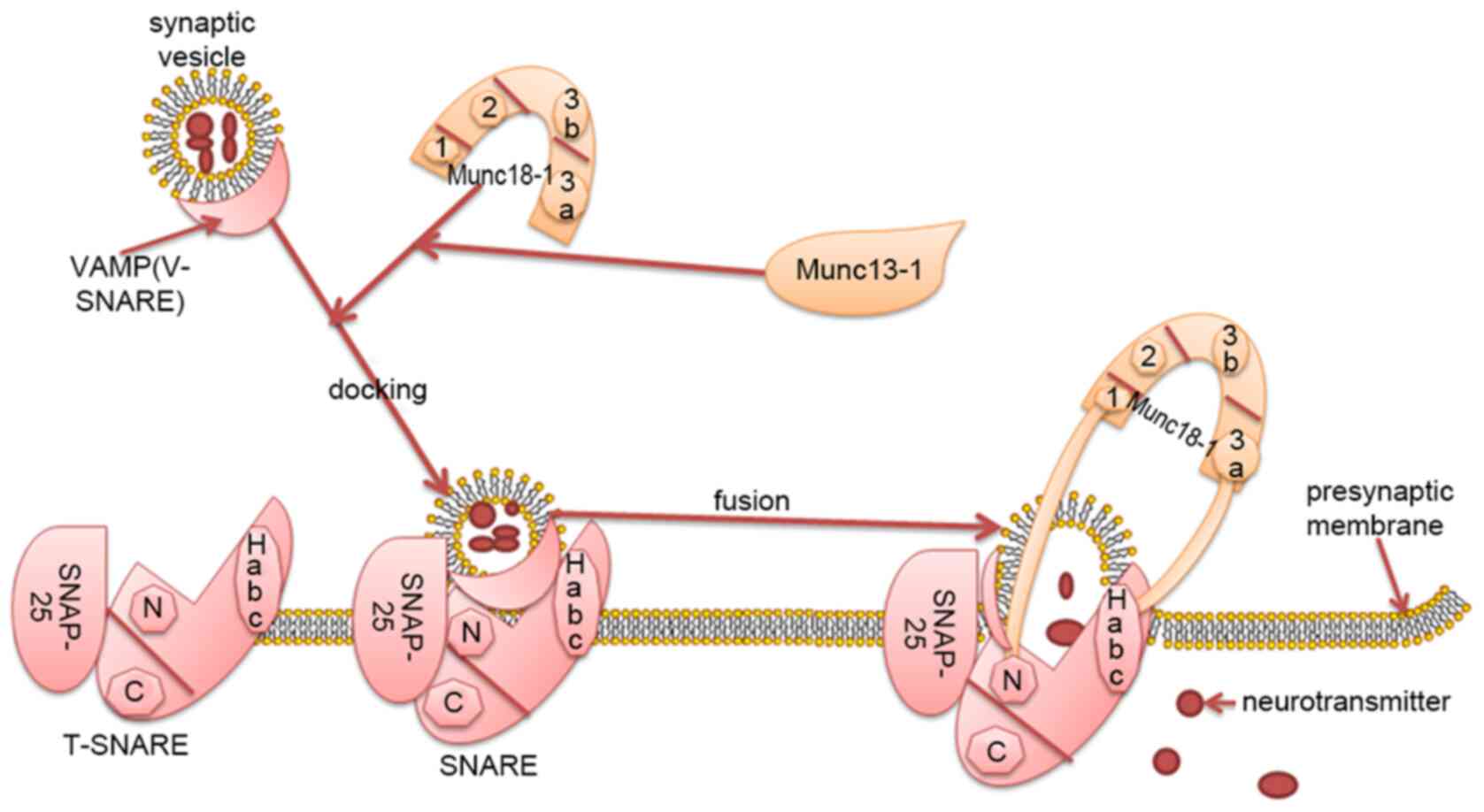
The release of neurotransmitter caused by the fusion
of synaptic vesicles and the presynaptic membrane is an important
process in the transmission of neuronal information (1). This process is regulated by a series
of key proteins, including soluble N-ethylmaleimide-sensitive
factor attachment protein receptor (SNARE), protein transport
protein Sec1 (Sec1)/Mammalian uncoordinated (Munc) 18,
N-ethylmaleimide sensitive factor (NSF), soluble NSF adaptor
protein (SNAP), synaptotagmin-1 and Munc13, among others (2). SNARE protein is the core complex that
establishes membrane fusion, which is mainly composed of SNAP25,
vesicular-associated membrane protein and syntaxin-1. SNAP25 and
syntaxin-1 form the target membrane of the vesicle protein
(T-SNARE), which binds to the synaptic vesicle protein (3). SNARE complex assembly is an important
step in the fusion process of synaptic vesicles and the presynaptic
membrane (4). SNARE
protein-mediated synaptic vesicle fusion and secretion are
regulated by a variety of proteins, of which Sec1/Munc18 (SM)
serves an important role. Previous studies have revealed that the
absence of the SM protein can inhibit membrane fusion in different
systems, such as the endocrine and the vascular system (5–8).
The SM protein family is an indispensable regulatory
protein for membrane fusion. It is a highly conserved polypeptide
chain with a molecular weight of 60–70 kDa and a length of ~600
amino acids (9). The human genome
contains 7 homologous SM proteins, of which syntaxin-binding
protein (Munc18)-1, Munc18-2 and Munc18-3 are involved in
secretion, with Munc18-1 being highly expressed in neurons and
neuroendocrine cells (10).
Munc18-1 is a synaptic fusion protein binding protein and is
encoded by the syntaxin-binding protein 1 gene (STXBP1), which is
located on chromosome 9q34.11 and contains 20 exons (11). The protein encoded by the STXBP1
gene is highly conserved in evolution. The occurrence of missense,
nonsense, frameshift and splicing mutations can lead to
insufficient protein haploids, impaired protein stability and
cortical excitation imbalances that affect learning and memory
(12–14).
Syntaxin-1 is the core protein of SNARE and Munc18-1
primarily regulates vesicle fusion by interacting with syntaxin-1
(Habc domain; N-terminal short peptide) (15). The tertiary structure of Munc18-1 is
complex, exhibiting an arched structure, consisting of four closely
connected domains named 1, 2, 3a and 3b (domains 1, 2, 3a and 3b
form the arch), domains 1 and 3a form an arched gap, domains 3a
contacts with the Habc of syntaxin-1. Domain 1 is located on the
other side of the arched gap and binds to the N-terminal peptide of
syntaxin-1 (Fig. 1) (16).
Munc18-1 promotes or inhibits the assembly of the
SNARE complex primarily by combining with syntaxin-1 (Habc domain;
N-terminal short peptide) in the ‘open’ and ‘closed’ conformations
(17). Munc18-1 regulates vesicle
fusion by interacting with the SNARE complex, affects the number of
vesicles released and the supply of vesicles, and participates in
the transmission of various neurotransmitters, including dopamine,
glutamate and γ-aminobutyric acid (GABA) (Fig. 1) (18,19).
Previous studies have demonstrated that the
N-terminal peptide and Habc domain of syntaxin-1 serve an important
role in synaptic vesicle fusion. The N-terminal peptide is
essential for vesicle fusion itself, whilst the Habc domain
regulates this fusion by forming a closed syntaxin-1 conformation
(20,21). The current hypothesis is that
Munc18-1 and syntaxin-1 have several binding modes: i) Munc18-1
binds to syntaxin-1 (via the Habc domain) in a ‘closed’
conformation, stabilizing the conformation of syntaxin-1 and
inhibiting the assembly of the SNARE complex (16); ii) Munc18-1 combined with ‘closed’
conformation syntaxin-1 serves as a template for SNARE assembly,
with Munc13-1 helping to open the syntaxin-1 ‘closed’ conformation.
Munc13-1 Bridges synaptic vesicles and presynaptic membrane fusion,
and Munc18-1 and Munc13-1 coordinate the assembly of the T-SNARE
complex together in an NSF-SNAP resistant manner (22); and iii) Munc18-1 combined with the
‘open’ conformation ofsyntaxin-1 (N-terminal short peptide)
initiates and stimulates SNARE-mediated membrane fusion (16). The regulatory mechanism of Munc18-1
is complex, involving promotion and inhibition of SNARE assembly;
however, the promotion process is the more dominant of the two.
Munc18-1 can function to regulate brain-derived
neurotrophic factors, participate in the development of synapses
and affect cognitive functions (23,24).
Synaptic plasticity is the basic process responsible for learning
and memory (25). Previous studies
have determined that the expression of Munc18-1 was decreased in
the hippocampus of STXBP1 heterozygous knockout mice. This impaired
synaptic plasticity, synaptic vesicle release rate and vesicle pool
production, leading to impaired learning and memory (26–28).
Activation of the diacylglycerol/protein kinase C (PKC) pathway can
enhance neuronal synaptic transmission (29,30).
In addition to the phosphorylation of Munc18-1, which transfers
synaptic vesicles to active regions, the PKC pathway further
accelerates the assembly of the SNARE complex (31,32).
Synaptotagmin-1 is a synaptic-associated protein, the
phosphorylation of which is an important step in the PKC pathway of
synaptic transmission (33).
Together with Munc18-1 phosphorylation, they represent an important
aspect of synaptic plasticity, enhancing synaptic transmission and
promoting neurotransmitter release (34,35).
The functional destruction of Munc18-1 affects the
development of the cerebral cortex, which may cause
neurodevelopmental disorders by affecting brain-derived
neurotrophic factor and the downstream tropomyosin receptor kinase
B pathway (36). While the
functions of the exocytotic proteins SNARE and Munc18-1 have
garnered increasing attention, studies have demonstrated that
Munc18-1 promotes insulin secretion by interacting with SNARE, and
the decline of its expression leads to insulin secretion disorders
(37,38). Thyroid hormone is very important for
normal brain development. Previous studies have determined that in
mice with mild spatial cognitive impairment, the spatial learning
and memory of old and middle-aged mice decreases. Additionally, the
expression of serum thyroxine is decreased in these animals, which
increases Munc18-1 levels, demonstrating a negative correlation and
indicating that Munc18-1 negatively regulates the role of thyroxine
in brain development (39–41).
Munc18-1 is a key regulatory protein of transmission
and its physiological function has been increasingly studied
(6). The abnormal expression of
Munc18-1 is involved in the pathogenesis of various neurological
diseases and is closely associated with epileptic encephalopathy,
autism, schizophrenia (SCZ), Parkinsons disease, Alzheimers disease
(AD), multiple sclerosis (MS), Duchennes muscular dystrophy and
neuronal ceroid lipofuscinosis (Table
I) (42–50).
Epileptic encephalopathy is a severe brain
dysfunction with poor prognosis that is commonly observed in
infants and children, and is often characterized by frequent
seizures, intellectual disability and dyskinesia (51). The etiology of epileptic
encephalopathy is not completely understood, and it has been
reported that ~70% of epileptic encephalopathy cases are associated
with genetic factors (52,53). An increasing number of studies have
reported that STXBP1exhibitsmultiple mutation types in severe
epileptic encephalopathy, including missense and splicing
mutations, which lead to single protein insufficient ploidy,
impaired protein stability, unbalanced cortical excitement,
impaired synaptic plasticity and GABA-ergic transmission barriers
that adversely affect learning and memory (28,43).
The treatment of epileptic encephalopathy is challenging. However,
studies have reported that drugs such as Keppra and
adreno-cortico-tropic-hormone can prevent epilepsy to a certain
extent, exerting no significant effect on other clinical symptoms,
such as psychomotor disability and cognitive dysfunction (54–57).
Ohtahara syndrome, also known as epileptic
encephalopathy in early infants, is a severe seizure related to
stunting and mental disability. Its other neurological
characteristics include autism spectrum disorder and dyskinesia
(14). STXBP1 is the pathogenic
gene of Ohtahara syndrome. Saitsu et al (58) identified that STXBP1 was mutated in
children with this syndrome in 2008. With research on genetic
diagnosis, an increasing number of studies have identified gene
mutations in Ohtahara syndrome (59,60). A
previous study demonstrated that the clinical manifestations in
children <3 months were accompanied with tonic spasm, and the
electroencephalograms of most children exhibited burst suppression
in Ohtahara syndrome (61). The
main clinical manifestations of West syndrome include nodding
spasm, high irregularities in electroencephalograms and severe
intellectual disability. Since Otsuka et al (62) first reported the STXBP1 mutation in
West syndrome in 2010, following studies then identified STXBP1
nonsense and splice site mutations through clinical studies
(63). In addition, STXBP1
mutations were identified in Dravet syndrome and Lennox-Gastaut
syndrome, although evidence of these mutations in Ohtahara syndrome
is not sufficient (64,65). Rett syndrome (RTT) is a
neurodevelopmental disorder typically caused by mutations of
methyl-CpG binding protein 2 (MECP2). However, Yuge et al
(66) reported in 2018 that a
STXBP1 mutation was detected in a case diagnosed with RTT.
Moreover, a 9-year-old female with RTT and severe cognitive
impairment exhibited no MECP2 mutation, but a STXBP1mutation was
identified. MECP2 gene mutations result in the decrease of
glutamate and gamma-aminobutyric acid receptor densities (17). It is hypothesized that deficits in
synaptic plasticity may, to some extent, indicate an association
between MECP2 and the pathogenicity of STXBP1 mutations (17).
Autism is a common neurodevelopmental disorder with
a high disability rate and strong genetic basis (67). At present, while almost all patients
with STXBP1 mutations exhibit certain clinical phenotypes,
including seizures and mental disabilities, ~20% have autism and
can demonstrate increased aggressive behavior (68,69). A
study by Dachtler et al (45) used a synaptic adhesion protein
α-neurexin II-deficient mouse model of autism, and through protein
quantitative analysis, detemined that Munc18-1 encoded by STXBP1
was significantly reduced in the hippocampus of mice. Synaptic
adhesion proteins serve a key role in the formation and maintenance
of synapses, and are involved in mediating synaptic plasticity
(70). Decreased expression of
Munc18-1 regulates the expression of α-neurexin II, impairs
synaptic plasticity and participates in the occurrence of autism
(71). In a mouse model of
heterozygous STXBP1 knockout, it was revealed that the number of
vesicles in the synaptic active region was reduced, and
glutamatergic and GABA-ergic transmission were impaired. These mice
demonstrated increased anxiety and impaired emotional learning
(72). In addition, Miyamoto et
al (73) reported that the
normalization of excitatory synaptic transmission in STXBP1
knockout mice, which exhibit various degrees of cognitive
impairment and aggressiveness, improved Munc18-1 haploid
deficiency-related aggression. It is speculated that Munc18-1 may
be involved in the clinical phenotype of autism, potentially
providing novel treatment methods.
SCZ is a severe mental disorder with unclear causes.
The main hypotheses suggest that it is mediated by overactive
dopamine activity, 5-hydroxytryptamine and norepinephrine neural
pathway dysfunction, excitatory amino acid dysfunction and the
neuropeptide hypothesis. Neuropeptide protein is involved in the
process of neurodevelopment, and low expression is involved in SCZ
through the release of glutamate and γ-aminobutyric acid) (74). A proteomic analysis of the brain
tissue of patients with SCZ by Behan et al (46) determined that Munc18-1 was highly
expressed. Compared with patients who did not use anti-SCZ drugs,
Munc18-1expression in patients who used these drugs decreased.
Additionally, in a mouse model of SCZ utilized by Urigüen et
al (75), a high expression of
Munc18-1 was identified in the gray matter of the brain. Similarly,
a high expression of Munc18-1 was detected in STXBP1 transgenic
mice, which exhibited symptoms of SCZ, decreased expressions of
dopamine receptors, decreased expressions of membrane transporters
and decreased brain gray matter volume (75). Munc18-1 participates in the
pathogenesis of SCZ by regulating excitatory neurotransmitters;
however, the cause and mechanism of associated brain volume
reduction is unclear.
Parkinsons disease is a degenerative disease of the
nervous system. Previous studies have demonstrated that the
abnormal expression of α-synuclein and self-replication aggregation
serve a key role in the degenerative changes of Parkinsons disease
neurons (76,77). Munc18-1 serves other biological
functions in addition to regulating neurotransmitter transmission.
For example, Lanoue et al (78) revealed that Munc18-1 participates in
various neurodegenerative diseases, including Parkinsons disease,
by controlling the self-replication and aggregation of α-synuclein.
Neurodevelopment in epileptic encephalopathy may also be affected
by synuclein (78). The
physiological functions of α-synuclein are complex; α-synuclein
self-replication and aggregation inhibit exocytosis, affect vesicle
secretion and regulate synaptic plasticity (76,79). A
previous study revealed that Munc18-1 acts as a chaperone to
regulate the replication and aggregation of α-synuclein, the
decreased expression of Munc18-1 increases the aggregation tendency
of α-synuclein, while the increased expression of Munc18-1 reverses
this affect (80). At present, the
specific pathways underlying the Munc18-1-associated regulation of
α-synuclein are unclear, and further research is required to
further understand and effectively treat Parkinsons disease.
AD is a progressive neurodegenerative disorder with
an insidious onset. Its etiology includes the deposition of
abnormal amyloid β extracellular plaques and intracellular
neurofibrillary tangles containing hyperphosphorylated Tau protein,
which lead to impaired synaptic plasticity (81). Previous studies have demonstrated
that at the GABA-ergic presynaptic inhibition terminal, the
expression of the Munc18-1 long splice variant is closely
associated with cognitive function. Furthermore, the deletion of
the Munc18-1 long splice variant increases the risk of AD (47,82).
In a study assessing the effect of pomegranate on an AD mouse
model, it was determined that pomegranate can enhance synaptic
plasticity by increasing the expression of Munc18-1, SNAP25 and
synaptophysin, improving spatial learning impairment in mice
(83). Additionally, a study has
determined that when comparing the brain tissue of healthy
individuals and deceased patients with AD of the same age, brain
tissue following AD-associated death is rich in highly
phosphorylated Tau protein and Munc18-1 is abnormally expressed
(84). The current treatment
methods used for patients with AD remain poor, and further
understanding of Munc18-1 may provide a novel target.
The most common type of white matter injury is
demyelination, which includes myelin loss and axonal abnormalities
(85). MS is an autoimmune disease
characterized by the inflammatory demyelination of white matter in
the central nervous system (86). A
previous study reported that in a murine model of multiple
sclerosis, the expression of Munc18-1 and axon structural protein
α-internexin increases, and the expression of glutamate
decarboxylase decreases, mediating immunity to myelin damage
(48). An increased expression of
α-internexin, axon disintegration, the Munc18-1 regulation of
glutamatergic transmission and glutamate excitotoxicity further
increase axon damage (87).
However, there are few studies that assessMunc18-1 in MS and the
specific underlying pathogenic mechanism remains unknown.
Duchenne muscular dystrophy is a disease that
affects the neuromuscular system. Progressive skeletal muscle
atrophy is the main complication, and developmental cognitive
deficits and behavioral abnormalities are its main clinical
features (88). Murphy et al
(49) performed proteomics on the
brain tissue of mice with Duchenne muscular dystrophy, the results
of which revealed that the expression of Munc18-1 was significantly
decreased. A further study revealed that the expression of Munc18-1
was significantly decreased in the brains of children with neuronal
ceroid lipofuscinosis (50). These
studies indicate that Munc18-1 may be involved in the pathogenesis
of Duchenne muscular dystrophy and neurodegenerative lysosomal
diseases. Combined with the characteristics of these diseases,
impaired synaptic plasticity and neurodevelopmental pathways serve
an important role in these diseases. Munc18-1 may participate in
the pathogenesis of disease by regulating synaptic plasticity and
neurodevelopment; however there are few related studies to confirm
this, and the specific underlying mechanisms require further
exploration.
Munc18-1 has powerful biological functions and can
regulate the balance of synaptic excitatory neurotransmitter and
inhibitory neurotransmitter transmission, and mediate disease
processes at increased or decreased expressions. Decreased Munc18-1
expression also damages synaptic plasticity and affects learning
and memory. Neuronal transmission requires the participation of
multiple neurotransmitters, and Munc18-1 regulates neurotransmitter
transmission by regulating the SNARE complex. This process is
complicated and requires the participation of multiple regulatory
proteins. The relationship between the expression of Munc18-1 and
specific neurotransmitter regulation mechanisms remains unclear and
requires further study. An enhanced understanding of the function
of Munc18-1 may elucidate novel therapeutic strategies for the
treatment of neurological diseases.
Not applicable.
The present study was supported by the National Key
R&D Program of China (grant nos. 2017YFA 0104201 and 2017YFA
0104200), the National Science Foundation of China (grant nos.
82071353, 82001593, 81330016, 81630038 and 81771634), and the
Science and Technology Bureau of Chengdu City (grant no.
2015-HM01-00424-SF).
Not applicable.
FT and DX conceived the present study. FT, DX, LC
and XL provided the software. FT, DX, HG and XL provided the study
resources. HG and XL generated the figure. FT and DX drafted the
manuscript. FT, DX, HG, HG and XL reviewed and edited the
manuscript. XL supervised the study. DL and XL confirm the
authenticity of all the data. All authors read and approved the
final manuscript.
Not applicable.
Not applicable.
All authors declare that they have no competing
interests.
|
1
|
Brose N, Brunger A, Cafiso D, Chapman ER,
Diao J, Hughson FM, Jackson MB, Jahn R, Lindau M, Ma C, et al:
Synaptic vesicle fusion: today and beyond. Nat Struct Mol Biol.
26:663–668. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Snead D and Eliezer D: Intrinsically
disordered proteins in synaptic vesicle trafficking and release. J
Biol Chem. 294:3325–3342. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ruete MC, Zarelli VEP, Masone D, de Paola
M, Bustos DM and Tomes CN: A connection between reversible tyrosine
phosphorylation and SNARE complex disassembly activity of
N-ethylmaleimide-sensitive factor unveiled by the phosphomimetic
mutant N-ethylmaleimide-sensitive factor-Y83E. Mol Hum Reprod.
25:344–358. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bombardier JP and Munson M: Three steps
forward, two steps back: Mechanistic insights into the assembly and
disassembly of the SNARE complex. Curr Opin Chem Biol. 29:66–71.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kavanagh DM, Smyth AM, Martin KJ, Dun A,
Brown ER, Gordon S, Smillie KJ, Chamberlain LH, Wilson RS, Yang L,
et al: A molecular toggle after exocytosis sequesters the
presynaptic syntaxin1a molecules involved in prior vesicle fusion.
Nat Commun. 5:57742014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jorgačevski J and Zorec R: Munc18-1,
exocytotic fusion pore regulation and local membrane anisotropy.
Commun Integr Biol. 5:74–77. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Han GA, Malintan NT, Saw NM, Li L, Han L,
Meunier FA, Collins BM and Sugita S: Munc18-1 domain-1 controls
vesicle docking and secretion by interacting with syntaxin-1 and
chaperoning it to the plasma membrane. Mol Biol Cell. 22:4134–4149.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Graham ME, Prescott GR, Johnson JR, Jones
M, Walmesley A, Haynes LP, Morgan A, Burgoyne RD and Barclay JW:
Structure-function study of mammalian Munc18-1 and C.
elegans UNC-18 implicates domain 3b in the regulation of
exocytosis. PLoS One. 6:e179992011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rizo J and Südhof TC: The membrane fusion
enigma: SNAREs, Sec1/Munc18 proteins, and their accomplices--guilty
as charged? Annu Rev Cell Dev Biol. 28:279–308. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pons-Vizcarra M, Kurps J, Tawfik B,
Sørensen JB, van Weering JRT and Verhage M: MUNC18-1 regulates the
submembrane F-actin network, independently of syntaxin1 targeting,
via hydrophobicity in β-sheet 10. J Cell Sci.
132:jcs2346742019.https://doi.org/10.1242/jcs.234674 View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hamdan FF, Piton A, Gauthier J, Lortie A,
Dubeau F, Dobrzeniecka S, Spiegelman D, Noreau A, Pellerin S, Côté
M, et al: De novo STXBP1 mutations in mental retardation and
nonsyndromic epilepsy. Ann Neurol. 65:748–753. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Deciphering Developmental Disorders Study:
Prevalence and architecture of de novo mutations in developmental
disorders. Nature. 542:433–438. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Weckhuysen S, Holmgren P, Hendrickx R,
Jansen AC, Hasaerts D, Dielman C, de Bellescize J, Boutry-Kryza N,
Lesca G, Von Spiczak S, et al: Reduction of seizure frequency after
epilepsy surgery in a patient with STXBP1 encephalopathy and
clinical description of six novel mutation carriers. Epilepsia.
54:e74–e80. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kovacevic J, Maroteaux G, Schut D, Loos M,
Dubey M, Pitsch J, Remmelink E, Koopmans B, Crowley J, Cornelisse
LN, et al: Protein instability, haploinsufficiency, and cortical
hyper-excitability underlie STXBP1 encephalopathy. Brain.
141:1350–1374. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Eisemann TJ, Allen F, Lau K, Shimamura GR,
Jeffrey PD and Hughson FM: The Sec1/Munc18 protein Vps45 holds the
Qa-SNARE Tlg2 in an open conformation. eLife. 9:e607242020.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lee S, Shin J, Jung Y, Son H, Shin J,
Jeong C, Kweon DH and Shin YK: Munc18-1 induces conformational
changes of syntaxin-1 in multiple intermediates for SNARE assembly.
Sci Rep. 10:116232020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Romaniello R, Saettini F, Panzeri E,
Arrigoni F, Bassi MT and Borgatti R: A de-novo STXBP1 gene mutation
in a patient showing the Rett syndrome phenotype. Neuroreport.
26:254–257. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gil-Pisa I, Munarriz-Cuezva E,
Ramos-Miguel A, Urigüen L, Meana JJ and García-Sevilla JA:
Regulation of munc18-1 and syntaxin-1A interactive partners in
schizophrenia prefrontal cortex: Down-regulation of munc18-1a
isoform and 75 kDa SNARE complex after antipsychotic treatment. Int
J Neuropsychopharmacol. 15:573–588. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ramos-Miguel A, Beasley CL, Dwork AJ, Mann
JJ, Rosoklija G, Barr AM and Honer WG: Increased SNARE
Protein-Protein Interactions in Orbitofrontal and Anterior
Cingulate Cortices in Schizophrenia. Biological Psychiatry.
78:361–373. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhou P, Pang ZP, Yang X, Zhang Y,
Rosenmund C, Bacaj T and Südhof TC: Syntaxin-1 N-peptide and
Habc-domain perform distinct essential functions in synaptic
vesicle fusion. EMBO J. 32:159–171. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jiang X, Zhang Z, Cheng K, Wu Q, Jiang L,
Pielak GJ, Liu M and Li C: Membrane-mediated disorder-to-order
transition of SNAP25 flexible linker facilitates its interaction
with syntaxin-1 and SNARE-complex assembly. FASEB J. 33:7985–7994.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sitarska E, Xu J, Park S, Liu X, Quade B,
Stepien K, Sugita K, Brautigam CA, Sugita S and Rizo J:
Autoinhibition of Munc18-1 modulates synaptobrevin binding and
helps to enable Munc13-dependent regulation of membrane fusion.
eLife. 6:e242782017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lee YI, Kim YG, Pyeon HJ, Ahn JC, Logan S,
Orock A, Joo KM, Lőrincz A and Deák F: Dysregulation of the
SNARE-binding protein Munc18-1 impairs BDNF secretion and synaptic
neurotransmission: A novel interventional target to protect the
aging brain. Geroscience. 41:109–123. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Peng Y, Lee J, Rowland K, Wen Y, Hua H,
Carlson N, Lavania S, Parrish JZ and Kim MD: Regulation of dendrite
growth and maintenance by exocytosis. J Cell Sci. 128:4279–4292.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
De Pittà M, Brunel N and Volterra A:
Astrocytes: Orchestrating synaptic plasticity? Neuroscience.
323:43–61. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lammertse HCA, van Berkel AA, Iacomino M,
Toonen RF, Striano P, Gambardella A, Verhage M and Zara F:
Homozygous STXBP1 variant causes encephalopathy and
gain-of-function in synaptic transmission. Brain. 143:441–451.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Meijer M, Cijsouw T, Toonen RF and Verhage
M: Synaptic effects of Munc18-1 alternative splicing in excitatory
hippocampal neurons. PLoS One. 10:e01389502015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Orock A, Logan S and Deak F: Munc18-1
haploinsufficiency impairs learning and memory by reduced synaptic
vesicular release in a model of Ohtahara syndrome. Mol Cell
Neurosci. 88:33–42. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang CC, Weyrer C, Paturu M, Fioravante D
and Regehr WG: Calcium-dependent protein kinase C is not required
for post-tetanic potentiation at the hippocampal CA3 to CA1
synapse. J Neurosci. 36:6393–6402. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Galván EJ, Cosgrove KE, Mauna JC, Card JP,
Thiels E, Meriney SD and Barrionuevo G: Critical involvement of
postsynaptic protein kinase activation in long-term potentiation at
hippocampal mossy fiber synapses on CA3 interneurons. J Neurosci.
30:2844–2855. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Barclay JW, Craig TJ, Fisher RJ, Ciufo LF,
Evans GJ, Morgan A and Burgoyne RD: Phosphorylation of Munc18 by
protein kinase C regulates the kinetics of exocytosis. J Biol Chem.
278:10538–10545. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wierda KD, Toonen RF, de Wit H, Brussaard
AB and Verhage M: Interdependence of PKC-dependent and
PKC-independent pathways for presynaptic plasticity. Neuron.
54:275–290. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cijsouw T, Weber JP, Broeke JH, Broek JA,
Schut D, Kroon T, Saarloos I, Verhage M and Toonen RF: Munc18-1
redistributes in nerve terminals in an activity- and PKC-dependent
manner. J Cell Biol. 204:759–775. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
de Jong AP, Meijer M, Saarloos I,
Cornelisse LN, Toonen RF, Sørensen JB and Verhage M:
Phosphorylation of synaptotagmin-1 controls a post-priming step in
PKC-dependent presynaptic plasticity. Proc Natl Acad Sci USA.
113:5095–5100. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Genc O, Kochubey O, Toonen RF, Verhage M
and Schneggenburger R: Munc18-1 is a dynamically regulated PKC
target during short-term enhancement of transmitter release. eLife.
3:e017152014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hamada N, Iwamoto I, Tabata H and Nagata
KI: MUNC18-1 gene abnormalities are involved in neurodevelopmental
disorders through defective cortical architecture during brain
development. Acta Neuropathol Commun. 5:922017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lang H, Ai Z, You Z, Wan Y, Guo W, Xiao J
and Jin X: Characterization of miR-218/322-Stxbp1 pathway in the
process of insulin secretion. J Mol Endocrinol. 54:65–73. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Oh E, Kalwat MA, Kim MJ, Verhage M and
Thurmond DC: Munc18-1 regulates first-phase insulin release by
promoting granule docking to multiple syntaxin isoforms. J Biol
Chem. 287:25821–25833. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cao L, Wang F, Yang QG, Jiang W, Wang C,
Chen YP and Chen GH: Reduced thyroid hormones with increased
hippocampal SNAP-25 and Munc18-1 might involve cognitive impairment
during aging. Behav Brain Res. 229:131–137. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cao L, Jiang W, Wang F, Yang QG, Wang C,
Chen YP and Chen GH: The reduced serum free triiodothyronine and
increased dorsal hippocampal SNAP-25 and Munc18-1 had existed in
middle-aged CD-1 mice with mild spatial cognitive impairment. Brain
Res. 1540:9–20. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zevenbergen C, Groeneweg S, Swagemakers
SMA, de Jong A, Medici-Van den Herik E, Rispens M, Klootwijk W,
Medici M, de Rijke YB, Meima ME, et al: Functional analysis of
genetic variation in the SECIS element of thyroid hormone
activating type 2 deiodinase. J Clin Endocrinol Metab.
104:1369–1377. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Grone BP, Marchese M, Hamling KR, Kumar
MG, Krasniak CS, Sicca F, Santorelli FM, Patel M and Baraban SC:
Epilepsy, behavioral abnormalities, and physiological comorbidities
in syntaxin-binding protein 1 (STXBP1) mutant Zebrafish. PLoS One.
11:e01511482016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ortega-Moreno L, Giráldez BG, Verdú A,
García-Campos O, Sánchez-Martín G, Serratosa JM and Guerrero-López
R: Novel mutation in STXBP1 gene in a patient with non-lesional
Ohtahara syndrome. Neurologia. 31:523–527. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Dachtler J, Ivorra JL, Rowland TE, Lever
C, Rodgers RJ and Clapcote SJ: Heterozygous deletion of α-neurexin
I or α-neurexin II results in behaviors relevant to autism and
schizophrenia. Behav Neurosci. 129:765–776. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Dachtler J, Glasper J, Cohen RN, Ivorra
JL, Swiffen DJ, Jackson AJ, Harte MK, Rodgers RJ and Clapcote SJ:
Deletion of α-neurexin II results in autism-related behaviors in
mice. Transl Psychiatry. 4:e4842014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Behan AT, Byrne C, Dunn MJ, Cagney G and
Cotter DR: Proteomic analysis of membrane microdomain-associated
proteins in the dorsolateral prefrontal cortex in schizophrenia and
bipolar disorder reveals alterations in LAMP, STXBP1 and BASP1
protein expression. Mol Psychiatry. 14:601–613. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lanoue V, Chai YJ, Brouillet JZ,
Weckhuysen S, Palmer EE, Collins BM and Meunier FA: STXBP1
encephalopathy: Connecting neurodevelopmental disorders with
α-synucleinopathies? Neurology. 93:114–123. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Linker RA, Brechlin P, Jesse S, Steinacker
P, Lee DH, Asif AR, Jahn O, Tumani H, Gold R and Otto M: Proteome
profiling in murine models of multiple sclerosis: Identification of
stage specific markers and culprits for tissue damage. PLoS One.
4:e76242009. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Murphy S, Zweyer M, Henry M, Meleady P,
Mundegar RR, Swandulla D and Ohlendieck K: Label-free mass
spectrometric analysis reveals complex changes in the brain
proteome from the mdx-4cv mouse model of Duchenne muscular
dystrophy. Clin Proteomics. 12:272015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Sleat DE, Tannous A, Sohar I, Wiseman JA,
Zheng H, Qian M, Zhao C, Xin W, Barone R, Sims KB, et al: Proteomic
analysis of brain and cerebrospinal fluid from the three major
forms of neuronal ceroid lipofuscinosis reveals potential
biomarkers. J Proteome Res. 16:3787–3804. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Scheffer IE, Berkovic S, Capovilla G,
Connolly MB, French J, Guilhoto L, Hirsch E, Jain S, Mathern GW,
Moshé SL, et al: ILAE classification of the epilepsies: Position
paper of the ILAE Commission for Classification and Terminology.
Epilepsia. 58:512–521. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Hunter MB, Yoong M, Sumpter RE, Verity K,
Shetty J, McLellan A, Jones J, Quigley A, Tallur KK and Chin RFM:
Neurobehavioral problems in children with early-onset epilepsy: A
population-based study. Epilepsy Behav. 93:87–93. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Mercimek-Mahmutoglu S, Patel J, Cordeiro
D, Hewson S, Callen D, Donner EJ, Hahn CD, Kannu P, Kobayashi J,
Minassian BA, et al: Diagnostic yield of genetic testing in
epileptic encephalopathy in childhood. Epilepsia. 56:707–716. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Liu S, Wang L, Cai XT, Zhou H, Yu D and
Wang Z: Therapeutic benefits of ACTH and levetiracetam in STXBP1
encephalopathy with a de novo mutation: A case report and
literature review. Medicine (Baltimore). 97:e06632018. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Li T, Cheng M, Wang J, Hong S, Li M, Liao
S, Xie L and Jiang L: De novo mutations of STXBP1 in Chinese
children with early onset epileptic encephalopathy. Genes Brain
Behav. 17:e124922018. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Stamberger H, Weckhuysen S and De Jonghe
P: STXBP1 as a therapeutic target for epileptic encephalopathy.
Expert Opin Ther Targets. 21:1027–1036. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Dilena R, Striano P, Traverso M, Viri M,
Cristofori G, Tadini L, Barbieri S, Romeo A and Zara F: Dramatic
effect of levetiracetam in early-onset epileptic encephalopathy due
to STXBP1 mutation. Brain Dev. 38:128–131. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Saitsu H, Kato M, Mizuguchi T, Hamada K,
Osaka H, Tohyama J, Uruno K, Kumada S, Nishiyama K, Nishimura A, et
al: De novo mutations in the gene encoding STXBP1 (MUNC18-1) cause
early infantile epileptic encephalopathy. Nat Genet. 40:782–788.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
59
|
Lee S, Kim SH, Kim B, Lee ST, Choi JR, Kim
HD, Lee JS and Kang HC: Genetic diagnosis and clinical
characteristics by etiological classification in early-onset
epileptic encephalopathy with burst suppression pattern. Epilepsy
Res. 163:1063232020. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Mitta N, Menon RN, McTague A,
Radhakrishnan A, Sundaram S, Cherian A, Madhavilatha GK, Mannan AU,
Nampoothiri S and Thomas SV: Genotype-phenotype correlates of
infantile-onset developmental & epileptic encephalopathy
syndromes in South India: A single centre experience. Epilepsy Res.
166:1063982020. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Di Meglio C, Lesca G, Villeneuve N,
Lacoste C, Abidi A, Cacciagli P, Altuzarra C, Roubertie A, Afenjar
A, Renaldo-Robin F, et al: Epileptic patients with de novo STXBP1
mutations: Key clinical features based on 24 cases. Epilepsia.
56:1931–1940. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Otsuka M, Oguni H, Liang JS, Ikeda H, Imai
K, Hirasawa K, Imai K, Tachikawa E, Shimojima K, Osawa M, et al:
STXBP1 mutations cause not only Ohtahara syndrome but also West
syndrome--result of Japanese cohort study. Epilepsia. 51:2449–2452.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Boutry-Kryza N, Labalme A, Ville D, de
Bellescize J, Touraine R, Prieur F, Dimassi S, Poulat AL, Till M,
Rossi M, et al: Molecular characterization of a cohort of 73
patients with infantile spasms syndrome. Eur J Med Genet. 58:51–58.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Steel D, Symonds JD, Zuberi SM and
Brunklaus A: Dravet syndrome and its mimics: Beyond SCN1A.
Epilepsia. 58:1807–1816. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Mastrangelo M: Lennox-Gastaut Syndrome: A
state of the art review. Neuropediatrics. 48:143–151. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Yuge K, Iwama K, Yonee C, Matsufuji M,
Sano N, Saikusa T, Yae Y, Yamashita Y, Mizuguchi T, Matsumoto N, et
al: A novel STXBP1 mutation causes typical Rett syndrome in a
Japanese girl. Brain Dev. 40:493–497. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Li J, Lin X, Wang M, Hu Y, Xue K, Gu S, Lv
L, Huang S and Xie W: Potential role of genomic imprinted genes and
brain developmental related genes in autism. BMC Med Genomics.
13:542020. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Stamberger H, Nikanorova M, Willemsen MH,
Accorsi P, Angriman M, Baier H, Benkel-Herrenbrueck I, Benoit V,
Budetta M, Caliebe A, et al: STXBP1 encephalopathy: A
neurodevelopmental disorder including epilepsy. Neurology.
86:954–962. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Jiang YH, Yuen RK, Jin X, Wang M, Chen N,
Wu X, Ju J, Mei J, Shi Y, He M, et al: Detection of clinically
relevant genetic variants in autism spectrum disorder by
whole-genome sequencing. Am J Hum Genet. 93:249–263. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Dudanova I, Tabuchi K, Rohlmann A, Südhof
TC and Missler M: Deletion of α-neurexins does not cause a major
impairment of axonal pathfinding or synapse formation. J Comp
Neurol. 502:261–274. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Toonen RF, Wierda K, Sons MS, de Wit H,
Cornelisse LN, Brussaard A, Plomp JJ and Verhage M: Munc18-1
expression levels control synapse recovery by regulating readily
releasable pool size. Proc Natl Acad Sci USA. 103:18332–18337.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
7Chamma I, Sainlos M and Thoumine O:
Biophysical mechanisms underlying the membrane trafficking of
synaptic adhesion molecules. Neuropharmacology. 169:1075552020.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Miyamoto H, Shimohata A, Abe M, Abe T,
Mazaki E, Amano K, Suzuki T, Tatsukawa T, Itohara S, Sakimura K, et
al: Potentiation of excitatory synaptic transmission ameliorates
aggression in mice with Stxbp1 haploinsufficiency. Hum Mol Genet.
26:4961–4974. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Valton V, Romaniuk L, Douglas Steele J,
Lawrie S and Seriès P: Comprehensive review: Computational
modelling of schizophrenia. Neurosci Biobehav Rev. 83:631–646.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Urigüen L, Gil-Pisa I, Munarriz-Cuezva E,
Berrocoso E, Pascau J, Soto-Montenegro ML, Gutiérrez-Adán A,
Pintado B, Madrigal JL, Castro E, et al: Behavioral, neurochemical
and morphological changes induced by the overexpression of
munc18-1a in brain of mice: Relevance to schizophrenia. Transl
Psychiatry. 3:e2212013. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Kim ST, Moon W, Chae Y, Kim YJ, Lee H and
Park HJ: The effect of electroaucpuncture for
1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced proteomic
changes in the mouse striatum. J Physiol Sci. 60:27–34. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Burré J, Sharma M and Südhof TC: Cell
biology and pathophysiology of α-synuclein. Cold Spring Harb
Perspect Med. 8:a0240912018. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Lanoue V, Chai YJ, Brouillet JZ,
Weckhuysen S, Palmer EE, Collins BM and Meunier FA: STXBP1
encephalopathy: Connecting neurodevelopmental disorders with
alpha-synucleinopathies? Neurology. 93:114–123. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Huang CC, Chiu TY, Lee TY, Hsieh HJ, Lin
CC and Kao LS: Soluble α-synuclein facilitates priming and fusion
by releasing Ca2+ from the thapsigargin-sensitive
Ca2+ pool in PC12 cells. J Cell Sci.
131:jcs2130172018.https://doi.org/10.1242/jcs.213017 View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Chai YJ, Sierecki E, Tomatis VM, Gormal
RS, Giles N, Morrow IC, Xia D, Götz J, Parton RG, Collins BM, et
al: Munc18-1 is a molecular chaperone for α-synuclein, controlling
its self-replicating aggregation. J Cell Biol. 214:705–718. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Braidy N, Essa MM, Poljak A, Selvaraju S,
Al-Adawi S, Manivasagm T, Thenmozhi AJ, Ooi L, Sachdev P and
Guillemin GJ: Consumption of pomegranates improves synaptic
function in a transgenic mice model of Alzheimer's disease.
Oncotarget. 7:64589–64604. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Ramos-Miguel A, Hercher C, Beasley CL,
Barr AM, Bayer TA, Falkai P, Leurgans SE, Schneider JA, Bennett DA
and Honer WG: Loss of Munc18-1 long splice variant in GABAergic
terminals is associated with cognitive decline and increased risk
of dementia in a community sample. Mol Neurodegener. 10:652015.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Liu X, Hao W, Qin Y, Decker Y, Wang X,
Burkart M, Schötz K, Menger MD, Fassbender K and Liu Y: Long-term
treatment with Ginkgo biloba extract EGb 761 improves
symptoms and pathology in a transgenic mouse model of Alzheimer's
disease. Brain Behav Immun. 46:121–131. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Donovan LE, Higginbotham L, Dammer EB,
Gearing M, Rees HD, Xia Q, Duong DM, Seyfried NT, Lah JJ and Levey
AI: Analysis of a membrane-enriched proteome from postmortem human
brain tissue in Alzheimer's disease. Proteomics Clin Appl.
6:201–211. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Miyamoto N, Maki T, Shindo A, Liang AC,
Maeda M, Egawa N, Itoh K, Lo EK, Lok J, Ihara M, et al: Astrocytes
promote oligodendrogenesis after white matter damage via
brain-derived neurotrophic factor. J Neurosci. 35:14002–14008.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Deiva K: Pediatric onset multiple
sclerosis. Rev Neurol (Paris). 176:30–36. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Werner P, Pitt D and Raine CS: Glutamate
excitotoxicity--a mechanism for axonal damage and oligodendrocyte
death in Multiple Sclerosis? J Neural Transm Suppl. 60:375–385.
2000.
|
|
88
|
Doorenweerd N, Dumas EM, Ghariq E, Schmid
S, Straathof CS, Roest AA, Wokke BH, van Zwet EW, Webb AG,
Hendriksen JG, et al: Decreased cerebral perfusion in Duchenne
muscular dystrophy patients. Neuromuscul Disord. 27:29–37. 2017.
View Article : Google Scholar : PubMed/NCBI
|