Introduction
With the development of medical technology, there
has been increased early prevention and diagnosis of cancer,
improved awareness of cancer prevention and significantly reduced
cancer mortality worldwide. Over the past 20 years, the number of
cancer-related mortalities has decreased by ~2.4 million ((1). However, lung cancer remains the number
one cause of mortality by cancer (2). The 5-year survival rate of patients
with lung cancer is only 18%, and early diagnosis of lung cancer
can decrease the mortality rate by 10–50 times. However, most
patients with lung cancer are diagnosed at an advanced stage
(3).
According to the 2017 Global Diabetes Map (8th
edition), ~425 million adults (20–79 years) worldwide suffer from
diabetes, with China accounting for 114.4 million cases, ranking
first in the world (4). Several
studies have reported that diabetes is a risk factor for lung
cancer and leads to poor prognosis (5–7).
Therefore, it is important to identify effective methods to treat
patients with lung cancer with diabetes.
The synuclein γ (SNCG) gene encodes the γ-synuclein
protein that is considered to be the third member of the synuclein
family (α, β and γ), which are reported to be involved in the
pathogenesis of neurodegenerative diseases. SNCG is also known as
breast cancer specific gene 1, and is highly expressed in advanced
breast cancer tissues (8). In
recent years, studies have revealed that SNCG expression is
abnormally high in breast (9,10),
esophageal (11), colorectal
(12), bladder (13), ovarian (9), lung cancer (14) and other malignant tumors. Moreover,
there is close association between SNCG expression and development
of malignant tumors. A study (15)
has shown that SNCG and insulin-like growth factor 1 (IGF-1)
receptor (IGF-1R) are mutually regulated in various types of
tumors, such as breast, hepatoma, colon cancer and lung cancer.
Inhibition of SNCG may inhibit IGF-1/IGF-1R-induced cell
proliferation and migration; furthermore, IGF-1R inhibitors can
block the abnormal expression of SNCG in cancer cells (15).
IGF-1 and IGF-1R are important growth factors in the
body. IGF-1 can be expressed in all parts of the body, while it is
mainly synthesized and secreted by the liver and exerts an
insulin-like effect on tissue cells (16). IGF-1R, which is highly expressed in
most human tumor tissues, can promote the growth and proliferation,
while inhibit the apoptosis of tumor cells (17). Most diabetic patients are
accompanied by hyperinsulinemia, with a certain level of
IGF-binding protein inhibition and relatively increased levels of
free IGF-1 (18). Moreover, IGF-1
is highly expressed in lung, colorectal, breast, prostate cancer
and other cancer types, which promotes the binding of IGF-1 and
IGF-1R, thereby activating the IGF-1/IGF-1R signaling pathway to
promote the development of tumors (19).
Therefore, the present study aimed to investigate
whether SNCG could affect the proliferation of lung cancer cells
induced by high glucose. The results may provide further background
for the treatment of lung cancer, especially for patients with lung
cancer with diabetes.
Materials and methods
Cell culture and high glucose
induction
Lung cancer cell lines (A549, NCI-H1975 and SK-MES-1
cells) were obtained from the American Type Culture Collection. HBE
cells were purchased from Procell Life Science & Technology
Co., Ltd. HBE cells and lung cancer cell lines were cultured in
RPMI-1640 medium (Thermo Fisher Scientific, Inc.) containing 10%
FBS (Gibco; Thermo Fisher Scientific, Inc.) and 1% penicillin in 5%
CO2 at 37°C. HBE cells and lung cancer cells were
treated with 5.5 mM glucose (Beyotime Institute of Biotechnology)
and 25 mM glucose at 37°C for 24 and 48 h. The glucose
concentrations were determined according to previous studies
(20,21).
Cell transfection
Small interfering (si) RNA-SNCG was purchased from
Shanghai GeneChem Co., Ltd. siRNA-SNCG-#1 forward,
5′-GAAUGUUGUACAGAGCGUTT-3′ and reverse,
5′-CACGCUCUGUACAACAUUCTC-3′. siRNA-SNCG-#2 forward,
5′-CCUCUGCCUUGGACACCAUTT-3′ and reverse,
5′-AUGGUGUCCAAGGCAGAGGAG-3′. siRNA NC forward,
5′-UUCUCCGAACGUGUCACGUTT-3′ and reverse,
5′-CUUGAGGCUGUUGUCAUACTT-3′. According to the
Lipofectamine® 2000 specification, lung cancer cells
were transfected with siRNA NC (5 nM), siRNA1 (5 nM) and siRNA2 (5
nM) at 37°C for 24 h and the cell adherence reached ~50% for
transfection using Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.). Subsequent experiments were
performed 24 h after transfection.
Cell Counting Kit-8 (CCK-8) assay
HBE cells and lung cancer cells were seeded into
96-well plates (1×104 cells/well) and respectively
treated with 5.5 mM glucose and 25 mM glucose at 37°C for 24 h and
48 h. A549 cells respectively transfected with siRNA NC and
siRNA-SNCG were treated with 25 mM glucose, and A549 cells were
treated with AXL1717 (10 µM; MedChemExpress) and 25 mM glucose at
37°C for 24 and 48 h. According to the manufacturers instructions,
10 μl CCK-8 solution (Beyotime Institute of Biotechnology) was
added to each well, which was incubated at 37°C for 4 h. The
absorbance at a wavelength of 450 nm for each well was detected
using a ThermoMax microplate reader (Thermo Fisher Scientific,
Inc.) in the dark.
Reverse transcription-quantitative PCR
(RT-qPCR)
Following the indicated treatments,
TRIzol® reagent (Thermo Fisher Scientific, Inc.) was
used to extract total RNA from lung cancer cells. cDNA was
synthesized using the Transcriptor First Strand cDNA Synthesis kit
(Roche Molecular Diagnostics) at 42°C for 30 min. The expression
levels of SNCG, IGF-1 and IGF-1R were determined using the
SYBR-Green Real-Time PCR kit (Beyotime Institute of Biotechnology).
The thermocycling conditions were: 95°C for 25 sec, 95°C for 15
sec, 60°C for 25 sec, 72°C for 25 sec (35 cycles). The primer
sequences were as follows: SNCG forward, 5′-ATGGATGTCTTCAAGAAGGG-3′
and reverse, 5′-CTCTGTACAACATTCTCCTT-3′; IGF-1 forward,
5′-ACGCTCTTCAGTTCGTGTGT-3′ and reverse, 5′-CTTCAGCGGAGCACAGTACA-3′;
IGF-1R forward, 5′-GGTGTCCAATAACTACATTG-3′ and reverse,
5′-CAGCGGTTTGTGGTCCAGC-3′; and GAPDH forward,
5′-GAAGGTGAAGGTCGGAGT-3′ and reverse, 5′-GAAGATGGTGATGGGATT-3′. The
relative expression levels of SNCG, IGF-1 and IGF-1R were
calculated using the 2−ΔΔCq method with GAPDH as a
reference gene (22).
Clone formation assay
Following the indicated treatments, lung cancer
cells were inoculated into a 6-well plate at a density of
1×105 cells/well. The cells were placed in a 37°C cell
incubator for culture. The medium was changed once for 2–3 days.
After 15 days, the cells were washed with PBS three times, fixed
with 4% formaldehyde for 30 min at room temperature, stained with
crystal violet for 20 min at room temperature, washed with PBS and
observed under a light microscope after drying (magnification,
×100).
Wound healing assay
A549 cells were seeded in 6-well plates and A549
cells were treated with 5.5 mM glucose and 25 mM glucose for 48 h.
A549 cells respectively transfected with siRNA NC and siRNA-SNCG
were treated with 25 mM glucose, and A549 cells were treated with
AXL1717 (10 µM; MedChemExpress) and 25 mM glucose at 37°C for 48 h.
Once the cells were adherent, a scratch was made using a 200-µl
sterile pipette tip. Serum-free RPMI-1640 medium was then added for
further culture for 24 h, and wound healing was observed and
photographed under a light microscope magnification, ×100) 24 h
after the scratch was made. The online image analysis system of
WimScratch (Wimasis; http://www.wimasis.com/en/) used to measure the cell
scratch width.
ELISA
Cells were cultured after the indicated treatments
for 24 h and the cell supernatant was collected after
centrifugation (3,000 × g) at 4°C for 15 min. The levels of TNF-α
and IL-6 in cell supernatant were detected using respective TNF-α
(PT518) and IL-6 (PI330) ELISA kits (Beyotime Institute of
Biotechnology) according to the manufacturers instructions.
Western blot analysis
Following the indicated treatments, lung cancer
cells were lysed with cell lysis buffer (Beyotime Institute of
Biotechnology) and an ultrasonic cell disruptor. The cells were
centrifuged at 3,000 × g at 4°C for 15 min and the supernatant was
obtained. Protein concentration was quantified using the protein
concentration determination kit (Bio-Rad Laboratories, Inc.). A
total of 20 µg protein/lane was separated using 10% SDS-PAGE.
Separated proteins were transferred to a PVDF membrane at 100 V and
blocked with 5% skim milk powder for 2 h at room temperature.
Subsequently, membranes were incubated with SNCG (cat. no. ab52633;
1:1,000; Abcam), IGF-1 (cat. no. ab133542; 1:1,000; Abcam), IGF-1R
(cat. no. ab182408; 1:1,000; Abcam), ERK1/2 (cat. no. ab184699;
1:10,000; Abcam), phosphorylated (p)-ERK1/2 (cat. no. ab223500;
1:400; Abcam), c-JNK (cat. no. ab199380; 1:2,500; Abcam) and GAPDH
(cat. no. ab8245; 1:1,000; Abcam) antibodies at 4°C overnight.
Following three washes with TBS-Tween-20 (0.05%), the membranes
were then incubated with horseradish peroxidase-conjugated
secondary antibody (cat. no. 7074; 1:1,000; Cell Signaling
Technology, Inc.) at 25°C for 1 h. Protein bands were visualized
using an enhanced chemiluminescence system (Bio-Rad Clarity Western
EC; Bio-Rad Laboratories, Inc.). Relative protein expression was
quantified using Image-Pro Plus software (version 6.0; Media
Cybernetics, Inc.)
Statistical analysis
All experiments were repeated three times, and
experimental data are presented as the mean ± SD. SPSS 22.0 (IBM
Corp.) was used for statistical analysis of all data, and GraphPad
Prism 6 (GraphPad Software, Inc.) was used for mapping. One-way
ANOVA with Tukeys post hoc test was used for the comparison of mean
values between groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
Proliferation of lung cancer cells
increases after high glucose induction
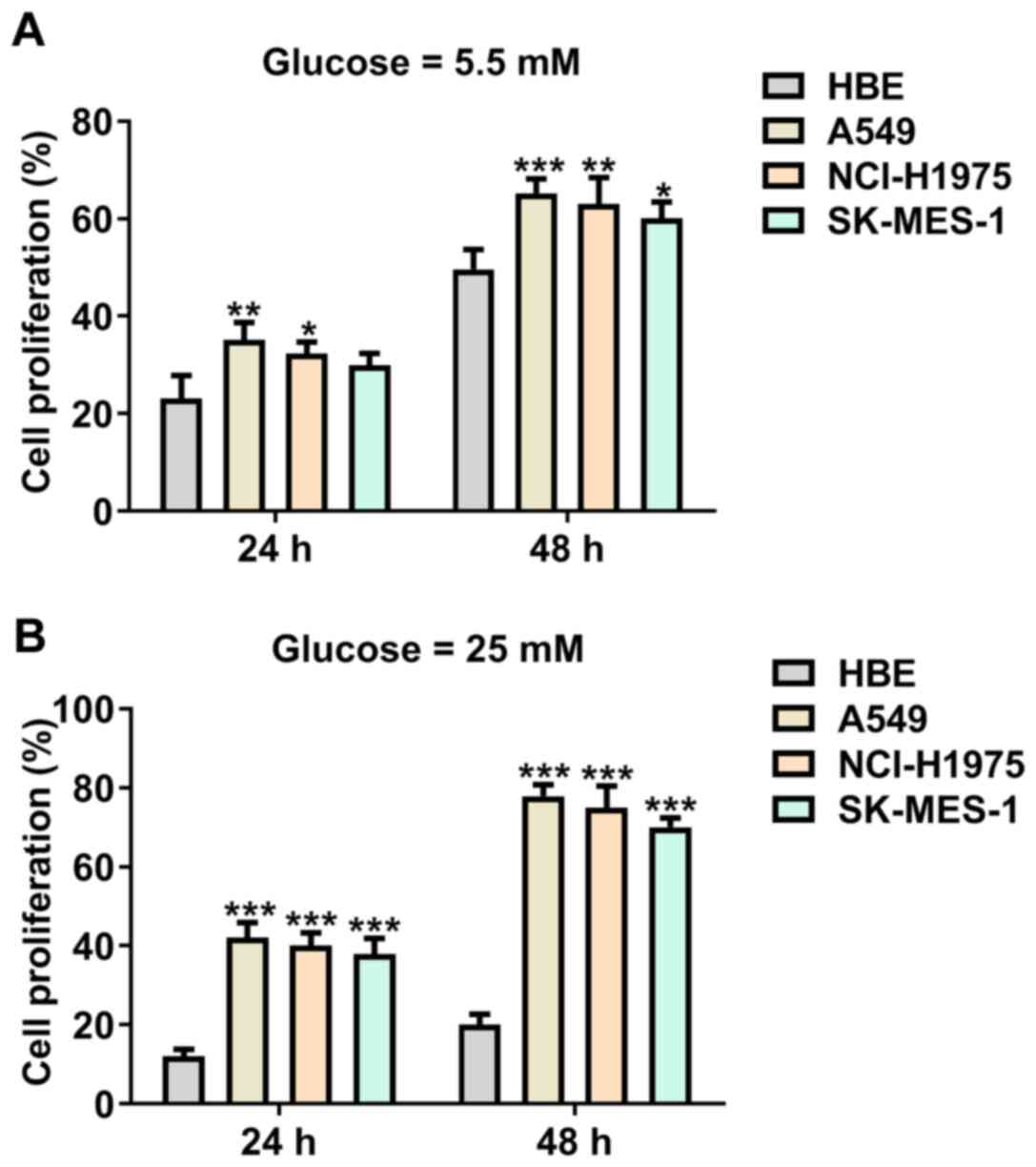
After HBE cells and lung cancer cells were treated
with 5.5 mM glucose, the proliferation of A549 and NCI-H1975 cells
was increased, while SK-MES-1 cell proliferation was not
significantly different compared with HBE cells at 24 h. The
proliferation of A549, NCI-H1975 and SK-MES-1 cells was
significantly increased compared with HBE cells at 48 h (Fig. 1A). As presented in Fig. 1B, treatment of 25 mM glucose
significantly increased the proliferation of A549, NCI-H1975 and
SK-MES-1 cells compared with HBE cells at 24 and 48 h.
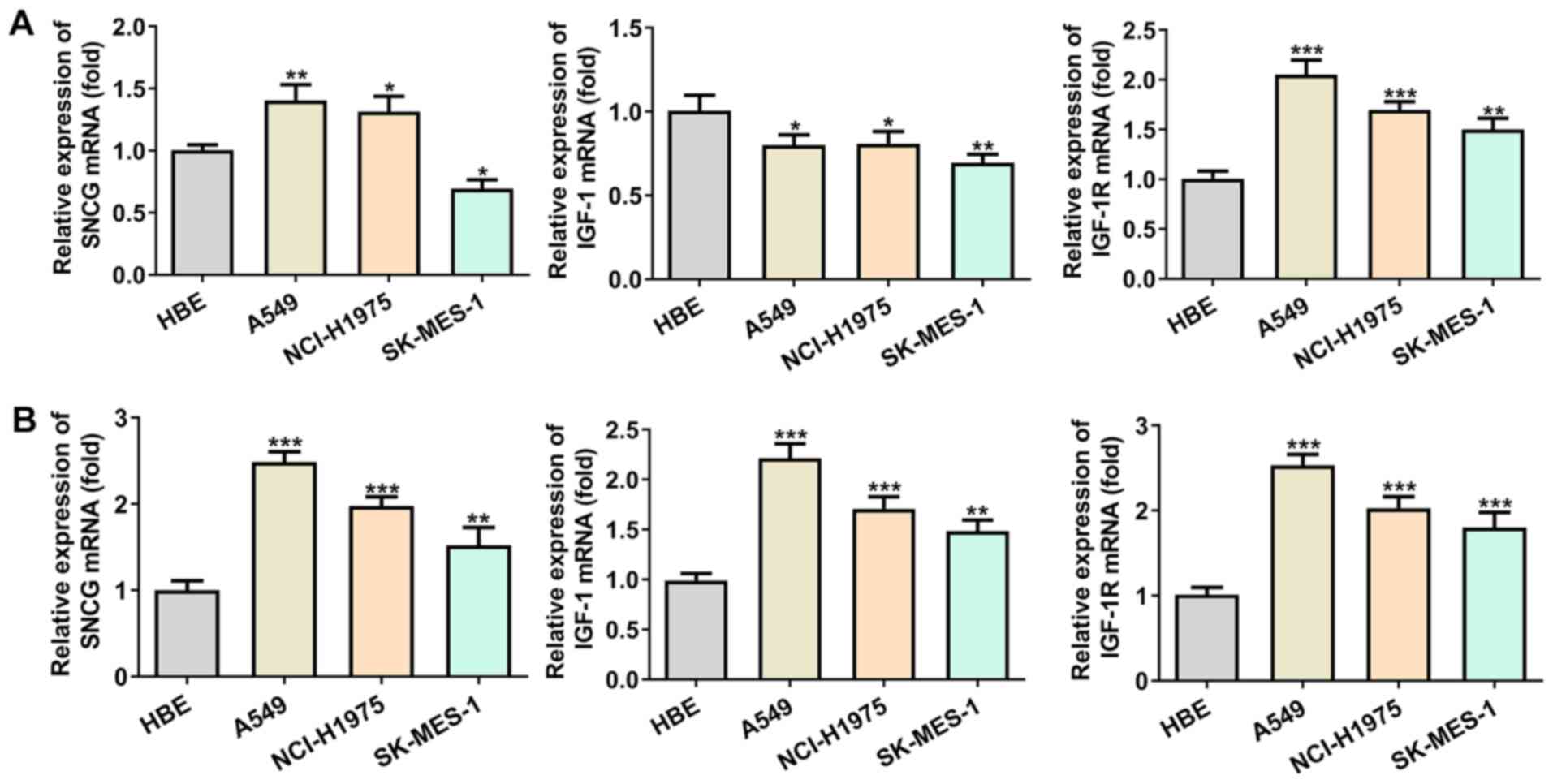
Expression levels of SNCG, IGF-1 and
IGF-1R in lung cancer cells increase after high glucose
induction
SNCG expression was increased in A549 and NCI-H1975
cells, while it was decreased in SK-MES-1 cells. IGF-1 expression
was decreased and that of IGF-1R was increased in A549, NCI-H1975
and SK-MES-1 cells. The trend of SNCG, IGF-1 and IGF-1R expression
levels was different in lung cancer cells compared with HBE cells
(Fig. 2A). Following treatment of
lung cancer cells with high glucose (25 mM), the expression levels
of SNCG, IGF-1 and IGF-1R in lung cancer cells were increased
compared with HBE cells; SNCG, IGF-1 and IGF-1R expression levels
were highest in A549 cells (Fig.
2B). Therefore, A549 cells were chosen for subsequent
experiments.
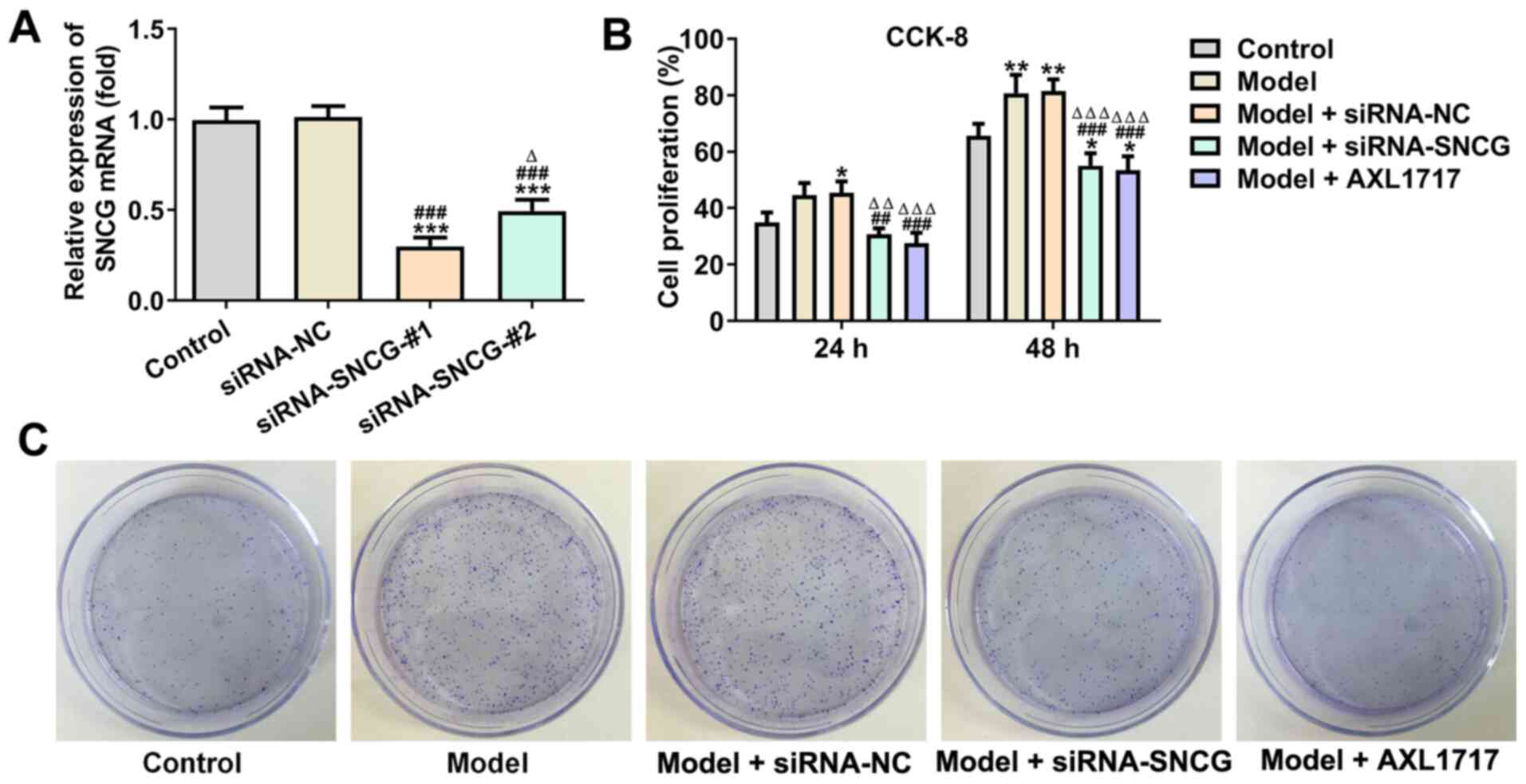
Knockdown of SNCG suppresses the
proliferation and clone formation ability of lung cancer cells
Following transfection of A549 cells with siRNA
negative control (NC), siRNA-SNCG-#1 and siRNA-SNCG-#2, SNCG
expression was downregulated in cells transfected with
siRNA-SNCG-#1 or siRNA-SNCG-#2, while its expression did not
significantly change in cells transfected with siRNA NC. SNCG
expression in A549 cells transfected with siRNA-SNCG-#1 was the
lowest; hence, siRNA-SNCG-#1 was selected for subsequent
experiments (Fig. 3A). After A549
cells were treated with high glucose (25 mM), A549 cell
proliferation was enhanced, which was not significantly affected by
siRNA NC transfection. Knockdown of SNCG suppressed the
proliferation of A549 cells, and the same effects were observed
following AXL1717 treatment (Fig.
3B). The results of the clone formation assay indicated that
the trend of clone formation ability of A549 cells in each group
was consistent with the trend of A549 cell proliferation (Fig. 3C).
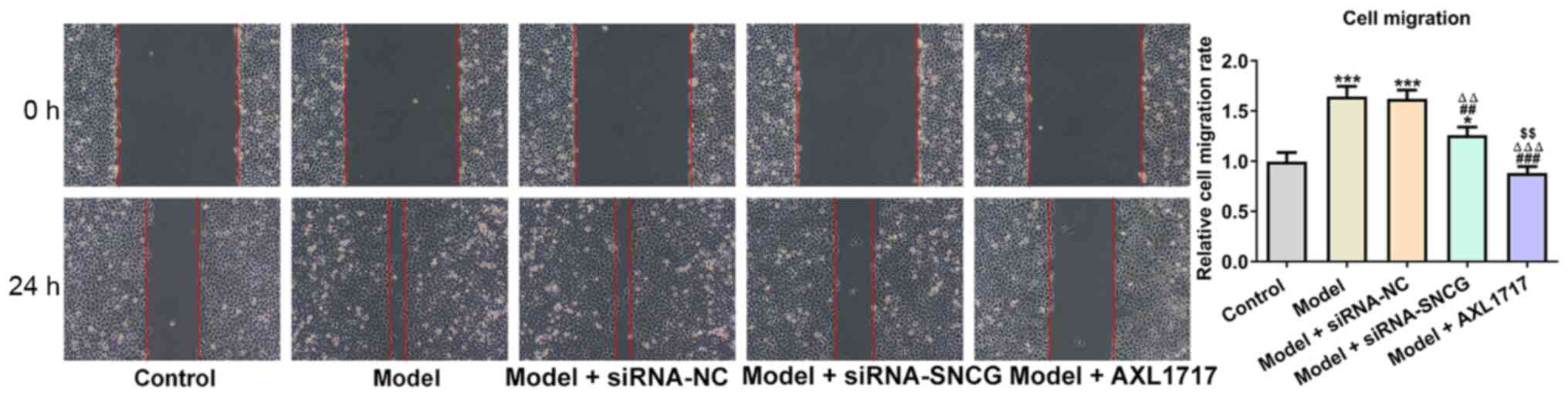
Knockdown of SNCG suppresses the
migration of lung cancer cells induced by high glucose
As presented in Fig.
4, high glucose (25 mM) treatment enhanced A549 cell migration,
an increase which was not affected when high glucose (25
mM)-induced A549 cells were transfected with siRNA NC. However,
knockdown of SNCG suppressed the migration of high glucose (25
mM)-induced A549 cells, and the same effects were observed
following AXL1717 treatment. AXL1717 treatment appeared to exhibit
a significantly greater effect on cell migration compared with
siRNA transfection (Fig. 4).
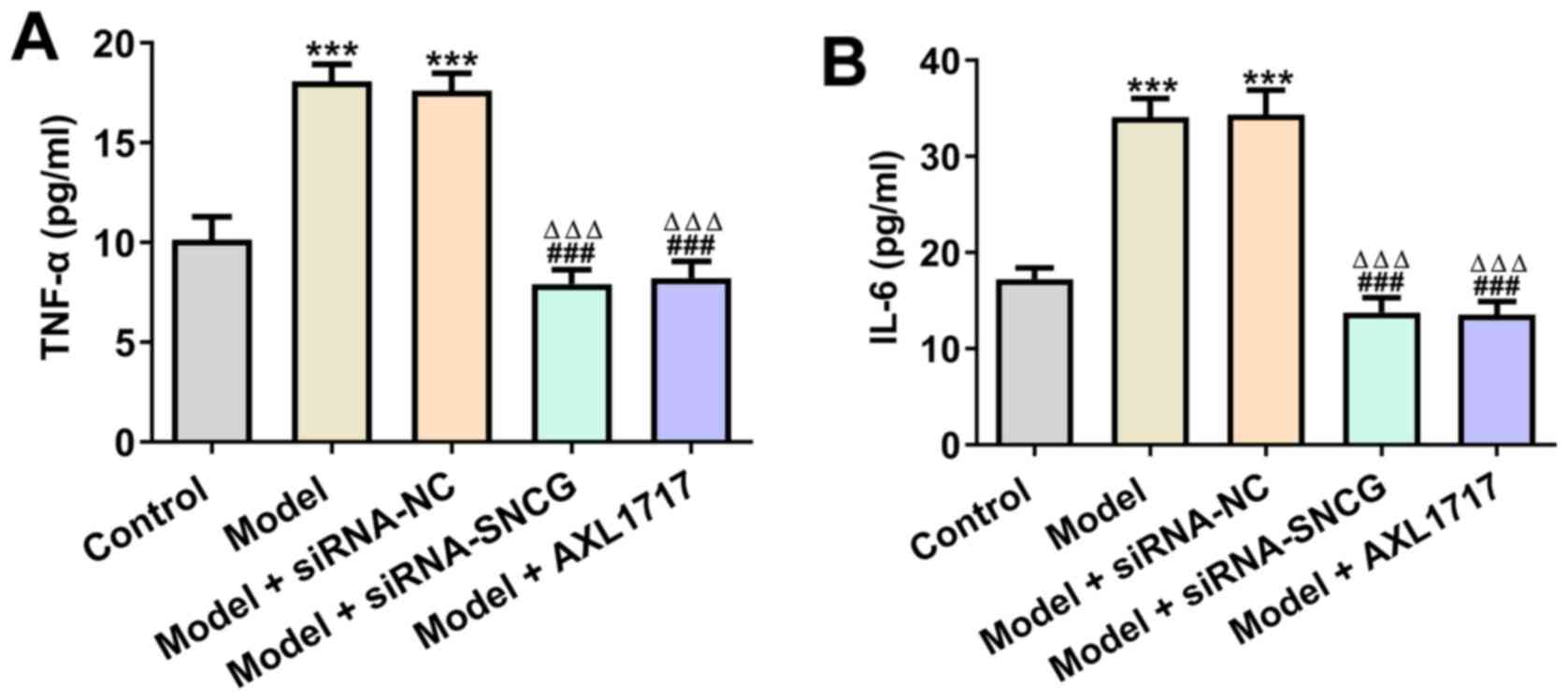
Knockdown of SNCG alleviates
inflammation in lung cancer cells induced by high glucose
The levels of TNF-α were significantly elevated in
high glucose (25 mM)-induced A549 cells, and siRNA NC transfection
did not affect the levels of TNF-α in these cells. Knockdown of
SNCG and AXL1717 treatment decreased TNF-α levels in high glucose
(25 mM)-induced A549 cells (Fig.
5A). High glucose (25 mM) upregulated the levels of IL-6 in
A549 cells and A549 cells transfected with siRNA NC. Moreover,
knockdown of SNCG reversed the promotive effects of high glucose
(25 mM) on the levels of IL-6, which was similar to AXL1717
treatment (Fig. 5B).
Knockdown of SNCG suppresses the
proliferation-associated signaling pathway
High glucose (25 mM) promoted the expression levels
of SNCG, IGF-1 and IGF-1R, which were not significantly altered
after siRNA NC transfection (Fig.
6A). The expression levels of SNCG, IGF-1 and IGF-1R in A549
cells induced by high glucose (25 mM) were decreased after siRNA
SNCG transfection or AXL1717 treatment. In addition, the inhibitory
effects of siRNA-SNCG transfection on SNCG expression were more
significant compared with AXL1717 treatment, while the inhibitory
effects of AXL1717 treatment on IGF-1R expression were more
significant compared with siRNA-SNCG transfection. As presented in
Fig. 6B, high glucose (25 mM)
upregulated the expression levels of ERK1/2 and p-ERK1/2, while JNK
expression was downregulated in A549 cells. It was identified that
knockdown of SNCG suppressed the expression levels of ERK1/2 and
p-ERK1/2, but promoted JNK expression in A549 cells induced by high
glucose (25 mM). The effects of AXL1717 treatment on the expression
levels of ERK1/2, p-ERK1/2 and JNK was consistent with that of SNCG
knockdown. AXL1717 treatment appeared to exhibit a significantly
greater effect on expression of SNCG, IGF-1R and JNK compared with
siRNA transfection.
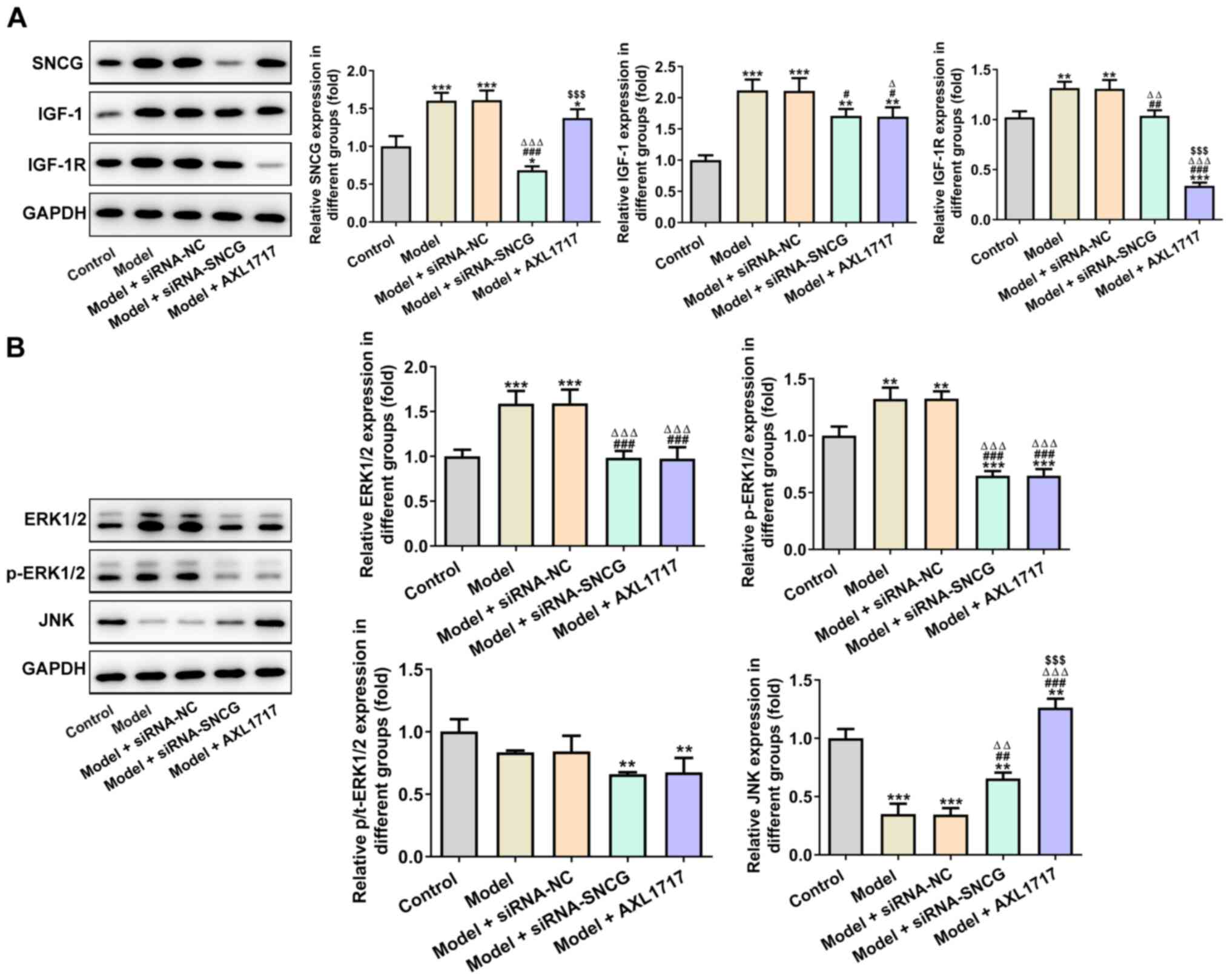 | Figure 6.Knockdown of SNCG suppresses the
proliferation-related signaling pathway. (A) Expression levels of
SNCG, IGF-1 and IGF-1R in A549 cells after indicated treatment were
analyzed using western blot analysis. (B) Expression levels of
ERK1/2, p-ERK1/2 and JNK in A549 cells after indicated treatment
were analyzed via western blot analysis. *P<0.05, **P<0.01
and ***P<0.001 vs. Control group; #P<0.05,
##P<0.01 and ###P<0.001 vs. Model
group; ∆P<0.05, ∆∆P<0.01 and
∆∆∆P<0.001 vs. Model + siRNA-NC group;
$$$P<0.001 vs. Model + siRNA-SNCG group. NC, negative
control; siRNA, small interfering RNA; SNCG, synuclein γ; p-,
phosphorylated; IGF-1, insulin-like growth factor 1; IGF-1R, IGF-1
receptor; t-, total. |
Discussion
With the yearly increase of Chinas aging population
and the prevalence of diabetes, the number of patients with lung
cancer with diabetes is also rising (23,24).
Diabetes can shorten the survival time of patients with lung
cancer, especially in patients with non-small cell lung cancer
undergoing surgical treatment (25,26).
Therefore, additional studies on patients with lung cancer with
diabetes should be performed, and further researches on the
mechanisms and possible interventions are required.
Insulin raises the levels of free and bioactive
IGF-1 in the bloodstream by inhibiting the livers production of
IGF-binding proteins (27). Most
human tumor cells have high expression levels of insulin receptors
and IGF-1R (28). High
concentrations of IGF-1 may increase the risk of multiple
epithelial neoplasms (29) and
influence the occurrence of breast cancer, endometrial cancer,
prostate cancer and other tumors by regulating sex hormones
(30). IGF-1 and IGF-1R can also
induce the transcription of the VEGF gene, upregulate VEGF
expression and enhance the formation of new blood vessels, thereby
promoting the occurrence and development of tumors (31). In the present study, the expression
levels of IGF-1 and IGF-1R were increased in lung cancer cells
induced by high glucose. AXL1717 served as an IGF-1R blocker to
suppress SNCG expression in A549 cells induced by high glucose. It
was found that AXL1717 treatment suppressed the proliferation,
clone formation and migration, as well as alleviated inflammation
in A549 cells induced by high glucose.
SNCG and IGF-1R are mutually regulated in various
types of tumors, including breast, hepatoma, colon and lung cancer
(15). Surgucheva et al
(32) reported that SNCG
overexpression may enhance MMP gene expression by upregulating the
MAPK pathway and phosphorylating AP-1, thereby promoting the
migration and invasion of tumor cells. SNCG overexpression can
stimulate proliferation and induce the metastasis of breast cancer
cells (33). Shen et al
(34) demonstrated that inhibition
of SNCG reduced the colony formation and invasion of breast cancer
cells, and downregulation of the SNCG gene could inhibit tumor
growth in vivo. In the present study, SNCG knockdown could
suppress cell proliferation, clone formation and migration, and
alleviated inflammation in A549 cells induced by high glucose.
ERK is a mitogen-transmitting signal transduction
protein that participates in signaling pathways associated with
proliferation, invasion, migration and apoptosis of a large
proportion of tumor cells (35).
The ERK/JNK signaling pathway is an important intracellular pathway
that regulates proliferation and apoptosis of tumor cells, and its
abnormal activation affects proliferation, differentiation and
apoptosis (36). Yang et al
(37) indicated that ERK
overexpression could enhance the proliferation, migration and
invasion of A549 cells. Moreover, ERK activates p90-ribosomal S6
kinase to inactivate the apoptotic protein Bad, thereby inhibiting
cell apoptosis in lung cancer (38). It has also been reported that
microRNA-23a could further inhibit the proliferation and migration
of A549 cells by activating the JNK signaling pathway (39). In the present study, the
proliferation, clone formation and migration of A549 cells induced
by high glucose was decreased when p-ERK1/2 expression was
downregulated and JNK expression was increased.
Downregulation of SNCG suppresses the proliferation,
migration and tumorigenesis of gastric cancer by inhibiting the
phosphorylation of AKT and ERK (40). Low-dose ProEGCG has also been
reported to significantly enhance phosphorylation of JNK and p38
MAPK, and inhibit phosphorylation of Akt and ERK in endometrial
cancer (41). The change in JNK was
opposite to the changes in Akt and ERK in cancer; these findings
are consistent with the present results. In the present study,
knockdown of SNCG suppressed the expression levels of ERK1/2 and
p-ERK1/2, while promoted JNK expression in A549 cells induced by
high glucose.
In conclusion, the present study demonstrated that
SNCG knockdown suppressed the proliferation, clone formation and
migration, as well as alleviated inflammation of A549 cells induced
by high glucose, which provides a background for the treatment of
patients with lung cancer with diabetes. However, there are
limitations to the present study. For instance, overexpression
experiments were not included in this study and such experiments
would strengthen the results of the present study. Moreover, there
was no animal model to further investigate the role of SNCG in lung
cancer under the condition of high glucose in vivo. These
limitations should be resolved in future studies.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors contributions
PG conceived and designed the experiments. JF
performed all the experiments with the help of CX and MY, and
analyzed the experimental data with the help of XZ. JF wrote the
manuscript, which was revised by PG. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare they have no competing
interests.
References
|
1
|
Torre LA, Trabert B, DeSantis CE, Miller
KD, Samimi G, Runowicz CD, Gaudet MM, Jemal A and Siegel RL:
Ovarian cancer statistics, 2018. CA Cancer J Clin. 68:284–296.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Edwards BK, Brown ML, Wingo PA, Howe HL,
Ward E, Ries LA, Schrag D, Jamison PM, Jemal A, Wu XC, et al:
Annual report to the nation on the status of cancer, 1975–2002,
featuring population-based trends in cancer treatment. J Natl
Cancer Inst. 97:1407–1427. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Han C, Zhang M, Luo X, Wang C, Yin L, Pang
C, Feng T, Ren Y, Wang B, Zhang L, et al: Secular trends in the
prevalence of type 2 diabetes in adults in China from 1995 to 2014:
A meta-analysis. J Diabetes. 9:450–461. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sona MF, Myung SK, Park K and
Jargalsaikhan G: Type 1 diabetes mellitus and risk of cancer: A
meta-analysis of observational studies. Jpn J Clin Oncol.
48:426–433. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Luo J, Chen YJ and Chang LJ: Fasting blood
glucose level and prognosis in non-small cell lung cancer (NSCLC)
patients. Lung Cancer. 76:242–247. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yu WS, Lee CY, Park SY, Suh JW, Narm KS,
Kim DJ, Chung KY and Lee JG: Prognostic factors for resected
non-small cell lung cancer in patients with type 2 diabetes
mellitus. J Surg Oncol. 117:985–993. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ji H, Liu YE, Jia T, Wang M, Liu J, Xiao
G, Joseph BK, Rosen C and Shi YE: Identification of a breast
cancer-specific gene, BCSG1, by direct differential cDNA
sequencing. Cancer Res. 57:759–764. 1997.PubMed/NCBI
|
|
9
|
Bruening W, Giasson BI, Klein-Szanto AJP,
Lee VM, Trojanowski JQ and Godwin AK: Synucleins are expressed in
the majority of breast and ovarian carcinomas and in preneoplastic
lesions of the ovary. Cancer. 88:2154–2163. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gu YM, Tan JX, Lu XW, Ding Y, Han X and
Sun YJ: BCSG1 methylation status and BCSG1 expression in breast
tissues derived from Chinese women with breast cancer. Oncology.
74:61–68. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tastekin D, Kargin S, Karabulut M, Yaldız
N, Tambas M, Gurdal N, Tatli AM, Arslan D, Gok AF and Aykan F:
Synuclein-gamma predicts poor clinical outcome in esophageal cancer
patients. Tumour Biol. 35:11871–11877. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ye Q, Wang TF, Peng YF, Xie J, Feng B, Qiu
MY, Li LH, Lu AG, Liu BY and Zheng MH: Expression of α-, β- and
γ-synuclein in colorectal cancer, and potential clinical
significance in progression of the disease. Oncol Rep. 23:429–436.
2010.PubMed/NCBI
|
|
13
|
Liu C, Shi B, Hao C, Wang Q, Lv Q, Xing N,
Shou J, Qu L, Gao Y, Qin C, et al: Urine gamma-synuclein as a
biomarker for the diagnosis of bladder cancer. Oncotarget.
7:43432–43441. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zheng Y, Jiang J, Zhao N, Liu R, Yang C,
Zhang J, Sun M and Zhang X: Expression of γ-synuclein in non-small
cell lung cancer. Zhongguo Laonianxue Zazhi. 31:4771–4773.
2011.
|
|
15
|
Li M, Yin Y, Hua H, Sun X, Luo T, Wang J
and Jiang Y: The reciprocal regulation of γ-synuclein and IGF-I
receptor expression creates a circuit that modulates IGF-I
signaling. J Biol Chem. 285:30480–30488. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Higashi Y, Sukhanov S, Anwar A, Shai SY
and Delafontaine P: Aging, atherosclerosis, and IGF-1. J Gerontol A
Biol Sci Med Sci. 67:626–639. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xu C: Insulin-like growth factor type 1
receptor and malignancy:An update. Journal of Modern Oncology.
23:706–709. 2015.
|
|
18
|
Annibalini G, Lucertini F, Agostini D,
Vallorani L, Gioacchini A, Barbieri E, Guescini M, Casadei L,
Passalia A, Del Sal M, et al: Concurrent aerobic and resistance
training has anti-inflammatory effects and increases both plasma
and leukocyte levels of IGF-1 in late middle-aged type 2 diabetic
patients. Oxid Med Cell Longev. 2017:39378422017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pollak M: The insulin and insulin-like
growth factor receptor family in neoplasia: An update. Nat Rev
Cancer. 12:159–169. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ding CZ, Guo XF, Wang GL, Wang HT, Xu GH,
Liu YY, Wu ZJ, Chen YH, Wang J and Wang WG: High glucose
contributes to the proliferation and migration of non-small cell
lung cancer cells via GAS5-TRIB3 axis. Biosci Rep.
38:BSR201710142018. View Article : Google Scholar
|
|
21
|
Zhang J, Luo W, Chi X, Zhang L, Ren Q,
Wang H and Zhang W: IGF2BP1 silencing inhibits proliferation and
induces apoptosis of high glucose-induced non-small cell lung
cancer cells by regulating Netrin-1. Arch Biochem Biophys.
693:1085812020. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hemminki K, Li X, Sundquist J and
Sundquist K: Risk of cancer following hospitalization for type 2
diabetes. Oncologist. 15:548–555. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cheng X and Han Y: Impact of diabetes
mellitus on lung cancer and its biological mechanism: A Literature
Review. Chin Gen Pract. 21:1779–1784. 2018.
|
|
25
|
Luo J, Hendryx M, Qi L, Ho GY and Margolis
KL: Pre-existing diabetes and lung cancer prognosis. Br J Cancer.
115:76–79. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhu L, Cao H, Zhang T, Shen H, Dong W,
Wang L and Du J: The effect of diabetes mellitus on lung cancer
prognosis: A PRISMA-compliant meta-analysis of cohort studies.
Medicine (Baltimore). 95:e35282016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Renehan AG, Frystyk J and Flyvbjerg A:
Obesity and cancer risk: The role of the insulin-IGF axis. Trends
Endocrinol Metab. 17:328–336. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Papa V, Pezzino V, Costantino A, Belfiore
A, Giuffrida D, Frittitta L, Vannelli GB, Brand R, Goldfine ID and
Vigneri R: Elevated insulin receptor content in human breast
cancer. J Clin Invest. 86:1503–1510. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pollak MN, Schernhammer ES and Hankinson
SE: Insulin-like growth factors and neoplasia. Nat Rev Cancer.
4:505–518. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Giovannucci E, Harlan DM, Archer MC,
Bergenstal RM, Gapstur SM, Habel LA, Pollak M, Regensteiner JG and
Yee D: Diabetes and cancer: A consensus report. Diabetes Care.
33:1674–1685. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li F, Liu Y, Ren L, Sun Q and Luo YX:
IGF-1 regulates Ang II and VEGF signaling pathways in retinal
neovascularization. Eur Rev Med Pharmacol Sci. 22:6175–6180.
2018.PubMed/NCBI
|
|
32
|
Surgucheva IG, Sivak JM, Fini ME, Palazzo
RE and Surguchov AP: Effect of gamma-synuclein overexpression on
matrix metalloproteinases in retinoblastoma Y79 cells. Arch Biochem
Biophys. 410:167–176. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jiang Y, Liu YE, Goldberg ID and Shi YE: γ
synuclein, a novel heat-shock protein-associated chaperone,
stimulates ligand-dependent estrogen receptor α signaling and
mammary tumorigenesis. Cancer Res. 64:4539–4546. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shen P-H, Fan Q-X, Li Y-W, Zhang W, He XK,
Wang Z and Zhang YH: SNCG shRNA suppressed breast cancer cell
xenograft formation and growth in nude mice. Chin Med J (Engl).
124:1524–1528. 2011.PubMed/NCBI
|
|
35
|
Alexa A, Gógl G, Glatz G, Garai Á, Zeke A,
Varga J, Dudás E, Jeszenői N, Bodor A, Hetényi C, et al: Structural
assembly of the signaling competent ERK2-RSK1 heterodimeric protein
kinase complex. Proc Natl Acad Sci USA. 112:2711–2716. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu K, Lu R, Zhao Q, Du J, Li Y, Zheng M
and Zhang S: Association and clinicopathologic significance of
p38MAPK-ERK-JNK-CDC25C with polyploid giant cancer cell formation.
Med Oncol. 37:62019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yang T, Wang Y, Zhan S, Jia Q and Wang X:
Effects of ERK gene overexpression on proliferation, migration and
invasion of lung cancer A549 Cells. J Ningxia Med Univ. 42:332–338.
2020.
|
|
38
|
Eisinger-Mathason TSK, Andrade J and
Lannigan DA: RSK in tumorigenesis: Connections to steroid
signaling. Steroids. 75:191–202. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jian W, Xue-zhong H, Jing L and Jing G:
The molecular mechanism of miR23a promotes apoptosis of lung cancer
cell line A549 Caspase-3 expression. Anat Res. 40:494–497.
2018.
|
|
40
|
Fan C, Liu J, Tian J, Zhang Y, Yan M and
Zhu C: siRNA Targeting of the SNCG gene inhibits the growth of
gastric carcinoma SGC7901 cells in vitro and in vivo by
downregulating the phosphorylation of AKT/ERK. Cytogenet Genome
Res. 154:209–216. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Man GCW, Wang J, Song Y, Wong JH, Zhao Y,
Lau TS, Leung KT, Chan TH, Wang H, Kwong J, et al: Therapeutic
potential of a novel prodrug of green tea extract in induction of
apoptosis via ERK/JNK and Akt signaling pathway in human
endometrial cancer. BMC Cancer. 20:9642020. View Article : Google Scholar : PubMed/NCBI
|