki G, Martin-Ruiz C and von Zglinicki T: Premature senescence
of mesothelial cells is associated with non-telomeric DNA damage.
Biochem Biophys Res Commun. 362:707–711. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yeager TR, Neumann AA, Englezou A,
Huschtscha LI, Noble JR and Reddel RR: Telomerase-negative
immortalized human cells contain a novel type of promyelocytic
leukemia (PML) body. Cancer Res. 59:4175–4179. 1999.PubMed/NCBI
|
|
36
|
Bryan TM, Englezou A, Gupta J, Bacchetti S
and Reddel RR: Telomere elongation in immortal human cells without
detectable telomerase activity. EMBO J. 14:4240–4248. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Perrem K, Colgin LM, Neumann AA, Yeager TR
and Reddel RR: Coexistence of alternative lengthening of telomeres
and telomerase in hTERT-transfected GM847 cells. Mol Cell Biol.
21:3862–3875. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Dunham MA, Neumann AA, Fasching CL and
Reddel RR: Telomere maintenance by recombination in human cells.
Nat Genet. 26:447–450. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
39
|
Varley H, Pickett HA, Foxon JL, Reddel RR
and Royle NJ: Molecular characterization of inter-telomere and
intra-telomere mutations in human ALT cells. Nat Genet. 30:301–305.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cesare AJ and Reddel RR: Alternative
lengthening of telomeres: Models, mechanisms and implications. Nat
Rev Genet. 11:319–330. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zencir S, Hsieh MH, Hsu JS, Ergun Y, Chou
GL, Li TK, Teng SC and Topcu Z: Selected Ellipticine derivatives,
known to target topoisomerase II, suppress the alternative
lengthening of telomere (ALT) pathway in telomerase-negative cells.
J Cancer Res Clin Oncol. 146:1671–1676. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kashubowska L: Telomere shortening and
ageing of immune systems. J Physiol Pharmacol. 59:169–186.
2008.PubMed/NCBI
|
|
43
|
Klapper W, Moosig F, Sotnikova A, Qian W,
Schroeder JO and Parwaresch R: Telomerase activity in B and T
lymphocytes of patients with systemic lupus erythematosus. Ann
Rheum Dis. 63:1681–1683. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Blinova EA, Zinnatova EV, Barkovskaya MSh,
Borisov VI, Sizikov AE, Kozhevnikov VS, Rubtsov NB and Kozlov VA:
Telomere length of individual chromosomes in patients with
rheumatoid arthritis. Bull Exp Biol Med. 160:779–782. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lin J, Xie J and Qian WB: Telomerase
activity and telomere length in CD4+, CD8+
and CD19+ lymphocytes from patients with systemic lupus
erythematosus. Zhejiang Da Xue Xue Bao Yi Xue Ban. 34:534–537.
2005.(In Chinese). PubMed/NCBI
|
|
46
|
Wu K, Higashi N, Hansen ER, Lund M, Bang K
and Thestrup-Pedersen K: Telomerase activity is increased and
telomere length shortened in T-cells from blood of patients with
atopic dermatitis and psoriasis. J Immunol. 165:4742–4747. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Bishop NA, Lu T and Yankner BA: Neural
mechanisms of ageing and cognitive decline. Nature. 464:529–535.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sun N, Youle RJ and Finkel T: The
mitochondrial basis of aging. Mol Cell. 61:654–666. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Vyas S, Zaganjor E and Haigis MC:
Mitochondria and cancer Cell. 166:555–566. 2016.
|
|
50
|
Wallace D: Mitochondria and cancer. Nat
Rev Cancer. 12:685–698. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kausar S, Wang F and Cui H: The role of
mitochondria in reactive oxygen species and its implications for
neurodegenerative diseases. Cells. 7:2742018. View Article : Google Scholar
|
|
52
|
Lane N and Martin W: The energetics of
genome complexity. Nature. 467:929–934. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Anderson S, Bankier AT, Barrell BG, de
Bruijn MH, Coulson AR, Drouin J, Eperon IC, Nierlich DP, Roe BA,
Sanger F, et al: Sequence and organization of the human
mitohondrial genome. Nature. 290:457–465. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Ojala D, Montoya J and Attardi G: tRNA
punctuation model of RNA processing in human mitochondria. Nature.
290:470–471. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Nagaike T, Suzuki T, Katoh T and Ueda T:
Human mitochondrial mRNAs are stabilized with polyadenylation
regulated by mitochondria-specific poly(A) polymerase and
polynucleotide phosphorylase. J Biol Chem. 280:19721–19727. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zhang X, Zuo X, Yang B, Li Z, Xue Y, Zhou
Y, Huang J, Zhao X, Zhou J, Yan Y, et al: MicroRNA directly
enhances mitochondrial translation during muscle differentiation.
Cell. 8:607–619. 2014. View Article : Google Scholar
|
|
57
|
Jacquier A: The complex euaryotic
transcriptome: Unexpected pervasive transcription and novel small
RNAs. Nat Rev Genet. 10:833–844. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Artlett CM, Black CM, Briggs DC, Stevens
CO and Welsh KI: Telomere reduction in scleroderma patients: A
possible cause for chromosomal instability. Br J Rheumatol.
35:732–737. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Mercer TR, Neph S, Dinger ME, Crawford J,
Smith MA, Shearwood AM, Haugen E, Bracken CP, Rackham O,
Stamatoyannopoulos JA, et al: The human mitochondrial
transcriptome. Cell. 146:645–658. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Wang G, Chen HW, Oktay Y, Zhang J, Allen
EL, Smith GM, Fan KC, Hong JS, French SW and McCaffery JM: PNPASE
regulates RNA import into mitochondria. Cell. 142:456–467. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Cheng Y, Liu P, Zheng Q, Gao G, Yuan J,
Wang P, Huang J, Xie L, Lu X, Tong T, et al: Mitochondrial
trafficking and processing of telomerase RNA TERC. Cell Rep.
24:2589–2595. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Haendeler J, Hoffmann J, Diehl JF, Vasa M,
Spyridopoulos I, Zeiher AM and Dimmeler S: Antioxidants inhibit
nuclear export of telomerase reverse transcriptase and delay
replicative senescence of endothelial cells. Circ Res. 94:768–775.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Santos JH, Meyer JN, Skorvaga M, Annab LA
and Van Houten B: Mitochondrial hTERT exacerbates
free-radical-mediated mtDNA damage. Aging Cell. 3:399–411. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
De Punder K, Heim C, Wadhwa PD and
Entringer S: Stress and immunosenescence: The role of telomerase.
Psychoneuroendocrinology. 101:87–100. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Vickers M, Brown GC, Cologne JB and
Kyoizumi S: Modelling heaemopoietic stem cell division by analysis
of mutant red cells. Br J Haematol. 2000:54–62. 2000. View Article : Google Scholar
|
|
66
|
Greenwood MJ and Lansdorp PM: Telomeres,
telomerase, and hematopoietic stem cell biology. Arch Med Res.
34:489–495. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Rufer N, Brümmendorf TH, Kolvraa S,
Bischoff C, Christensen K, Wadsworth L, Schulzer M and Lansdorp PM:
Telomere fluorescence measurements in granulocytes and T lymphocyte
subsets point to a high turnover of hematopoietic stem cells and
memory T cells in early childhood. J Exp Med. 190:157–167. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Verfaillie CM, Pera MF and Lansdorp PM:
Stem cells: Hype and reality. Hematology Am Soc Hematol Educ
Program. 2002:369–391. 2002. View Article : Google Scholar
|
|
69
|
Wynn RF, Cross MA, Hatton C, Will AM,
Lashford LS, Dexter TM and Testa NG: Accelerated telomere
shortening in young recipients of allogeneic bone-marrow
transplants. Lancet. 351:178–181. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Scheding S, Ersöz I, Hartmann U, Bartolvic
K, Balabanov S, Salama A, Kanz L and Brümmendorf TH: Peripheral
blood cell telomere length measurements indicate that hematopoietic
stem cell turnover is not significantly increased in whole blood
and apheresis PLT donors. Transfusion. 43:1089–1095. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Weng NP, Levine BL, June CH and Hodes RJ:
Human naive and memory T lymphocytes differ in telomeric length and
replicative potential. Proc Natl Nat Acad Sci USA. 92:11091–11094.
1995. View Article : Google Scholar
|
|
72
|
Jiang Y, Jahagirdar BN, Reinhardt RL,
Schwartz RE, Keene CD, Ortiz-Gonzalez XR, Reyes M, Lenvik T, Lund
T, Blackstad M, et al: Pluripotency of mesenchymal stem cells
derived from adult marrow. Nature. 418:41–49. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Simonsen JL, Rosada C, Serakinci N,
Justesen J, Stenderup K, Rattan SI, Jensen TG and Kassem M:
Telomerase expression extends the proliferative life-span and
maintains the osteogenic potential of human bone marrow stromal
cells. Nat Biotechnol. 20:592–596. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Shi S, Gronthos S, Chen S, Reddi A,
Counter CM, Robey PG and Wang CY: Bone formation by human postnatal
bone marrow stromal stem cells is enhanced by telomerase
expression. Nat Biotechnol. 20:587–591. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Satija NK, Gurudutta GU, Sharma S, Afrin
F, Gupta P, Verma YK, Singh VK and Tripathi RP: Mesenchymal stem
cells: Molecular targets for tissue engineering. Stem Cells Dev.
16:7–23. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Lessard J and Sauvageau G: Bmi-1
determines the proliferative capacity of normal and leukaemic stem
cells. Nature. 423:255–260. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Marx J: Cancer research. Mutant stem cells
may seed cancer. Science. 301:1308–1310. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Armanios M and Greider CW: Telomerase and
cancer stem cells. Cold Spring Harb Symp Quant Biol. 70:205–208.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Zimmermann S and Martens UM: Telomeres,
senescence, and hematopoietic stem cells. Cell Tissue Res.
331:79–90. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Kuhn G, Brustle O, Martens U, Wobus A and
Unsicker K: Stem cells: Established facts, open issues, and future
directions. Cell and Tissue Res. 331:1–3. 2008. View Article : Google Scholar
|
|
81
|
Yegorov YE, Vishnyakova KS, Terekhov SM
and Zelenin AV: Telomerization of diploid fibroblasts does not lead
to substantial changes in cell sensitivity to common cytostatics.
Biologicheskie Membrany. 23:370–375. 2006.
|
|
82
|
Saretzki G, Armstrong L, Leake A, Lako M
and von Zglinicki T: Stress defense in murine embryonic stem cells
is superior to that of various differentiated murine cells. Stem
Cells. 22:962–971. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Atkinson SP, Hoare SF, Glasspool RM and
Keith WN: Lack of telomerase gene expression in alternative
lengthening of telomere cells is associated with chromatin
remodeling of the hTR and hTERT gene promoters. Cancer Res.
65:7585–7590. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Serakinci N, Hoare SF, Kassem M, Atkinson
SP and Keith WN: Telomerase promoter reprogramming and interaction
with general transcription factors in the human mesenchymal stem
cell. Regen Med. 1:125–131. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Richter T, Saretzki G, Nelson G, Melcher
M, Olijslagers S and von Zglinicki T: TRF2 overexpression
diminishes repair of telomeric single-strand breaks and accelerates
telomere shortening in human fibroblasts. Mech Ageing Dev.
128:340–345. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Ito K, Hirao A, Arai F, Takubo K, Matsuoka
S, Miyamoto K, Ohmura M, Naka K, Hosokawa K, Ikeda Y and Suda T:
Reactive oxygen species act through p38 MAPK to limit the lifespan
of hematopoietic stem cells. Nat Med. 12:446–451. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Lonergan T, Brenner C and Bavister B:
Differentiation-related changes in mitochondrial properties as
indicators of stem cell competence. J Cell Physiol. 208:149–153.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Passos JF, von Zglinicki T and Kirkwood
TB: Mitochondria and ageing: Winning and losing in the numbers
game. Bioessays. 29:908–917. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Lee HC, Yin PH, Chi CW and Wei YH:
Increase in mitochondrial mass in human fibroblasts under oxidative
stress and during replicative cell senescence. J Biomed Sci.
9:517–526. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Walter M, Davies JP and Ioannou YA:
Telomerase immortalization upregulates Rab9 expression and restores
LDL cholesterol egress from Niemann-Pick C1 late endosomes. J Lipid
Res. 44:243–253. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Jiang LW, Maher VM, McCormick JJ and
Schindler M: Alkalinization of the lysosomes is correlated with Ras
transformation of murine and human fibroblasts. J Biol Chem.
265:4775–4777. 1990.PubMed/NCBI
|
|
92
|
Martinez-Botas J, Ferruelo AJ, Suarez Y,
Fernandez C, Gomez-Coronado D and Lasuncion MA: Dose-dependent
effects of lovastatin on cell cycle progression. Distinct
requirement of cholesterol and non-sterol mevalonate derivatives.
Biochim Biophys Acta. 1532:185–194. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Condon J, Yin S, Mayhew B, Word RA, Wright
WE, Shay JW and Rainey WE: Telomerase immortalization of human
myometrial cells. Biol Biol Reprod Reprod. 2002:506–514. 2002.
View Article : Google Scholar
|
|
94
|
McKee JA, Banik SS, Boyer MJ, Hamad NM,
Lawson JH, Niklason LE and Counter CM: Human arteries engineered in
vitro. EMBO Rep. 4:633–638. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Klinger RY, Blum JL, Hearn B, Lebow B and
Niklason LE: Relevance and safety of telomerase for human tissue
engineering. Proc Natl Nat Acad Sci USA. 2006:2500–2505. 2006.
View Article : Google Scholar
|
|
96
|
Maximow A: Der Lymphozyt als gemeinsame
Stammzelle der verschiedenen Blutelemente in der embryonalen
Entwicklung und im postfetalen Leben der Saugetiere.
(Demonstrationsvortrag, gehalten in der ausserordentlichen Sitzung
der Berliner, Hamatologischen Gesellschaft am 1. Jun 1909. Folia
Haematol. 8:125–134. 1909.(In German).
|
|
97
|
Simon NM, Smoller JW, McNamara KL, Maser
RS, Zalta AK, Pollack MH, Nierenberg AA, Fava M and Wong KK:
Telomere shortening and mood disorders: Preliminary support for a
chronic stress model of accelerated aging. Biol Psychiatr.
60:432–435. 2006. View Article : Google Scholar
|
|
98
|
Ramirez R, Carracedo J, Soriano S, Jimenez
R, Martin-Malo A, Rodriguez M, Blasco M and Aljama P: Stress
induced-induced premature senescence in mononuclear cells from
patients on long term-term hemodialysis. Am J Kidney Dis.
45:353–359. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Lung FW, Chen NC and Shu BC: Genetic
pathway of major depressive disorder in shortening telomeric
length. Psychiatr Genet. 17:195–199. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Epel ES, Lin J, Wilhelm FH, Wolkowitz OM,
Cawthon R, Adler NE, Dolbier C, Mendes WB and Blackburn EH: Cell
aging in relation to stress arousal and cardiovascular disease risk
factors. Psychoneuroendocrinology. 31:277–287. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Epel ES, Lin J, Dhabhar FS, Wolkowitz OM,
Puterman E, Karan L and Blackburn EH: Dynamics of telomerase
activity in response to acute psychological stress. Brain Behav
Immun. 24:531–539. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Zalli A, Carvalho LA, Lin J, Hamer M,
Erusalimsky JD, Blackburn EH and Steptoe A: Shorter telomeres with
high telomerase activity are associated with raised allostatic load
and impoverished psychosocial resources. Proc Natl Acad Sci USA.
111:4519–4524. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Aschbacher K, O'Donovan A, Wolkowitz A,
Dhabhar OM, Su FS and Epel E: Good stress, bad stress and oxidative
stress: Insights Insights from anticipatory cortisol reactivity.
Psychoneuroendocrinology. 38:1698–1708. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Spivak IM, Mikhelson VM and Spivak DL:
Telomere length, telomerase activity, stress, and aging. Adv
Gerontol. 6:29–35. 2016. View Article : Google Scholar
|
|
105
|
Lansdorp PM: Stress, social rank and
leukocyte telomere length. Aging Cell. 5:583–584. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Adams J, Martin Ruiz-Ruiz C, Pearce MS,
White MS, Parker L and von Zglinicki T: No association between
socio economic-economic status and white blood cell telomere
length. Aging Cell. 6:125–128. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Epel ES: Psychological and metabolic
stress: A recipe for accelerated cellular aging? Hormones. 8:7–22.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
O'Donovan A, Lin J, Tillie J, Dhabhar FS,
Wolkowitz OM, Blackburn EH and Epel ES: Pessimism correlates with
leukocyte telomere shortness and elevated interleukin-6 in
post-menopausal women. Brain Behav Immun. 23:446–449. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Lin J, Epel E and Blackburn E: Telomeres
and lifestyle factors: Roles in cellular aging. Mutat Res.
730:85–89. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Farzaneh-Far R, Lin J, Epel ES, Harris WS,
Blackburn EH and Whooley MA: Association of marine omega-3 fatty
acid levels with telomeric aging in patients with coronary heart
disease. JAMA. 303:250–257. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Tsuboi H, Shimoi K, Kinae N, Oguni I, Hori
R and Kobayashi F: Depressive symptoms are independently correlated
with lipid peroxidation in a female population: Comparison
Comparison with vitamins and carotenoids. J Psychosom Res.
56:53–58. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Forlenza MJ and Miller GE: Increased serum
levels of 8-hydroxy-2′-deoxyguanosine in clinical depression.
Psychosom Med. 68:1–7. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Von Zglinicki T: Oxidative stress shortens
telomeres. Trends Biochem Sci. 27:339–344. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Haendeler J, Hoffmann J, Brandes RP,
Zeiher AM and Dimmeler S: Hydrogen peroxide triggers nuclear export
of telomerase reverse transcriptase via Src kinase family-dependent
phosphorylation of tyrosine 707. Mol Cell Biol. 23:4598–4610. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Romieu I, Garcia-Esteban R, Sunyer J, Rios
C, Alcaraz- Zubeldia M, Velasco SR and Holguin F: The effect of
supplementation with omega-3 polyunsaturated fatty acids on markers
of oxidative stress in elderly exposed to PM(2.5). Environ Environ
Health Perspect. 116:1237–1242. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Graham JE, Christian LM and Kiecolt-Glaser
JK: Stress, age, and immune function: Toward a lifespan approach. J
Behav Med. 29:389–400. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Kiecolt-Glaser JK, Loving TJ, Stowell JR,
Malarkey WB, Lemeshow S, Dickinson SL and Glaser R: Hostile marital
interactions, proinflammatory cytokine production, and wound
healing. Arch Gen Psychiatry. 62:1377–1384. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Liu JP, Chen SM, Cong YS, Nicholls C, Zhou
SF, Tao ZZ and Li H: Regulation of telomerase activity by
apparently opposing elements. Ageing Res Rev. 9:45–256. 2010.
View Article : Google Scholar
|
|
119
|
Tu W, Zhang DK, Cheung PT, Tsao SW and Lau
YL: Effect of insulin-like growth factor 1 on PHA-stimulated cord
blood mononuclear cell telomerase activity. Br J Haematol.
104:785–794. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Haik S, Gauthier LR, Granotier C, Peyrin
JM, Lages CS, Dormont D and Boussin FD: Fibroblast growth factor 2
up regulates telomerase activity in neural precursor cells.
Oncogene. 19:2957–2966. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Kurz DJ, Hong Y, Trivier E, Huang HL,
Decary S, Zang GH, Luscher TF and Erusalimsky JD: Fibroblast growth
factor-2, but not vascular endothelial growth factor, upregulates
telomerase activity in human endothelial cells. Arterioscler Thromb
Vasc Biol. 23:748–754. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Carmeliet P and Jain RK: Angiogenesis in
cancer and other diseases. Nature. 407:249–257. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Trivier E, Kurz DJ, Hong Y, Huang HL and
Erusalimsky JD: Differential regulation of telomerase in
endothelial cells by fibroblast growth factor-2 and vascular
endothelial growth factor-a: Association with replicative life
span. Ann NY Acad Sci USA. 1019:111–115. 2004. View Article : Google Scholar
|
|
124
|
Bermudez Y, Yang H, Saunders BO, Cheng JQ,
Nicosia SV and Kruk PA: VEGF- and LPA-induced telomerase in human
ovarian cancer cells is Sp1-dependent. Gynecol Oncol. 106:526–537.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Nakagawa K, Eitsuka T, Inokuchi H and
Miyazawa T: DNA chip analysis of comprehensive food function:
Inhibition of angiogenesis and telomerase activity with unsaturated
vitamin E Tocotrienol. Biofactors. 21:5–10. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Bermudez Y, Ahmadi S, Lowell NE and Kruk
PA: Vitamin E suppresses telomerase activity in ovarian cancer
cells. Cancer Detect Prev. 31:119–128. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Jacobs EG, Kroenke C, Lin J, Epel ES,
Kenna HA, Blackburn EH and Rasgon NL: Accelerated cell aging in
female APOE-ε4 carriers: Implications Implications for hormone
therapy use. PLoS One. 8:e547132013. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Damjanovic AK, Yang Y, Glaser R,
Kiecolt-Glaser JK, Nguyen H, Laskowski B, Zou Y, Beversdorf DQ and
Weng NP: Accelerated telomere erosion is associated with a
declining immune function of caregivers of Alzheimer's′ disease
patients. J Immunol. 179:4249–4254. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Effros RB: Ageing and the immune system.
Novartis Found Symp. 235:130–149. 2001.PubMed/NCBI
|
|
130
|
Effros RB: Telomerase induction in T
cells: A cure for aging and disease? Exp Gerontol. 42:416–420.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Choi J, Fauce SR and Effros RB: Reduced
telomerase activity in human T-lymphocytes lymphocytes exposed to
cortisol. Brain Behav Immun. 22:600–605. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Ornish D, Lin J, Daubenmier J, Weidner G,
Epel E, Kemp C, Magbanua MJ, Marlin R, Yglecias L, Carroll PR and
Blackburn EH: Increased telomerase activity and comprehensive
lifestyle changes: A pilot study. Lancet Oncol. 9:1048–1057. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Puterman E, Lin J, Blackburn E, O'Donovan
A, Adler N and Epel E: The power of exercise: Buffering the effect
of chronic stress on telomere length. PLoS One. 5:e108372010.
View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Kiefer A, Lin J, Blackburn E and Epel E:
Dietary restraint and telomere length in pre- and postmenopausal
women. Psychosom Med. 70:845–849. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Puterman E, Epel ES, Lin J, Blackburn EH,
Gross JJ, Whooley MA and Cohen BE: Multisystem resiliency moderates
the major depression-telomere length association: Findings findings
from the heart and soul study. Brain Behav Immun. 3:65–73. 2013.
View Article : Google Scholar
|
|
136
|
Wolkowitz OM, Mellon SH, Epel ES, Lin J,
Dhabhar FS, Su Y, Reus VI, Rosser R, Burke HM, Kupferman E, et al:
Leukocyte telomere length in major depression: Correlations with
chronicity, inflammation and oxidative stress-preliminary findings.
PLoS One. 6:178372011. View Article : Google Scholar
|
|
137
|
De Punder K, Heim C, Przesdzing I, Wadhwa
PD and Entringer S: Characterization in humans of in vitro
leucocyte maximal telomerase activity capacity and association with
stress. Philos Trans R Soc B Biol Sci. 373:201604412018. View Article : Google Scholar
|
|
138
|
Bauer ME, Vedhara K, Perks P, Wilcock GK,
Lightman SL and Shanks N: Chronic stress in caregivers of dementia
patients is associated with reduced lymphocyte sensitivity to
glucocorticoids. J Neuroimmunol. 103:84–92. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Buchkovich KJ and Greider CW: Telomerase
regulation during entry into the cell cycle in normal human T
cells. Mol Biol Cell. 7:1443–1454. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Kawauchi K, Ihjima K and Yamada O: IL-2
increases human telomerase reverse transcriptase activity
transcriptionally and posttranslationally through
phosphatidylinositol 3′-kinase/Akt, heat shock protein 90, and
mammalian target of rapamycin in transformed NK cells. J Immunol.
174:5261–5269. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Valenzuela HF and Effros RB: Divergent
telomerase and CD28 expression patterns in human CD4 and CD8 T
cells following repeated encounters with the same antigenic
stimulus. Clin Immunol. 105:117–125. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Jacobs TL, Epel ES, Lin J, Blackburn EH,
Wolkowitz OM, Bridwell DA, Zanesco AP, Aichele SR, Sahdra BK,
MacLean KA, et al: Intensive meditation training, immune cell
telomerase activity, and psychological mediators.
Psychoneuroendocrinology. 36:664–681. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Vasilopoulos E, Fragkiadaki P, Kalliora C,
Fragou D, Docea AO, Vakonaki E, Tsoukalas D, Calina D, Buga AM,
Georgiadis G, et al: The association of female and male infertility
with telomere length (Review). Int J Mol Med. 44:375–389.
2019.PubMed/NCBI
|
|
144
|
Fragkiadaki P, Tsoukalas D, Fragkiadoulaki
I, Psycharakis C, Nikitovic D, Spandidos DA and Tsatsakis A:
Telomerase activity in pregnancy complications (Review). Mol Med
Rep. 14:16–21. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Cohen S, Janicki-Deverts D and Miller GE:
Psychological stress and disease. JAMA. 298:1685–1687. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Monaghan P: Organismal stress, telomeres
and life histories. J Exp Biol. 217:57–66. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Kiecolt-Glaser JK and Glaser R:
Psychological stress, telomeres, and telomerase. Brain Behav Immun.
24:529–530. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Lazarides C, Epel E, Lin J, Blackburn E,
Voelkle M, Buss C, Simhan H, Wadhwa P and Entringer S: Maternal
pro-inflammatory state during pregnancy and newborn leukocyte
telomere length: A prospective investigation. Brain Behav Immun.
80:419–426. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Shivappa N, Steck SE, Hurley TG, Hussey JR
and Hébert JR: Designing and developing a literature-derived,
population based dietary inflammatory index. Public Health Nutr.
17:1689–1696. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Shivappa N, Steck SE, Hurley TG, Hussey
JR, Ma Y, Ockene IS, Tabung F and Hébert JR: A population-based
dietary inflammatory index predicts levels of C-reactive protein in
the Seasonal Variation of Blood Cholesterol Study (SEASONS). Public
Health Nutr. 17:1825–1833. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Shivappa N, Hébert JR, Rietzschel ER, De
Buyzere ML, Langlois M, Debruyne E, Marcos A and Huybrechts I:
Associations between dietary inflammatory index and inflammatory
markers in the Asklepios Study. Br J Nutr. 113:665–671. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Shivappa N, Wirth MD, Hurley TG and Hébert
JR: Association between the dietary inflammatory index (DII) and
telomere length and C-reactive protein from the National Health and
Nutrition Examination Survey-1999-2002. Mol Nutr Food Res.
61:16006302017. View Article : Google Scholar
|
|
153
|
Simopoulos AP: The importance of the
omega-6/omega-3 fatty acid ratio in cardiovascular disease and
other chronic diseases. Exp Biol Med (Maywood). 233:674–688. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Papamichael MM, Shrestha SK, Itsiopoulos C
and Erbas B: The role of fish intake on asthma in children: A
meta-analysis of observational studies. Pediatr Allergy Immunol.
29:350–360. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Kiecolt-Glaser JK, Epel ES, Belury MA,
Andridge R, Lin J, Glaser R, Malarkey WB, Hwang BS and Blackburn E:
Omega-3 fatty acids, oxidative stress, and leukocyte telomere
length: A randomized controlled trial. Brain Behav Immun. 28:16–24.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Tsoukalas D, Fragkiadaki P, Docea AO,
Alegakis AK, Sarandi E, Vakonaki E, Salata E, Kouvid E, Nikitovic
D, Kovatsi L, et al: Association of nutraceutical supplements with
longer telomere length. Int J Mol Med. 44:218–226. 2019.PubMed/NCBI
|
|
157
|
Tsatsakis A, Tsoukalas D, Fragkiadaki P,
Vakonaki E, Tzatzarakis M, Sarandi E, Nikitovic D, Tsilimidos G and
Alegakis AK: Developing BIOTEL: A semi-automated spreadsheet for
estimating telomere length and biological age. Front Genet.
10:842019. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Tsoukalas D, Fragkiadaki P, Docea AO,
Alegakis AK, Sarandi E, Thanasoula M, Spandidos DA, Tsatsakis A,
Razgonova MP and Calina D: Discovery of potent telomerase
activators: Unfolding new therapeutic and anti-aging perspectives.
Mol Med Rep. 20:3701–3708. 2019.PubMed/NCBI
|
|
159
|
Fragkiadaki P, Nikitovic D, Kalliantasi K,
Sarandi E, Thanasoula M, Stivaktakis DP, Nepka C, Spandidos AD,
Tosounidis T and Tsatsakis A: Telomere length and telomerase
activity in osteoporosis and osteoarthritis. Exp Ther Med.
19:1626–1632. 2020.PubMed/NCBI
|
|
160
|
Olovnikov AM: How could the program of
aging be arranged? Russ J Gen Chem. 80:1482–1489. 2010. View Article : Google Scholar
|
|
161
|
Olovnikov AM: Chronographic theory of
development, aging, and origin of cancer: Role of chronomeres and
printomeres. Curr Aging Sci. 8:76–88. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Anisimov VN: Molecular and physiological
mechanisms of aging. 2:(2nd). (St. Petersburg, Nauka). 2003.(In
Russian).
|
|
163
|
Mitteldorf JJ: How does the body know how
old it is? Introducing the epigenetic clock hypothesis.
Biochemistry (Mosc). 78:1048–1053. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Severin FF, Feniouk BA and Skulachev VP:
Advanced glycation of cellular proteins as a possible basic
component of the ‘master biological clock’. Biochemistry (Mosc).
78:1043–1047. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Pisaruk AV: Ontogenetic clock:
Molecular-genetic mechanism. Adv Gerontol. 23:527–535. 2010.(In
Russian). PubMed/NCBI
|
|
166
|
Lund J, Tedesco P, Duke K, Wang J, Kim SK
and Johnson TE: Transcriptional profile of aging in C.
elegans. Curr Biol. 12:1566–1573. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Halaschek-Wiener J, Khattra JS, McKay S,
Pouzyrev A, Stott JM, Yang GS, Holt RA, Jones SJ, Marra MA,
Brooks-Wilson AR and Riddle DL: Analysis of long-lived C.
elegans daf-2 mutants using serial analysis of gene expression.
Genome Res. 15:603–615. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Golden TR and Melov S: Gene expression
changes associated with aging in C. elegans. WormBook: The
Online Review of C. elegans Biology. WormBook. (Pasadena,
CA). 2005-2018.
|
|
169
|
Herndon LA, Schmeissner PJ, Dudaronek JM,
Brown PA, Listner KM, Sakano Y, Paupard MC, Hall DH and Driscoll M:
Stochastic and genetic factors influence tissue-specific decline in
ageing C. elegans. Nature. 419:808–814. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Thaden JJ and Shmookler Reis RJ: Ammonia,
respiration, and longevity in nematodes: Insights on metabolic
regulation of life span from temporal rescaling. J Am Aging Assoc.
23:75–84. 2000.PubMed/NCBI
|
|
171
|
Olovnikov AM: Hypothesis: Lifespan is
regulated by chronomere DNA of the hypothalamus. J Alzheimers Dis.
11:241–252. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Andoniadou CL and Martinez-Barbera JP:
Developmental mechanisms directing early anterior forebrain
specification in vertebrates. Cell Mol Life Sci. 70:3739–3752.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Bayramov AV, Eroshkin FM, Martynova NY,
Ermakova GV, Solovieva EA and Zaraisky AG: Novel functions of
Noggin proteins: Inhibition of Activin/Nodal and Wnt signaling.
Development. 138:5345–5356. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Lewis DA: Development of the prefrontal
cortex during adolescence: Insights into vulnerable neural circuits
in schizophrenia. Neuropsychopharmacology. 16:385–398. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Rzeczkowska PA, Hou H, Wilson MD and
Palmert MR: Epigenetics: A new player in the regulation of
mammalian puberty. Neuroendocrinology. 99:139–155. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Lieber M and Blumenthal HT: Lifespan
changes in the index of cephalization. Exp Aging Res. 10:127–135.
1984. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Arendt T: Alzheimer's disease as a loss of
differentiation control in a subset of neurons that retain immature
features in the adult brain. Neurobiol Aging. 21:783–796. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Park DC, Polk TA, Park R, Minear M, Savage
A and Smith MR: Aging reduces neural specialization in ventral
visual cortex. Proc Natl Acad Sci USA. 101:13091–13095. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
179
|
Voss MW, Erickson KI, Chaddock L, Prakash
RS, Colcombe SJ, Morris KS, Doerksen S, Hu L, McAuley E and Kramer
AF: Dedifferentiation in the visual cortex: An fMRI investigation
of individual differences in older adults. Brain Res. 1244:121–131.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Rajah MN and D'Esposito M: Region-specific
changes in prefrontal function with age: A review of PET and fMRI
studies on working and episodic memory. Brain. 128:1964–1983. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
181
|
Dreher JC, Meyer-Lindenberg A, Kohn P and
Berman KF: Age-related changes in midbrain dopaminergic regulation
of the human reward system. Proc Natl Acad Sci USA.
105:15106–15111. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
182
|
Raz N, Lindenberger U, Rodrigue KM,
Kennedy KM, Head D, Williamson A, Dahle C, Gerstorf D and Acker JD:
Regional brain changes in aging healthy adults: General trends,
individual differences and modifiers. Cereb Cortex. 15:1676–1689.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Desgranges B, Kalpouzos G and Eustache F:
(Cerebral imaging in healthy aging: Contrast with Alzheimer
disease). Rev Neurol (Paris). 164 (Suppl 3):S102–S107. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
184
|
Andrews-Hanna JR, Snyder AZ, Vincent JL,
Lustig C, Head D, Raichle ME and Buckner RL: Disruption of
large-scale brain systems in advanced aging. Neuron. 56:924–935.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
185
|
Fotenos AF, Snyder AZ, Girton LE, Morris
JC and Buckner RL: Normative estimates of cross-sectional and
longitudinal brain volume decline in aging and AD. Neurology.
64:1032–1039. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
186
|
Fotenos AF, Mintun MA, Snyder AZ, Morris
JC and Buckner RL: Brain volume decline in aging: Evidence for a
relation between socioeconomic status, preclinical Alzheimer
disease, and reserve. Arch Neurol. 65:113–120. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
187
|
Yamanishi Y, Hiyama K, Maeda H, Ishioka S,
Murakami T, Hiyama E, Kurose Y, Shay JW and Yamakido M: Telomerase
activity in rheumatoid synovium correlates with the mononuclear
cell infiltration level and disease aggressiveness of rheumatoid
arthritis. J Rheumatol. 25:214–220. 1998.PubMed/NCBI
|
|
188
|
Tarhan F, Vural F, Rosova B, Aksu K,
Cogulu O, Keser G, Gunduz C, Tombuloglu M, Oder C, Karaca E and
Doganavsargil E: Telomerase activity in connective tissue diseases:
Elevated in rheumatoid arthritis, but markedly decreased in
systemic sclerosis. Rheumatol Int. 28:579–583. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
189
|
Kurosaka D, Yasuda J, Yoshida K, Yokoyama
T, Ozawa Y, Obayashi Y, Kingetsu I, Saito S and Yamada A:
Telomerase activity and telomere length of peripheral blood
mononuclear cells in SLE patients. Lupus. 12:591–599. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Hug A, Korporal M, Schröder I, Haas J,
Glatz K, Storch-Hagenlocher B and Wildemann B: Thymic export
function and T cell homeostasis in patients with relapsing
remitting multiple sclerosis. J Immunol. 171:432–437. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
191
|
Jang HS, Oh CK, Jo JH, Kim YS and Kwon KS:
Detection of telomerase activity in psoriasis lesional skin and
correlation with Ki-67 expression and suppression by retinoic acid.
J Korean Med Sci. 16:623–629. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
192
|
Ogoshi M, Le T, Shay JW and Taylor RS: In
situ hybridization analysis of the expression of human telomerase
RNA in normal and pathologic conditions of the skin. J Invest
Dermatol. 110:818–823. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
193
|
Taylor RS, Ramirez RD, Ogoshi M, Chaffins
M, Piatyszek MA and Shay JW: Detection of telomerase activity in
malignant and nonmalignant skin conditions. J Invest Dermatol.
106:759–765. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
194
|
Brümmendorf TH, Maciejewski JP, Mak J,
Young NS and Lansdorp PM: Telomere length in leukocyte
subpopulations of patients with aplastic anemia. Blood. 97:895–900.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
195
|
Fogarty PF, Yamaguchi H, Wiestner A,
Baerlocher GM, Sloand E, Zeng WS, Read EJ, Lansdorp PM and Young
NS: Late presentation of dyskeratosis congenita as apparently
acquired aplastic anaemia due to mutations in telomerase RNA.
Lancet. 362:1628–1630. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
196
|
Ly H: Genetic and environmental factors
influencing human diseases with telomere dysfunction. Int J Clin
Exp Med. 2:114–130. 2009.PubMed/NCBI
|
|
197
|
Young NS: Pathophysiologic mechanisms in
acquired aplastic anemia. Hematology Am Soc Hematol Educ Program.
2006:72–77. 2006. View Article : Google Scholar
|
|
198
|
Yamaguchi H, Calado RT, Ly H, Kajigaya S,
Baerlocher GM, Chanock SJ, Lansdorp PM and Young NS: Mutations in
TERT, the gene for telomerase reverse transcriptase, in aplastic
anemia. N Engl J Med. 352:1413–1424. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
199
|
Marrone A, Stevens D, Vulliamy T, Dokal I
and Mason PJ: Heterozygous telomerase RNA mutations found in
dyskeratosis congenita and aplastic anemia reduce telomerase
activity via haploinsufficiency. Blood. 104:3936–3942. 2004.
View Article : Google Scholar : PubMed/NCBI
|