Introduction
Hepatic ischemia-reperfusion (IR) injury is a common
complication that occurs due to a variety of factors, such as liver
transplantation, shock and trauma (1). Temporary blood flow deprivation
(ischemia) and restoration (reperfusion) of the organs are the
primary pathological processes occurring in IR (2). Liver parenchymal cell death is caused
by ischemic injury, which involves metabolic disorders and
oxidative stress. Moreover, inflammatory mediators cause further
damage during blood reperfusion (3). In addition, hepatic IR injury affects
the quality of donor livers and the prognosis of liver
transplantation (4).
The transforming growth factor-β (TGF-β) superfamily
exerts multiple biological functions via the secretion of inhibins,
activins and bone morphogenetic proteins (BMPs), which are involved
in regulating a range of biological processes, for example BMPs can
induce endochondral bone formation (5–7). TGF-β
is an important activated mediator of myofibroblasts, which
activates stellate cells to secrete collagen fibers, in turn
leading to liver fibrosis (8,9). The
abnormal expression of TGF-β not only promotes the proliferation
and migration of liver cancer cells, but it is also associated with
viral hepatitis, hepatic failure and other chronic hepatic diseases
(10–14). As the downstream effector of TGF-β,
SMAD family member 3 (Smad3) activation is induced by its
phosphorylation to phosphorylated-Smad3 (p-Smad3) and the signal is
transported to the nucleus, thus forming the classic TGF-β/Smad3
signal transduction pathway (15).
The Smad3 linker region can be phosphorylated by intracellular
kinases and affects TGF-β responses, such as tumor growth
inhibition (16).
A previous study reported that the TGF-β/Smad3
signaling pathway was activated and promoted ventricular remodeling
after IR injury in rats (17),
while mediating the protection of myocardial cells against IR
injury via the stimulation of sphingosine-1-phosphate (S1P)/S1P
receptor 1. TGF-β1/Smad3 has also been reported to exert an
important effect on cerebral ischemia. For example, Smad3 has been
shown to exhibit neuroprotective effects on the brain following IR
via the induction of anti-inflammatory and anti-apoptotic pathways
(18). Thus, it was hypothesized
that the TGF-β1/Smad3 signaling pathway may be able to protect
liver cells from IR injury. Therefore, the present study
investigated the role of TGF-β/Smad3 in achieving clinical
transformation in IR-induced acute liver injury and provide a novel
therapeutic target for the prognosis of liver surgery.
Materials and methods
Animals
In total, 20 129S2/SvPasCrl wild-type (WT) mice,
aged 6–8 weeks and weighing 22–25 g, were purchased from the
Laboratory Animal Center of Zhejiang University School of Medicine
(Hangzhou, China) and were randomized into sham-operated (SH-WT)
and IR injury (IR-WT) groups (n=10/group). A total of four
Smad3+/− heterozygous 129S2/SvPasCrl mice were donated
by The Second Military Medical University (Shanghai, China). Mating
of Smad3+/− mice was followed by sibling mating of
offspring-generated Smad3+/− mice, to obtain
Smad3+/+ mice and Smad3−/− mice for
experiments. The mouse genotypes were identified at 3 weeks of age,
and DNA was isolated from the toes of mice by heating to 100°C with
50 mM NaOH for 10 min. Then, 2X Phanta Max Master Mix (cat. no.
P515-01; Vazyme Biotech Co., Ltd.) was used for PCR amplification.
The amplification conditions were set as follows: Initial
denaturation at 95°C for 3 min, followed by 35 cycles at 95°C for
15 sec, 59°C for 15 sec and 72°C for 30 sec. Then, 3%
Tris-acetate-EDTA buffer agarose gel electrophoresis was used to
identify the Smad3 gene-deficient mouse genotype, and visualized
under the ChemiDoc™ System (cat. no. 1708265; Bio-Rad Laboratories,
Inc.). The primers used for genotyping are listed in Table I. The overall morphology of Smad3
mice was assessed and homozygous Smad3−/− mutant (MUT)
mice were found to be smaller than littermate Smad3+/−
and Smad3+/+ mice. Then, the mice were divided into four
groups: Smad3+/+ mice that underwent a sham operation
(SH-WT; n=10); Smad3+/+ mice with IR liver injury
(IR-WT; n=10); Smad3−/− mice that underwent a sham
operation (SH-MUT; n=5); and Smad3−/− mice with IR liver
injury (IR-MUT; n=5).
 | Table I.Primers used for genotyping and
reverse transcription-quantitative PCR. |
Table I.
Primers used for genotyping and
reverse transcription-quantitative PCR.
| Target gene | Forward primer
(5′→3′) | Reverse primer
(5′→3′) |
|---|
| Smad3 WT/MUT |
CCACTTCATTGCCATATGCCCTG |
|
| Smad3 WT |
CCCGAACAGTTGGATTCACACA |
|
| Smad3 MUT |
CCAGACTGCCTTGGGAAAAGC |
|
| β-actin |
GGCTGTATTCCCCTCCATCG |
CCAGTTGGTAACAATGCCATGT |
| TLR4 |
ATGGCATGGCTTACACCACC |
GAGGCCAATTTTGTCTCCACA |
| IFN-γ |
AGGTCAACAACCCACAGGTC |
ATCAGCAGCGACTCCTTTTC |
| TNF |
TAGCTCCCAGAAAAGCAAGC |
TTTTCTGGAGGGAGATGTGG |
| B220 |
GTTTTCGCTACATGACTGCACA |
AGGTTGTCCAACTGACATCTTTC |
| CD3E |
ATGCGGTGGAACACTTTCTGG |
GCACGTCAACTCTACACTGGT |
| Ly-6G |
CGCCCCACTACTCTGGACAATAC |
AAACCAGGCTGAACAGAAGCACCC |
All mice were maintained on a 12/12-h light/dark
cycle under controlled humidity (50±10%) and temperature (25±0.5°C)
in a specific pathogen-free environment and allowed free access to
standard chow and water in the Experimental Animal Center of Sir
Run Run Shaw Hospital, Zhejiang University School of Medicine. All
animal experiments included in this protocol adhere to the Animal
Research: Reporting In Vivo Experiments guidelines (19), and were approved by the Animal
Testing Ethics Committee of Sir Run Run Shaw Hospital, Zhejiang
University School of Medicine (approval no. 20171120-14).
Model of IR liver injury
The present study used a non-lethal segmental (70%)
liver IR model as previously described (20). In brief, a vertical incision was
made to each layer to expose the liver after the mouse was
anesthetized (4% chloral hydrate sodium; 400 mg/kg) by
intraperitoneal injection. The blood vessels, except those of the
caudate and right lobes, were occluded with the vascular clamp for
30 min to block the blood flow without injuring the remaining liver
tissues. Subsequently, the vascular clamp was gently removed and
the blood flow and liver tissues were assessed without injuring
before the wound was sutured. In the sham group, surgery was
performed by exposing the blood vessels only for 30 min without
blocking. Subsequently, recipient animals were intraperitoneally
injected with anesthesia (4% chloral hydrate sodium; 400 mg/kg) at
the end of the predetermined period (after 6 h of reperfusion), and
then the blood samples were collected from the orbital venous
plexus to assess serum alanine aminotransferase (ALT) levels. Mice
were then sacrificed by cervical dislocation, and the death of mice
was confirmed when the respiration and various reflexes had ceased.
A portion of the IR injury tissue was used for extraction and
detection of total protein and RNA. Then, ~1.0×1.0×1.0-cm liver
tissues were promptly fixed overnight in 4% paraformaldehyde (cat.
no. 6148; Sigma-Aldrich; Merck KGaA) at room temperature.
Histopathological evaluation
The hepatic tissues were fixed in 4%
paraformaldehyde for 8 h at room temperature, dehydrated, embedded
in paraffin, sectioned to 3-µm thickness, stained with hematoxylin
for 3 min, rinsed under running water for 5 min, stained with eosin
for 2 min and sealed at room temperature. Stained tissues were
viewed and imaged using a light microscope. The Suzuki histological
grading scores for liver damage were determined as described
previously (21). A score of 0
indicated minimal or no evidence of damage, 1 indicated mild damage
with cytoplasmic vacuolation and interstitial disorder, 2 indicated
moderate to severe damage with extensive nuclear pyknosis and
interstitial congestion, and 3 indicated serious necrosis with
disintegration of liver cells, hemorrhage and inflammatory cell
infiltration.
Detection of ALT levels
Blood samples were centrifuged at 4°C for 5 min at
1,500 × g after standing for 1 h at room temperature and the serum
was separated. Serum levels of ALT were determined using an ALT
Assay kit (cat. no. C009-2-1; Nanjing Jiancheng Biological
Technology), according to the manufacturer's instructions.
Reverse transcription-quantitative PCR
(RT-qPCR)
The fresh hepatic tissue was homogenized using an
automatic sample quick grinding machine (model JXFSTPRP-24;
Shanghai Jingxin Industrial Development Co., Ltd.). Total RNA was
extracted using TRIzol® reagent (cat. no. 10296-028;
Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. cDNA was synthesized from RNA using
the Hifair® II 1st Strand cDNA Synthesis SuperMix for RT
(cat. no. 11121ES60; Shanghai Yeasen Biotechnology Co., Ltd.). The
temperature conditions reverse transcription were as follows: 42°C
for 15 min and 85°C for 2 min. qPCR was performed using the
FastStart Universal SYBR Green Master (ROX) mix (cat. no.
04913914001; Roche Diagnostics), according to the manufacturer's
instructions. The amplification conditions were set as: Initial
denaturation at 95°C for 10 min, followed by 40 cycles at 95°C for
15 sec, 60°C for 1 min and 60°C for 30 sec. TsingKe Biological
Technology synthesized the primers for PCR (Table I). The 2−ΔΔCq method was
used to analyze the data (22).
Western blotting
RIPA lysis buffer (Beyotime Institute of
Biotechnology), supplemented with protease inhibitor cocktail (cat.
no. HY-K0011; MedChemExpress) and phosphatase inhibitor cocktail
(cat. no. HY-K0022; MedChemExpress), was used to extract total
proteins. Liver tissues were homogenized in protein lysis buffer on
ice and the protein concentrations were measured using a BCA
protein assay kit (cat. no. CW0014; CoWin Biosciences). The
proteins (20 µg) were loaded onto each lane and separated via 15%
SDS-PAGE, and then transferred onto a PVDF membrane (cat. no.
IPVH00010; MilliporeSigma). Then, 5% skimmed milk (BD Biosciences)
was used to block the membrane for 1 h using a rocking shaker at
room temperature; then, the membranes were incubated for 8 h, at a
minimum temperature of 4°C, with primary antibodies against Smad3
(1:1,000; cat. no. ab208182; Abcam), p-Smad3 (1:500; cat. no.
bs-3425R; BIOSS), TGF-β1 (1:1,000; cat. no. ab215715; Abcam),
β-actin (1:1,000; cat. no. 20536-1-AP; ProteinTech Group, Inc.) and
cleaved caspase-3 (1:2,000; cat. no. ab214430; Abcam). The
following day, the membrane was thoroughly rinsed three times in
western blot washing buffer (cat. no. CW0043S; CoWin Biosciences),
followed by incubation with an appropriate HRP-conjugated secondary
antibody (1:4,000; cat. no. FDR007; Fdbio Science) for 1 h at room
temperature. Finally, ECL reagents (cat. no. FD8030; Fdbio Science)
were used to assess the antigen-antibody complex on the membrane.
The bands were detected using the ChemiDoc™ Touch Imager (Bio-Rad
Laboratories, Inc.) and analysis was performed using ImageLab
software version 5.2 (Bio-Rad Laboratories, Inc.). The ratio of
phosphorylated protein/total protein was evaluated by ImageJ
version 1.0 software (National Institutes of Health).
Immunohistochemistry
The hepatic tissues were fixed in 4%
paraformaldehyde for 8 h at room temperature, fixed tissue was
processed using an autoprocessor machine (cat. no. ASP200S; Leica
Microsystems GmbH), and then the tissues were cut into 3-µm
sections and the antigen retrieval process was performed in sodium
citrate solution (cat. no. C1010; Beijing Solarbio Science &
Technology Co., Ltd.) in a high-pressure steam boiler for 10 min,
followed by incubation with 3% H2O2 for 15
min at room temperature and blocking with 10% goat serum (cat. no.
G9023; Sigma-Aldrich; Merck KGaA) in PBS for 1 h. Slides were
incubated with primary antibodies against CD45 (1:250; cat. no.
14-0454-85; eBioscience; Thermo Fisher Scientific, Inc.), Ki67
(1:10,000; cat. no. ab15580; Abcam) or F4/80 (1:250; cat. no.
4339486; Invitrogen; Thermo Fisher Scientific, Inc.) at 4°C
overnight. Next, the slides were restored to normal temperature and
washed with PBS three times and then processed using the GTvision
immunohistochemical kit (cat. no. GK600710; Shanghai, Gene Tech
Company Ltd.) according to the manufacturer's instructions. Slides
were incubated with anti-mouse/rabbit secondary antibodies,
included in the aforementioned kit, at room temperature for 30 min
from the working solution B of the kit, the DAB solution was
prepared according to the kit instructions and the reaction time
was controlled under the light microscope. After counterstaining
with hematoxylin (same procedure as aforementioned), it was sealed
with neutral resin (cat. no. MB0722; Dalian Meilun Biology
Technology Co., Ltd.). Stained tissues were viewed and imaged under
a light microscope.
TUNEL staining
The hepatic tissues were fixed in 4%
paraformaldehyde for 8 h at room temperature, fixed tissue was
processed using an autoprocessor machine, and then the tissues were
cut into 3-µm sections. Hepatocyte apoptosis in paraffin-embedded
sections was determined with the TUNEL kit (cat. no. KGA703;
Nanjing KeyGen Biotech Co., Ltd.) according to the manufacturer's
instructions, as previously described (20). After staining the nuclei with
hematoxylin (same procedure as aforementioned), it was sealed with
neutral resin. Stained tissues were viewed and imaged using a light
microscope (23).
Statistical analysis
GraphPad Prism 8.00 software (GraphPad Software,
Inc.) was used for all statistical analyses. Data are presented as
the mean ± SEM of a minimum of three independent experiments.
Differences among multiple groups were analyzed using one-way ANOVA
followed by Tukey's post hoc test. Ordinal data were analyzed using
non-parametric tests, Mann-Whitney U or Kruskal-Wallis followed by
Dunn's post hoc test. Data were analyzed using SPSS version 17.0
(SPSS, Inc.). P<0.05 was considered to indicate a statistically
significant difference.
Results
TGF-β1/Smad3 signaling is activated
during hepatic IR injury in 129S2/SvPasCrl WT mice
Firstly, the hepatic IR injury model was established
in mice. Subsequently, serum ALT levels were determined and H&E
staining was performed. As presented in Fig. 1A, the hepatic cells were arranged in
neat rows and the lobular structure in the SH-WT group was still
clear; however, the swelling and mild vacuolation in the IR-WT
group contrasted with that seen in the SH-WT group. Moreover, the
Suzuki histological grading value was significantly increased
following liver IR injury in the IR-WT group compared with that in
the SH-WT group (Fig. 1B). It was
revealed that the levels of ALT were significantly increased in the
hepatic IR injury group compared with those in the SH-WT group
(Fig. 1C). These results indicated
hepatocellular damage occurred and confirmed that the mouse model
of liver IR injury had been successfully established.
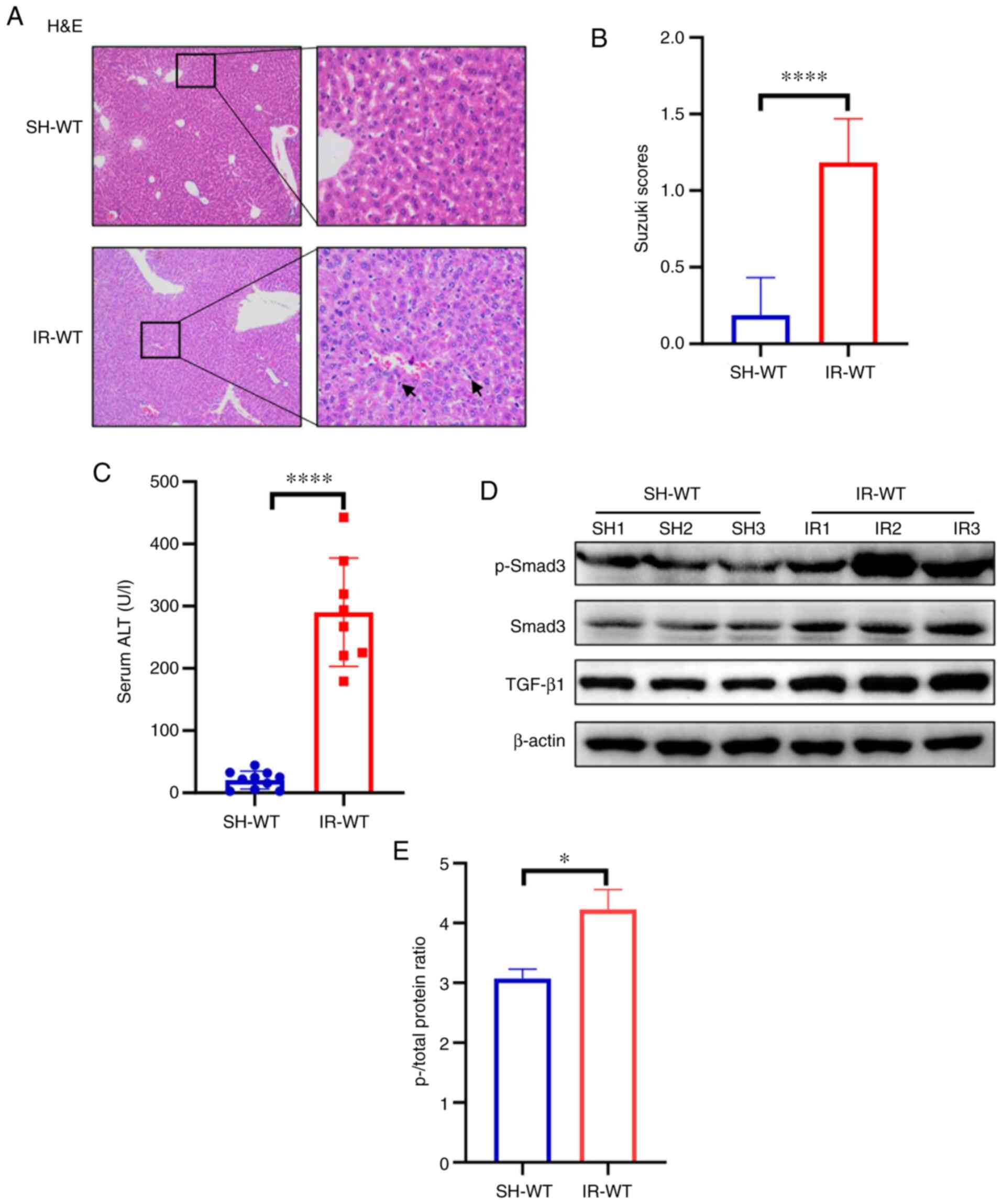 | Figure 1.TGF-β1/Smad3 signaling is activated
during hepatic IR injury in 129S2/SvPasCrl WT mice. (A) Liver
pathological changes were analyzed via histopathological evaluation
in hepatic IR injury tissue (magnification, ×100 and ×400). The
black arrows indicate the positive inflammatory cells. (B) Suzuki
injury score from H&E staining. (C) Levels of the transaminase
ALT were detected in serum samples from the IR-WT and SH-WT groups.
(D) Protein expression levels of TGF-β1, Smad3 and p-Smad3 were
determined in hepatic tissues by western blotting. SH1, SH2 and SH3
indicate three randomly selected samples from the SH-WT group. IR1,
IR2 and IR3 indicate three randomly selected samples from the IR-WT
group. (E) ImageJ software was used to perform grayscale analysis,
to evaluate the ratio of p- vs. total protein. β-actin was used as
the internal reference. *P<0.05, ****P<0.0001. IR-WT, IR
injury in 129S2/SvPasCrl WT mice (n=10); SH-WT, sham operation in
129S2/SvPasCrl WT mice (n=10). ALT, alanine aminotransferase; IR,
ischemia-reperfusion; p-, phosphorylated; Smad3, SMAD family member
3; TGF-β1, transforming growth factor-β1; WT, wild-type. |
Furthermore, the expression levels of TGF-β1, Smad3
and p-Smad3 were increased in the hepatic tissue homogenates of
mice in the IR-WT group compared with those in the SH-WT group
(Fig. 1D). ImageJ software was used
to perform grayscale analysis; the ratio of p- and total protein
indicated that p-Smad3 protein expression was significantly
increased in the IR-WT group compared with the SH-WT group
(Fig. 1E). These results suggested
that the TGF-β/Smad3 signaling pathway was activated during liver
IR injury in mice.
Aggravated hepatic IR injury in Smad3
gene-deficient mice
To determine the effect of the TGF-β/Smad3 signaling
pathway on the induction of hepatic IR damage in mice, Smad3
gene-deficient mice were prepared in order to block the
TGF-β1/Smad3 signaling pathway. The Smad3 gene was expressed during
embryonic development (24), Smad3
expression was detected in adult Smad3−/− mice liver
tissues (data not shown). There was no obvious phenotypic
difference between the Smad3+/+ and Smad3+/−
heterozygous littermates, whereas 70% of the Smad3−/−
mice were smaller prior to weaning (Fig. 2A). The PCR analysis revealed a small
fragment of 284 bp in Smad3−/− mice, and a large
fragment of 431 bp in the Smad3+/+ mice, which confirmed
the genotype of the mice (Fig.
2B).
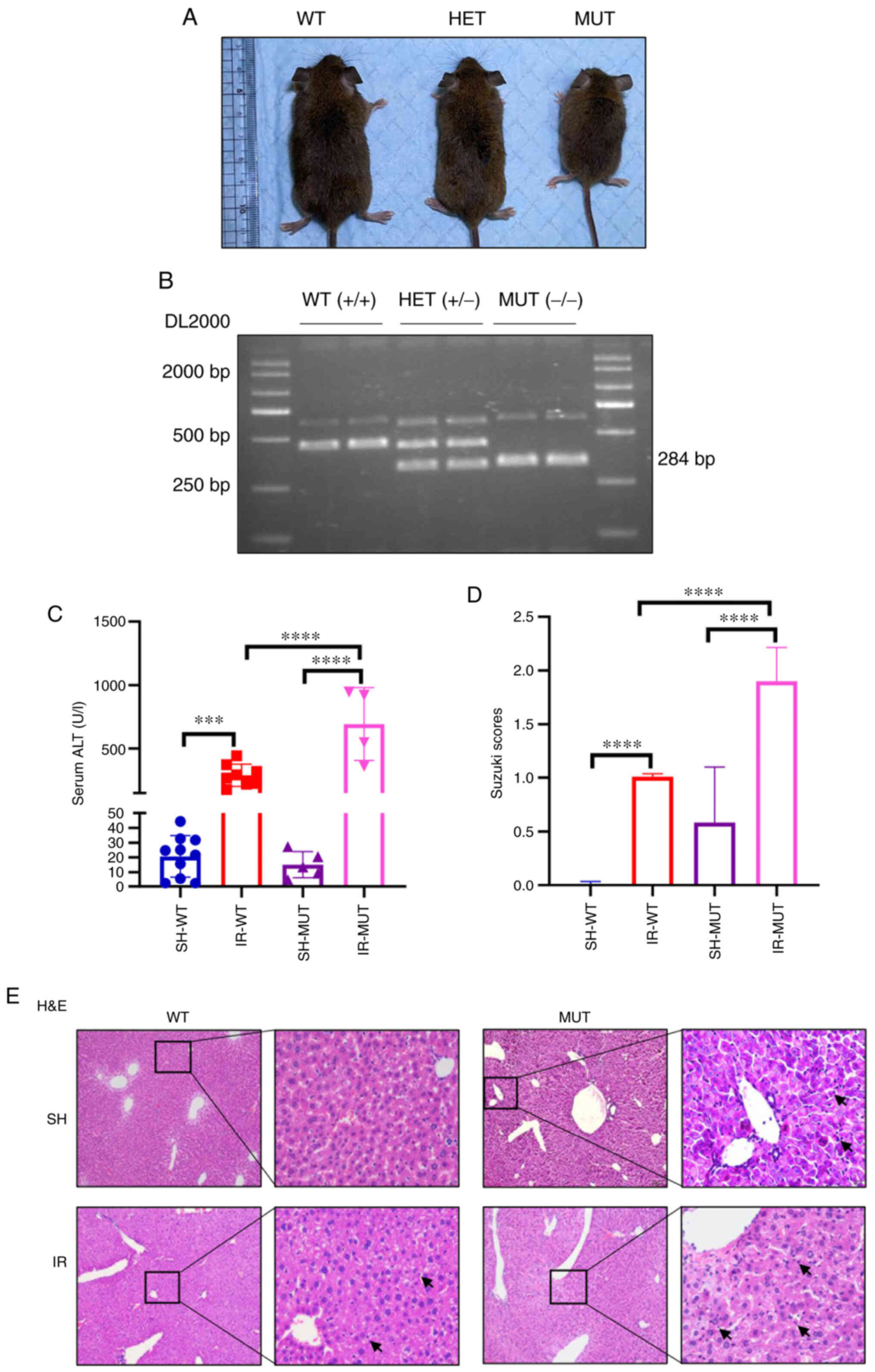 | Figure 2.Aggravated hepatic IR injury in
Smad3−/− mice. (A) Smad3−/− MUT mice appeared
smaller than littermate WT mice or HET mice. (B) Genotyping was
performed via agarose gel electrophoresis. The 431 bp band is the
WT transcript and the 284 bp band is the MUT transcript. (C) Levels
of serum ALT were detected. (D) Suzuki injury score from H&E
staining. (E) Liver pathological changes were analyzed via
histopathological evaluation in hepatic tissue (magnification, ×100
and ×400). The black arrows indicate the positive inflammatory
cells. ***P<0.001, ****P<0.0001. IR-WT, IR injury in Smad3 WT
mice (n=10); SH-WT, sham operation in Smad3 WT mice (n=10); IR-MUT,
IR injury in Smad3 MUT mice (n=5); SH-MUT, sham operation in Smad3
MUT mice (n=5). ALT, alanine aminotransferase; HET, heterozygote;
IR, ischemia-reperfusion; MUT, mutant; Smad3, SMAD family member 3;
WT, wild-type. |
To further identify which component served an
important role in the mouse liver following IR injury, the current
study investigated whether TGF-β/Smad3 possessed a protective
effect. The results indicated that the levels of serum ALT were
increased in the IR-MUT group compared with those in the SH-MUT,
SH-WT and IR-WT groups (Fig. 2C).
The pathological changes after liver injury in the IR-MUT group
were more notable than in the IR-WT group, these changes included
swelling, mild vacuolation and hepatic sinus hyperemia (Fig. 2E). In addition, the Suzuki
histological grading score was significantly higher in the IR-MUT
group compared with that in the IR-WT group (Fig. 2D).
Apoptosis is increased in Smad3
gene-deficient mice following liver IR injury
To further examine the hepatic cell apoptosis and
proliferation in Smad3−/− mice following liver IR
injury, TUNEL and Ki67 staining assays were conducted. The
expression levels of cleaved caspase-3 were detected via western
blotting, and caspase-3 expression levels were measured using
immunohistochemistry. Elevated expression of cleaved caspase-3 was
detected in the IR-MUT group compared with IR-WT group (Fig. 3A). Furthermore, increased staining
of caspase-3-positive hepatocytes was found in the hepatic lobule
portal area of the IR-MUT group compared with that in the IR-WT
group (Fig. 3B). It was also
identified that the staining of TUNEL-positive hepatocytes was
increased in the IR-MUT mice compared with that in the IR-WT group
(Fig. 3C). Although a high level of
proliferation was observed in both the IR groups in comparison with
that in the sham-operated mice, no differences were observed
between the IR-WT and IR-MUT groups based on Ki67 staining results
(Fig. 3D). These findings
demonstrated that Smad3 gene deficiency may aggravate liver IR
damage by promoting hepatocyte apoptosis in mice.
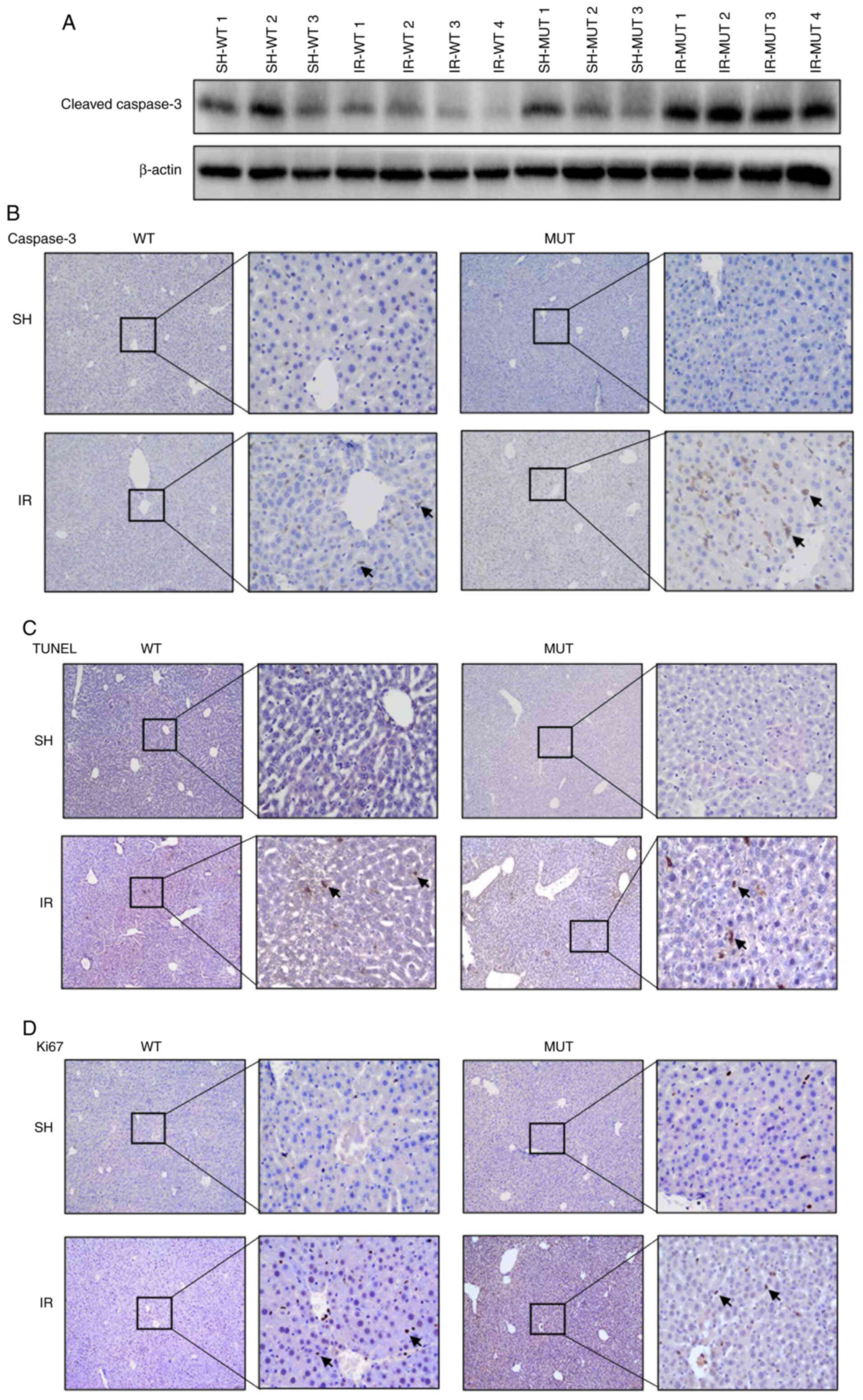 | Figure 3.Apoptosis is increased in
Smad3−/− mice following hepatic IR injury. (A) Protein
expression levels of cleaved caspase-3 were determined via western
blot analysis. SH-WT1, SH-WT2 and SH-WT3 indicate three randomly
selected samples from the SH-WT group. IR-WT1, IR-WT2, IR-WT3 and
IR-WT4 indicate three randomly selected samples from the IR-WT
group. SH-MUT1, SH-MUT2 and SH-MUT3 indicate three randomly
selected samples from the SH-WT group. IR-MUT1, IR-MUT2, IR-MUT3
and IR-MUT4 indicate three randomly selected samples from the IR-WT
group. (B) Caspase-3 staining analysis in hepatic tissue from WT
and Smad3 MUT mice. The positive cells are colored brown (black
arrows; magnification, ×100 and ×400). (C) TUNEL staining indicated
liver cell apoptosis in WT and Smad3 MUT mice. The positive cells
are colored brown (black arrows; magnification, ×100 and ×400). (D)
Ki67 staining analysis in hepatic tissue of WT and Smad3 MUT mice.
The positive cells are colored brown (black arrows; magnification,
×100 and ×400). IR-WT, IR injury in Smad3 WT mice (n=10); SH-WT,
sham-operated in Smad3 WT mice (n=10); IR-MUT, IR injury in Smad3
MUT mice (n=5); SH-MUT, sham-operated in Smad3 MUT mice (n=5). IR,
ischemia-reperfusion; MUT, mutant; Smad3, SMAD family member 3; WT,
wild-type. |
Inflammatory cell infiltration in
Smad3 gene-deficient mice following liver IR injury
IR not only causes damage to liver parenchymal
cells, but also causes the infiltration of inflammatory cells and
the secretion of inflammatory factors (25). To further examine the possible
mechanism via which Smad3 knockout could aggravate liver injury,
inflammatory cells, including inflammatory neutrophils [lymphocyte
antigen 6 complex locus G6D (Ly-6G)], leukocyte infiltration (CD3)
and macrophage cells (F4/80) were analyzed. The results revealed
increased staining of CD45- (Fig.
4A) and F4/80-positive (Fig.
4B) cells in the hepatic tissues of the IR-MUT group compared
with that in the IR-WT group.
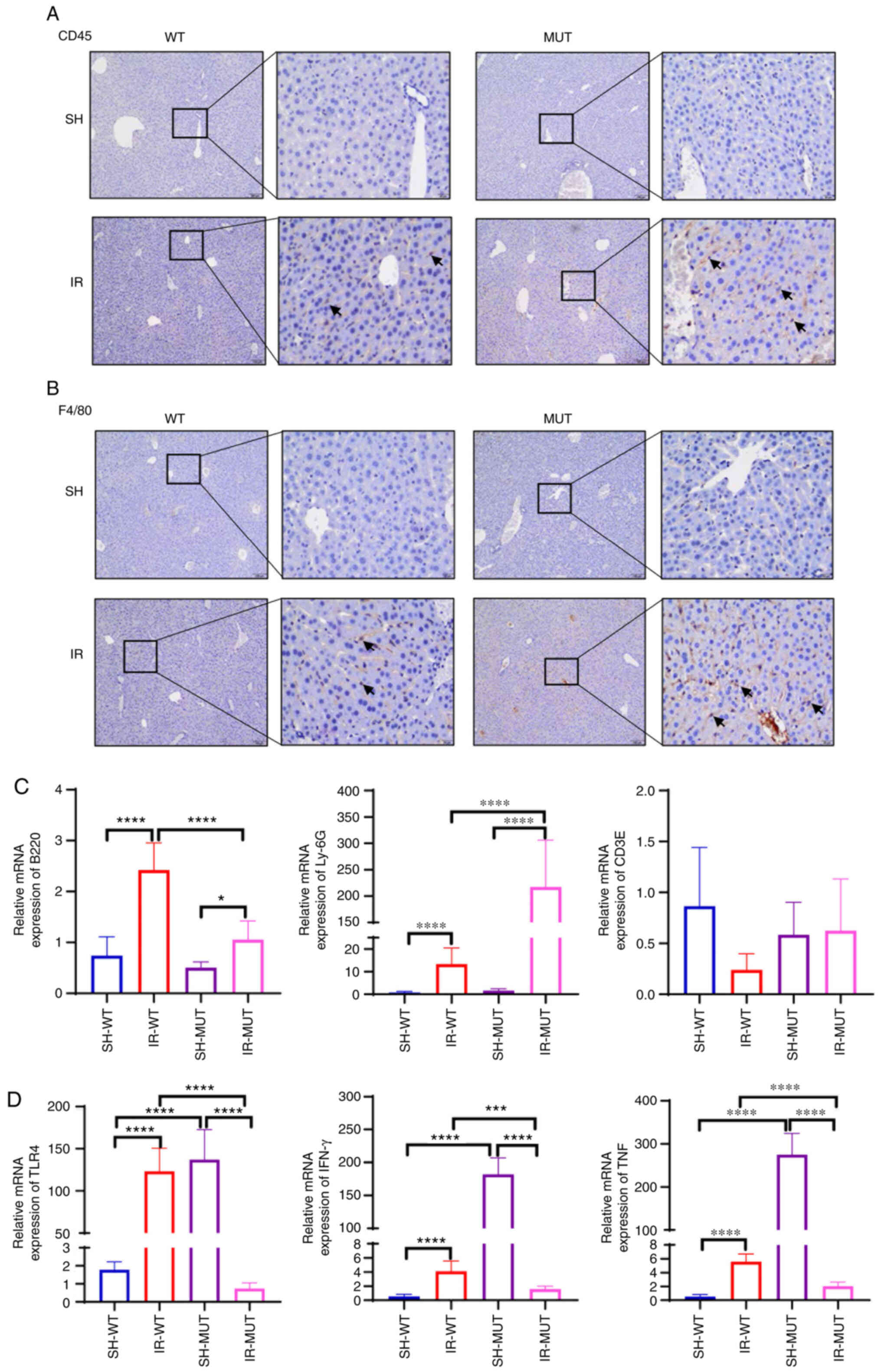 | Figure 4.Inflammatory cell infiltration in
Smad3−/− mice following liver IR injury. (A) CD45
staining analysis in hepatic tissue from WT and Smad3 MUT mice. The
positive cells are colored brown (black arrows; magnification, ×100
and ×400). (B) F4/80 staining analysis in hepatic tissue from WT
and Smad3 MUT mice. The positive cells are colored brown (black
arrows; magnification, ×100 and ×400). (C) B220, Ly-6G and CD3E
mRNA expression levels were assessed in hepatic tissue homogenates
via reverse transcription-quantitative PCR. The results of the
relative mRNA expression levels of represent at least triplicate
determinations. (D) mRNA expression levels of TLR4, IFN-γ and TNF
were assessed in the liver tissues. The graph represents the
relative mRNA expression levels from triplicate determinations.
*P<0.05, ***P<0.001, ****P<0.0001. IR-WT, IR injury in
Smad3 WT mice (n=10); SH-WT, sham-operated in Smad3 WT mice (n=10);
IR-MUT, IR injury in Smad3 MUT mice (n=5); SH-MUT, sham-operated in
Smad3 MUT mice (n=5). IR, ischemia-reperfusion; Ly-6G, lymphocyte
antigen 6 complex locus G6D; MUT, mutant; Smad3, SMAD family member
3; TLR, Toll-like receptor; WT, wild-type. |
To further verify the inflammatory cell infiltration
in Smad3−/− mice following liver IR injury, the mRNA
expression levels of B220, CD3E and Ly-6G were assessed. The
results indicated that the mRNA expression levels of Ly-6G were
significantly increased, whereas B220 mRNA expression levels were
decreased in the hepatic tissue homogenates of the IR-MUT group
compared with those in the IR-WT group; and there was no
significant difference in CD3E mRNA expression (Fig. 4C). Additionally, it was observed
that Toll-like receptor (TLR)4, TNF and IFN-γ mRNA expression
levels were significantly increased in the IR-WT group compared
with SH-WT groups; and TLR4, TNF and IFN-γ mRNA expression levels
were significantly increased in the SH-MUT group compared with
SH-WT groups; however, the opposite result was observed in
Smad3−/− mice, in which the expression levels of TLR4,
IFN-γ and TNF mRNA in the IR-MUT group were decreased compared with
the SH-MUT group (Fig. 4D). These
results suggested that the TGF-β/Smad3 signaling pathway may
directly or indirectly regulate the TLR4 signaling pathway. These
findings demonstrated that inflammatory cell infiltration was
significantly aggravated in Smad3 gene-deficient mice following
liver IR injury.
Discussion
IR injury in the liver is one of the most severe
side effects of liver surgery and transplantation, and is also the
main factor affecting the quality of the transplanted liver
(24). Tissue IR injury can occur
during organ harvesting and peri-transplantation. It has been shown
that microcirculatory dysfunction and immune adjustment are
associated with the pathogenesis of liver IR damage (26,27).
Previous studies have reported that neutrophil inflammatory cells
were detected following IR injury in the liver (28–30).
In conditions such as ischemia, hypoxia and IR liver injury, a
large number of oxygen free radicals can be produced, which induce
the oxidative stress response and liver damage (31). The present study demonstrated that
the TGF-β/Smad3 signaling pathway was activated after liver IR
injury in a mouse model of liver IR and revealed that TGF-β/Smad3
signaling pathway activation may serve a role in protection of
hepatic cells, as the attenuation of this pathway caused aggravated
hepatic cell injury.
TGF-β1 is one of three isoforms of the TGF-β
superfamily (32). TGF-β/Smad3 has
been revealed to be associated with IR injury in several organs.
For example, the interaction of Wnt/β-catenin and TGF-β/Smad
signaling pathways was shown to exert neuroprotective effect in
rats with cerebral IR injury (33).
Moreover, in a previous study, microRNA-211 suppressed apoptosis
and relieved kidney injury following IR by targeting the
TGF-β/Smad3 signaling pathway (34). It has also been shown that TGF-β1
may contribute to isoflurane post-conditioning against cerebral IR
injury by inhibiting the JNK signaling pathway (35). In the present study, it was
demonstrated that TGF-β1 was more highly expressed in hepatic
tissue derived from mice with liver IR injury compared with that in
the sham-operated group.
Considering that TGF-β is the strongest fibrotic
factor and pro-inflammatory factor, which can also aggravate IR
injury (36), the present study
aimed to reduce the effect of IR injury by blocking TGF-β/Smad
signaling. As homozygous TGF-β1 (37,38)
and Smad2 mutations in mice were all embryonic lethal (39,40),
the current study selected Smad3 gene-deficient mice (41,42) as
a model for further examination. This strain of Smad3 MUT mice was
reported to have deficient TGF-β signaling, and studies using the
TGF-β responsive reporter 3TP-Lux failed to show any activation by
the MUT construct (24,42), which indicated that TGF-β signaling
was attenuated in these mice. However, the present results
indicated that IR injury was more severe in Smad3 gene-deficient
mice, with increased hepatocyte apoptosis and higher inflammatory
cell infiltration. The results showed that endogenous deletion of
the TGF-β/Smad3 signaling pathway could aggravate IR injury, which
indirectly demonstrated that the TGF-β/Smad3 signaling pathway may
have a protective role against hepatic IR injury in liver tissue.
In order to further demonstrate the role of this pathway, it would
be useful to interfere with the TGF-β/Smad3 signaling pathway using
TGF-β neutralizing antibodies or inhibitors in future validation
experiments.
TLR4 is an intermediary agent of inflammation and
tissue damage in different IR damage models, such as hepatic
(43), renal (44) and pulmonary (45) models. The activated TLR4 signaling
pathway can promote an increase in the secretion of TNF-α, IFN-β
and other inflammatory cytokines, thus causing increased blood
reperfusion injury (46). A
previous study observed increased TLR4 mRNA expression in
Smad3−/− mice, and found that TLR4 was associated with
lipopolysaccharide (LPS) hyperresponsiveness, leading to the
increased expression of inflammatory cytokines (47). Therefore, the changes in the
expression of TLR4 and other inflammatory cytokines, such as TNF
and IFN-γ, in Smad3+/+ mice after hepatic IR injury in
the present study supported the aforementioned conclusions. In
addition, in the sham operated group, the expression levels of
TLR4, TNF and IFN-γ mRNA were significantly increased in
Smad3−/− mice compared with in the Smad3+/+.
These results indicated that the TGF-β/Smad3 signaling pathway acts
as an immunosuppressive factor that has a direct or indirect
negative regulatory effect on the TLR4 signaling pathway, and loss
of the TGF-β/Smad3 signaling pathway can promote TLR4-mediated
inflammatory injury. Significant increases in the expression of
TLR4, TNF and IFN-γ inflammatory cytokine genes were observed in
Smad3−/− mice, indicating that endogenous deletion of
the Smad3 gene in mice can lead to hyperreactivity of LPS in
vivo, which can increase the secretion of inflammatory
cytokines and hyperendotoxemia. In addition, mRNA expression levels
of TLR4, TNF and IFN-γ were significantly downregulated in
Smad3−/− mice after IR injury. It was suggested that
Smad3−/− mice were resistant to IR injury and the
associated endotoxic shock, resulting in an immune non-response,
which may be associated with the loss of the TGF-β/Smad3 signaling
pathway and persistently high LPS responses. Although the
underlying mechanism between TGF-β/Smad3 signaling and TLR4
signaling is unclear, it is clear that the loss of the negative
regulatory effects of TGF-β/Smad3, through either environmental or
endogenous stimuli, can trigger the activation of TLR4 and
downstream elements, and lead to an imbalance in the number of
inflammatory cells.
In conclusion, the present study demonstrated that
the TGF-β/Smad3 signaling pathway could protect against IR
injury-induced damage in liver tissue repair and immune response.
However, whether exogenous intervention treatment can reduce IR
injury in the early stages of disease still needs further
clarification, which provides a novel research direction for the
prevention of IR injury.
Acknowledgements
Not applicable.
Funding
This work was supported by the National Natural
Science Foundation of China (grant nos. 81702854 and 81700551), and
the Natural Science Foundation of Guangxi Zhuang Autonomous Region
(grant no. 2018GXNBSFBA138033).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
HL and XS conceived and performed the experiments,
analyzed the data, prepared the figures, authored or reviewed
drafts of the paper, and approved the final draft. YTo, TJ and XS
conceived and directed the experiments, and reviewed and approved
the final draft. YTo, TJ and XS confirm the authenticity of all the
raw data. XL designed and guided the project research, and examined
and approved the final manuscripts. YF and YTa conducted the
experiments, reviewed and modified drafts of the paper, and
approved the final draft. JY, TQ and RM performed the animal
experiments, and approved the final draft. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments included in this protocol
adhere to the Animal Research: Reporting In Vivo Experiments
guidelines, and have been approved by the Animal Testing Ethics
Committee of Sir Run Run Shaw Hospital, Zhejiang University School
of Medicine (approval no. 20171120-14; Hangzhou, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Takasu C, Vaziri ND, Li S, Robles L, Vo K,
Takasu M, Pham C, Farzaneh SH, Shimada M, Stamos MJ, et al:
Treatment with dimethyl fumarate ameliorates liver
ischemia/reperfusion injury. World J Gastroenterol. 23:4508–4516.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Peralta C, Jiménez-Castro MB and
Gracia-Sancho J: Hepatic ischemia and reperfusion injury: Effects
on the liver sinusoidal milieu. J Hepatol. 59:1094–1106. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shen XD, Ke B, Zhai Y, Amersi F, Gao F,
Anselmo DM, Busuttil RW and Kupiec-Weglinski JW: CD154-CD40 T-cell
costimulation pathway is required in the mechanism of hepatic
ischemia/reperfusion injury, and its blockade facilitates and
depends on heme oxygenase-1 mediated cytoprotection.
Transplantation. 74:315–319. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zabala V, Boylan JM, Thevenot P, Frank A,
Senthoor D, Iyengar V, Kim H, Cohen A, Gruppuso PA and Sanders JA:
Transcriptional changes during hepatic ischemia-reperfusion in the
rat. PLoS One. 14:e02270382019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Suzuki E, Ochiai-Shino H, Aoki H, Onodera
S, Saito A, Saito A and Azuma T: Akt activation is required for
TGF-β1-induced osteoblast differentiation of MC3T3-E1
pre-osteoblasts. PLoS One. 9:e1125662014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Derynck R and Budi EH: Specificity,
versatility, and control of TGF-β family signaling. Sci Signal.
12:eaav51832019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Brown KA, Pietenpol JA and Moses HL: A
tale of two proteins: Differential roles and regulation of Smad2
and Smad3 in TGF-beta signaling. J Cell Biochem. 101:9–33. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zou GL, Zuo S, Lu S, Hu RH, Lu YY, Yang J,
Deng KS, Wu YT, Mu M, Zhu JJ, et al: Bone morphogenetic protein-7
represses hepatic stellate cell activation and liver fibrosis via
regulation of TGF-β/Smad signaling pathway. World J Gastroenterol.
25:4222–4234. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Inagaki Y and Okazaki I: Emerging insights
into Transforming growth factor beta Smad signal in hepatic
fibrogenesis. Gut. 56:284–292. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li T, Zhao S, Song B, Wei Z, Lu G, Zhou J
and Huo T: Effects of transforming growth factor β-1 infected human
bone marrow mesenchymal stem cells on high- and low-metastatic
potential hepatocellular carcinoma. Eur J Med Res. 20:562015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Porowski D, Wirkowska A, Hryniewiecka E,
Wyzgał J, Pacholczyk M and Pączek L: Liver Failure Impairs the
Intrahepatic Elimination of Interleukin-6, Tumor Necrosis
Factor-Alpha, Hepatocyte Growth Factor, and Transforming Growth
Factor-Beta. BioMed Res Int. 2015:9340652015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang RQ, Mi HM, Li H, Zhao SX, Jia YH and
Nan YM: Modulation of IKKβ/NF-κB and TGF-β1/Smad via Fuzheng Huayu
recipe involves in prevention of nutritional steatohepatitis and
fibrosis in mice. Iran J Basic Med Sci. 18:404–411. 2015.PubMed/NCBI
|
|
13
|
Park SO, Kumar M and Gupta S: TGF-β and
iron differently alter HBV replication in human hepatocytes through
TGF-β/BMP signaling and cellular microRNA expression. PLoS One.
7:e392762012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Akhmetshina A, Palumbo K, Dees C, Bergmann
C, Venalis P, Zerr P, Horn A, Kireva T, Beyer C, Zwerina J, et al:
Activation of canonical Wnt signalling is required for
TGF-β-mediated fibrosis. Nat Commun. 3:7352012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Feng XH and Derynck R: Specificity and
versatility in tgf-beta signaling through Smads. Annu Rev Cell Dev
Biol. 21:659–693. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ooshima A, Park J and Kim SJ:
Phosphorylation status at Smad3 linker region modulates
transforming growth factor-β-induced epithelial-mesenchymal
transition and cancer progression. Cancer Sci. 110:481–488. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu ZY, Pan HW, Cao Y, Zheng J, Zhang Y,
Tang Y, He J, Hu YJ, Wang CL, Zou QC, et al: Downregulated
microRNA-330 suppresses left ventricular remodeling via the
TGF-β1/Smad3 signaling pathway by targeting SRY in mice with
myocardial ischemia-reperfusion injury. J Cell Physiol.
234:11440–11450. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu FF, Liu CY, Li XP, Zheng SZ, Li QQ,
Liu Q and Song L: Neuroprotective effects of SMADs in a rat model
of cerebral ischemia/reperfusion. Neural Regen Res. 10:438–444.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Percie du Sert N, Ahluwalia A, Alam S,
Avey MT, Baker M, Browne WJ, Clark A, Cuthill IC, Dirnagl U,
Emerson M, et al: Reporting animal research: Explanation and
elaboration for the ARRIVE guidelines 2.0. PLoS Biol.
18:e30004112020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu Y, Lu T, Zhang C, Xu J, Xue Z,
Busuttil RW, Xu N, Xia Q, Kupiec-Weglinski JW and Ji H: Activation
of YAP attenuates hepatic damage and fibrosis in liver
ischemia-reperfusion injury. J Hepatol. 71:719–730. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Malý O, Zajak J, Hyšpler R, Turek Z,
Astapenko D, Jun D, Váňová N, Kohout A, Radochová V, Kotek J, et
al: Inhalation of molecular hydrogen prevents ischemia-reperfusion
liver damage during major liver resection. Ann Transl Med.
7:7742019. View Article : Google Scholar
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Han L, Wang JN, Cao XQ, Sun CX and Du X:
An-te-xiao capsule inhibits tumor growth in non-small cell lung
cancer by targeting angiogenesis. Biomed Pharmacother. 108:941–951.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang X, Letterio JJ, Lechleider RJ, Chen
L, Hayman R, Gu H, Roberts AB and Deng C: Targeted disruption of
SMAD3 results in impaired mucosal immunity and diminished T cell
responsiveness to TGF-beta. EMBO J. 18:1280–1291. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bravatà V, Cammarata FP, Minafra L,
Pisciotta P, Scazzone C, Manti L, Savoca G, Petringa G, Cirrone
GAP, Cuttone G, et al: Proton-irradiated breast cells: Molecular
points of view. J Radiat Res (Tokyo). 60:451–465. 2019. View Article : Google Scholar
|
|
26
|
Zhai Y, Petrowsky H, Hong JC, Busuttil RW
and Kupiec-Weglinski JW: Ischaemia-reperfusion injury in liver
transplantation--from bench to bedside. Nat Rev Gastroenterol
Hepatol. 10:79–89. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jiménez-Castro MB, Cornide-Petronio ME,
Gracia-Sancho J and Peralta C: Inflammasome-Mediated Inflammation
in Liver Ischemia-Reperfusion Injury. Cells. 8:E11312019.
View Article : Google Scholar
|
|
28
|
Lee PY, Wang JX, Parisini E, Dascher CC
and Nigrovic PA: Ly6 family proteins in neutrophil biology. J
Leukoc Biol. 94:585–594. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xiang S, Chen K, Xu L, Wang T and Guo C:
Bergenin Exerts Hepatoprotective Effects by Inhibiting the Release
of Inflammatory Factors, Apoptosis and Autophagy via the PPAR-γ
Pathway. Drug Des Devel Ther. 14:129–143. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Palomino-Schätzlein M, Simó R, Hernández
C, Ciudin A, Mateos-Gregorio P, Hernández-Mijares A, Pineda-Lucena
A and Herance JR: Metabolic fingerprint of insulin resistance in
human polymorphonuclear leucocytes. PLoS One. 13:e01993512018.
View Article : Google Scholar
|
|
31
|
Cutrn JC, Perrelli MG, Cavalieri B,
Peralta C, Rosell Catafau J and Poli G: Microvascular dysfunction
induced by reperfusion injury and protective effect of ischemic
preconditioning. Free Radic Biol Med. 33:1200–1208. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bielecka-Dabrowa A, Gluba-Brzózka A,
Michalska-Kasiczak M, Misztal M, Rysz J and Banach M: The
multi-biomarker approach for heart failure in patients with
hypertension. Int J Mol Sci. 16:10715–10733. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang G, Ge M, Han Z, Wang S, Yin J, Peng
L, Xu F, Zhang Q, Dai Z, Xie L, et al: Wnt/β-catenin signaling
pathway contributes to isoflurane postconditioning against cerebral
ischemia-reperfusion injury and is possibly related to the
transforming growth factorβ1/Smad3 signaling pathway. Biomed
Pharmacother. 110:420–430. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shang J, Sun S, Zhang L, Hao F and Zhang
D: miR-211 alleviates ischaemia/reperfusion-induced kidney injury
by targeting TGFβR2/TGF-β/SMAD3 pathway. Bioengineered. 11:547–557.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang S, Yin J, Ge M, Dai Z, Li Y, Si J, Ma
K, Li L and Yao S: Transforming growth-beta 1 contributes to
isoflurane postconditioning against cerebral ischemia-reperfusion
injury by regulating the c-Jun N-terminal kinase signaling pathway.
Biomed Pharmacother. 78:280–290. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yang T, Zhang X, Ma C and Chen Y:
TGF-β/Smad3 pathway enhances the cardio-protection of S1R/SIPR1 in
in vitro ischemia-reperfusion myocardial cell model. Exp Ther Med.
16:178–184. 2018.PubMed/NCBI
|
|
37
|
Larsson J, Goumans MJ, Sjöstrand LJ, van
Rooijen MA, Ward D, Levéen P, Xu X, ten Dijke P, Mummery CL and
Karlsson S: Abnormal angiogenesis but intact hematopoietic
potential in TGF-beta type I receptor-deficient mice. EMBO J.
20:1663–1673. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Vander Ark A, Cao J and Li X: TGF-β
receptors: In and beyond TGF-β signaling. Cell Signal. 52:112–120.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Schepers D, Tortora G, Morisaki H,
MacCarrick G, Lindsay M, Liang D, Mehta SG, Hague J, Verhagen J,
van de Laar I, et al: A mutation update on the LDS-associated genes
TGFB2/3 and SMAD2/3. Hum Mutat. 39:621–634. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Waldrip WR, Bikoff EK, Hoodless PA, Wrana
JL and Robertson EJ: Smad2 signaling in extraembryonic tissues
determines anterior-posterior polarity of the early mouse embryo.
Cell. 92:797–808. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Stewart AG, Thomas B and Koff J: TGF-β:
Master regulator of inflammation and fibrosis. Respirology.
23:1096–1097. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Goumans MJ and Mummery C: Functional
analysis of the TGFbeta receptor/Smad pathway through gene ablation
in mice. Int J Dev Biol. 44:253–265. 2000.PubMed/NCBI
|
|
43
|
Taylor KR, Trowbridge JM, Rudisill JA,
Termeer CC, Simon JC and Gallo RL: Hyaluronan fragments stimulate
endothelial recognition of injury through TLR4. J Biol Chem.
279:17079–17084. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wu H, Chen G, Wyburn KR, Yin J, Bertolino
P, Eris JM, Alexander SI, Sharland AF and Chadban SJ: TLR4
activation mediates kidney ischemia/reperfusion injury. J Clin
Invest. 117:2847–2859. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Imai Y, Kuba K, Neely GG,
Yaghubian-Malhami R, Perkmann T, van Loo G, Ermolaeva M, Veldhuizen
R, Leung YH, Wang H, et al: Identification of oxidative stress and
Toll-like receptor 4 signaling as a key pathway of acute lung
injury. Cell. 133:235–249. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Liu Q and Zhang Y: PRDX1 enhances cerebral
ischemia-reperfusion injury through activation of TLR4-regulated
inflammation and apoptosis. Biochem Biophys Res Commun.
519:453–461. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
McCartney-Francis N, Jin W and Wahl SM:
Aberrant Toll receptor expression and endotoxin hypersensitivity in
mice lacking a functional TGF-beta 1 signaling pathway. J Immunol.
172:3814–3821. 2004. View Article : Google Scholar : PubMed/NCBI
|














