Stem cells are a group of progenitor cells
characterized by self-renewal and differentiation ability. Within
their niche or local microenvironment, stem cells communicate via
physical and mechanical cues to regulate cell fate and behavior
(1). Early research has focused on
biochemical signals produced by chemical or solution media when
discussing the molecular signaling mechanisms underlying stem cell
self-renewal, growth and differentiation (2). However, it is hypothesized that cells
perceive their microenvironment not only through soluble signals
but also via physical and mechanical cues, such as extracellular
matrix (ECM) stiffness or confined adhesiveness (3). Similar to intrinsic and extrinsic
biochemical factors, mechanical cues resulting from both
intracellularly generated and externally applied forces have a
broad range of effects on stem cell fate. For example, MSCs have
the ability to differentiate into neuroblast, chondrocyte,
osteoblast, adipocytes and numerous other cell types within
matrices, which mimic the stiffness of their native substrate
(4–6). Also, the extracellular matrix
structure such as nanotopographic structures or fluid shear stress
can affect differentiation of stem cells (7,8).
Studies have shown that yes-associated protein/transcriptional
coactivator with PDZ-binding motif (YAP/TAZ) serves an important
role in biomechanical and mechanical signaling that affects
self-renewal and differentiation of stem cells (3,9). The
present review summarizes the current knowledge of the mechanisms
involved in YAP/TAZ regulation on the physical and mechanical
microenvironment and its potential effects on stem cell
differentiation.
Stem cells are a paradigm model system in the
mechanotransduction field; it has been shown that differentiation
of stem cells into distinct cell fates is dictated by the physical
features of the cellular microenvironment (10). Numerous studies have shown that
YAP/TAZ is a key regulator of mechanical properties of the stem
cell microenvironment (11–14). Various biophysical stimuli from
external forces, such as cyclic stretching, shear stress and
acoustic tweezing, also modulate stem cell fate via YAP/TAZ
(15–18).
Flow shear stress can be defined as a function of
strain rate, which is the frictional force generated by the flow of
fluid on a contact surface (19).
More specifically, fluid shear stress is defined as stress (i.e.,
force per unit area) applied by a fluid to the tangential direction
of the contact surface. Fluid shear stress serves an important role
in the differentiation of embryonic and mesenchymal stem cells
(MSCs) (20).
MSCs are mechanically regulated by shear stress
generated in tissues by extravascular liquid flow. To date, the
regulation of YAP by shear stress has rarely been characterized in
the literature. YAP is regulated by fluid shear stress in
osteoblasts and stem cells (8,21).
Zhong et al (8) found that
increased YAP expression levels, triggered by flow shear stress,
increased osteogenesis and decreased adipogenesis of MSCs and
initiated dedifferentiation of chondrocytes. Furthermore, culturing
MSCs on microfluidic chips that mimic interstitial flow shear
stress increases YAP and TAZ activity in an Rho-Rho-associated
coiled-coil containing protein kinase (ROCK)-dependent manner,
enhancing osteogenic and fibrochondrogenic differentiation
(8,22). Dupont et al (3) used rigid vs. highly elastic
micropillars to assess the effects of cell-generated mechanical
force; on the elastic substrate, cytoplasmic localization of
YAP/TAZ was markedly increased. Collectively, it has been indicated
that YAP/TAZ responds to cytoskeletal tension (3). Wang et al (23) used a parallel flow chamber to
perform in vitro shear stress experiments and a partial
carotid ligation model in apolipoprotein E-deficient mice to
perform in vivo flow perturbation experiments. They proved
that the activation and expression levels of YAP/TAZ were lower in
the laminar flow region of the straight segment of thoracic aorta
than in the disturbed flow region of the inner curvature of aortic
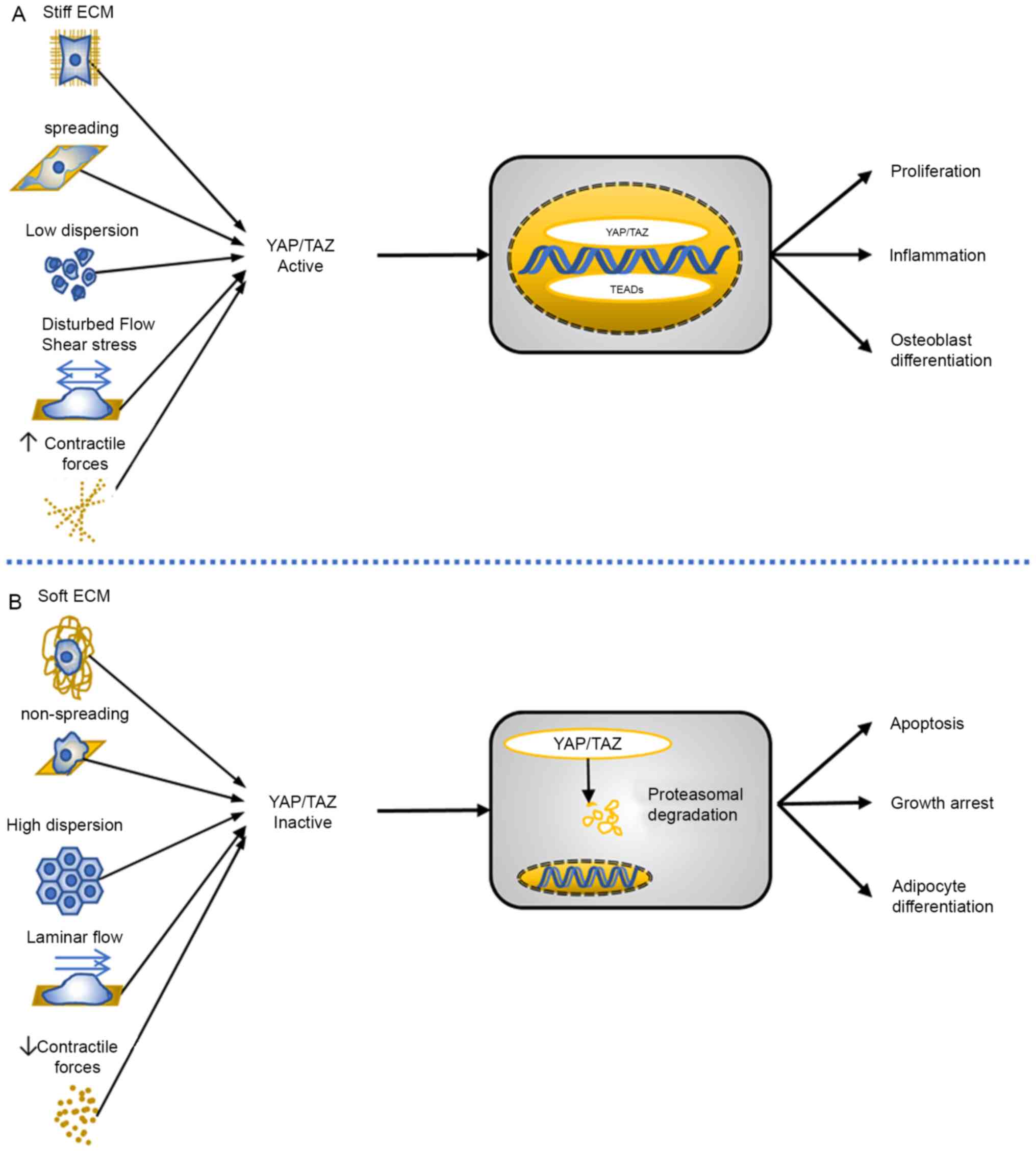
arch (Fig. 1) (23).
Each organ of the human body has a specific ECM
stiffness. The role of mechanical cues needs to be understood in
the context of the multicellular organization that characterizes
natural tissue (24,25). The importance of ECM sensing
elasticity has been demonstrated in fibroblasts, mesenchymal and
epidermal stem cells and other areas of cells (5,26–28).
For example, gelatinous polyacrylamide gel has been used as an
artificial cell matrix for in vitro adherence culture of
cells. The cell matrix was induced to take on a different stiffness
by adjusting the proportion of chemical components and human
epidermal stem cells were cultured in the matrix. The results
showed that different levels of stiffness had different effects on
the shape of cell growth (5,29).
When cells were cultured on a soft matrix, which is similar to
brain tissue, they exhibited characteristics of neurons; when cells
were cultured on a middle-stiffness matrix, which is similar to
muscle tissue, they showed characteristics of myocytes; and when
cells were cultured on a hard matrix, which is similar to collagen,
they showed characteristics of osteoblastic differentiation
(30). Thus, the varying elastic
properties of the different tissue types seem to influence tissue
regeneration.
Different basal shapes of cell culture have an
important effect on cell proliferation and differentiation. In
vitro cell culture experiments have demonstrated that cell
proliferation is fastest in the outer region of the culture dish
(31). In round dishes, this area
is the outermost circle, while in square dishes the fastest
proliferation occurs in the four corners. There is also research
suggesting that the topological terrain of the substrate serves an
important role in regulating the stemness of stem cells. Clones
formed in a groove and column are relatively flat, whereas clones
formed in a hexagonal pattern are more rounded (25,32).
Based on these reports, it is hypothesized that ECM shape induces
cell geometry and regulates cell fate and behavior. In similar
studies, microprinted ECM islands of different sizes were
engineered to control the extent of spreading of a single
endothelial cell (28,33,34).
It was observed that well-spread cells proliferated, whereas cells
confined to small adhesive areas did not proliferate and instead
underwent apoptosis. It has been proven that this effect was due to
changes in cell shape, rather than due to the extent of cell-ECM
contact (13).
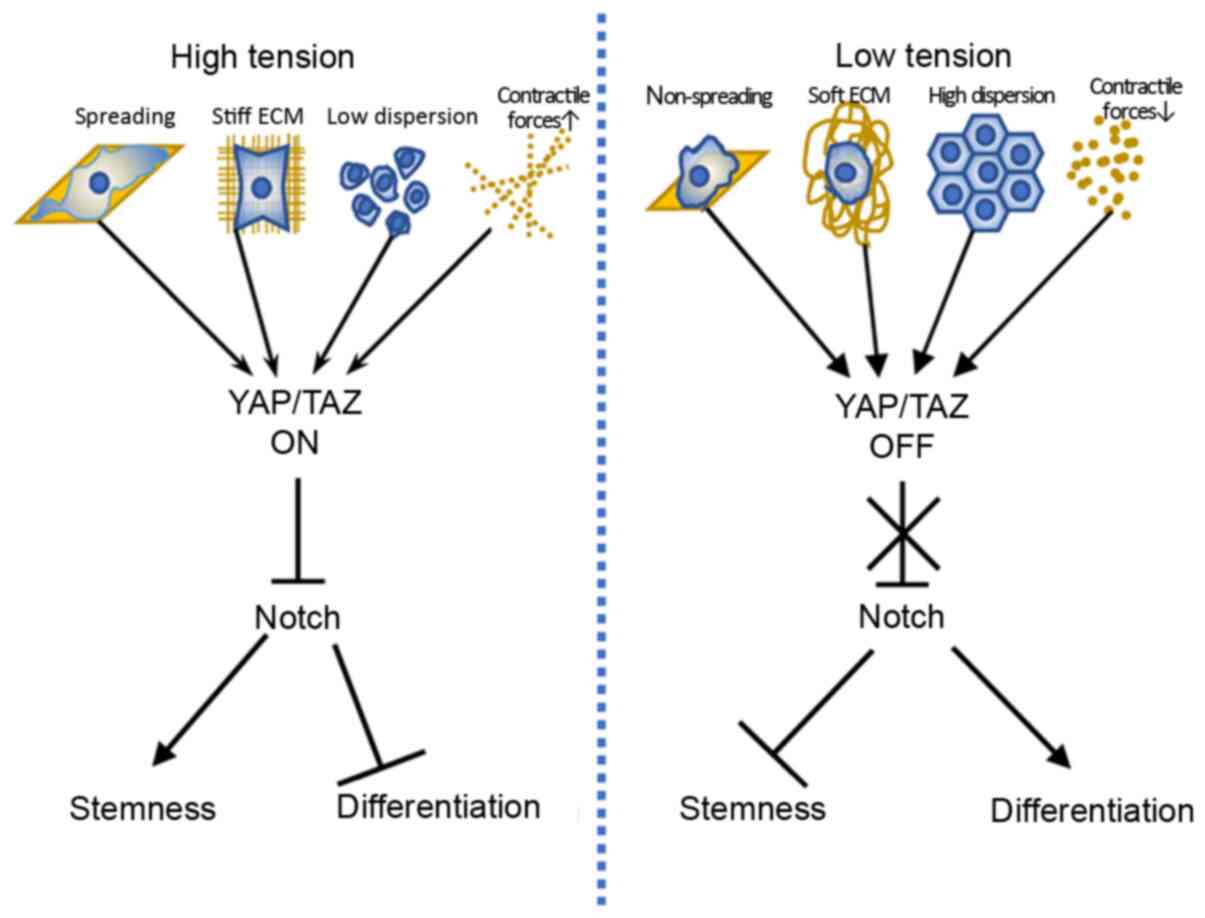
It has been proposed that cell fate induced by stiff
ECM requires YAP/TAZ function and, conversely, cell fate associated
with soft ECM requires its inactivation (Fig. 1) (35–37).
The activity of YAP/TAZ in cells grown on stiff hydrogels is
comparable to that of cells grown on plastics, whereas growing
cells on soft matrices inhibits YAP/TAZ activity to levels
comparable with that of short interfering RNA-mediated YAP/TAZ
depletion (3).
The cytoskeleton, within the cytoplasm, is a
fundamental structure for mediating force transmission (38,39).
It contains filamentous actin, intermediate filaments and
microtubules that transmit force over long distances in the cell.
Various types of external force applied to the cell can be
transformed into internal force via the cytoskeleton. Therefore,
force transmission contains both an external component and an
internal component and is completed via the cytoskeleton (40). Focal adhesions, a type of
multiprotein clustered from integrins, provide a direct physical
link between the ECM and cytoskeletal adaptors, consequently
connecting the ECM with the actin cytoskeleton (24). The organization of the cytoskeleton
sustains cell shape; however, the dynamics of actin microfilaments
are the focus of much research on mechanotransduction (41,42).
A number of stem cells differentiate into distinct
lineages depending on local cues present in their environment.
Previous studies have suggested that changes in cell shape regulate
the degree of development of lineage-specific markers, or
differentiation, in pre-committed preadipocytes or preosteoblasts
(10,48). Different basal shapes of cell
culture have an important effect on cell proliferation and
differentiation. Average focal-adhesion stress per cell increases
with micropost rigidity for both human MSCs and umbilical vein
endothelial cells but to different magnitudes in each cell type
(49,50). These differences in focal adhesion
stress between cell types suggest that there may be multiple ways
for cells to mechanically adapt to their environment (49).
Cells seeded on stiff hydrogels or large islands
show increased cell spreading and greater cell-ECM contact. YAP/TAZ
was localized in the nucleus when cells were seeded on
micropillars, indicating that YAP/TAZ are primarily regulated by
cell spreading imposed by the ECM (51,52).
Morphometric analysis of cell populations revealed a strong
correlation between focal adhesions and cell spreading, regardless
of micropost rigidity (49). These
findings indicate how ECM is able to mediate its effects on stem
cell morphology and stemness, and show the role of
mechanotransduction and stiffness on cell fate determination
(53,54).
Changes in shape and morphological characteristics
occur during and after cell differentiation. Previous studies have
proven that there is an important connection between cell
morphology and proliferation, survival and differentiation
(55,56). Cell shape may impact the state
transition of cell from life to death or between proliferation and
quiescence (57).
Fibronectin-coated micropillars have been used to
test whether YAP/TAZ is regulated by cell spreading independently
of the total amount of ECM (3,49). It
has been shown that the actual area available for cell-ECM
interaction is only ~10% of their projected area and YAP/TAZ
remained nuclear on micropillars, indicating that YAP/TAZ is
primarily regulated by cell spreading imposed by the ECM (Fig. 1) (3,51).
Similarly, Wada et al (58)
proposed a model by using a microdomain culture system in which the
cell area of a single cell was defined, while cell-cell contact was
prevented to show that cell morphology alone can modulate YAP
activity.
The geometrical morphology and area of cell spread
are associated with proliferation, differentiation and migration of
bone marrow MSCs. When the width of the cells is narrow, the
proliferation of bone marrow MSCs is inhibited and cell migration
is enhanced when the cell area is limited (59–61).
A growing body of research has revealed that YAP/TAZ
serves a central role in delivering information of mechanical
environments surrounding cells to the nucleus transcriptional
machinery via Hippo and non-Hippo mechanisms; this is an underlying
principle for YAP/TAZ-mediated regulation of biological functions
(9,62,63).
YAP also interacts with components of other signaling pathways,
which serve a role in mechanotransduction (64–66).
However, the focus of most research has been the mechanism by which
mechanical signals regulate YAP/TAZ activity.
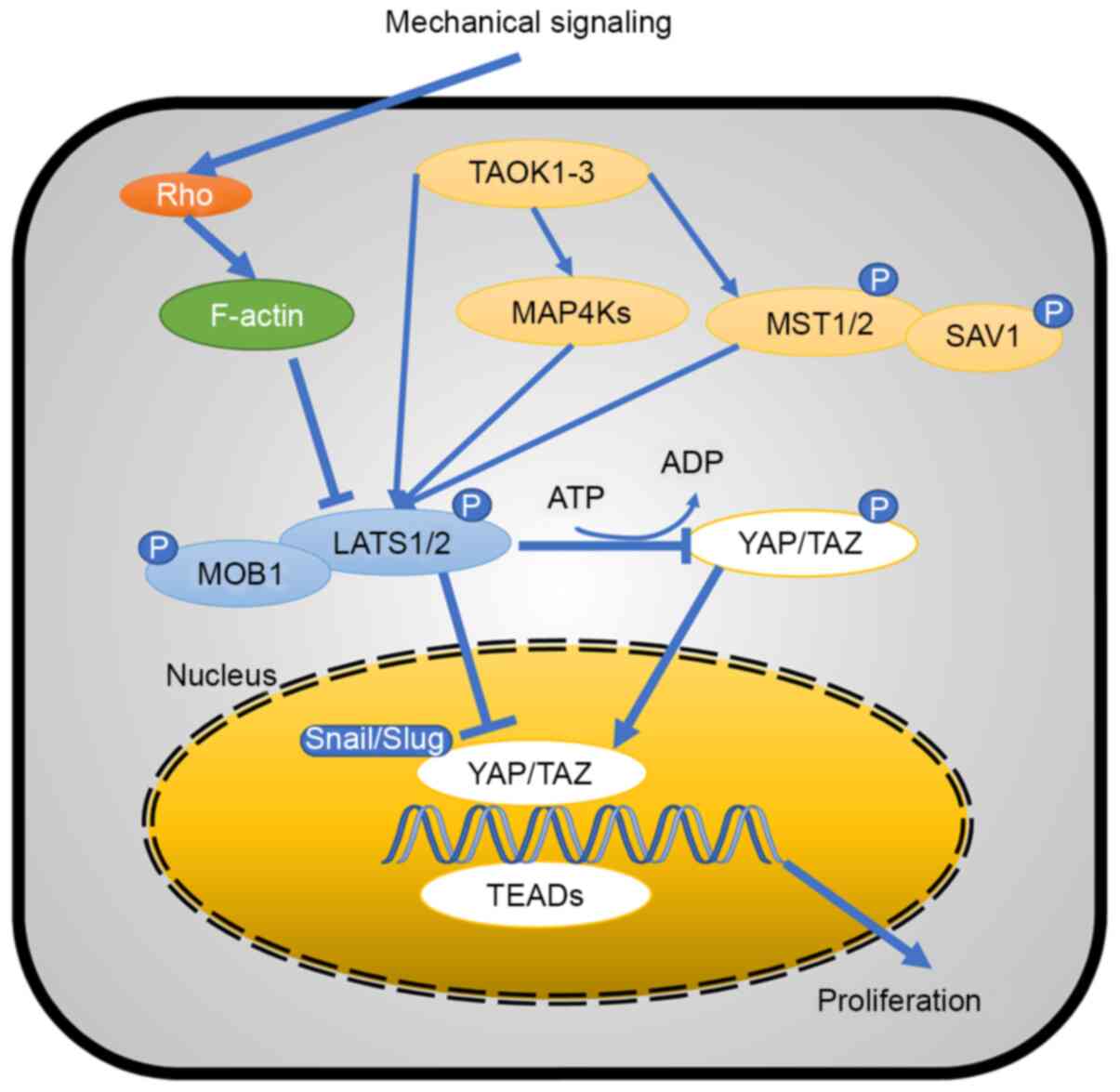
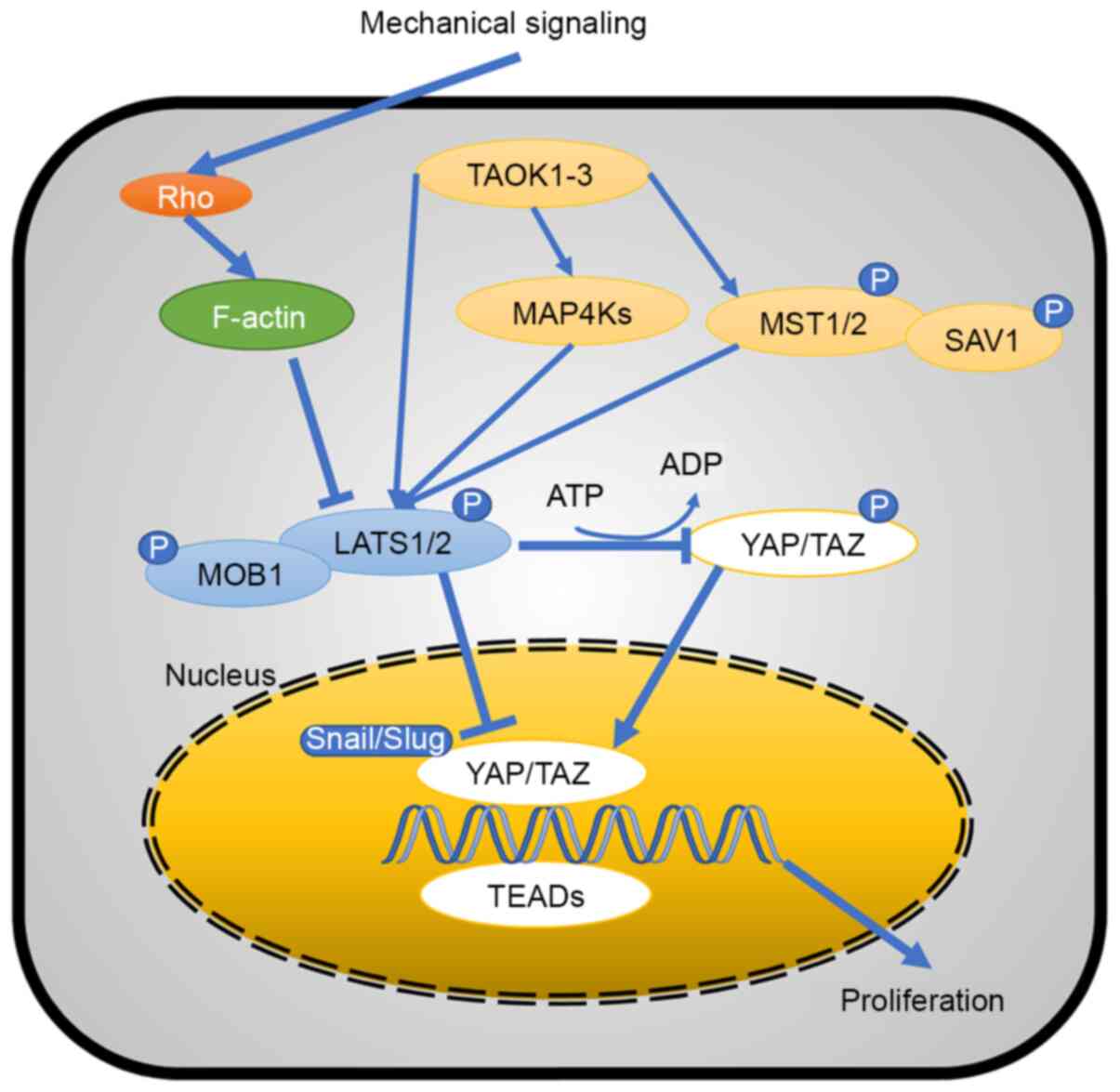
The Hippo pathway serves a key role in the
regulation of tissue homeostasis, abnormalities of which are
associated with human cancer. Research has provided an overview of
the functional importance of numerous Hippo pathway components. It
has been demonstrated that neurofibromin 2 and Ras Homolog Family
Member A (RhoA) are important regulators of YAP/TAZ, and TAO kinase
1/3 are direct kinases for large tumor suppressor kinase (LATS) 1/2
(67). Phosphorylation activates
LATS1/2, resulting in its activity as a kinase of YAP/TAZ (68). Hippo core components, as well as YAP
and TAZ activity, are regulated by a variety of mechanisms
(Fig. 2) (69–71).
There is a requirement for balanced amounts of YAP/TAZ in neural
stem and progenitor cells, which are involved in controlling the
correct expansion of the progenitor pool and timely
differentiation; this may be ensured by a crosstalk system between
CDK5 regulatory subunit-associated protein 2 and the Hippo pathway
(72). YAP/TAZ transfers into the
nucleus and participates in proliferation with the transcriptional
enhanced associate domain (TEADs) following dephosphorylation by
phosphatases, including protein phosphatase 1A and 2A (73–75).
Mechanical signals act on the Rho protein to promote
the effect of filamentous (F-)actin, which resists LATS1/2. LATS1/2
has a YAP/TAZ complex that inhibits phosphorylation of the
Snail/Slug-YAP/TAZ complex in the nucleus and cytoplasm, while
phosphorylated YAP/TAZ in the cytoplasm promotes the Snail-YAP/TAZ
complex formation. Therefore, mechanical signaling controls the
function of YAP/TAZ via the classic Hippo pathway (Fig. 2).
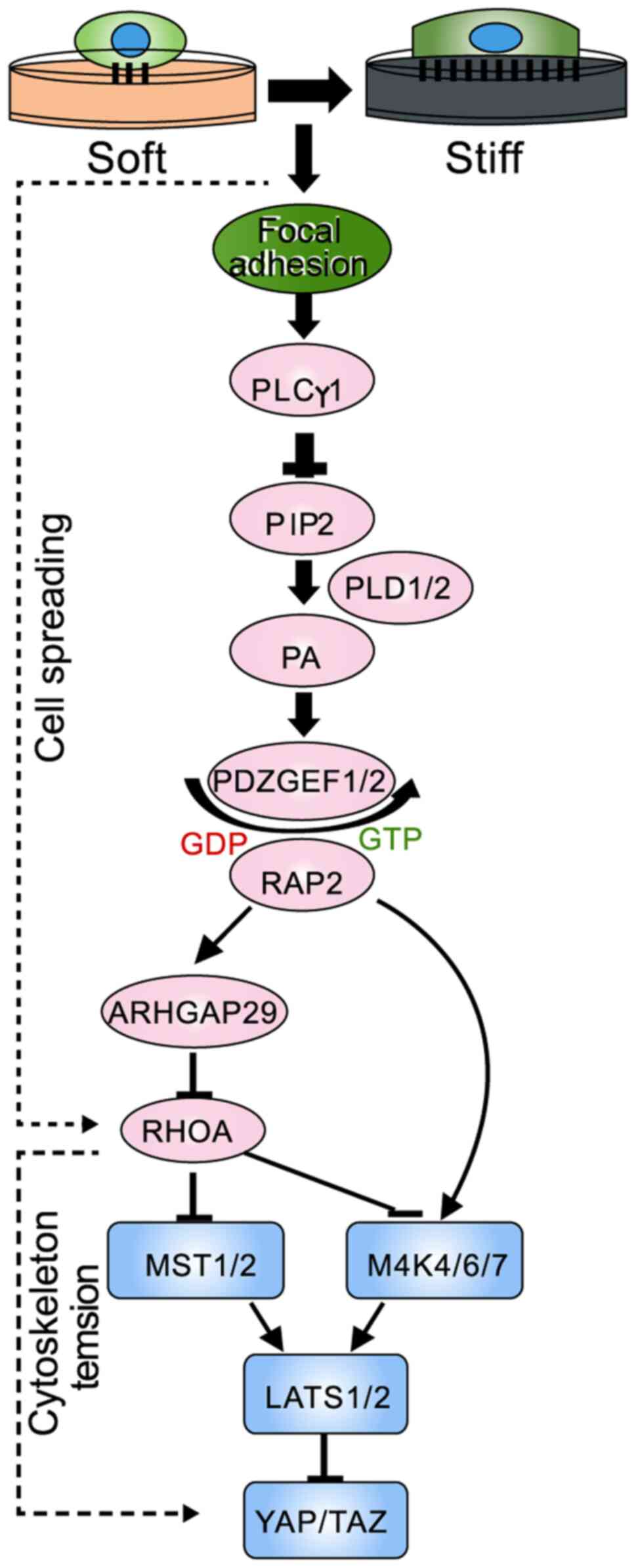
Ras-related protein 2 (RAP2) GTPase has been
identified as a key intracellular mechanotransducer that relays
extracellular mechanical signals for transcriptional regulation via
the Hippo pathway (76). ECM
stiffness acts via RAP2 and its downstream Hippo kinase cascade to
modulate a YAP/TAZ-mediated mechanoresponsive transcriptome. The
identification of this signaling axis provides mechanistic insights
into how cellular machinery is driven by mechanical stimuli
(Fig. 3). In addition, another
study validated the role of AT-rich interaction domain
(ARID)1A-containing SWitch/sucrose non-fermentable (SWI/SNF)
complex as a mechanoregulated inhibitor of YAP/TAZ in vivo
(77). In environments with high
levels of mechanical stress, nuclear F-actin binds to
ARID1A-SWI/SNF, thereby relieving YAP/TAZ from SWI/SNF inhibition
in favor of an association between TEAD and YAP/TAZ.
Tissue stiffness can be a predictor of tumor
development and carries a high risk of pathogenesis of malignant
transformation (78,79); to the best of our knowledge,
however, the association between tissue rigidity and tumor
pathogenesis has not been fully characterized. The situation is
similar for stem cell self-renewal and regulation of
differentiation (1). Research has
shown that tumor rigidity reflects an increase in stromal stiffness
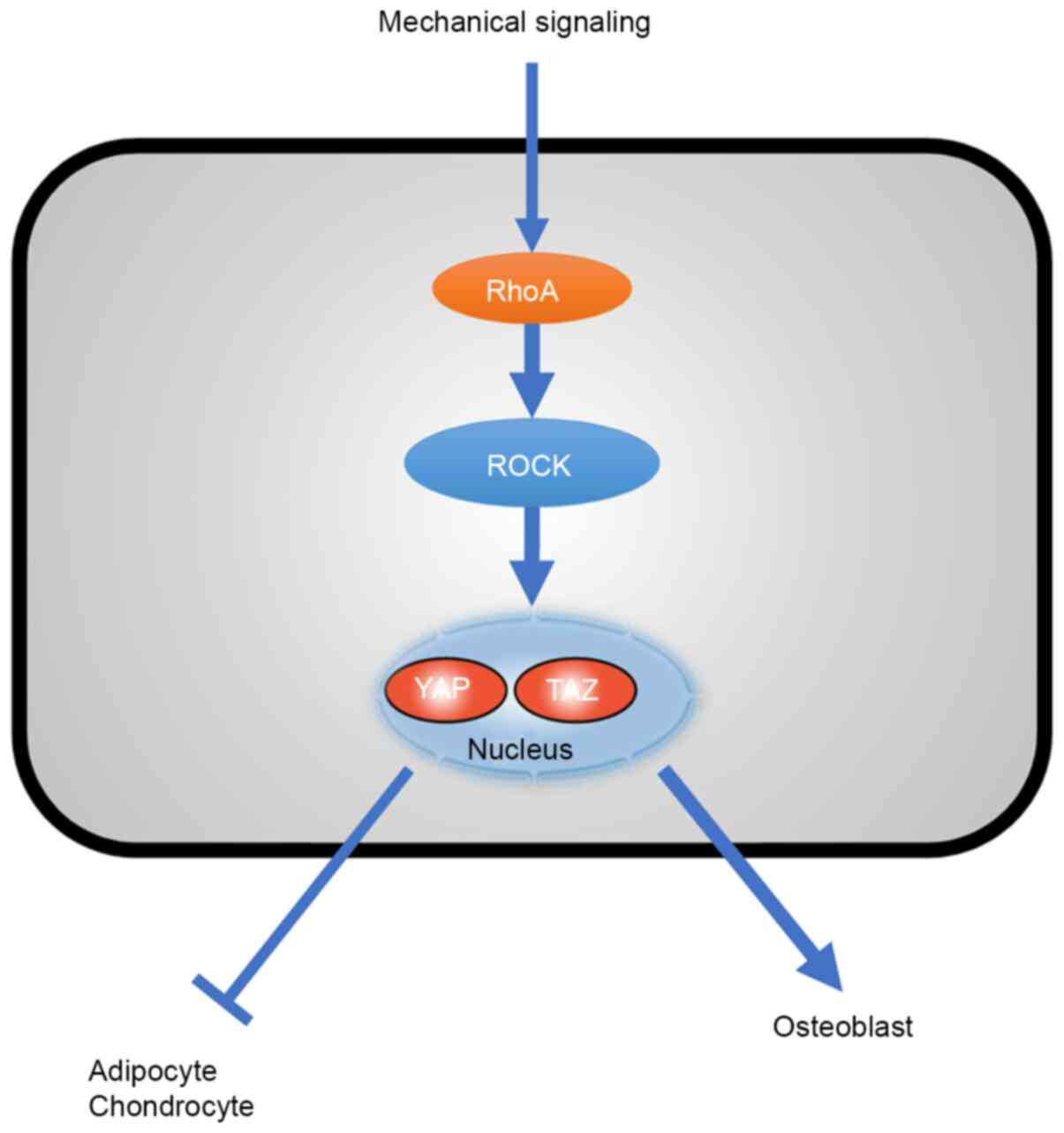
and tumor cell tension (26). Rho
GTPase-dependent ROCK activation upregulates cytoskeletal tension
by triggering the assembly of actomyosin networks (80). RhoA-mediated adipogenesis depends on
a circular or expanded shape, whereas in of RhoA-mediated
osteogenesis, RhoA effector ROCK constitutive activation induces
osteogenesis independent of cell shape (10). This RhoA-ROCK commitment signal
requires actin-myosin to produce tension. Studies have shown that
the commitment of stem cells to various lineages is modulated by
different physical cues embodied by cytoskeletal tension, cell
shape and RhoA signaling (Fig. 4)
(10,81,82).
It has been demonstrated that membrane type 1
(MT1)-MMP directs skeletal stem cell lineage commitment and
differentiation by controlling ECM remodeling, in turn promoting
cytoskeletal tension via the integrin-RhoA-ROCK pathway, thereby
triggering nuclear localization of YAP/TAZ co-transcriptional
activators, which activates a program that favors osteogenic
differentiation over alternative adipogenic and chondrogenic cell
fate (52).
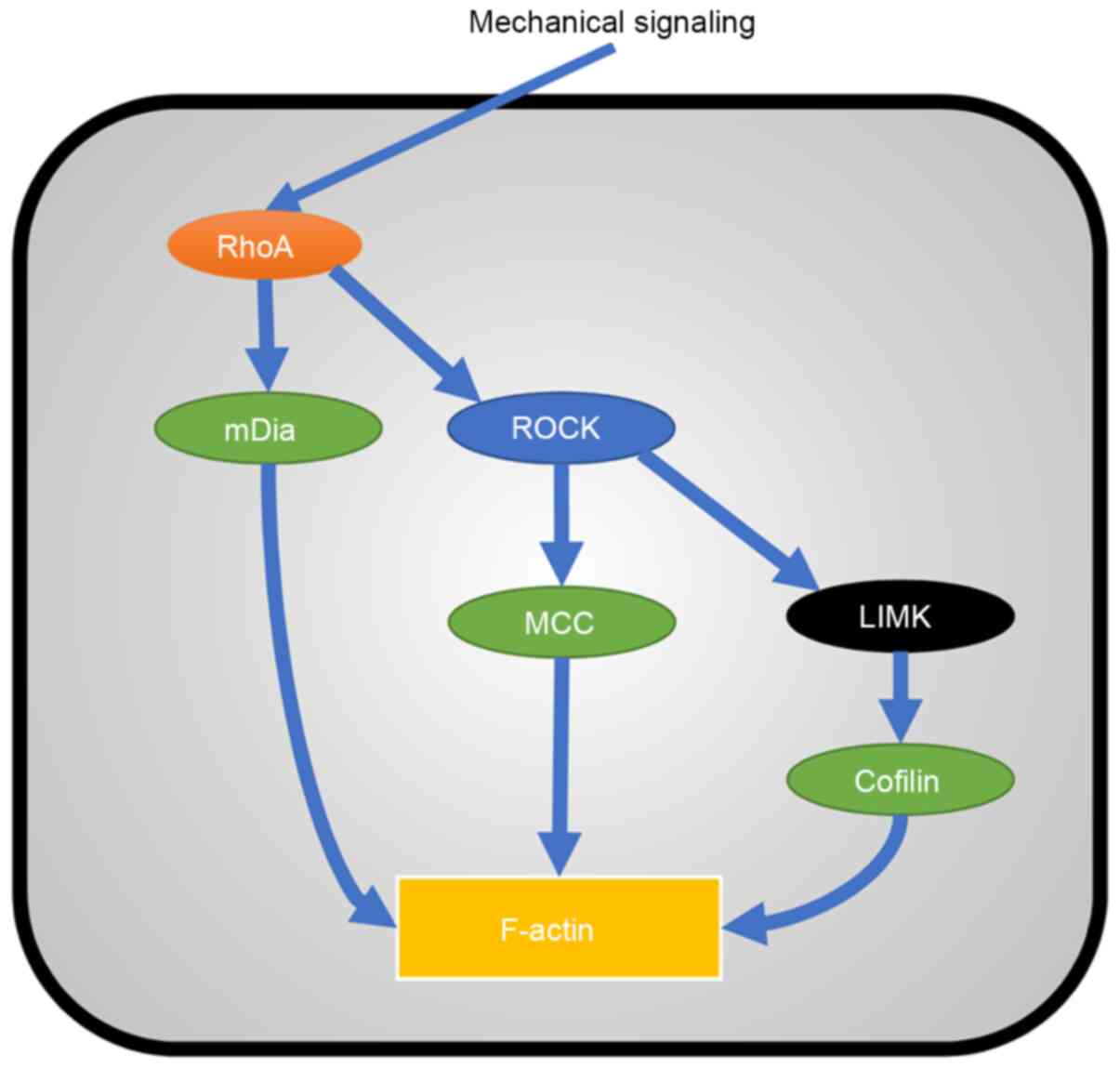
Both TGF-β canonical and non-canonical signaling
cascades simultaneously occur via crosstalk of core pathway
components and combined utilization of SMAD/non-SMAD transcription
factors (83). Tissue mechanics are
translated into the activation or inhibition of transcription
factors and their co-activating factors (84). Previous research has investigated
the impact of morphological alterations on cellular signaling of
TGFβ-1 (85,86). From the non-canonical TGF-β
signaling pathway, Rho and ROCK activate F-actin and RhoA activates
ROCK and mammalian diaphanous (mDia); ROCK activates MCC regulator
of Wnt signaling pathway (MCC) and LIM domain kinase (LIMK)
(87); LIMK activates Cofilin; and
mDia, MCC and Cofilin activate F-actin (88). This is a cascade amplification
process (Fig. 5). Canonical
TGFβ/bone morphogenetic protein (BMP) signaling acts via SMADs and
has also been shown to be sensitive to mechanical inputs into the
cell (89). It has been confirmed
that YAP/TAZ are mechanoregulators of TGF-β-SMAD signaling
(66). YAP/TAZ interaction with
SMAD2/3 serves a key role in SMAD nuclear-cytoplasmic shuttling
(90). YAP nuclear exclusion
sequesters the SMAD2/3 protein to the cytoplasm and therefore
suppresses TGF-β signaling (91,92).
YAP binds to activated SMAD1 protein to enhance BMP-induced
transcriptional activity (93). In
addition, YAP/TAZ functions as an endogenous repressor of SMAD7
expression to modulate TGF-β signaling (94).
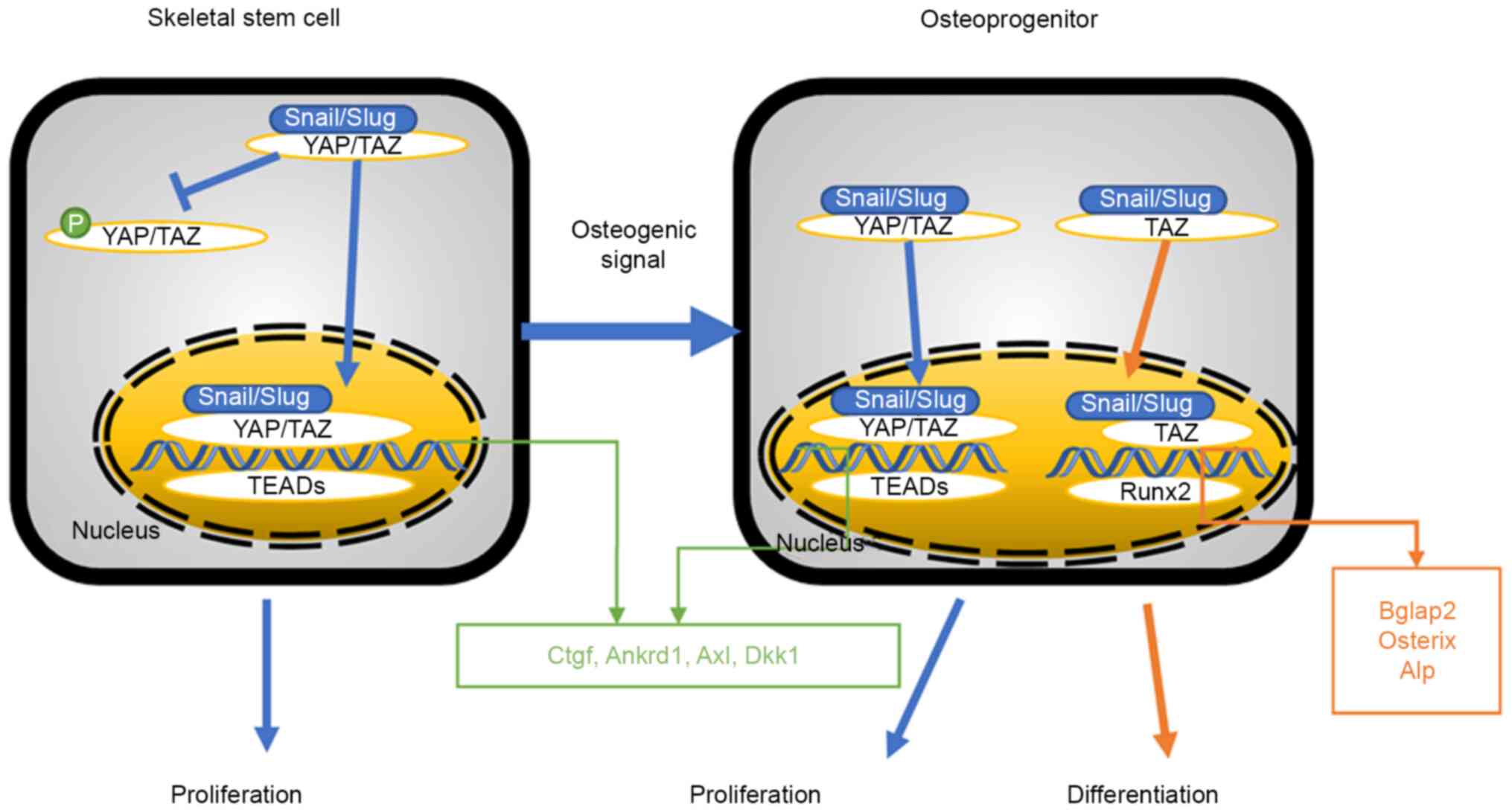
Bone-marrow-derived skeletal stem cell (SSC)
self-renewal and function is key to bone development, in
vivo balance and repair (95).
Mechanically, Snail/segment regulates SSC function by forming a
complex with transcription co-activators YAP and TAZ and
suppressing Hippo pathway-dependent adjustment of the YAP/TAZ
signal cascade. In turn, the Snail/Slug-YAP/TAZ axis activates a
series of YAP/TAZ-TEAD/Runx2 downstream targets that control the
stability and osteogenesis of SSCs (65). In conclusion, these results indicate
that SSCs mobilize the Snail/Slug-YAP/TAZ complex to control stem
cell function (65).
The Snail/Slug-YAP/TAZ complex promotes YAP/TAZ
phosphorylation in the cytoplasm and Snail/Slug-YAP/TAZ-TEADs
complex in the nucleus, thus inducing expression of connective
tissue growth factor, ankyrin repeat domain 1, axonin l and
Dickkopf-related protein 1, as well as cell proliferation (96). This is established in SSCs and
osteoprogenitors. In osteoprogenitors, the Snail/Slug-TAZ complex
promotes the Snail/Slug-TAZ-Runx2 complex in the nucleus (97), thereby inducing the expression of
bone γ-carboxyglutamate protein 2, Osterix and Alp, and inducing
differentiation (Fig. 6).
Functional interactions between Hippo and the Sonic
hedgehog (Shh) signaling pathway have been observed in
medulloblastoma and neural differentiation (98,99).
Shh expression in the regenerating limb bud in Xenopus can
be decreased under the functional inhibition of YAP (100). Furthermore, the Hedgehog pathway
acts upstream of the Hippo pathway in regulating follicle stem cell
maintenance in the Drosophila ovary, in which the Hedgehog
pathway regulates Yorkie activity via a post-translational
mechanism (101). It has been
observed that in non-alcoholic steatohepatitis-associated liver
injury and inflammation, injured hepatocytes release Shh ligands,
which promotes the YAP-induced accumulation of reactive-appearing
ductular cells (102). However, in
healthy liver, the activation of YAP does not result in its growth
(103). YAP/TAZ inhibition of
smooth muscle cell differentiation is mediated by the Hedgehog
pathway, but signal transduction in mouse embryonic pluripotent
mesenchymal cells is not (104,105). This suggests a working interaction
between YAP and Shh signal transduction.
It has been reported that following loss of cell
attachment to the basement membrane, epidermis cells begin a
terminal differentiation program until they fall off from the
tissue form surface (106–109). As YAP/TAZ serves an important role
in organ size control during the period of embryonic development
(44,110–114) and the notch signaling pathway is
core in the paradigm of the epidermis, it is hypothesized that
YAP/TAZ and notch regulation have a synergistic effect with each
other to control stem cell preservation vs. differentiation and
intercellular communication (Fig.
7) (109). This was also
confirmed by Totaro et al (109), who demonstrated that notch was
downstream of YAP/TAZ and that inhibition of YAP/TAZ preserved the
undifferentiated state of human epidermal stem cells, although the
specific mechanism remains unclear.
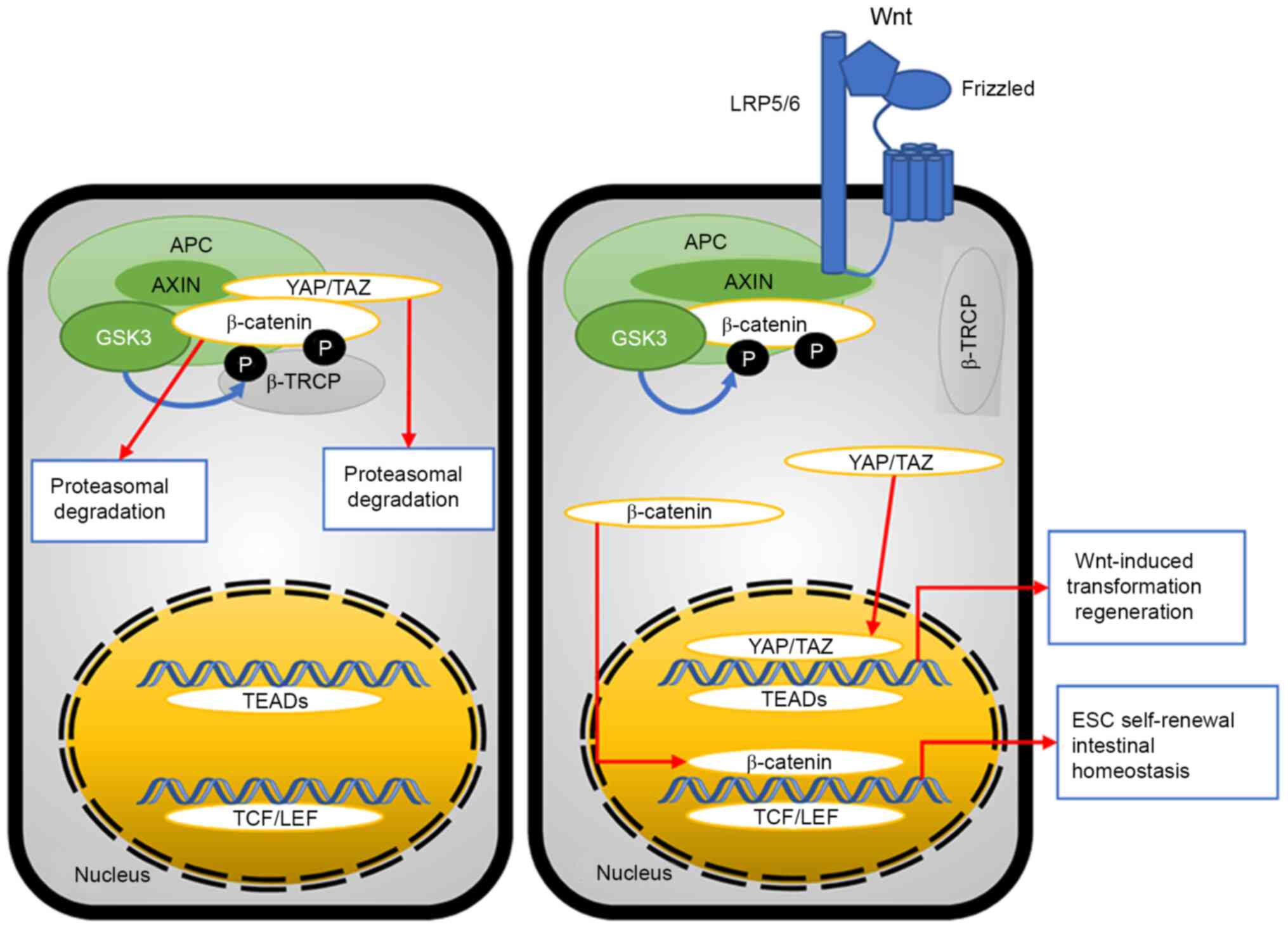
YAP/TAZ participates in the Wnt/β-catenin pathway,
which is involved in cell proliferation, tumorigenesis,
regeneration and stem cell expansion (115–117). It has been confirmed that the
primary purpose of the YAP/TAZ-associated Wnt/β pathway is control
of β-catenin (118,119). In cells lacking the Wnt signal,
YAP and TAZ are sequestered in the β-catenin destruction complex
and they play an essential role for phospho-β-catenin degradation
by recruiting β-transducin repeats-containing proteins (β-TRCP)
(118). As a result, there is no
reason that YAP/TAZ has to be banded with TEADs or β-catenin has to
be banded with TCF/LEF in the nucleus to perform their respective
roles. (Fig. 8). In cells
possessing the Wnt signal, β-TRCP is released, as well as β-catenin
and YAP/TAZ; YAP/TAZ and β-catenin can transfer into the nucleus to
form YAP/TAZ-TEAD and β-catenin-TCF/LEF complexes, resulting in
Wnt-induced transformation regeneration and ESC self-renewal
(117,120). In addition, YAP/TAZ is the key
downstream effector of alternative Wnt signaling to induce cell
migration and osteogenic differentiation, as well as negative
regulation of canonical Wnt/β-catenin signaling (119). The functional interactions between
Wnt activation and YAP localization have also been demonstrated in
maintaining undifferentiated mouse ESCs (64,121).
Cytoplasmic YAP/TAZ can sequester β-catenin, resulting in the loss
of self-renewal ability and differentiation activation of ESCs
(121).
The JNK pathway, one of the major signaling
cassettes of the MAPK signaling pathway, serves an important role
in cytoskeletal modulation, apoptosis and cell proliferation
(122–124). JNK is a stress-activated kinase,
which can be activated by stretch-induced actin stress (123,125). LATS activity can be decreased by
cyclic stretch stress due to increased LIM domain-containing
1-LATS1 banding through activation of JNK, consequently, leading to
elevated YAP activity (126). In
addition, YAP/TAZ activation induced by disturbed flow shear stress
can promote inflammation and atherogenesis by enhancing JNK
activity (127).
Future research will investigate the mechanisms by
which YAP/TAZ modulates cell mechanics to enable cell function and
the role of YAP/TAZ in ECM shape-induced stem cell function and
shape/geometry.
Not applicable.
The present review was supported by the National
Natural Science Foundation of China (grant no. 81601619) and
Science Research Foundation of Education Department of Liaoning
(grant no. LZ2019008).
Not applicable.
YL and WZ contributed to the conception of the
review, wrote the manuscript, performed the literature search and
prepared the original draft. JW performed the literature search and
constructed the figures. All the authors read and approved the
final manuscript. Data sharing is not applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Vining KH and Mooney DJ: Mechanical forces
direct stem cell behaviour in development and regeneration. Nat Rev
Mol Cell Biol. 18:728–742. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Demehri S and Kopan R: Notch signaling in
bulge stem cells is not required for selection of hair follicle
fate. Development. 136:891–896. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dupont S, Morsut L, Aragona M, Enzo E,
Giulitti S, Cordenonsi M, Zanconato F, Le Digabel J, Forcato M,
Bicciato S, et al: Role of YAP/TAZ in mechanotransduction. Nature.
474:179–183. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lee JH, Park HK and Kim KS: Intrinsic and
extrinsic mechanical properties related to the differentiation of
mesenchymal stem cells. Biochem Biophys Res Commun. 473:752–757.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Engler AJ, Sen S, Sweeney HL and Discher
DE: Matrix elasticity directs stem cell lineage specification.
Cell. 126:677–689. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li D, Zhou J, Chowdhury F, Cheng J, Wang N
and Wang F: Role of mechanical factors in fate decisions of stem
cells. Regen Med. 6:229–240. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Oh S, Brammer KS, Li YS, Teng D, Engler
AJ, Chien S and Jin S: Stem cell fate dictated solely by altered
nanotube dimension. Proc Natl Acad Sci USA. 106:2130–2135. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhong W, Tian K, Zheng X, Li L, Zhang W,
Wang S and Qin J: Mesenchymal stem cell and chondrocyte fates in a
multishear microdevice are regulated by Yes-associated protein.
Stem Cells Deve. 22:2083–2093. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ishihara E and Nishina H: Role of
Hippo-YAP/TAZ signaling pathway in mechanotransduction. Clin
Calcium. 26:1751–1756. 2016.(In Japanese). PubMed/NCBI
|
|
10
|
McBeath R, Pirone DM, Nelson CM,
Bhadriraju K and Chen CS: Cell shape, cytoskeletal tension, and
RhoA regulate stem cell lineage commitment. Dev Cell. 6:483–495.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dupont S: Role of YAP/TAZ in cell-matrix
adhesion-mediated signalling and mechanotransduction. Exp Cell Res.
343:42–53. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pocaterra A, Romani P and Dupont S:
YAP/TAZ functions and their regulation at a glance. J Cell Sci.
133:jcs2304252020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pan JX, Xiong L, Zhao K, Zeng P, Wang B,
Tang FL, Sun D, Guo HH, Yang X, Cui S, et al: YAP promotes
osteogenesis and suppresses adipogenic differentiation by
regulating β-catenin signaling. Bone Res. 6:182018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Oliver-De La Cruz J, Nardone G, Vrbsky J,
Pompeiano A, Perestrelo AR, Capradossi F, Melajová K, Filipensky P
and Forte G: Substrate mechanics controls adipogenesis through YAP
phosphorylation by dictating cell spreading. Biomaterials.
205:64–80. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yang Y, Wang BK, Chang ML, Wan ZQ and Han
GL: Cyclic stretch enhances osteogenic differentiation of human
periodontal ligament cells via YAP activation. BioMed Res Int.
2018:21748242018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kim NG, Koh E, Chen X and Gumbiner BM:
E-cadherin mediates contact inhibition of proliferation through
Hippo signaling-pathway components. Proc Natl Acad Sci USA.
108:11930–11935. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xue X, Hong X, Li Z, Deng CX and Fu J:
Acoustic tweezing cytometry enhances osteogenesis of human
mesenchymal stem cells through cytoskeletal contractility and YAP
activation. Biomaterials. 134:22–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hu JK, Du W, Shelton SJ, Oldham MC,
DiPersio CM and Klein OD: An FAK-YAP-mTOR signaling axis regulates
stem cell-based tissue renewal in mice. Cell Stem Cell.
21:91–106.e6. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lecarpentier E, Bhatt M, Bertin GI,
Deloison B, Salomon LJ, Deloron P, Fournier T, Barakat AI and
Tsatsaris V: Computational fluid dynamic simulations of maternal
circulation: Wall shear stress in the human placenta and its
biological implications. PLoS One. 11:e01472622016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Adamo L and Garcia-Cardeña G: Directed
stem cell differentiation by fluid mechanical forces. Antioxid
Redox Signal. 15:1463–1473. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kaneko K, Ito M, Naoe Y, Lacy-Hulbert A
and Ikeda K: Integrin alphav in the mechanical response of
osteoblast lineage cells. Biochem Biophys Res Commun. 447:352–357.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhong W, Zhang W, Wang S and Qin J:
Regulation of fibrochondrogenesis of mesenchymal stem cells in an
integrated microfluidic platform embedded with biomimetic
nanofibrous scaffolds. PLoS One. 8:e612832013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang KC, Yeh YT, Nguyen P, Limqueco E,
Lopez J, Thorossian S, Guan KL, Li YJ and Chien S: Flow-dependent
YAP/TAZ activities regulate endothelial phenotypes and
atherosclerosis. Proc Natl Acad Sci USA. 113:11525–11530. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Halder G, Dupont S and Piccolo S:
Transduction of mechanical and cytoskeletal cues by YAP and TAZ.
Nat Rev Mol Cell Biol. 13:591–600. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang Y, Wang G, Luo X, Qiu J and Tang C:
Substrate stiffness regulates the proliferation, migration, and
differentiation of epidermal cells. Burns. 38:414–420. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Paszek MJ, Zahir N, Johnson KR, Lakins JN,
Rozenberg GI, Gefen A, Reinhart-King CA, Margulies SS, Dembo M,
Boettiger D, et al: Tensional homeostasis and the malignant
phenotype. Cancer Cell. 8:241–254. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Discher DE, Janmey P and Wang YL: Tissue
cells feel and respond to the stiffness of their substrate.
Science. 310:1139–1143. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Connelly JT, Gautrot JE, Trappmann B, Tan
DW, Donati G, Huck WT and Watt FM: Actin and serum response factor
transduce physical cues from the microenvironment to regulate
epidermal stem cell fate decisions. Nat Cell Biol. 12:711–718.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li Z, Gong Y, Sun S, Du Y, Lü D, Liu X and
Long M: Differential regulation of stiffness, topography, and
dimension of substrates in rat mesenchymal stem cells.
Biomaterials. 34:7616–7625. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hadden WJ, Young JL, Holle AW, McFetridge
ML, Kim DY, Wijesinghe P, Taylor-Weiner H, Wen JH, Lee AR, Bieback
K, et al: Stem cell migration and mechanotransduction on linear
stiffness gradient hydrogels. Proc Natl Acad Sci USA.
114:5647–5652. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Nelson CM and Bissell MJ: Modeling dynamic
reciprocity: Engineering three-dimensional culture models of breast
architecture, function, and neoplastic transformation. Semin Cancer
Biol. 15:342–352. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Witkowska-Zimny M, Walenko K, Wrobel E,
Mrowka P, Mikulska A and Przybylski J: Effect of substrate
stiffness on the osteogenic differentiation of bone marrow stem
cells and bone-derived cells. Cell Biol Int. 37:608–616. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Brusatin G, Panciera T, Gandin A, Citron A
and Piccolo S: Biomaterials and engineered microenvironments to
control YAP/TAZ-dependent cell behaviour. Nat Mater. 17:1063–1075.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Singhvi R, Kumar A, Lopez GP,
Stephanopoulos GN, Wang DI, Whitesides GM and Ingber DE:
Engineering cell shape and function. Science. 264:696–698. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kuroda M, Wada H, Kimura Y, Ueda K and
Kioka N: Vinculin promotes nuclear localization of TAZ to inhibit
ECM stiffness-dependent differentiation into adipocytes. J Cell
Sci. 130:989–1002. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Musah S, Morin SA, Wrighton PJ, Zwick DB,
Jin S and Kiessling LL: Glycosaminoglycan-binding hydrogels enable
mechanical control of human pluripotent stem cell self-renewal. ACS
Nano. 6:10168–10177. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Caliari SR, Vega SL, Kwon M, Soulas EM and
Burdick JA: Dimensionality and spreading influence MSC YAP/TAZ
signaling in hydrogel environments. Biomaterials. 103:314–323.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang N, Butler JP and Ingber DE:
Mechanotransduction across the cell surface and through the
cytoskeleton. Science. 260:1124–1127. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Eyckmans J, Boudou T, Yu X and Chen CS: A
hitchhiker's guide to mechanobiology. Deve Cell. 21:35–47. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Dogterom M, Kerssemakers JW, Romet-Lemonne
G and Janson ME: Force generation by dynamic microtubules. Curr
Opin Cell Biol. 17:67–74. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Vogel V and Sheetz M: Local force and
geometry sensing regulate cell functions. Nat Rev Mol Cell Biol.
7:265–275. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Schwartz MA: Integrins and extracellular
matrix in mechanotransduction. Cold Spring Harb Perspect Biol.
2:a0050662010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Fernandez BG, Gaspar P, Bras-Pereira C,
Jezowska B, Rebelo SR and Janody F: Actin-Capping Protein and the
Hippo pathway regulate F-actin and tissue growth in Drosophila.
Development. 138:2337–2346. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sansores-Garcia L, Bossuyt W, Wada K,
Yonemura S, Tao C, Sasaki H and Halder G: Modulating F-actin
organization induces organ growth by affecting the Hippo pathway.
EMBO J. 30:2325–2335. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chakraborty S, Njah K, Pobbati AV, Lim YB,
Raju A, Lakshmanan M, Tergaonkar V, Lim CT and Hong W: Agrin as a
Mechanotransduction signal regulating YAP through the hippo
pathway. Cell Rep. 18:2464–2479. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Elosegui-Artola A, Andreu I, Beedle AEM,
Lezamiz A, Uroz M, Kosmalska AJ, Oria R, Kechagia JZ, Rico-Lastres
P, Le Roux AL, et al: Force triggers YAP nuclear entry by
regulating transport across nuclear pores. Cell. 171:1397–1410.e14.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Dasgupta I and McCollum D: Control of
cellular responses to mechanical cues through YAP/TAZ regulation. J
Biol Chem. 294:17693–17706. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Cawthorn WP, Scheller EL and MacDougald
OA: Adipose tissue stem cells meet preadipocyte commitment: Going
back to the future. J Lipid Res. 53:227–246. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Fu J, Wang YK, Yang MT, Desai RA, Yu X,
Liu Z and Chen CS: Mechanical regulation of cell function with
geometrically modulated elastomeric substrates. Nat Methods.
7:733–736. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Geng Y and Wang Z: Review of cellular
mechanotransduction on micropost substrates. Med Biol Eng Comput.
54:249–271. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Sero JE and Bakal C: Multiparametric
analysis of cell shape demonstrates that beta-PIX directly couples
YAP activation to extracellular matrix adhesion. Cell Syst.
4:84–96.e86. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Tang Y, Rowe RG, Botvinick EL, Kurup A,
Putnam AJ, Seiki M, Weaver VM, Keller ET, Goldstein S, Dai J, et
al: MT1-MMP-dependent control of skeletal stem cell commitment via
a β1-integrin/YAP/TAZ signaling axis. Dev Cell. 25:402–416. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Gattazzo F, Urciuolo A and Bonaldo P:
Extracellular matrix: A dynamic microenvironment for stem cell
niche. Biochim Biophys Acta. 1840:2506–2519. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Lu D, Luo C, Zhang C, Li Z and Long M:
Differential regulation of morphology and stemness of mouse
embryonic stem cells by substrate stiffness and topography.
Biomaterials. 35:3945–3955. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Pucci B, Kasten M and Giordano A: Cell
cycle and apoptosis. Neoplasia. 2:291–299. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Pittenger MF, Discher DE, Peault BM,
Phinney DG, Hare JM and Caplan AI: Mesenchymal stem cell
perspective: Cell biology to clinical progress. NPJ Regen Med.
4:222019. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Chen CS, Mrksich M, Huang S, Whitesides GM
and Ingber DE: Geometric control of cell life and death. Science.
276:1425–1428. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Wada K, Itoga K, Okano T, Yonemura S and
Sasaki H: Hippo pathway regulation by cell morphology and stress
fibers. Development. 138:3907–3914. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Pek YS, Wan AC and Ying JY: The effect of
matrix stiffness on mesenchymal stem cell differentiation in a 3D
thixotropic gel. Biomaterials. 31:385–391. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Burke DP and Kelly DJ: Substrate stiffness
and oxygen as regulators of stem cell differentiation during
skeletal tissue regeneration: A mechanobiological model. PLoS One.
7:e407372012. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Tse JR and Engler AJ: Stiffness gradients
mimicking in vivo tissue variation regulate mesenchymal stem cell
fate. PLoS One. 6:e159782011. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Panciera T, Azzolin L, Cordenonsi M and
Piccolo S: Mechanobiology of YAP and TAZ in physiology and disease.
Nat Rev Mol Cell Biol. 18:758–770. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Hansen CG, Moroishi T and Guan KL: YAP and
TAZ: A nexus for Hippo signaling and beyond. Trends Cell Biol.
25:499–513. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Bejoy J, Song L and Li Y: Wnt-YAP
interactions in the neural fate of human pluripotent stem cells and
the implications for neural organoid formation. Organogenesis.
12:1–15. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Tang Y, Feinberg T, Keller ET, Li XY and
Weiss SJ: Snail/Slug binding interactions with YAP/TAZ control
skeletal stem cell self-renewal and differentiation. Nat Cell Biol.
18:917–929. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Szeto SG, Narimatsu M, Lu M, He X, Sidiqi
AM, Tolosa MF, Chan L, De Freitas K, Bialik JF, Majumder S, et al:
YAP/TAZ Are mechanoregulators of TGF-β-Smad signaling and renal
fibrogenesis. J Am Soc Nephrol. 27:3117–3128. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Plouffe SW, Meng Z, Lin KC, Lin B, Hong
AW, Chun JV and Guan KL: Characterization of hippo pathway
components by gene inactivation. Mol Cell. 64:993–1008. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Zhao B, Wei X, Li W, Udan RS, Yang Q, Kim
J, Xie J, Ikenoue T, Yu J, Li L, et al: Inactivation of YAP
oncoprotein by the Hippo pathway is involved in cell contact
inhibition and tissue growth control. Genes Dev. 21:2747–2761.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Bae JS, Kim SM and Lee H: The Hippo
signaling pathway provides novel anti-cancer drug targets.
Oncotarget. 8:16084–16098. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Wang C, Gu C, Jeong KJ, Zhang D, Guo W, Lu
Y, Ju Z, Panupinthu N, Yang JY, Gagea MM, et al: YAP/TAZ-mediated
upregulation of GAB2 leads to increased sensitivity to growth
factor-induced activation of the PI3K pathway. Cancer Res.
77:1637–1648. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Wang L, Luo JY, Li B, Tian XY, Chen LJ,
Huang Y, Liu J, Deng D, Lau CW, Wan S, et al: Integrin-YAP/TAZ-JNK
cascade mediates atheroprotective effect of unidirectional shear
flow. Nature. 540:579–582. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Sukumaran SK, Stumpf M, Salamon S, Ahmad
I, Bhattacharya K, Fischer S, Müller R, Altmüller J, Budde B,
Thiele H, et al: CDK5RAP2 interaction with components of the Hippo
signaling pathway may play a role in primary microcephaly. Mol
Genet Genomics. 292:365–383. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Schlegelmilch K, Mohseni M, Kirak O,
Pruszak J, Rodriguez JR, Zhou D, Kreger BT, Vasioukhin V, Avruch J,
Brummelkamp TR and Camargo FD: Yap1 acts downstream of α-catenin to
control epidermal proliferation. Cell. 144:782–795. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Wang P, Bai Y, Song B, Wang Y, Liu D, Lai
Y, Bi X and Yuan Z: PP1A-mediated dephosphorylation positively
regulates YAP2 activity. PLoS One. 6:e242882011. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Denis D, Rouleau C and Schaffhausen BS: A
transformation-defective polyomavirus middle T antigen with a novel
defect in PI3 kinase signaling. J Virol. 91:e01774–16. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Meng Z, Qiu Y, Lin KC, Kumar A, Placone
JK, Fang C, Wang KC, Lu S, Pan M, Hong AW, et al: RAP2 mediates
mechanoresponses of the Hippo pathway. Nature. 560:655–660. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Chang L, Azzolin L, Di Biagio D, Zanconato
F, Battilana G, Lucon Xiccato R, Aragona M, Giulitti S, Panciera T,
Gandin A, et al: The SWI/SNF complex is a mechanoregulated
inhibitor of YAP and TAZ. Nature. 563:265–269. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Singh A, Brito I and Lammerding J: Beyond
tissue stiffness and bioadhesivity: Advanced biomaterials to model
tumor microenvironments and drug resistance. Trends Cancer.
4:281–291. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Lopez JI, Mouw JK and Weaver VM:
Biomechanical regulation of cell orientation and fate. Oncogene.
27:6981–6993. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Hoon JL, Tan MH and Koh CG: The regulation
of cellular responses to mechanical cues by Rho GTPases. Cells.
5:172016. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Spector AA and Grayson WL: Stem cell fate
decision making: Modeling approaches. ACS Biomater Sci Eng.
3:2702–2711. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Wu RX, Yin Y, He XT, Li X and Chen FM:
Engineering a cell home for stem cell homing and accommodation. Adv
Biosyst. 1:e17000042017. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Costanza B, Umelo IA, Bellier J,
Castronovo V and Turtoi A: Stromal modulators of TGF-β in cancer. J
Clin Med. 6:72017. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Janmey PA, Wells RG, Assoian RK and
McCulloch CA: From tissue mechanics to transcription factors.
Differentiation. 86:112–120. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Muehlich S, Rehm M, Ebenau A and
Goppelt-Struebe M: Synergistic induction of CTGF by cytochalasin D
and TGFbeta-1 in primary human renal epithelial cells: Role of
transcriptional regulators MKL1, YAP/TAZ and Smad2/3. Cell Signal.
29:31–40. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Rana MK, Aloisio FM, Choi C and Barber DL:
Formin-dependent TGF-β signaling for epithelial to mesenchymal
transition. Mol Biol Cell. 29:1465–1475. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Ng LF, Kaur P, Bunnag N, Suresh J, Sung
ICH, Tan QH, Gruber J and Tolwinski NS: WNT signaling in disease.
Cells. 8:8262019. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Mezzacappa C, Komiya Y and Habas R:
Activation and function of small GTPases Rho, Rac, and Cdc42 during
gastrulation. Methods Mol Biol. 839:119–131. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Maeda T, Sakabe T, Sunaga A, Sakai K,
Rivera AL, Keene DR, Sasaki T, Stavnezer E, Iannotti J, Schweitzer
R, et al: Conversion of mechanical force into TGF-β-mediated
biochemical signals. Curr Biol. 21:933–941. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Varelas X, Sakuma R, Samavarchi-Tehrani P,
Peerani R, Rao BM, Dembowy J, Yaffe MB, Zandstra PW and Wrana JL:
TAZ controls Smad nucleocytoplasmic shuttling and regulates human
embryonic stem-cell self-renewal. Nat Cell Biol. 10:837–848. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Varelas X, Samavarchi-Tehrani P, Narimatsu
M, Weiss A, Cockburn K, Larsen BG, Rossant J and Wrana JL: The
Crumbs complex couples cell density sensing to Hippo-dependent
control of the TGF-β-SMAD pathway. Dev Cell. 19:831–844. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Narimatsu M, Samavarchi-Tehrani P, Varelas
X and Wrana JL: Distinct polarity cues direct Taz/Yap and TGFβ
receptor localization to differentially control TGFβ-induced Smad
signaling. Dev Cell. 32:652–656. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Alarcon C, Zaromytidou AI, Xi Q, Gao S, Yu
J, Fujisawa S, Barlas A, Miller AN, Manova-Todorova K, Macias MJ,
et al: Nuclear CDKs drive Smad transcriptional activation and
turnover in BMP and TGF-beta pathways. Cell. 139:757–769. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Qin Z, Xia W, Fisher GJ, Voorhees JJ and
Quan T: YAP/TAZ regulates TGF-β/Smad3 signaling by induction of
Smad7 via AP-1 in human skin dermal fibroblasts. Cell Commun
Signal. 16:182018. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Serowoky MA, Arata CE, Crump JG and
Mariani FV: Skeletal stem cells: Insights into maintaining and
regenerating the skeleton. Development. 147:dev1793252020.
View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Tang Y and Weiss SJ: Snail/Slug-YAP/TAZ
complexes cooperatively regulate mesenchymal stem cell function and
bone formation. Cell Cycle. 16:399–405. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Kovar H, Bierbaumer L and Radic-Sarikas B:
The YAP/TAZ pathway in osteogenesis and bone sarcoma pathogenesis.
Cells. 9:9722020. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Fernandez LA, Northcott PA, Dalton J,
Fraga C, Ellison D, Angers S, Taylor MD and Kenney AM: YAP1 is
amplified and up-regulated in hedgehog-associated medulloblastomas
and mediates Sonic hedgehog-driven neural precursor proliferation.
Genes Dev. 23:2729–2741. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Lin YT, Ding JY, Li MY, Yeh TS, Wang TW
and Yu JY: YAP regulates neuronal differentiation through Sonic
hedgehog signaling pathway. Exp Cell Res. 318:1877–1888. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Hayashi S, Tamura K and Yokoyama H: Yap1,
transcription regulator in the Hippo signaling pathway, is required
for Xenopus limb bud regeneration. Dev Biol. 388:57–67. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Hsu TH, Yang CY, Yeh TH, Huang YC, Wang TW
and Yu JY: The Hippo pathway acts downstream of the Hedgehog
signaling to regulate follicle stem cell maintenance in the
Drosophila ovary. Sci Rep. 7:44802017. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Machado MV, Michelotti GA, Pereira TA, Xie
G, Premont R, Cortez-Pinto H and Diehl AM: Accumulation of duct
cells with activated YAP parallels fibrosis progression in
non-alcoholic fatty liver disease. J Hepatol. 63:962–970. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Yimlamai D, Christodoulou C, Galli GG,
Yanger K, Pepe-Mooney B, Gurung B, Shrestha K, Cahan P, Stanger BZ
and Camargo FD: Hippo pathway activity influences liver cell fate.
Cell. 157:1324–1338. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Cotton JL, Li Q, Ma L, Park JS, Wang J, Ou
J, Zhu LJ, Ip YT, Johnson RL and Mao J: YAP/TAZ and hedgehog
coordinate growth and patterning in gastrointestinal mesenchyme.
Dev Cell. 43:35–47.e4. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Heng BC, Zhang X, Aubel D, Bai Y, Li X,
Wei Y, Fussenegger M and Deng X: Role of YAP/TAZ in cell lineage
fate determination and related signaling pathways. Front Cell Dev
Biol. 8:7352020. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Blanpain C and Fuchs E: Epidermal
homeostasis: A balancing act of stem cells in the skin. Nat Rev Mol
Cell Biol. 10:207–217. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Simpson CL, Patel DM and Green KJ:
Deconstructing the skin: Cytoarchitectural determinants of
epidermal morphogenesis. Nat Rev Mol Cell Biol. 12:565–580. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Watt FM, Estrach S and Ambler CA:
Epidermal Notch signalling: Differentiation, cancer and adhesion.
Curr Opin Cell Biol. 20:171–179. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Totaro A, Castellan M, Battilana G,
Zanconato F, Azzolin L, Giulitti S, Cordenonsi M and Piccolo S:
YAP/TAZ link cell mechanics to Notch signalling to control
epidermal stem cell fate. Nat Commun. 8:152062017. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Low BC, Pan CQ, Shivashankar GV,
Bershadsky A, Sudol M and Sheetz M: YAP/TAZ as mechanosensors and
mechanotransducers in regulating organ size and tumor growth. FEBS
Lett. 588:2663–2670. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Piccolo S, Cordenonsi M and Dupont S:
Molecular pathways: YAP and TAZ take center stage in organ growth
and tumorigenesis. Clin Cancer Res. 19:4925–4930. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Hayashi S, Yokoyama H and Tamura K: Roles
of Hippo signaling pathway in size control of organ regeneration.
Dev Growth Differ. 57:341–351. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Ramos A and Camargo FD: The Hippo
signaling pathway and stem cell biology. Trends Cell Biol.
22:339–346. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Mo JS, Park HW and Guan KL: The Hippo
signaling pathway in stem cell biology and cancer. EMBO Rep.
15:642–656. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Hans C: Wnt/beta-catenin signaling in
development and disease. Cell. 127:469–480. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Niehrs C and Acebron SP: Mitotic and
mitogenic Wnt signalling. EMBO J. 31:2705–2713. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Piccolo S, Dupont S and Cordenonsi M: The
biology of YAP/TAZ: Hippo signaling and beyond. Physiol Rev.
94:1287–1312. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Azzolin L, Panciera T, Soligo S, Enzo E,
Bicciato S, Dupont S, Bresolin S, Frasson C, Basso G, Guzzardo V,
et al: YAP/TAZ incorporation in the β-catenin destruction complex
orchestrates the Wnt response. Cell. 158:157–170. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Park HW, Kim YC, Yu B, Moroishi T, Mo JS,
Plouffe SW, Meng Z, Lin KC, Yu FX, Alexander CM, et al: Alternative
Wnt signaling activates YAP/TAZ. Cell. 162:780–794. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Chen X, Yuan W, Li Y, Luo J and Hou N:
Role of Hippo-YAP1/TAZ pathway and its crosstalk in cardiac
biology. Int J Biol Sci. 16:2454–2463. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Sato N, Meijer L, Skaltsounis L, Greengard
P and Brivanlou AH: Maintenance of pluripotency in human and mouse
embryonic stem cells through activation of Wnt signaling by a
pharmacological GSK-3-specific inhibitor. Nat Med. 10:55–63. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Weston CR and Davis RJ: The JNK signal
transduction pathway. Curr Opin Cell Biol. 19:142–149. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Chen F: JNK-induced apoptosis,
compensatory growth, and cancer stem cells. Cancer Res. 72:379–386.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Bogoyevitch MA and Kobe B: Uses for JNK:
The many and varied substrates of the c-Jun N-terminal kinases.
Microbiol Mol Biol Rev. 70:1061–1095. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Kaunas R, Usami S and Chien S: Regulation
of stretch-induced JNK activation by stress fiber orientation. Cell
Signal. 18:1924–1931. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Codelia VA, Sun G and Irvine KD:
Regulation of YAP by mechanical strain through Jnk and Hippo
signaling. Curr Biol. 24:2012–2017. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Wang L, Luo JY, Li B, Tian XY, Chen LJ,
Huang Y, Liu J, Deng D, Lau CW, Wan S, et al: Integrin-YAP/TAZ-JNK
cascade mediates atheroprotective effect of unidirectional shear
flow. Nature. 540:579–582. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Plouffe SW, Hong AW and Guan KL: Disease
implications of the Hippo/YAP pathway. Trends Mol Med. 21:212–222.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Martinez B, Yang Y, Harker DMR, Farrar C,
Mukundan H, Nath P and Mascareñas D: YAP/TAZ related BioMechano
signal transduction and cancer metastasis. Front Cell Dev Biol.
7:1992019. View Article : Google Scholar : PubMed/NCBI
|