Introduction
Exosomes are small lipid bilayer-surrounded
extracellular vesicles released from cells into the extracellular
space or biological fluids (1,2). The
biological fluids exosomes are found in abundance in include blood,
saliva, milk, urine, semen, and bile juice (3). These vesicles are carriers of active
or non-autonomous function biomolecules, such as proteins, lipids,
DNA, mRNA and non-coding regulatory RNA. Exosomal markers include
microRNAs like miR-21 and miR-141, plus various proteins that
belong in functional groups such as tetraspanins (CD9, CD63 and
CD81), heat shock proteins (Hsp70, Hsp73 and Hsp90) and membrane
transporters (GTPases) (4).
Exosomes, via their cargo or surface composition, are
signals/mediators of systemic homeostasis and stress for specific
cell-to-cell or tissue-to-tissue communication (5). They are an integral part of the later
phase of the cellular stress response, i.e. the stress-induced
senescence-like phenotype, as well as the senescence-associated
secretory phenotype. Because of their expanding roles in the
endocrine regulation of homeostasis and stress, and their
involvement in human pathophysiology, exosomes are now the
epicenter of a new field, termed ‘exosomics’ (3).
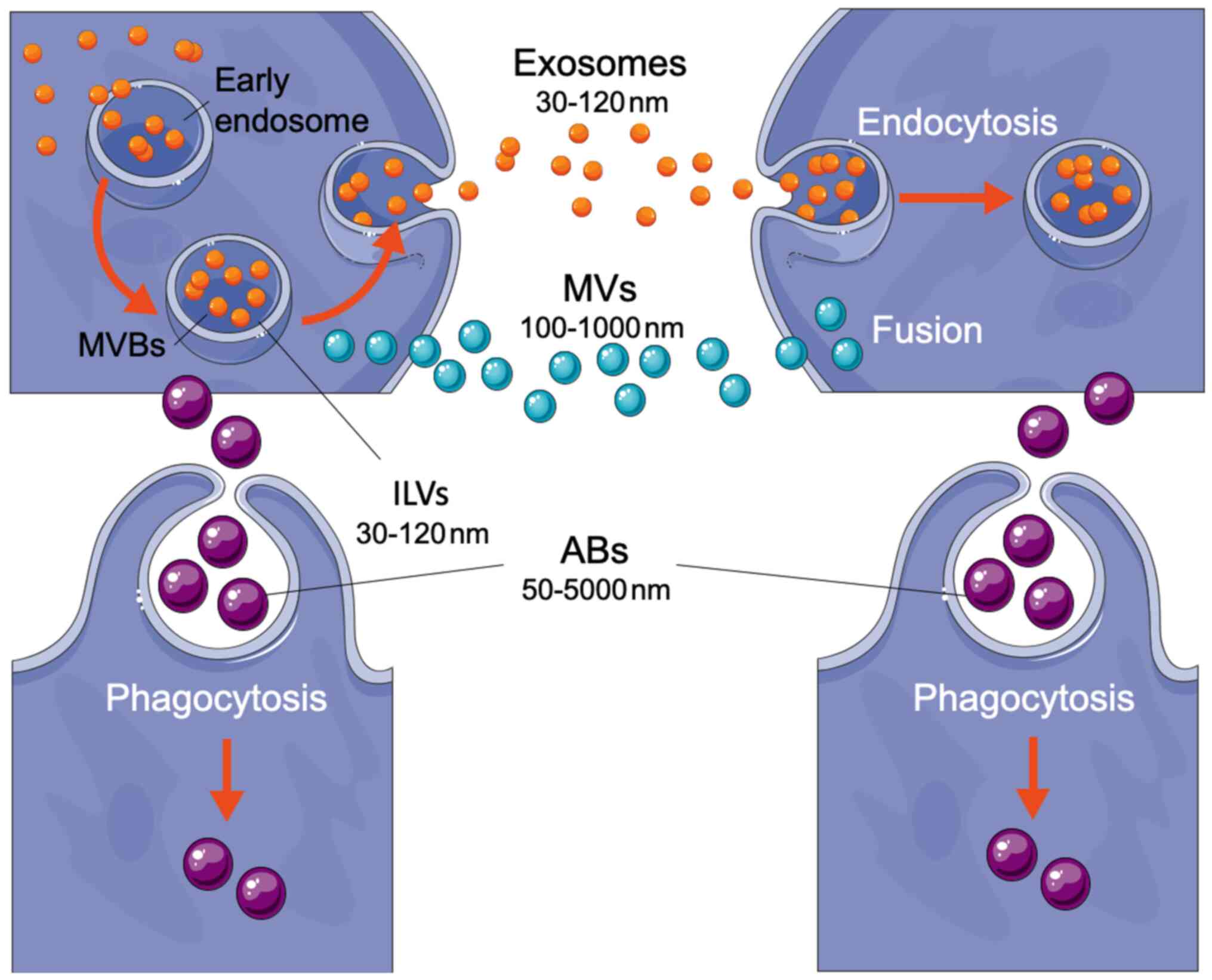
Extracellular vesicles (EVs) comprise a large
spectrum of lipid bilayer-covered micro-particles; based on their
size-range, they can be broadly classified into three distinct
classes: microvesicles (MVs), exosomes and apoptotic
bodies (ABs) (Fig. 1) (6). Microvesicles have a diameter ranging
from 100 nm to 1 µm and are released by cell membrane budding.
Exosomes have a diameter in the range of 30–120 nm. They are
derived by a targeted mechanism from the cell endocytic compartment
and are formed and stored within the intracellular
multivesicular bodies (MVBs). Lastly, apoptotic bodies have
a diameter from 50 nm to 5 µm, the latter a dimension similar to
that of platelets (2–3 µm). They originate from the blebbing of
dying cells (1,7,8).
Exosomes have been heavily researched in the past
few years. Numerous technologies have been developed both for the
isolation and characterization of these vesicles (4). Exosome isolation methods include
differential centrifugation, density gradient centrifugation, size
exclusion chromatography, ultrafiltration, polymer-based
precipitation, immunological separation and isolation by sieving
(4). The characterization
technologies used for exosomes make use of biophysical methods,
such as optical particle tracking that measures from 10 nm to 2 µm,
molecular methods, such as Raman spectrometry that provides the
chemical structure of the exosomes, and microfluidic methods that
make use of immunoisolation and targeted protein analysis of
circulating exosomes (4,9). Exosomes appear to have major roles in
both the physiology and pathophysiology of multicellular organisms
(10). When they are released under
physiological conditions, exosomes are essential for proper immune
system function, while under pathological circumstances, they
potentiate cellular stress and damage (11). Some of the diseases associated with
exosomes are the cardiometabolic and neurodegenerative disorders
and cancer (11). Finally, exosomes
have a significant role in viral infections (12). Virus-infected cells release exosomes
that contain a variety of viral and host cellular factors that can
modify recipient host cell responses (12).
The research on exosomes is expanding the
traditional view on endocrine and neuroendocrine signaling
mechanisms. Traditionally, endocrine mechanisms include specialized
groups of cells, the glands, their molecular mediators, which are
secreted in the general circulation, i.e., the hormones and the
distant target tissues of the latter (13). Hormones are unique chemical
messengers that can be recognized by specific receptors in their
target tissues (13). As exosomes
carry bioactive molecules from donor cells and express surface
proteins that allow them to transfer their contents into their
target cells, they themselves or their cargo may function as
‘hormones’. They can, thus, participate as such in an
autocrine, i.e., local signals between the same type of
cells, paracrine, i.e., local signals between different
types of cells, or endocrine, i.e., distant signals between
any type of cell reaching their target cells via the systemic or
local circulation (14,15).
Therefore, taking into consideration the involvement
of exosomes in current endocrine research, we suggest that they can
provide useful information and lead to a large number of potential
clinical applications. Some of these applications include their use
as biomarkers and as a novel method of message or drug delivery
(16).
An in-depth view of exosomes
Biogenesis
The classification of EVs is mainly based on the
mechanisms of their biogenesis. Exosome biogenesis begins within
the endosomal system, apoptotic bodies are released by cells
undergoing programmed death, i.e. apoptosis, while microvesicles
are derived directly from plasma membrane blebbing (17). Early endosomes grow into late
endosomes by acidification, changes in their protein content, and
increased ability to fuse with other membranes. Late endosomes then
form multivesicular bodies (MVBs) through reverse budding,
during which the endosomal membrane invaginates to form
intraluminal vesicles (ILVs) (18). Multivesicular bodies transport
molecules that have been marked by mono-ubiquitination or by
tetraspanins. The endosomal sorting complexes required for
transport (ESCRT) mediate one of the main mechanisms of
incorporation of molecules into ILVs (19). MVBs may then follow different
pathways: i) directed to lysosomes for content degradation; ii)
transported to the plasma membrane to fuse with it and release the
ILVs outside the cell as exosomes; iii) directed to the plasma
membrane to present antigen-loaded major histocompatibility complex
(MHC) class II molecules; and iv) recycled (18–20).
It appears that different types of cells may be
under conditions and/or exposed to factors that promote the release
of some MVBs as exosomes. Apart from sorting through the
aforementioned ESCRT, another possible mechanism is one in which
cargo is sorted to specific lipid-raft-enriched microdomains that
are associated with ILVs destined for exosome release (20). This possibility is supported by the
fact that post-translational modifications, such as
N-glycosylation, direct proteins to specific microdomains on the
limiting membrane of late endosomes to sort into ILVs also destined
for exosome release (20).
Multivesicular bodies' transport within the cell and
towards the cell membrane is based on interactions with actin and
microtubes of the cytoskeleton (21). Many proteins partake in MVB fusion
with the plasma membrane and subsequent exosome release.
Specifically, the GTPase family of Rab proteins appears to be of
high importance in regulating vesicle tethering and fusion
(22). This protein family action
on exosome release seems to be cell-specific. Different cells
possibly use different Rab proteins to regulate exosome release in
response to membrane fusion (21).
Another group of proteins that may have an important role in
membrane fusion and exosome secretion are the SNAP receptor
proteins (SNARE), which are regulators of vesicle docking and
fusion (22). The involvement of
these proteins in the release of the final exosomes in response to
membrane fusion is through the formation of a SNARE complex
composed of proteins containing three-fourth coiled-coil helices
(21).
Cell-to-cell interactions and the
microenvironment
As already mentioned, the role of exosomes as
generic or specific carriers of signals and regulatory molecules
through a pathway of intercellular vesicle traffic is one of the
main reasons for the explosion of exosome research (23). After their release, a number of set
events characterize exosome function. First, they undergo a change
in their environment, as they move from the cytoplasm and cell
surface to the extracellular fluid. Subsequently, exosomes interact
with the plasma and endocytic membranes of target cells. Last, the
ensuing fusions between exosomes and cell membranes complete the
exosome signaling pathway (24).
All EVs showcase a number of common attributes. On
release, some extracellular vesicles cannot remain intact, their
membranes break down, and their bioactive cargo is expelled into
the extracellular space. The released contents-which may include
bioactive molecules, such as interleukin-1β, growth factors, and
tissue factors-bind their corresponding receptors in adjacent cells
and activate rapid responses (25).
Most EVs, though, maintain their structure and, therefore, survive
in the extracellular fluid, even for long periods of time (24). Extracellular vesicles that persist
in biological fluids may promote degradation of the extracellular
matrix by their transmembrane proteases, an important mechanism for
the circulation of macrophages and for tissue invasion by cancer
cells (25). After their release,
EVs can accumulate in the extracellular spaces close to
intercellular junctions, moving between and through cells so they
may leave the initial fluid and move to adjacent areas or tissues
(24). Later on, these
extracellular vesicles do not interact generally with any type of
cell, but rather showcase a preference for specifically targeted
cells (25). The mechanisms that
dictate this ability of EVs are still under research.
A few mechanisms have been proposed specifically for
exosome-mediated cell-to-cell interaction: i) exosome membrane
proteins may interact with their corresponding membrane receptors
on target cells, activating intracellular signaling; ii) exosome
membrane proteins may be cleaved by proteases, and the resultant
soluble fragments may act as ligands that bind to cell surface
receptors; or iii) exosomes may be internalized by their target
cells, releasing their cargo and activating downstream events
within the receiving cells (26).
In the last-mentioned mechanism, exosome cargo
delivery is responsible for the transport of genetic information
between cells (27). Specifically,
the RNA cargo of exosomes, including both mRNAs and noncoding (nc)
RNAs, seems to be of high importance. This RNA appears to be
functional in target cells and can modulate gene activity and/or
protein production (28).
Particularly, microRNAs, a class of regulatory small endogenous
non-coding RNAs, have been thoroughly researched for their possible
clinical applications (29–31).
Biological processes
Exosome function and heterogeneity depend on the
cell of origin and the conditions at the time of exosome generation
in the originating tissues or cells (32). Exosomes partake in a wide number of
biological processes, beyond inter-cellular communication. Some of
these processes include angiogenesis, antigen presentation,
inflammation, cellular homeostasis, apoptosis, and cell
differentiation (32).
Angiogenesis
Angiogenesis is a process in which new blood vessels
arise from preexisting vasculature (33). Microcirculation is vital for tissue
homeostasis by regulating the supply of oxygen and nutrients and
removing products of cellular metabolism. As exosomes can carry
several biomolecules to target cells and alter their phenotype, it
is not surprising that they can be carriers of pro- or
anti-angiogenic signaling molecules (34). Exosomes derived by mesenchymal stem
cells (MSC-derived exosomes) may carry microRNAs with angiogenic
effect, such as miR-21 which activates the ACT and ERK pathways of
the recipient cell and promotes vascular endothelial growth factor
(VEGF), a signaling molecule which promotes the growth of new blood
vessels (35). On the other hand,
platelet-derived exosomes from human septic patients contain the
p22phox and gp91phox subunits of the NADPH oxidase, which is
associated with endothelial cell apoptosis and dysfunction, thus
showcasing anti-angiogenic effects (34). A pathological condition that
exploits exosome role in angiogenesis is cancer. Cancer cells are
in need of a constant supply of nutrients, oxygen, and growth
factors. Angiogenesis provides new blood vessels that can supply
cancer cells with the requirements mentioned above for survival.
These blood vessels can promote tumor growth and metastasis
formation. Exosomes seem to be mediators of such mechanisms because
tumor cell-derived exosomes seem to promote angiogenesis (34).
Immunity
Exosomes secreted by antigen-presenting cells
(APCs), such as dendritic cells (DCs), carry molecules involved in
antigen presentation, for instance, MHC class I and II molecules
and molecules involved in T-cell stimulation, such as CD86
(36). Particularly, antigens
internalized by DCs and other professional APCs are processed into
peptides and are incorporated into MHC class I (peptide-bound MHC
I/pMHCI) or MHC class II (peptide-bound MHCII/pMHCII) molecules,
which can later be transported to the extracellular space in
extracellular vesicles such as exosomes (37). MHC class II molecules that are found
on antigen-presenting cells-derived exosomes can stimulate
CD4+ T-cells (36).
Specifically, in the case of DC-derived exosomes, antigen-specific
pMHCII found on the surface of the mentioned exosomes can activate
the T-cell antigen receptor on the CD4+ T-cells and
either stimulate them to proliferate directly or indirectly through
the help of additional DCs. The need for additional DCs is
dependent on the abundance of co-stimulatory molecules expressed in
the exosome membrane (37).
CD4+ T-cells have an important role in adaptive
immunity, where they mediate immune responses through the secretion
of specific cytokines (38). MHC
class I molecules found on APC-derived exosomes could potentially
activate CD8+ T-cells (36). Indeed, it has been shown that
DC-derived exosome feature functional pMHCI molecules and can
activate CD8+ T-cell clones either alone or when
incubated with DCs that express allogeneic MHC class I molecules
(39). Naïve CD8+
T-cells can react to pathogens through expansion and
differentiation into cytotoxic effector cells that can wander to
all corners of the body to clear infections (40).
Inflammation
Exosome cargo has been shown to alter recipient cell
functions, including changes in inflammatory responses (41). The above, along with the fact that
different immune cells, such as macrophages, can both produce
exosomes and receive mentioned vesicles, implies an important role
for exosomes in inflammation (41).
Exosomes have been associated with a number of pro-inflammatory
cytokines, such as tumor necrosis factor-alpha (TNF-α),
interleukin-1 beta (IL-1β) and interleukin-6 (IL-6), while they
have also been associated with microRNAs showcasing
anti-inflammatory actin such as miR-155 and miR-146a (41). Moreover, exosomes have also been
associated with inflammation-related molecules, like small heat
shock proteins (HSPs) (42). Some
of these proteins are induced in response to stressful events,
while others are expressed at a constant rate (42).
Apoptosis
The secretion of exosomes plays an essential role in
maintaining cellular homeostasis of exosome-secreting cells
(43). It is speculated that EVs
may remove harmful cytoplasmic DNA from cells. The accumulation of
such DNA causes activation of the cytoplasmic DNA-sensing machinery
that promotes the innate immune response. This accumulation could
lead to activation of reactive oxygen species (ROS)-dependent DNA
damage response, resulting in cell cycle arrest and/or apoptosis in
physiological human cells (43).
Apoptosis is an evolutionarily conserved mechanism
of programmed cell death in multicellular organisms (44). These organisms need to tightly
regulate their number of cells, which is achieved, among other
mechanisms, through coupling of cell division and apoptosis control
(45). Under normal circumstances,
apoptosis helps maintain homeostasis by eliminating unwanted cells
as a response to particular molecular signals. Apoptotic mechanisms
are mainly characterized by condensation of chromatin material, DNA
fragmentation in the nucleus, and cell shrinkage (46). Exosomes can induce cell cycle arrest
and apoptosis in specific cells (32). A pathological condition making use
of this exosomal ability includes exosomes derived from liver
metastases of colorectal cancer. These exosomes can inhibit cell
proliferation and induce T-cell apoptosis by carrying the Fas
ligand (32). Moreover, apoptotic
cells themselves release exosomes with specific attributes
(47), through which they
communicate with neighboring cells. These apoptotic
cell-derived-exosomes seem to regulate inflammation and immune
response (47,48). Specifically, apoptotic cell-derived
exosomes produced from endothelial cells carry ncRNAs, which can
stimulate retinoic acid-inducible gene I-like receptors (RIG-I-like
receptors) and toll-like receptors (TLRs) to promote inflammation,
while apoptotic cell-derived exosomes produced from thymocytes
induces TGFβ in macrophages and suppresses immune response
(48).
Differentiation
Cell differentiation is a vital process in
multicellular organisms and is involved in several biological
functions, such as development, growth and reproduction (49). Exosomes, as significant mediators of
intercellular communication, have been implied to participate in
stem cell differentiation (50). A
prime example is exosomes derived from cells cultured under
osteogenic conditions (osteogenic exosomes), which can be
endocytosed by human marrow-derived stromal cells (HMSCs) and lead
to cell differentiation. Specifically, osteogenic exosomes lead to
an increase in the expression levels of growth factors bone
morphogenetic protein 9 (BMP9) and transforming growth factor β1
(TGFβ1), which have been described as good inducers of osteogenic
differentiation of MSCs (50).
Stress
Living organisms have to cope with various
challenges during their lifespan. In order to cope with such
challenges, living organisms are programmed to maintain an inner
biological equilibrium, between an abiotic (chemical/physical)
environment and the holobiome of all interacting life species. This
inner equilibrium or homeostasis is vital for proper organism
function (51). Unforeseen internal
or external stimuli, the stressors, can threaten homeostasis, and
the state of threatened, or perceived as such, homeostasis is
called stress. Exposure to stressors induces a highly adaptive and
integrated response system, the stress system, whose main goal is
the re-establishment of homeostasis (51,52).
Precise communication among organs and cells is essential in
maintaining homeostasis and responding to stressors. A system that
includes precise communication mechanisms affiliated with the
maintenance of homeostasis is the endocrine system (53). Exosomes are important mediators of
inter-cellular communication and, therefore, partake in mechanisms
related to homeostasis. These vesicles may act as important
components of a complex endocrine system that provides endocrine,
paracrine and autocrine signals (54).
Exosomes have a role in stress physiology (55). Supporting this concept is the fact
that exosomes ubiquitously express membrane proteins that are
associated with stress, like Hsp72, which also exert
pro-inflammatory effects (56). It
has been demonstrated that exposure to acute stressors in the
absence of injury, inflammation, or disease can alter exosome
composition to boost innate immunity (55). Such alterations may include changes
in both exosome membrane proteins and the vesicular microRNA
cargo.
These circulating stress-modified exosomes may have
an immunomodulatory role (55).
Evidence shows that in the absence of stress or pathology, exosomes
generally downregulate immune processes. This may be due to their
microRNA cargo, which includes molecules, such as mir-126, that
exert anti-inflammatory effects (56). Stress exposure, on the other hand,
increases the Hsp72 content and reduces the microRNA cargo of
blood-borne exosomes, changing their role from immunosuppressive to
immunostimulatory (56).
Cell-to-virus interaction
As mentioned above, exosomes have an essential role
in viral pathogenesis and immunity (12). Under physiological conditions,
exosomes can stimulate immune cells and present viral antigens in
response to viral infection (12).
In pathological conditions, viral infection alters the composition
of exosomes and their functions (57). Viruses appear to co-opt exosome
communication through a number of mechanisms that promote viral
propagation. In the case of the hepatitis C virus (HCV), the viral
genome can enter ILVs and be secreted later within exosomes, which,
thus, act as infectious particles (58). In the case of other viruses, such as
the Epstein-Barr virus (EBV), exosomes may carry viral proteins
that promote infectivity (59).
Furthermore, in EBV infections, exosomes derived from infected
cells may carry host proteins that modulate the immune response and
boost infectivity (60). Another
mechanism used by the human immunodeficiency virus (HIV) involves
the incorporation of viral nucleic acid in exosomes, which can
improve and sustain the production of the HIV in infected cells
(59). Last, exosomes derived from
viruses, such as the herpes simplex virus 1 (HSV1), may carry
microRNAs that eliminate production of host proteins associated
with anti-viral responses (59).
An intriguing observation on the connection between
exosomes and viruses is that they display a number of similarities
in their formation, architecture and molecular mechanisms of
action, including the biogenesis and transportation of functional
nucleic acids and proteins (12).
In regards to biogenesis, enveloped viruses, like HIV, make use of
the ESCRT machinery to facilitate viral budding (61). Furthermore, as exosomes and viruses
can deliver active biomolecules from one cell to another, they both
have been implicated in playing a major role in evolution (62).
On the topic of evolution, exosomes may contain
proteins encoded by endogenous retroviruses (ERVs), which are
genetic remnants of ancient retroviral infections (23). None of the human endogenous
retroviruses are truly functional, but some of the proteins they
encode have evolved to execute physiological functions, while
aberrant expression of such proteins has been associated with
various diseases (23). Moreover,
the presence of functional genetic material in exosomes and
viruses, indicates that they may function as potential mediators of
horizontal gene transfer, not only between cells of the same
organism but also across different species (63). All of the above similarities provide
an incentive to further research the connection between exosomes
and viruses, both in an evolutionary and molecular manner.
Disorders of the endocrine system and
exosomes
Dysregulation of the endocrine system has been
associated with a large number of pathological conditions. Some of
these include obesity, type 2 diabetes mellitus (T2DM), disorders
of the reproductive system, and various forms of cancer, such as
breast, testicular, and ovarian cancer (64). Exosomes have been implicated in many
of these and other pathological conditions.
Obesity is a chronic disorder characterized by
excessive accumulation of body fat that increases health risks
(65). Exosomes have a role in
obesity and its metabolic complications through their action as
mediators of communication between adipose tissue, skeletal muscle,
liver and immune cells (66).
Studies have shown that adipose-derived exosome microRNAs are
differentially expressed between lean and obese individuals
(66). Furthermore, research in
animal models of obesity indicates that treatment of lean mice with
exosomes containing obesity-associated microRNA mimics, induces
central obesity and hepatic steatosis (67). A suggested mechanism via which
obesity leads to health complications is through the induction of
inflammation in major organs, including adipose tissue and tissues
of the cardiovascular system (66).
Exosomes may play a role in these mechanisms, as they have both
pro-inflammatory and anti-inflammatory properties (55,56).
Type 2 diabetes mellitus is a metabolic disease that
has been associated with both obesity and presence of exosomes
(68). This disease is
characterized by hyperglycemia, insulin resistance, and insulin
deficiency (69). T2DM is
associated with the metabolic syndrome, a condition related to
obesity and characterized by insulin resistance, dyslipidemia,
hypertension and blood hypercoagulability (70). Exosomes released from
obesity-related adipose tissue contain different microRNA cargo
compared to those secreted by healthy adipose tissue and seem to
significantly decrease insulin sensitivity and glucose uptake
(71). The mechanisms via which
these abnormal exosomes influence T2DM may include activation of
inflammation, downregulation of glucose transporter type 4, and
disruption of insulin receptor signaling (72).
Exosomes have also been implicated in a number of
reproduction-related pathologies, like endometriosis and the
polycystic ovary syndrome (PCOS) (73). Endometriosis is a gynecological
disorder characterized by the presence of tissue that resembles the
endometrium outside the uterine cavity (74). In this condition, studies implicate
stromal endometrial-derived exosomes, which modulate the immune
surveillance response, and allow ectopic endometrial tissue to
attach to peritoneal surfaces. This ectopic endometrial tissue may
continue to release exosomes that help it evade the immune system
and survive (75).
Polycystic ovary syndrome is a common gynecological
metabolic and reproductive disorder associated with menstrual
dysfunction, androgen excess and decreased fertility (76). Exosomes derived from human
follicular fluid display differences in their microRNA cargo
between PCOS patients and controls (77). Bioinformatic analyses indicate that
these differences may result in alterations in amino acid
biosynthesis and metabolism (77).
An increase in exosome release and alterations in
their composition may promote cancer progression and metastases
(78). Specifically, exosomes
released from tumor cells seem to have an essential role in cancer.
These exosomes induce oncogenesis, reprogram stromal cells,
modulate the immune system, and, as already mentioned above,
promote angiogenesis (79).
Particularly, tumor-derived exosomes can carry oncogenic and
anti-apoptotic molecules among cells, promoting oncogenesis
(79). Additionally, tumor-derived
exosomes play an essential role in transforming normal stromal
cells to cancer-associated fibroblasts through transforming growth
factor beta (TGFβ) signaling (80).
Cancer-associated fibroblasts are key regulators of the tumor
microenvironment and can modulate cancer metastases, immune system
function, production of growth factors, and angiogenesis (81). Last, tumor-derived exosomes can act
directly on immune cells to produce immunosuppression (80).
Exosome-based personalized medical
applications
Precision medicine's goal is to provide the
appropriate treatment to the fitting patient at the right time. In
precision medicine, molecular factors may be used to provide
precise diagnosis and, thus, lead to a more accurate treatment
(82). Exosomes can be considered
such factors with some of their potential applications, including
their use as biomarkers or therapeutic biomolecule carriers
(83). Biomarkers are cellular,
biochemical, or molecular parameters that can be objectively
measured to serve as accurate indicators of a biological state
(84).
The key attribute exosomes showcase that can be
exploited in therapeutic applications is that their origin dictates
their cargo and role in cell signaling (85,86).
Levels of exosome population subgroups can reflect many aspects of
cell-to-cell communication, such as the state of cellular
homeostasis, the targeted pathway and tissue, and the dynamics of
the extracellular environment. Furthermore, exosome cargo contains
a mixture of active signals and non-autonomous genetic information
protected by the lipid bilayer of the vesicle. Finally, exosomes
are highly stable and, thus, can regulate the stress response at a
systemic level by storing and transferring enriched amounts of
regulatory information, as well as markers of stress, while
retaining a cell- and state-type of specificity for low abundant
proteins and RNAs (86), which is
related to their cell of origin and the function(s) that they
mediate.
Exosomes can be used as therapeutic agents due to
their exceptional biocompatibility and bio-specificity, both as
vehicles and as cargo, especially when combined with
patient-derived embryonic stem cells (hESCs) (87). A prime example of ESC-derived
exosomes showcasing therapeutic abilities analogous to their cell
of origin is that of mesenchymal stem cell derived exosomes
(88). MSC-derived exosomes can be
used in tissue injury repair and can reduce the inherent health
risks related to administering viable cells (88). On the other hand, exosomes can be
modified to deliver drugs and vaccines. A prime example is loading
exosomes with anticancer agents, an approach that has been shown to
have therapeutic effects in animal models (89). Last, it should be mentioned that the
diverse abilities of exosomes should be taken into consideration
when they are used as therapeutic agents, as they may have adverse
effects on a patient's physiological mechanisms, including those of
the immune system, intermediary metabolism, and others (90).
Conclusion
Exosomes have a vital and largely unknown role in
inter-cellular signaling. These vesicles seem to partake in a large
number of biological functions, while they have also been
associated with several diseases. It is, therefore, not surprising
that there has been a high interest in exosome research over the
recent years. Endocrinology encompasses most branches of biology
and medicine and we can now add exosomes as endocrine cell-to-cell
communication components. Thus, exosomal endocrinology represents
yet another aspect of endocrinology, an essential field of research
that may help improve understanding, prognosis, diagnosis, and
treatment of many diseases.
Acknowledgements
Not applicable.
Funding
Funding was received from Microsoft Azure for
Genomics Research (grant no. CRM:0740983), Amazon Web Services
Cloud for Genomics Research (grant no. 309211522729),
AdjustEBOVGP-Dx (grant no. RIA2018EF-2081; Biochemical Adjustments
of native EBOV Glycoprotein in Patient Sample to Unmask target
Epitopes for Rapid Diagnostic Testing), a European & Developing
Countries Clinical Trials Partnership (EDCTP2) under the Horizon
2020 ‘Research and Innovation Actions’ DESCA.
Availability of data and materials
Not applicable.
Authors' contributions
DV and GPC conceived of the current article. DV, TM,
NN, AE, AG, FB and GPC have all contributed to the writing,
drafting, revising, editing and reviewing of the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests
References
|
1
|
Doyle LM and Wang MZ: Overview of
extracellular vesicles, their origin, composition, purpose, and
methods for exosome isolation and analysis. Cells. 8:7272019.
View Article : Google Scholar
|
|
2
|
Lawson C, Vicencio JM, Yellon DM and
Davidson SM: Microvesicles and exosomes: New players in metabolic
and cardiovascular disease. J Endocrinol. 228:R57–R71. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Patel GK, Khan MA, Zubair H, Srivastava
SK, Khushman Md, Singh S and Singh AP: Comparative analysis of
exosome isolation methods using culture supernatant for optimum
yield, purity and downstream applications. Sci Rep. 9:53352019.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yakimchuk K: Exosomes: Isolation and
characterization methods and specific markers. Mater Methods.
5:14502015.//dx.doi.org/10.13070/mm.en.5.1450. View Article : Google Scholar
|
|
5
|
Maacha S, Bhat AA, Jimenez L, Raza A,
Haris M, Uddin S and Grivel JC: Extracellular vesicles-mediated
intercellular communication: Roles in the tumor microenvironment
and anti-cancer drug resistance. Mol Cancer. 18:552019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yanez-Mo M, Siljander PR, Andreu Z, Zavec
AB, Borras FE, Buzas EI, Buzas K, Casal E, Cappello F, Carvalho J,
et al: Biological properties of extracellular vesicles and their
physiological functions. J Extracell Vesicles. 4:270662015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Villa F, Quarto R and Tasso R:
Extracellular vesicles as natural, safe and efficient drug delivery
systems. Pharmaceutics. 11:5572019. View Article : Google Scholar
|
|
8
|
Zarka R, Horev MB, Volberg T, Neubauer S,
Kessler H, Spatz JP and Geiger B: Differential modulation of
platelet adhesion and spreading by adhesive ligand density. Nano
Lett. 19:1418–1427. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
He M, Crow J, Roth M, Zeng Y and Godwin
AK: Integrated immunoisolation and protein analysis of circulating
exosomes using microfluidic technology. Lab Chip. 14:3773–3780.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Camussi G, Deregibus MC, Bruno S,
Cantaluppi V and Biancone L: Exosomes/microvesicles as a mechanism
of cell-to-cell communication. Kidney Int. 78:838–848. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Isola AL and Chen S: Exosomes: The
messengers of health and disease. Curr Neuropharmacol. 15:157–165.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Crenshaw BJ, Gu L, Sims B and Matthews QL:
Exosome biogenesis and biological function in response to viral
infections. Open Virol J. 12:134–148. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nussey SS and Whitehead SA: Endocrinology:
An Integrated Approach. CRC Press; 2001, View Article : Google Scholar
|
|
14
|
Thery C, Ostrowski M and Segura E:
Membrane vesicles as conveyors of immune responses. Nat Rev
Immunol. 9:581–593. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zduriencikova M, Gronesova P, Cholujova D
and Sedlak J: Potential biomarkers of exosomal cargo in endocrine
signaling. Endocr Regul. 49:141–50. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Qin J and Xu Q: Functions and application
of exosomes. Acta Pol Pharm. 71:537–543. 2014.PubMed/NCBI
|
|
17
|
Willms E, Cabañas C, Mäger I, Wood MJA and
Vader P: Extracellular vesicle heterogeneity: Subpopulations,
isolation techniques, and diverse functions in cancer progression.
Front Immunol. 9:7382018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hessvik NP and Llorente A: Current
knowledge on exosome biogenesis and release. Cell Mol Life Sci.
75:193–208. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Turegano-Lopez M, Santuy A, DeFelipe J and
Merchan-Perez A: Size, shape, and distribution of multivesicular
bodies in the juvenile rat somatosensory cortex: A 3D electron
microscopy study. Cereb Cortex. 30:18872020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tang Y and Dawn B: Mesenchymal Stem Cell
Derived Exosomes: The Potential for Translational Nanomedicine. 1st
edition. Academic Press; 2015
|
|
21
|
Catalano M and O'Driscoll L: Inhibiting
extracellular vesicles formation and release: A review of EV
inhibitors. J Extracell Vesicles. 9:17032442020. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Beer KB and Wehman AM: Mechanisms and
functions of extracellular vesicle release in vivo: What we can
learn from flies and worms. Cell Adh Migr. 11:135–50. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pegtel DM and Gould SJ: Exosomes. Annu Rev
Biochem. 88:487–514. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Meldolesi J: Exosomes and ectosomes in
intercellular communication. Curr Biol. 28:R435–R44. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cocucci E and Meldolesi J: Ectosomes and
exosomes: Shedding the confusion between extracellular vesicles.
Trends Cell Biol. 25:364–372. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Urbanelli L, Magini A, Buratta S, Brozzi
A, Sagini K, Polchi A, Tancini B and Emiliani C: Signaling pathways
in exosomes biogenesis, secretion and fate. Genes (Basel).
4:152–170. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
de la Torre Gomez C, Goreham RV, Bech
Serra JJ, Nann T and Kussmann M: ‘Exosomics’: A review of
biophysics, biology and biochemistry of exosomes with a focus on
human breast milk. Front Genet. 9:922018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Valadi H, Ekstrom K, Bossios A, Sjostrand
M, Lee JJ and Lotvall JO: Exosome-mediated transfer of mRNAs and
microRNAs is a novel mechanism of genetic exchange between cells.
Nat Cell Biol. 9:654–659. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Huang Y, Shen XJ, Zou Q, Wang SP, Tang SM
and Zhang GZ: Biological functions of microRNAs: A review. J
Physiol Biochem. 67:129–139. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang M, Yu F, Ding H, Wang Y, Li P and
Wang K: Emerging function and clinical values of exosomal MicroRNAs
in cancer. Mol Ther Nucleic Acids. 16:791–804. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Beuzelin D and Kaeffer B: Exosomes and
miRNA-loaded biomimetic nanovehicles, a focus on their potentials
preventing type-2 diabetes linked to metabolic syndrome. Frontiers
in immunology. 9:27112018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gurunathan S, Kang MH, Jeyaraj M, Qasim M
and Kim JH: Review of the isolation, characterization, biological
function, and multifarious therapeutic approaches of exosomes.
Cells. 8:3072019. View Article : Google Scholar
|
|
33
|
Tahergorabi Z and Khazaei M: A review on
angiogenesis and its assays. Iran J Basic Med Sci. 15:1110–1126.
2012.PubMed/NCBI
|
|
34
|
Ribeiro MF, Zhu H, Millard RW and Fan GC:
Exosomes function in pro- and anti-angiogenesis. Curr Angiogenes.
2:54–59. 2013.PubMed/NCBI
|
|
35
|
Zimta AA, Baru O, Badea M, Buduru SD and
Berindan-Neagoe I: The role of angiogenesis and pro-angiogenic
exosomes in regenerative dentistry. Int J Mol Sci. 20:4062019.
View Article : Google Scholar
|
|
36
|
Pawankar R, Holgate S and Rosenwasser L:
Allergy Frontiers: Future Perspectives. 6. Springer; 2010
|
|
37
|
Leone DA, Rees AJ and Kain R: Dendritic
cells and routing cargo into exosomes. Immunol Cell Biol.
2018.(Ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Luckheeram RV, Zhou R, Verma AD and Xia B:
CD4+ T cells: Differentiation and functions. Clin Dev
Immunol. 2012:9251352012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Théry C, Ostrowski M and Segura E:
Membrane vesicles as conveyors of immune responses. Nat Rev
Immunol. 9:581–593. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhang N and Bevan MJ: CD8+ T
cells: Foot soldiers of the immune system. Immunity. 35:161–168.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chan BD, Wong WY, Lee MM, Cho WC, Yee BK,
Kwan YW and Tai WC: Exosomes in inflammation and inflammatory
disease. Proteomics. 19:e18001492019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Reddy VS, Madala SK, Trinath J and Reddy
GB: Extracellular small heat shock proteins: Exosomal biogenesis
and function. Cell Stress Chaperones. 23:441–454. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Takahashi A, Okada R, Nagao K, Kawamata Y,
Hanyu A, Yoshimoto S, Takasugi M, Watanabe S, Kanemaki MT, Obuse C
and Hara E: Exosomes maintain cellular homeostasis by excreting
harmful DNA from cells. Nat Commun. 8:152872017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Singh R, Letai A and Sarosiek K:
Regulation of apoptosis in health and disease: The balancing act of
BCL-2 family proteins. Nat Rev Mol Cell Biol. 20:175–193. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Alberts B: Molecular biology of the cell,
6th edition. CRC Press; 2017
|
|
46
|
Jan R and Chaudhry GE: Understanding
apoptosis and apoptotic pathways targeted cancer therapeutics. Adv
Pharm Bull. 9:205–218. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Caruso S and Poon IKH: Apoptotic
cell-derived extracellular vesicles: More than just debris. Front
Immunol. 9:14862018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kakarla R, Hur J, Kim YJ, Kim J and Chwae
YJ: Apoptotic cell-derived exosomes: Messages from dying cells. Exp
Mol Med. 52:1–6. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Sanchez Alvarado A and Yamanaka S:
Rethinking differentiation: Stem cells, regeneration, and
plasticity. Cell. 157:110–119. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Narayanan R, Huang CC and Ravindran S:
Hijacking the cellular mail: Exosome mediated differentiation of
mesenchymal stem cells. Stem Cells Int. 2016:38086742016.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Chrousos GP: Stress and disorders of the
stress system. Nat Rev Endocrinol. 5:374–381. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Nicolaides NC, Kyratzi E,
Lamprokostopoulou A, Chrousos GP and Charmandari E: Stress, the
stress system and the role of glucocorticoids.
Neuroimmunomodulation. 22:6–19. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Hiller-Sturmhofel S and Bartke A: The
endocrine system: An overview. Alcohol Health Res World.
22:153–164. 1998.PubMed/NCBI
|
|
54
|
Zhang HG and Grizzle WE: Exosomes: A novel
pathway of local and distant intercellular communication that
facilitates the growth and metastasis of neoplastic lesions. Am J
Pathol. 184:28–41. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Beninson LA and Fleshner M: Exosomes: An
emerging factor in stress-induced immunomodulation. Semin Immunol.
26:394–401. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Fleshner M and Crane CR: Exosomes, DAMPs
and miRNA: Features of stress physiology and immune homeostasis.
Trends Immunol. 38:768–776. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Anderson M, Kashanchi F and Jacobson S:
Role of exosomes in human retroviral mediated disorders. J
Neuroimmune Pharmacol. 13:279–291. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Anderson MR, Kashanchi F and Jacobson S:
Exosomes in viral disease. Neurotherapeutics. 13:535–546. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Urbanelli L, Buratta S, Tancini B, Sagini
K, Delo F, Porcellati S and Emiliani C: The role of extracellular
vesicles in viral infection and transmission. Vaccines (Basel).
7:1022019. View Article : Google Scholar
|
|
60
|
Keryer-Bibens C, Pioche-Durieu C,
Villemant C, Souquere S, Nishi N, Hirashima M, Middeldorp J and
Busson P: Exosomes released by EBV-infected nasopharyngeal
carcinoma cells convey the viral latent membrane protein 1 and the
immunomodulatory protein galectin 9. BMC Cancer. 6:2832006.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Votteler J and Sundquist WI: Virus budding
and the ESCRT pathway. Cell Host Microbe. 14:232–241. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Nolte-'t Hoen E, Cremer T, Gallo RC and
Margolis LB: Extracellular vesicles and viruses: Are they close
relatives? Proc Natl Acad Sci USA. 113:9155–9161. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Kawamura Y, Yamamoto Y, Sato TA and Ochiya
T: Extracellular vesicles as trans-genomic agents: Emerging roles
in disease and evolution. Cancer Sci. 108:824–830. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Maqbool F, Mostafalou S, Bahadar H and
Abdollahi M: Review of endocrine disorders associated with
environmental toxicants and possible involved mechanisms. Life Sci.
145:265–273. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Agha M and Agha R: The rising prevalence
of obesity: part A: Impact on public health. Int J Surg Oncol (N
Y). 2:e172017. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Kim A, Shah AS and Nakamura T:
Extracellular Vesicles: A potential novel regulator of obesity and
its associated complications. Children (Basel). 5:1522018.
|
|
67
|
Castaño C, Kalko S, Novials A and Párrizas
M: Obesity-associated exosomal miRNAs modulate glucose and lipid
metabolism in mice. Proc Natl Acad Sci USA. 115:12158–12163. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Zhang B, Yang Y, Xiang L, Zhao Z and Ye R:
Adipose-derived exosomes: A novel adipokine in obesity-associated
diabetes. J Cell Physiol. 234:16692–16702. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Olokoba AB, Obateru OA and Olokoba LB:
Type 2 diabetes mellitus: A review of current trends. Oman Med J.
27:269–273. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Nath D, Heemels MT and Anson L: Obesity
and diabetes. Nature. 444:8392006. View Article : Google Scholar
|
|
71
|
Dang SY, Leng Y, Wang ZX, Xiao X, Zhang X,
Wen T, Gong HZ, Hong A and Ma Y: Exosomal transfer of obesity
adipose tissue for decreased miR-141-3p mediate insulin resistance
of hepatocytes. Int J Biol Sci. 15:351–368. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Xiao Y, Zheng L, Zou X, Wang J, Zhong J
and Zhong T: Extracellular vesicles in type 2 diabetes mellitus:
Key roles in pathogenesis, complications, and therapy. J Extracell
Vesicles. 8:16256772019. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Simon C, Greening DW, Bolumar D, Balaguer
N, Salamonsen LA and Vilella F: Extracellular vesicles in human
reproduction in health and disease. Endocr Rev. 39:292–332. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Zondervan KT, Becker CM, Koga K, Missmer
SA, Taylor RN and Vigano P: Endometriosis. Nat Rev Dis Primers.
4:92018. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Schjenken JE, Panir K, Robertson SA and
Hull ML: Exosome-mediated intracellular signalling impacts the
development of endometriosis-new avenues for endometriosis
research. Mol Hum Reprod. 25:2–4. 2018. View Article : Google Scholar
|
|
76
|
El Hayek S, Bitar L, Hamdar LH, Mirza FG
and Daoud G: Poly cystic ovarian syndrome: An updated overview.
Front Physiol. 7:1242016. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Hu J, Tang T, Zeng Z, Wu J, Tan X and Yan
J: The expression of small RNAs in exosomes of follicular fluid
altered in human polycystic ovarian syndrome. PeerJ. 8:e86402020.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Rajagopal C and Harikumar KB: The origin
and functions of exosomes in cancer. Front Oncol. 8:662018.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Tai YL, Chen KC, Hsieh JT and Shen TL:
Exosomes in cancer development and clinical applications. Cancer
Sci. 109:2364–3674. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Tung KH, Ernstoff MS, Allen C and an Shu
SL: A Review of Exosomes and their Role in The Tumor
Microenvironment and Host-Tumor ‘Macroenvironment’. J Immunol Sci.
3:4–8. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Sahai E, Astsaturov I, Cukierman E,
DeNardo DG, Egeblad M, Evans RM, Fearon D, Greten FR, Hingorani SR,
Hunter T, et al: A framework for advancing our understanding of
cancer-associated fibroblasts. Nat Rev Cancer. 20:174–186. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Currie G and Delles C: Precision medicine
and personalized medicine in cardiovascular disease. Adv Exp Med
Biol. 1065:589–605. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Zhang Y, Liu Y, Liu H and Tang WH:
Exosomes: Biogenesis, biologic function and clinical potential.
Cell Biosci. 9:192019. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Mayeux R: Biomarkers: Potential uses and
limitations. NeuroRx. 1:182–188. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Sancho-Albero M, Navascués N, Mendoza G,
Sebastián V, Arruebo M, Martín-Duque P and Santamaría J: Exosome
origin determines cell targeting and the transfer of therapeutic
nanoparticles towards target cells. J Nanobiotechnology. 17:162019.
View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Boukouris S and Mathivanan S: Exosomes in
bodily fluids are a highly stable resource of disease biomarkers.
Proteomics Clin Appl. 9:358–367. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Liu C and Su C: Design strategies and
application progress of therapeutic exosomes. Theranostics.
9:1015–1028. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Zhao T, Sun F, Liu J, Ding T, She J, Mao
F, Xu W, Qian H and Yan Y: Emerging role of mesenchymal stem
cell-derived exosomes in regenerative medicine. Curr Stem Cell Res
Ther. 14:482–494. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Luan X, Sansanaphongpricha K, Myers I,
Chen H, Yuan H and Sun D: Engineering exosomes as refined
biological nanoplatforms for drug delivery. Acta Pharmacol Sin.
38:754–763. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Conlan RS, Pisano S, Oliveira MI, Ferrari
M and Mendes Pinto I: Exosomes as reconfigurable therapeutic
systems. Trends Mol Med. 23:636–650. 2017. View Article : Google Scholar : PubMed/NCBI
|