Introduction
Atherosclerosis is a primary cause of myocardial
infarction worldwide and atherosclerosis was present in ~60% of the
people >60 years old in European and American countries in 2019
(1,2). Atherosclerosis is associated with
several risk factors, including dysfunction of human umbilical vein
endothelial cells (HUVECs) (3).
Inappropriate proliferation, apoptosis, injury and migration of
HUVECs lead to the progression of atherosclerosis (4,5).
Therefore, the maintenance of the function of HUVECs is important
for the treatment of atherosclerosis.
Circular (circ)RNAs are a class of endogenous RNA
molecules with a stable closed loop structure (6). circRNAs serve important regulatory
roles in cell biological functions, such as protein synthesis, gene
expression and post-transcriptional modification (6). circRNAs have been shown to play a key
role in the function of HUVECs during the progression of
atherosclerosis. For example, Qin et al (7) demonstrated that knockdown of
hsa_circ_0003645 expression alleviates inflammation and apoptosis
in oxidized low-density lipoprotein (oxLDL)-induced HUVECs.
Moreover, Wang et al (8)
indicated that circ_0124644 exacerbates oxLDL-induced injury in
HUVECs via sponging miR-149-5p in atherosclerosis. In addition,
hsa_circ_0001445 expression is notably downregulated in
atherosclerosis (9). However, the
function of hsa_circ_0001445 in HUVECs remains unclear.
Serine/arginine-rich splicing factors (SRSFs) are
well-characterized for their roles in atherosclerosis and are
composed of one or two RNA recognition motifs and a
serine/arginine-rich domain (10).
Among these SRSFs, SRSF1 is a prototypical splicing factor that
specifically binds to exonic enhancers and stimulates splicing
(11). In addition, it has been
reported that SRSF1 binds to hsa_circ_0001445 (12). However, the association between
hsa_circ_0001445 and SRSF1 remains unclear.
Based on this evidence, the present study aimed to
investigate the biological function of hsa_circ_0001445 in
atherosclerosis in vitro.
Materials and methods
Cell culture and treatment
HUVECs (cat. no. CRL1730) were obtained from the
American Type Culture Collection and cultured in RPMI-1640 medium,
supplemented with 10% FBS and 2 mM glutamine (Sigma-Aldrich; Merck
KGaA) at 37°C, 5% CO2. In order to establish an in
vitro model of atherosclerosis, HUVECs were treated with oxLDL
(50, 75 or 100 µg/ml) for 48 h at 37°C.
Cell transfection
The pcDNA3.1 vector (1 µg/µl),
pDNA3.1-hsa_circ_0001445 (1 µg/µl) and pDNA3.1-SRSF1 (1 µg/µl) were
purchased from Guangzhou RiboBio Co., Ltd., and transfected into
HUVECs using Lipofectamine® 2000 (Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions.
After 24 h of transfection, transfected cells were used in
subsequent experiments. The efficiency of hsa_circ_0001445
overexpression was detected by reverse transcription-quantitative
(RT-q)PCR and the efficiency of SRSF1 overexpression was
investigated by RT-qPCR and western blotting.
RT-qPCR
Total RNA was extracted from cell lines with
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). Total RNA was reverse transcribed into cDNA using the
PrimeScript RT reagent kit (Takara Bio, Inc.) according to the
manufacturer's instructions. Then, RT-qPCR was performed using a
SYBR® Premix Ex Taq™ II kit (Takara Bio, Inc.) on a
7900HT system (Applied Biosystems; Thermo Fisher Scientific, Inc.)
using the following thermocycling conditions: 60°C for 1 min, 90°C
for 15 min, followed by 40 cycles of 90°C for 15 sec and 55°C for
60 sec. β-actin was used as the internal control. The primer
sequences used were as follows: hsa_circ_0001445 forward,
5′-GAGAAAAACAAAAGGGAGGCTT-3′ and reverse,
5′-TTGAAGCAAACACATGTGTTGC-3′; SRSF1 forward,
5′-GCGACGGCTATGATTACGATG-3′ and reverse,
5′-ACATACATCACCTGCTTCACGC-3′; and β-actin forward,
5′-GTCCACCGCAAATGCTTCTA-3′ and reverse, 5′-TGCTGTCACCTTCACCGTTC-3′.
The 2−ΔΔCq method (13)
was used to measure the relative expression levels of genes.
Cell Counting Kit (CCK)-8 assay
HUVECs were seeded in 96-well plates
(5×103 per well) overnight. The cells were treated with
100 µg/ml oxLDL, hsa_circ_0001445 OE or oxLDL + hsa_circ_0001445 OE
for 0, 24, 48 and 72 h. A total of 10 µl CCK-8 (Beyotime Institute
of Biotechnology) was added to each well and the cells were
incubated for 2 h at 37°C. Finally, the absorbance of HUVECs was
measured at 450 nm using a microplate reader (Thermo Fisher
Scientific, Inc.).
Oil red O staining of lipid
accumulation in cells
Culture media was removed from HUVECs and cells were
washed twice with PBS. The cells were then fixed with 4%
paraformaldehyde at room temperature for 30 min. Oil Red O working
solution was prepared fresh from 0.5% (w/v) stock solution, which
was diluted with water at a ratio of 6:4 (Oil Red O:water). Cells
were then incubated with Oil Red O for 30 min at room temperature,
washed gently with PBS three times to remove excess non-specific
staining and observed under a light microscope (magnification,
×200; Olympus Corporation).
RNA pull-down assay
RNA pull-down assay was performed using Biotin RNA
Labeling Mix (Roche Diagnostics) to transcribe and label
probe-control (ctrl) or probe-hsa_circ_0001445 in vitro. An
RNA structure buffer (Thermo Fisher Scientific, Inc.) was used to
induce secondary structure formation from the biotin-labeled RNAs.
Streptavidin beads (Thermo Fisher Scientific, Inc.) were washed
three times with 500 µl RNA immunoprecipitation washing buffer
(Thermo Fisher Scientific, Inc.) and added to the biotinylated RNA
samples at 4°C overnight. The mixture was separated by a magnetic
field to obtain streptavidin bead-RNA complexes. Subsequently,
lysates of HUVECs (60 µl) were added to the complexes and incubated
on a rotating platform at room temperature for 1 h. The incubated
mixture was separated with a magnetic field to obtain streptavidin
bead-RNA-protein complexes.
In vitro angiogenesis assay
Angiogenesis assay was conducted using 6-well plates
that were pre-coated with Matrigel (BD Biosciences) at 37°C for 24
h. Subsequently, 1×105 HUVECs were seeded into each
well. Following 24 h cell culture at 37°C, the formation of
capillary-like structures was photographed and the branchpoints
were counted. The tube length was analyzed by the AxioVision Rel
software version 4.8 (Carl Zeiss AG).
Transwell assay
Transwell chambers (8 mm pore; Corning, Inc.) were
used to determine the rate of cell migration. The aforementioned
cell culture medium containing 4×105 cells/200 µl was
added into the upper chamber. Cell culture medium supplemented with
10% FBS was added to the lower chamber of the well. Following
culture at 37°C for 24 h, the migrated cells were fixed with 4%
paraformaldehyde for 20 min at room temperature, followed by
staining with 4% crystal violet solution for 5 min at room
temperature. The migrated cells were counted using a light
microscope (200× magnification; Olympus Corporation).
Western blotting
Total protein was isolated from cell lysates using
RIPA buffer (Shanghai GenePharma Co., Ltd.) and quantified by BCA
protein assay kit (Beyotime Institute of Biotechnology). The
proteins (40 µg per lane) were resolved on 10% SDS-PAGE and
subsequently transferred onto PVDF (Bio-Rad Laboratories, Inc.)
membranes. Following blocking with 5% skimmed milk in TBST (0.05%
Tween-20) at room temperature for 1 h, the membranes were incubated
with primary antibodies (all 1:1,000) at 4°C overnight. The
following day, membranes were incubated with HRP-conjugated
secondary anti-rabbit antibody (cat. no. ab7090; 1:5,000 Abcam) at
room temperature for 1 h. The membranes were scanned using an
Odyssey Imaging system and analyzed with Odyssey v2.0 software
(LI-COR Biosciences). The primary antibodies used in the present
study were as follows: Anti-β-catenin (cat. no. ab223075; Abcam),
anti-SRSF1 (cat. no. 32-4500; Thermo Fisher Scientific, Inc.),
anti-E-cadherin (cat. no. ab40772; Abcam), anti-Bcl-2 (cat. no.
ab182858; Abcam), anti-Bax (cat. no. ab182733; Abcam) and
anti-β-actin (cat. no. ab8226; Abcam). β-actin was used as an
internal control. Enhanced chemiluminescence reagent (Thermo Fisher
Scientific, Inc.) was used to visualize the protein bands. ImageJ
software (version 2.0; National Institutes of Health) was used to
quantify the intensity of the bands.
Measurement of mitochondrial membrane
potential (MMP)
The changes in MMP were assessed via
5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethyl-imidacarbocyanine (JC-1)
staining using a FACScan flow cytometer (BD Biosciences). The cells
were plated into 6-well plates (3×105 cells/well)
overnight, harvested, washed twice with PBS and finally resuspended
in 0.5 ml complete medium containing 2 µM JC-1 at 37°C for 20
min.
Immunofluorescence analysis
HUVECs (5×103) were seeded in 24-well
plates and incubated overnight. The following day, the cells were
treated with oxLDL, hsa_circ_0001445 OE, oxLDL + hsa_circ_0001445
OE or oxLDL + hsa_circ_0001445 OE + SRSF1 OE at 37°C for 72 h,
blocked with 10% goat serum (Invitrogen; Thermo Fisher Scientific,
Inc.) for 30 min at room temperature and finally incubated with
anti-5-ethynyl-2′-deoxyuridine (EdU) antibody (1:1,000; cat. no.
ab222421; Abcam) at 4°C overnight, followed by incubation with
HRP-conjugated goat anti-rabbit IgG (1:5,000; ab7090; Abcam) at
37°C for 1 h. Subsequently, the nuclei were stained with DAPI (5
µg/µl, Beyotime Institute of Biotechnology) for 5 min at room
temperature. Finally, the cells were observed using a fluorescence
microscope (magnification, ×200; Olympus CX23; Olympus
Corporation). The images was analyzed by ImageJ software (version
2.0; National Institutes of Health).
Cell apoptosis analysis
HUVECs were trypsinized, washed with PBS and
resuspended in Annexin V binding buffer. The cells were stained
with 5 µl FITC (BD Biosciences) and 5 µl propidium iodide (BD
Biosciences) at 4°C for 15 min. Finally, they were analyzed using
flow cytometry (BD Biosciences) to assess the cell apoptotic rate.
The data was quantified by FlowJo (v7.6.5; FlowJo LLC).
Statistical analysis
Each experiment was performed in three independent
repeats. The data are presented as the mean ± SD. Comparisons
between two groups were analyzed using unpaired Student's t-test.
Comparisons between multiple groups were performed using one-way
ANOVA followed by post hoc Tukey's test. The analysis was performed
using GraphPad Prism 7 (GraphPad Software, Inc.). P<0.05 was
considered to indicate a statistically significant difference.
Results
oxLDL-induced inhibition of HUVEC
proliferation is reversed by hsa_circ_0001445 OE
In order to mimic atherosclerosis in vitro,
HUVECs were treated with different concentrations of oxLDL (50, 75
or 100 µg/ml). oxLDL significantly inhibited the expression levels
of hsa_circ_0001445 in HUVECs in a dose-dependent manner (Fig. 1A). Based on this evidence, 100 µg/ml
oxLDL was selected for use in subsequent experiments. Expression
levels of hsa_circ_0001445 in HUVECs were notably upregulated by
pcDNA3.1-hsa_circ_0001445 (Fig.
1B). HUVEC viability was significantly decreased by oxLDL; this
effect was reversed in the presence of hsa_circ_0001445 OE
(Fig. 1C). hsa_circ_0001445 OE
reversed the antiproliferative effect of oxLDL on HUVECs (Fig. 1D). Taken together, the results
indicated that oxLDL-induced inhibition of HUVEC proliferation was
reversed by hsa_circ_0001445 OE.
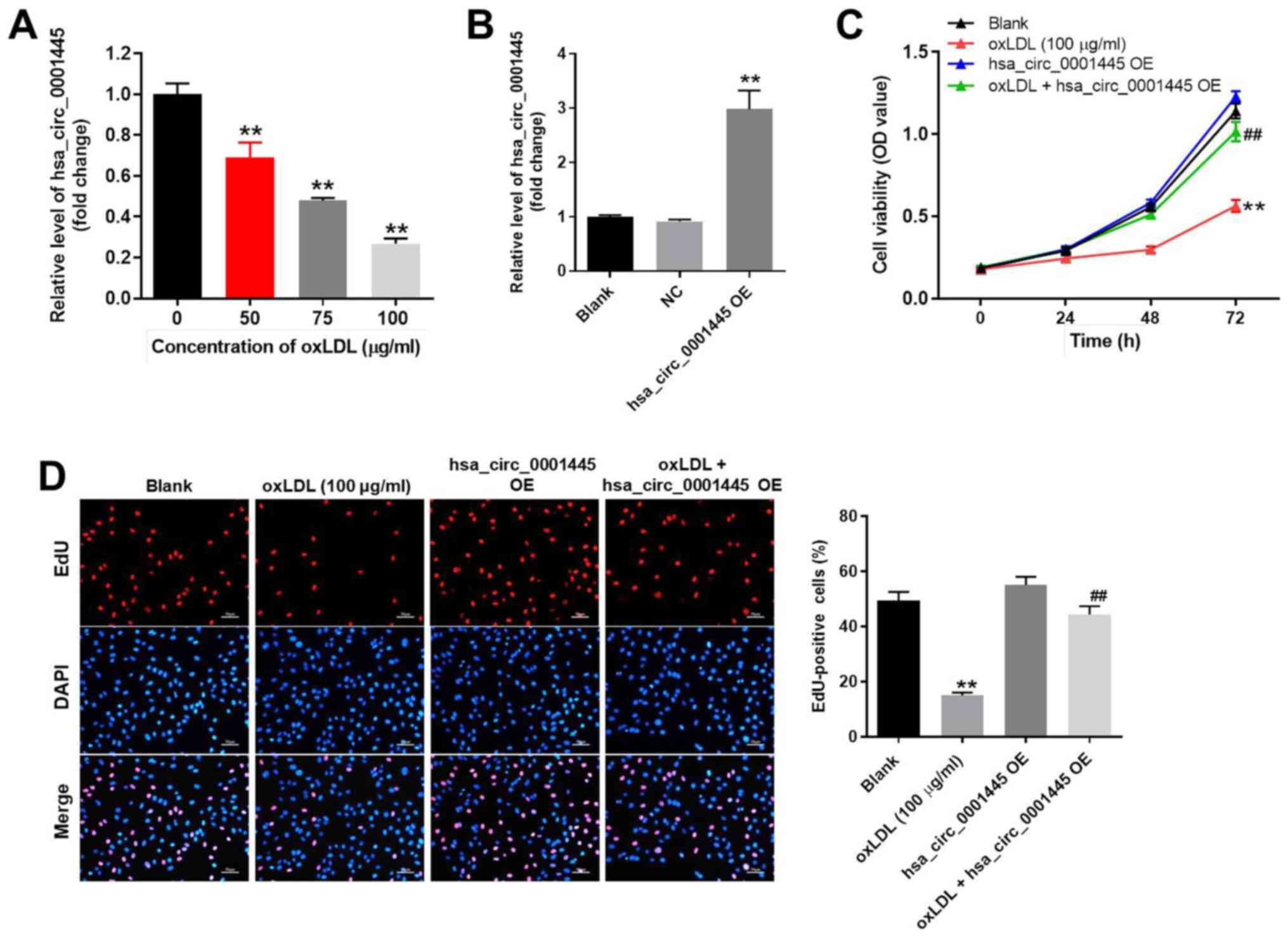 | Figure 1.hsa_circ_0001445 OE promotes
proliferation of oxLDL-treated HUVECs. (A) HUVECs were treated with
0, 50, 75 or 100 µg/ml oxLDL for 48 h. Then, the expression of
hsa_circ_0001445 in HUVECs was detected by RT-qPCR. (B) HUVECs were
transfected with hsa_circ_0001445 OE for 24 h. Then, the expression
of hsa_circ_0001445 in HUVECs was measured by RT-qPCR. (C) HUVECs
were treated with oxLDL (100 µg/ml), hsa_circ_0001445 OE or oxLDL +
hsa_circ_0001445 OE for 0, 24, 48 or 72 h. The viability of HUVECs
was detected by Cell Counting Kit-8 assay. (D) Proliferation of
HUVECs was detected by EdU staining (red). The number of
EdU-positive cells was calculated. Blue immunofluorescence
indicates DAPI. **P<0.01 vs. blank, ##P<0.01 vs.
oxLDL (100 µg/ml). OE, overexpression; oxLDL, oxidized low-density
lipoprotein; HUVECs, human umbilical vein endothelial cells; RT-q,
reverse transcription-quantitative; EdU, 5-ethynyl-2′-deoxyuridine;
OD, optical density; NC, negative control. |
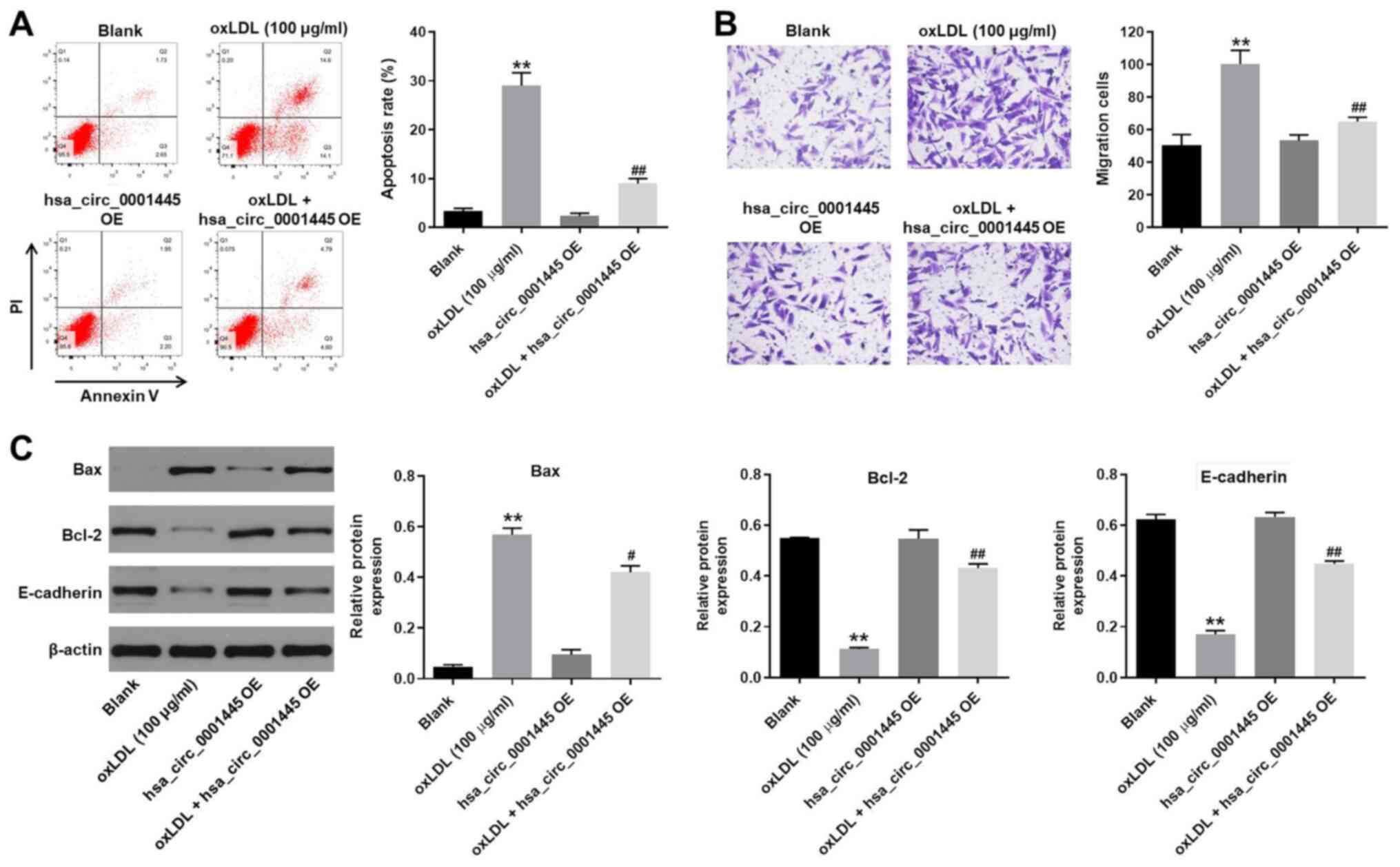
hsa_circ_0001445 OE reverses
oxLDL-induced apoptosis and migration of HUVECs
In order to detect cell apoptosis, flow cytometry
was performed. oxLDL induced apoptosis of HUVECs, while its
apoptotic effect was significantly inhibited by hsa_circ_0001445 OE
(Fig. 2A). oxLDL-induced increase
in cell migration was inhibited by hsa_circ_0001445 OE (Fig. 2B). In addition, the expression
levels of Bax in HUVECs were upregulated by oxLDL, while this
effect was reversed by hsa_circ_0001445 OE (Fig. 2C). By contrast, the oxLDL-induced
decrease in Bcl-2 and E-cadherin expression levels was partially
rescued in the presence of hsa_circ_0001445 OE (Fig. 2C). Taken together, the data
demonstrated that hsa_circ_0001445 OE reversed oxLDL-induced
apoptosis and migration of HUVECs.
hsa_circ_0001445 OE inhibits
progression of angiogenesis in vitro
The role of hsa_circ_0001445 in atherosclerosis was
investigated by angiogenesis assay. The branch points of
capillary-like structures in HUVECs were significantly decreased by
oxLDL treatment, while this effect was improved in the presence of
hsa_circ_0001445 OE (Fig. 3A).
Similarly, oxLDL-induced decrease in tube length of HUVECs was
reversed by hsa_circ_0001445 OE (Fig.
3A). In addition, hsa_circ_0001445 OE relieved oxLDL-induced
lipid accumulation and mitochondrial injury in HUVECs (Fig. 3B and C) and significantly inhibited
the progression of angiogenesis in vitro.
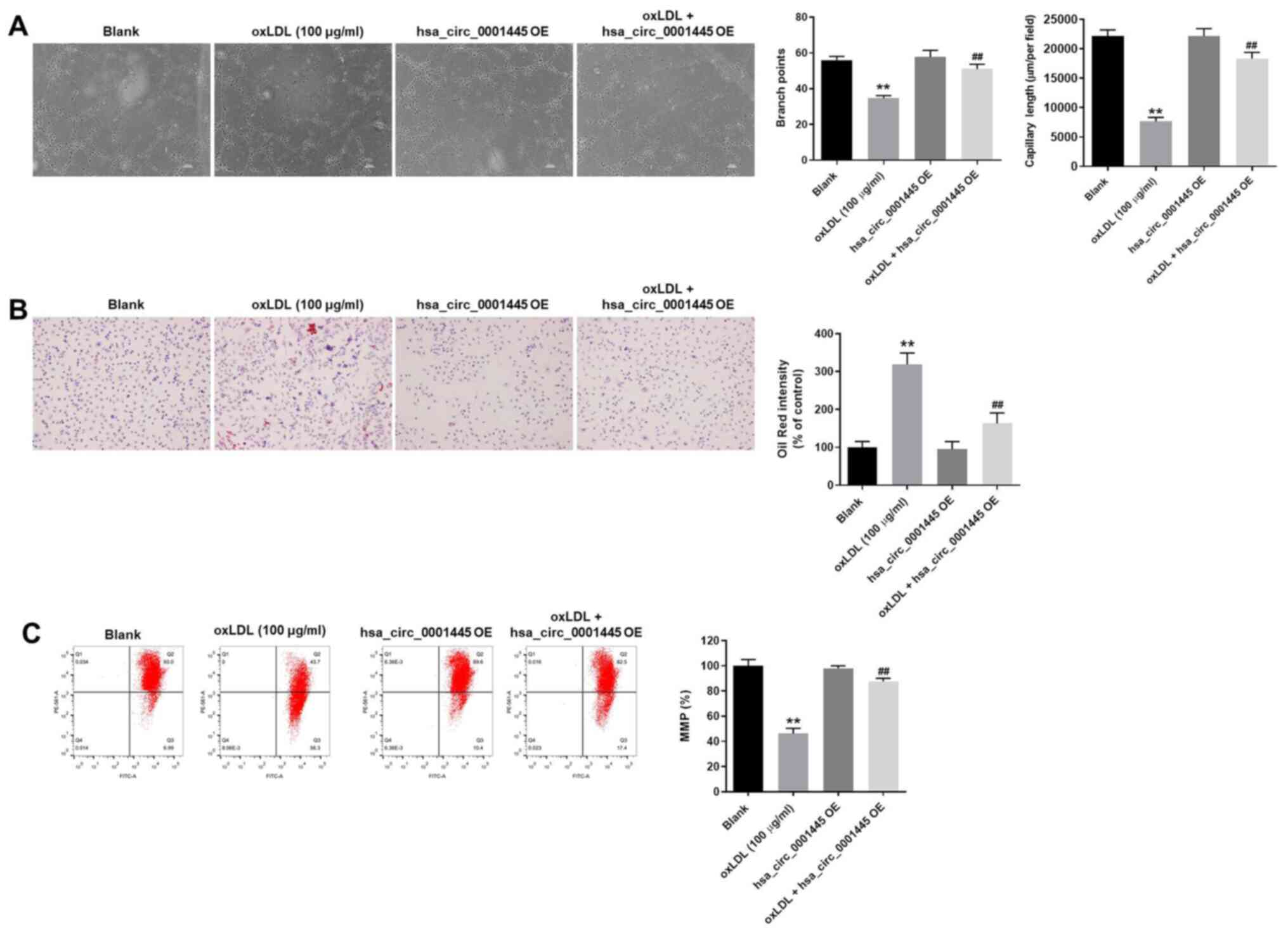 | Figure 3.hsa_circ_0001445 OE inhibits
progression of angiogenesis in vitro. (A) Formation of
capillary-like structures was photographed in HUVECs. The branch
points were calculated. The capillary length was tested. (B) Lipid
accumulation in HUVECs was tested by Oil Red O staining. (C)
Changes in MMP were measured by
5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethyl-imidacarbocyanine
staining using a FACSan flow cytometer. The ratio of MMP was
calculated. **P<0.01 vs. blank, ##P<0.01 vs. oxLDL
(100 µg/ml). OE, overexpression; HUVECs, human umbilical vein
endothelial cells; MMP, mitochondrial membrane potential. |
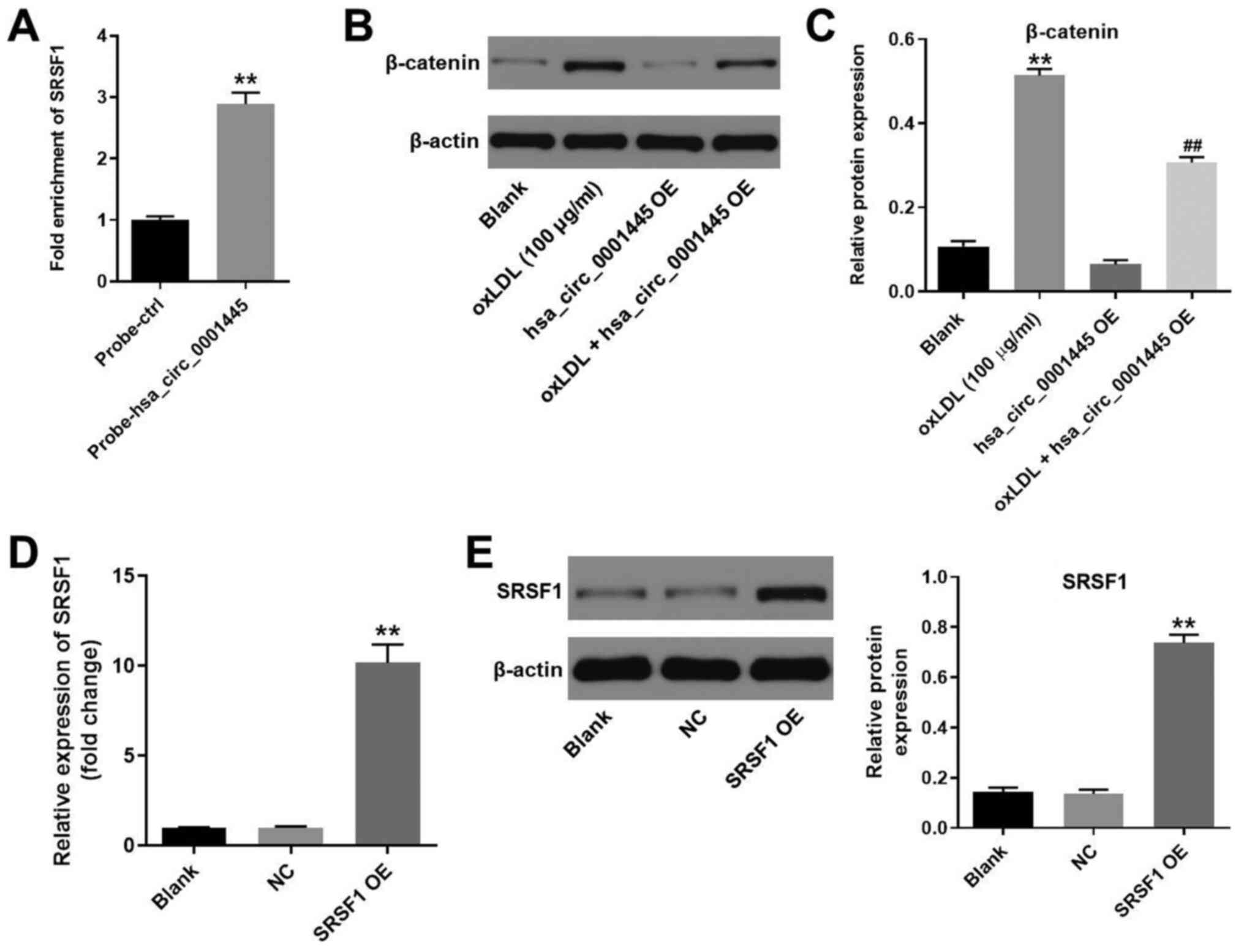
hsa_circ_0001445 binds to SRSF1 and
reverses oxLDL-induced activation of β-catenin
It has been shown that hsa_circ_0001445 binds to
SRSF1 (12,14). Therefore, the present study
investigated the association between hsa_circ_0001445 expression
levels and SRSF1 in HUVECs by RNA pull-down assay. As expected, the
fold enrichment of SRSF1 in HUVECs was increased in
probe-hsa_circ_0001445 compared with that in probe-ctrl cells
(Fig. 4A). Moreover, the protein
levels of β-catenin in HUVECs were upregulated by oxLDL (Fig. 4B and C). However, the effects of
oxLDL on β-catenin expression were partially inhibited in the
presence of hsa_circ_0001445 OE (Fig.
4B and C). HUVECs were transfected with pcDNA3.1-SRSF1, and the
efficiency of cell transfection was tested. Expression of SRSF1 in
HUVECs was significantly upregulated by pcDNA3.1-SRSF1 (Fig. 4D and E). In summary, the data
demonstrated that hsa_circ_0001445 reversed oxLDL-induced
activation of β-catenin via binding to SRSF1.
hsa_circ_0001445 OE reverses
oxLDL-induced HUVEC proliferation inhibition by binding to
SRSF1
In order to assess the association between
hsa_circ_0001445 and SRSF1, EdU staining and flow cytometry were
performed. hsa_circ_0001445 OE reversed the antiproliferative
effect of oxLDL on HUVECs, while SRSF1 OE reversed this effect
(Fig. 5A). Moreover, SRSF1 OE
partially rescued the antiapoptotic effects of hsa_circ_0001445 on
oxLDL-treated HUVECs (Fig. 5B),
whereas oxLDL-induced activation of β-catenin was significantly
inhibited in the presence of hsa_circ_0001445 OE. hsa_circ_0001445
OE exerted limited effects on the expression of SRSF1 in
oxLDL-treated HUVECs (Fig. 5C). By
contrast, protein levels of E-cadherin in HUVECs were inhibited in
the presence of oxLDL, while this effect was significantly reversed
by hsa_circ_0001445 OE (Fig. 5C).
Furthermore, the effects of hsa_circ_0001445 on SRSF1, E-cadherin
and β-catenin were significantly inhibited by SRSF1 OE (Fig. 5C). Taken together, the results
indicated that hsa_circ_0001445 OE reversed oxLDL-induced
inhibition of HUVEC proliferation via binding to SRSF1.
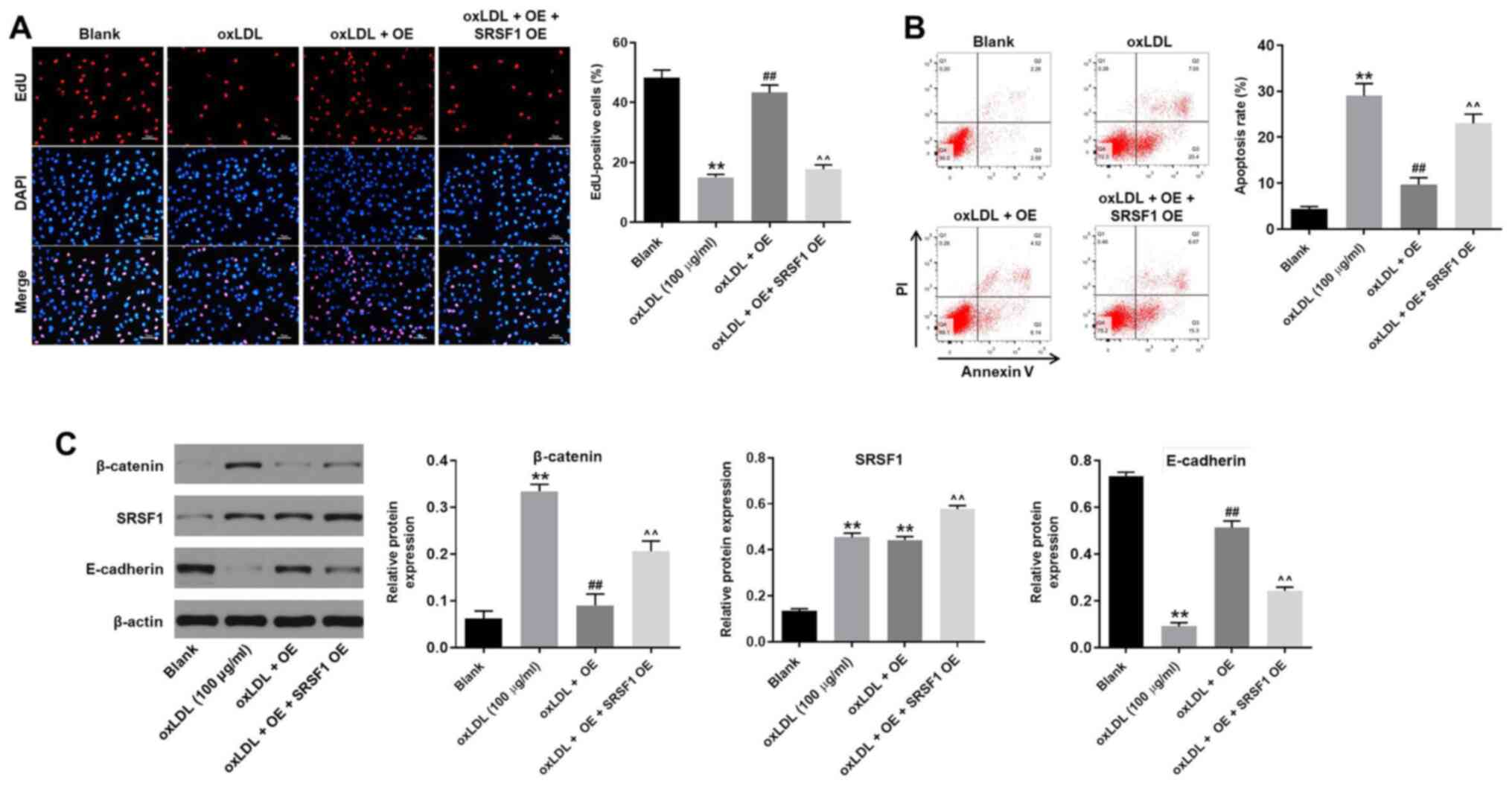 | Figure 5.hsa_circ_0001445 OE reverses
oxLDL-induced inhibition of HUVEC growth via binding to SRSF1. (A)
HUVECs were treated with oxLDL (100 µg/ml), hsa_circ_0001445 OE,
oxLDL + hsa_circ_0001445 OE or oxLDL + hsa_circ_0001445 OE + SRSF1
OE. Then, the proliferation of HUVECs was tested by EdU staining.
(B) Apoptosis of HUVECs was tested by flow cytometry. (C) Protein
expression levels of β-catenin, SRSF1 and E-cadherin in HUVECs were
detected by western blot analysis. The relative expression levels
were normalized to β-actin. **P<0.01 vs. blank,
##P<0.01 vs. oxLDL (100 µg/ml),
^^P<0.01 vs. oxLDL + OE. OE, overexpression; oxLDL,
oxidized low-density lipoprotein; HUVEC, human umbilical vein
endothelial cells; SRSF1, serine/arginine-rich splicing factor 1;
EdU, 5-ethynyl-2′-deoxyuridine; E-cadherin, epithelial
cadherin. |
Discussion
Atherosclerosis is a complex multicellular process
involving vascular injury and atheroma formation, which is
associated with vascular inflammation, leading to increased
expression levels of pro-inflammatory cytokines, pentraxin-3,
monocyte chemotactic protein, vascular cell adhesion molecule-1,
intercellular adhesion molecule-1 and E-selectin in monocytes and
endothelial cells (15,16). During the progression of
atherosclerosis, oxLDL induces lipid accumulation in macrophages,
thus leading to the formation of foam cells (17). In the present study, it was
demonstrated that hsa_circ_0001445 expression was significantly
decreased in oxLDL-treated HUVECs. It has been previously shown
that hsa_circ_0001445 participates in the development of certain
diseases. For example, hsa_circ_0001445 serves as a key biomarker
for osteoporosis in postmenopausal women (18) and can modulate VEGFA mRNA splicing
and angiogenesis in glioblastoma multiforme by binding to SRSF1
(12). The present study
investigated the biological function of hsa_circ_0001445 in
atherosclerosis, revealing that hsa_circ_0001445 may serve as an
inhibitor during the progression of this disease.
The present study investigated the mechanism by
which hsa_circ_0001445 OE mediates the progression of
atherosclerosis. In this study, SRSF1 could bind to
hsa_circ_0001445. SRSF1 is involved in several cellular processes.
For instance, LncRNA GASAL1 can interact with SRSF1 to regulate
trophoblast cell proliferation, invasion, and apoptosis via the
mTOR signaling (19); Sun and Hu
(20) found that SRSF1 upregulation
could induce the apoptosis of cardiomyocytes. It has been reported
that SRSF1 is a risk factor involved in coronary disease, including
atherosclerosis (21). In addition,
previous studies have shown that hsa_circ_0001445 binds to SRSF1 in
glioblastoma multiforme (12,14).
The present study demonstrated the association between
hsa_circ_0001445 and SRSF1 in HUVECs, confirming that
hsa_circ_0001445 reversed oxLDL-induced inhibition of HUVEC
proliferation by binding to SRRSF1.
The association between SRSF1 and β-catenin has been
previously reported (22,23). SRSF1 recruit β-catenin and promote
its expression (23,24). In the present study,
hsa_circ_0001445 OE inhibited the expression of β-catenin in
oxLDL-treated HUVECs. Therefore, the present study revealed the
association between SRSF1 and β-catenin in oxLDL-treated HUVECs,
suggesting that hsa_circ_0001445 OE decreased expression levels of
β-catenin via binding to SRSF1. Moreover, Wnt/β-catenin signaling
has been reported to serve an important role in the development of
coronary artery disease, including atherosclerosis (25,26).
Wu et al (27) demonstrated
that inactivation of the Wnt/β-catenin signaling pathway reverses
oxLDL-induced inhibition of HUVEC proliferation. Our research
indicates that hsa_circ_0001445 reverses oxLDL-induced inhibition
of HUVEC proliferation via inactivation of β-catenin. It has also
been shown that activation of Wnt/β-catenin reverses oxLDL-induced
foam cell formation in macrophages during the development of
atherosclerosis (28). Therefore,
more studies are required to investigate the association between
β-catenin and atherosclerosis. E-cadherin is a crucial mediator in
the epithelial-mesenchymal transition (EMT) process (29,30)
and it has been shown that β-catenin negatively regulates
expression of E-cadherin, thereby promoting EMT (31,32).
The findings of the present study indicated that E-cadherin
expression in oxLDL-HUVECs was upregulated by hsa_circ_0001445 OE.
In summary, the data suggested that hsa_circ_0001445 OE reversed
oxLDL-induced inhibition of HUVEC proliferation via activation of
the SRSF1/β-catenin/EMT axis.
There were certain limitations to the present study.
The sponging of specific miRNAs by hsa_circ_0001445 in HUVECs was
not investigated. Only in vitro data were presented and the
analysis lacked in vivo results. The effect of oxLDL +
hsa_circ_0001445 OE + SRSF1 OE on angiogenesis and migration of
cells needs to be further investigated. Further in vivo
assays using multiple cell types are also required to validate the
results of the present study. Therefore, additional investigations
are needed in future.
Taken together, the data from the present study
demonstrated that hsa_circ_0001445 OE reversed oxLDL-induced
inhibition of HUVEC proliferation via activation of the
SRSF1/β-catenin/EMT axis. The findings may provide information for
the identification of potential treatment options for
atherosclerosis.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LD conceived and supervised the study. GL designed
the study. GL, SC and SX performed the experiments and analyzed the
data. LD and GL confirmed the authenticity of all the raw data. All
authors reviewed the results and read and approved the final
version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Liang H, Chen M, Qi F, Shi L, Duan Z, Yang
R, He J, Lou B, Li Y and Yang Q: The proatherosclerotic function of
indoleamine 2, 3-dioxygenase 1 in the developmental stage of
atherosclerosis. Signal Transduct Target Ther. 4:232019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yang W, Yin R, Zhu X, Yang S, Wang J, Zhou
Z, Pan X and Ma A: Mesenchymal stem-cell-derived exosomal miR-145
inhibits atherosclerosis by targeting JAM-A. Mol Ther Nucleic
Acids. 23:119–131. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Luo X, Fu H, Xu C, Dong Y, Wu Z, Li D, Sun
Y, Shen M, Wang L, Li Z and Duan Y: Efficient treatment of
atherosclerosis by dexamethasone acetate and rapamycin Co-Loaded
mPEG-DSPE calcium phosphate nanoparticles. J Biomed Nanotechnol.
16:810–826. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yin J, Hou X and Yang S: MicroRNA-338-3p
promotes ox-LDL-induced endothelial cell injury through targeting
BAMBI and activating TGF-β/Smad pathway. J Cell Physiol.
234:11577–11586. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dang H, Song B, Dong R and Zhang H:
Atorvastatin reverses the dysfunction of human umbilical vein
endothelial cells induced by angiotensin II. Exp Ther Med.
16:5286–5297. 2018.PubMed/NCBI
|
|
6
|
Han BH, Song CH, Yoon JJ, Kim HY, Seo CS,
Kang DG, Lee YJ and Lee HS: Anti-vascular inflammatory effect of
ethanol extract from securinega suffruticosa in human umbilical
vein endothelial cells. Nutrients. 12:34482020. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Qin M, Wang W, Zhou H, Wang X, Wang F and
Wang H: Circular RNA circ_0003645 silencing alleviates inflammation
and apoptosis via the NF-κB pathway in endothelial cells induced by
oxLDL. Gene. 755:1449002020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang G, Li Y, Liu Z, Ma X, Li M, Lu Q, Li
Y, Lu Z, Niu L, Fan Z and Lei Z: Circular RNA circ_0124644
exacerbates the ox-LDL-induced endothelial injury in human vascular
endothelial cells through regulating PAPP-A by acting as a sponge
of miR-149-5p. Mol Cell Biochem. 471:51–61. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Vilades D, Martinez-Camblor P,
Ferrero-Gregori A, Bar C, Lu D, Xiao K, Vea A, Nasarre L, Sanchez
Vega J, Leta R, et al: Plasma circular RNA hsa_circ_0001445 and
coronary artery disease: Performance as a biomarker. FASEB J.
34:4403–4414. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Long JC and Caceres JF: The SR protein
family of splicing factors: Master regulators of gene expression.
Biochem J. 417:15–27. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Graveley BR and Maniatis T:
Arginine/serine-rich domains of SR proteins can function as
activators of pre-mRNA splicing. Mol Cell. 1:765–771. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Barbagallo D, Caponnetto A, Brex D,
Mirabella F, Barbagallo C, Lauretta G, Morrone A, Certo F, Broggi
G, Caltabiano R, et al: CircSMARCA5 regulates VEGFA mRNA splicing
and angiogenesis in glioblastoma multiforme through the binding of
SRSF1. Cancers (Basel). 11:1942019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Barbagallo D, Caponnetto A, Cirnigliaro M,
Brex D, Barbagallo C, D'Angeli F, Morrone A, Caltabiano R,
Barbagallo GM, Ragusa M, et al: CircSMARCA5 inhibits migration of
glioblastoma multiforme cells by regulating a molecular axis
involving splicing factors SRSF1/SRSF3/PTB. Int J Mol Sci.
19:4802018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Huang JW, Jiang X, Li ZL and Jiang CR:
MicroRNA-328-5p alleviates macrophage lipid accumulation through
the histone deacetylase 3/ATP-binding cassette transporter A1
pathway. Lipids. Mar 4–2021.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Christophersen DV, Moller P, Thomsen MB,
Lykkesfeldt J, Loft S, Wallin H, Vogel U and Jacobsen NR:
Accelerated atherosclerosis caused by serum amyloid A response in
lungs of ApoE(−/-) mice. FASEB J. 35:e213072021. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dubland JA: Role of inflammatory cytokines
in genesis and treatment of atherosclerosis. Looking at foam cells
through a different lens. Trends Cardiovasc Med. Mar 3–2021.(Epub
ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xiang S, Wu Y, Shi H, Xue L, Luo K and
Ding Y: Circular RNA hsa_circ_0001445 in plasma as a novel
biomarker for osteoporosis in postmenopausal women. Biomark Med.
14:1599–1607. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu J, Zhang Q and Ma N: LncRNA GASAL1
interacts with SRSF1 to regulate trophoblast cell proliferation,
invasion, and apoptosis via the mTOR signaling pathway. Cell
Transplant. 29:9636897209651822020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sun Y and Hu ZQ: LncRNA HOTAIR aggravates
myocardial ischemia-reperfusion injury by sponging microRNA-126 to
upregulate SRSF1. Eur Rev Med Pharmacol Sci. 24:9046–9054.
2020.PubMed/NCBI
|
|
21
|
Tejedor JR, Tilgner H, Iannone C, Guigo R
and Valcarcel J: Role of six single nucleotide polymorphisms, risk
factors in coronary disease, in OLR1 alternative splicing. RNA.
21:1187–1202. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hu M, Wang R, Li X, Fan M, Lin J, Zhen J,
Chen L and Lv Z: LncRNA MALAT1 is dysregulated in diabetic
nephropathy and involved in high glucose-induced podocyte injury
via its interplay with beta-catenin. J Cell Mol Med. 21:2732–2747.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fu Y, Huang B, Shi Z, Han J, Wang Y,
Huangfu J and Wu W: SRSF1 and SRSF9 RNA binding proteins promote
Wnt signalling-mediated tumorigenesis by enhancing beta-catenin
biosynthesis. EMBO Mol Med. 5:737–750. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Thorsen K, Mansilla F, Schepeler T, Øster
B, Rasmussen MH, Dyrskjøt L, Karni R, Akerman M, Krainer AR,
Laurberg S, et al: Alternative splicing of SLC39A14 in colorectal
cancer is regulated by the Wnt pathway. Mol Cell Proteomics.
10:M110.002998. 2011. View Article : Google Scholar
|
|
25
|
Liu Y, Neogi A and Mani A: The role of Wnt
signalling in development of coronary artery disease and its risk
factors. Open Biol. 10:2001282020. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Singla B, Lin HP, Chen A, Ahn W, Ghoshal
P, Cherian-Shaw M, White J, Stansfield BK and Csanyi G: Role of
R-spondin 2 in arterial lymphangiogenesis and atherosclerosis.
Cardiovasc Res. Aug 4–2020.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wu H, Liu T and Hou H: Knockdown of
LINC00657 inhibits ox-LDL-induced endothelial cell injury by
regulating miR-30c-5p/Wnt7b/β-catenin. Mol Cell Biochem.
472:145–155. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu Y, Wei M, Liu G, Song C, Yang M, Cao Z
and Zheng M: Silencing protease-activated Receptor-2 alleviates
ox-LDL-induced lipid accumulation, inflammation and apoptosis via
activation of Wnt/β-catenin signaling. Gen Physiol Biophys.
39:437–448. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hu C, Li M, Guo T, Wang S, Huang W, Yang
K, Liao Z, Wang J, Zhang F and Wang H: Anti-metastasis activity of
curcumin against breast cancer via the inhibition of stem cell-like
properties and EMT. Phytomedicine. 58:1527402019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhao L, Li J, Liu Y, Zhou W, Shan Y, Fan
X, Zhou X, Shan B, Song Y and Zhan Q: Flotillin1 promotes EMT of
human small cell lung cancer via TGF-β signaling pathway. Cancer
Biol Med. 15:400–414. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang W and He B: MiR-760 inhibits the
progression of non-small cell lung cancer through blocking
ROS1/Ras/Raf/MEK/ERK pathway. Biosci Rep. Apr 29–2020.(Epub ahead
of print).
|
|
32
|
Yang D, Li Q, Shang R, Yao L, Wu L, Zhang
M, Zhang L, Xu M, Lu Z, Zhou J, et al: WNT4 secreted by tumor
tissues promotes tumor progression in colorectal cancer by
activation of the Wnt/β-catenin signalling pathway. J Exp Clin
Cancer Res. 39:2512020. View Article : Google Scholar : PubMed/NCBI
|