Introduction
Osteoporosis is a systemic skeletal disease
characterized by reduced bone density, which leads to an increased
risk of fracture (1). Osteoporosis
is considered a public health problem, the prevalence of which is
increasing worldwide (2). Bone
homeostasis requires maintenance of a balance between
osteoclast-induced bone resorption and osteoblast-induced bone
formation; however, an increase in osteoclast number and activity
can result in an imbalance in bone remodeling (3). This imbalance can result in
osteoclast-mediated bone diseases, such as rheumatoid arthritis,
periodontal disease, Paget's disease and osteoporosis (4).
Bisphosphonate and hormone replacement therapy with
estrogen are the most widely used treatments for postmenopausal
osteoporosis (5). However,
long-term treatment with bisphosphonate is unsuitable for several
patients with osteoporosis owing to serious side effects, such as
osteonecrosis of the jaw, atrial fibrillation, esophageal cancer,
musculoskeletal pain and atypical fractures (6–9). In
addition, hormone replacement therapy has unwanted side effects,
such as an increased risk of developing ovarian cancer and breast
cancer (10–12). Thus, it is essential to discover
novel effective and safe treatments for osteoporosis.
Osteoclasts are multinucleated giant cells derived
from monocyte/macrophage hematopoietic precursor cells (13). Receptor activator of nuclear factor
(NF)-κB (RANK) ligand (RANKL) is an essential cytokine of
osteoclast differentiation (14).
Binding of RANKL to RANK induces recruitment of the adaptor protein
tumor necrosis factor (TNF) receptor-associated factor (TRAF6).
TRAF6 induces the translocation of NF-κB (15,16).
These signaling pathways are responsible for the activation of key
transcription factors, such as c-Fos and nuclear factor of
activated T cell, cytoplasmic 1 (NFATc1), resulting in increased
expression of osteoclast-specific genes, including
tartrate-resistant acid phosphatase (TRAP/Acp5), ATPase H+
transporting V0 subunit D2 (ATP6v0d2/Atp6v0d2),
osteoclast-associated immunoglobulin-like receptor
(OSCAR/Oscar), cathepsin K (CTK/Ctsk) and matrix
metalloproteinase-9 (MMP-9/Mmp9) (17–20).
Melandrium firmum (Siebold & Zucc.)
Rohrbach (MFR) is the dried aerial portion of Melandrii Herba
Rohrbach, a member of the Caryophyllaceae family. MFR is known as
‘Wangbulryuhaeng’ in Korea and has been traditionally used to treat
gynecological conditions, such as breast cancer and lactation
disorders (21). Previous studies
have shown that the methanol extract of MFR exhibits an
anti-inflammatory effect on lipopolysaccharide (LPS)-induced
proinflammatory cytokines, such as TNF-α and interleukin (IL)-1β
(22,23). According to previous studies,
inhibition of TNF-α and IL-1β has been associated with metabolic
bone disease, such as postmenopausal osteoporosis and rheumatoid
arthritis. IL-1β has been revealed to be regulated by RANKL to
increase activity and promote osteoclast formation (24,25).
In addition, TNF-α may serve an important role in regulating bone
homeostasis by stimulating osteoclast formation and inhibiting
osteoblast function (26,27). Furthermore, vitexin, another active
compound found in MFR, has been shown to inhibit osteoclastogenesis
and prevent LPS-induced osteolysis (28). Based on these previous findings, it
was hypothesized that MFR may potentially attenuate osteoclast
differentiation, thus preventing bone loss in osteoporosis.
However, to the best of our knowledge, the effect of MFR on
osteoclast differentiation and osteoporosis is yet to be
explained.
In the present study, the effect of MFR on
osteoclast differentiation in RANKL-induced mouse macrophage RAW
264.7 cells and bone loss in an ovariectomized (OVX) rat model of
osteoporosis was assessed.
Materials and methods
Reagents
DMEM was purchased from Welgene, Inc. Minimum
essential medium-α (α-MEM), penicillin/streptomycin (P/S) and
Dulbecco's PBS (DPBS) were obtained from Gibco; Thermo Fisher
Scientific, Inc. FBS was purchased from Atlas Biologicals. RANKL
was purchased from PeproTech, Inc. CellTiter 96 Aqueous
non-radioactive cell proliferation (MTS) assay was purchased from
Promega Corporation. Bicinchoninic acid (BCA) solution, phosphatase
inhibitor cocktail, DAPI, 17β-estradiol (E2) and
alendronate (ALN) were obtained from Sigma-Aldrich; Merck KGaA.
Osteo Assay Surface multiple well plates (cat. no. 3989) were
obtained from Corning, Inc.. PCR primers were synthesized by
Genotech Corp. Acti-stain™ 488 Fluorescent Phalloidin was purchased
from Cytoskeleton, Inc.. The primary antibodies and secondary
antibodies used in the present study were: β-actin (cat. no.
sc-8432; Santa Cruz Biotechnology, Inc.), c-Fos (cat. no. sc-447;
Santa Cruz Biotechnology, Inc.), NFATc1 (cat. no. 556602; BD
Biosciences), MMP-9 (cat. no. ab38898; Abcam), CTK (cat. no.
ab19027; Abcam), TRAF6 (cat. no. sc-8409; Santa Cruz Biotechnology,
Inc.) and peroxidase AffiniPure Goat Anti-Mouse IgG (cat no.
115-035-062; Jackson ImmunoResearch Laboratories, Inc.) and
peroxidase AffiniPure Goat Anti-Rabbit IgG (cat no. 115-035-144;
Jackson ImmunoResearch Laboratories, Inc). All other reagents used
were of analytical grade.
Preparation of MFR extract
MFR was purchased from Omniherb. MFR was prepared by
decocting 300 g of the dried herb in 1.5 l boiling distilled water
(D.W) for 2 h. The extract was filtered using filter paper (no. 3;
Whatman plc; GE Healthcare Life Sciences) and collected in a rotary
evaporator at 55°C. The extract was lyophilized and 19 g dried
powder was obtained (yield ratio, 12.7%).
High-performance liquid chromatography
(HPLC) analysis
Vitexin (cat. no. 49513; Sigma-Aldrich; Merck KGaA)
is the active ingredient in MFR (29); therefore, to quantitatively evaluate
the MFR extract, HPLC was performed with vitexin used as an
internal standard. Vitexin was prepared in DMSO. Analysis was
performed using a UV detector (2996 Waters 2695; Waters
Corporation); separation was carried out on an Xbridge-C18 with a
C18 guard column (250 × 4.6 mm; 5 µm; Waters Corporation) and
proceeded at 30°C for 50 min at a flow rate of 1 ml/min. Vitexin
was detected at 335 nm. Samples were injected in a volume of 10 µl.
The mobile phase consisted of acetonitrile (A) and water (B), at a
composition of 10% A from 0–10 min and 50% A from 10–30 min.
RAW 264.7 cell culture and
cytotoxicity assay
RAW 264.7 cells are mouse monocyte-macrophage like
cells (30). RAW 264.7 cells were
obtained from Korean Cell Line Bank; Korean Cell Line Research
Foundation. RAW 264.7 cells were cultured in DMEM supplemented with
10% FBS and 1% P/S in a cell incubator at 37°C and 5%
CO2. To examine cell viability, the cells were seeded in
a 96-well plate at 5×103 cells/well and incubated for 24
h. RAW 264.7 cells were treated with MFR (12.5, 25, 50 and 100
µg/ml) at 37°C for 24 h. After treatment, 20 µl/well MTS solution
was added and the cells were incubated at 37°C for 2 h. To
determine cell viability of mature osteoclasts, RAW 264.7 cells
were seeded in a 96-well plate at 5×103 cells/well and
were treated with or without RANKL (100 ng/ml) for 5 days.
Subsequently, cells were treated with MFR (12.5, 25, 50 and 100
µg/ml) for 1 day, after which, 20 µl MTS solution was added and
incubated at 37°C for 2 h. Cell viability was measured using an
enzyme-linked immunosorbent assay (ELISA) reader at a wavelength of
490 nm.
TRAP staining and measurement of TRAP
levels
A total of 5×103 RAW 264.7 cells/well
were seeded in a 96-well plate. After 24 h, to induce osteoclast
differentiation of RAW 264.7 cells, cells were treated with RANKL
(100 ng/ml) and MFR (12.5, 25, 50 and 100 µg/ml) at 37°C for 5
days. The medium was replaced every 2 days. In order to compare the
inhibitory effect of MFR and vitexin on osteoclast differentiation,
cells were seeded in the same manner as mentioned previously. After
24 h, the medium was replaced with RANKL (100 ng/ml) and MFR (50
and 100 µg/ml) or vitexin (0.0753 and 0.147 µg/ml) at 37°C for 5
days. Subsequently, the cells were washed and fixed with 4%
formalin at room temperature for 10 min. TRAP staining was
performed using a TRAP kit (Sigma-Aldrich; Merck KGaA) according to
the manufacturer's protocol. The number of TRAP-positive cells was
counted using an inverted light microscope (Olympus Corporation;
magnification, ×100).
TRAP is secreted in large quantities by osteoclasts
and TRAP levels are considered a biochemical marker of osteoclast
function (31). Therefore, the
present study also analyzed TRAP levels to determine the effect of
MFR on osteoclast function. In order to measure the TRAP levels, 50
µl differentiation medium was obtained and transferred to a new
plate. Subsequently, an equal volume of TRAP solution (4.93 mg
4-Nitrophenyl phosphate disodium salt hexahydrate in 850 µl 0.5 M
acetate solution and 150 µl tartrate solution) was added to the
differentiation medium and incubated at 37°C for 1 h. Subsequently,
the reaction was terminated using 0.5 M NaOH and TRAP levels were
measured using an ELISA reader at a wavelength of 405 nm.
Filamentous actin (F-actin) ring
formation and pit formation
To determine F-actin ring formation,
5×103 cells/well were seeded in a 96-well plate, and
treated with RANKL (100 ng/ml) and MFR (12.5, 25, 50 and 100 µg/ml)
at 37°C for 5 days. The medium was replaced every 2 days. The cells
were fixed with 4% paraformaldehyde at room temperature for 20 min
and permeabilized with 0.1% Triton X-100 in PBS at room temperature
for 5 min. The cells were then stained using Acti-stain™ 488
Fluorescent Phalloidin at room temperature in the dark for 30 min.
Cells were washed with PBS and nuclei were counterstained with
DAPI. Images were captured using fluorescence microscopy (Cellena;
Logos Biosystems; magnification, ×200).
To examine pit formation, the cells were seeded in
Osteo Assay Surface multiple well plates at 5×103
cells/well. Subsequently, the cells were treated with RANKL (100
ng/ml) and MFR (12.5, 25, 50 and 100 µg/ml) at 37°C for 5 days. The
differentiation medium was replaced every 2 days. Subsequently,
cells were washed and removed using deionized water with 4% sodium
hypochlorite. Images of resorbed areas were captured using an
inverted light microscope (Olympus Corporation; magnification,
×200). Total pit areas were analyzed using ImageJ version 1.46
(National Institutes of Health).
Western blotting
To examine the effect of MFR on osteoclast
differentiation-associated transcription factors, 5×105
RAW 264.7 cells/well were seeded in a 60-mm dish and treated with
RANKL (100 ng/ml) and MFR (12.5, 25, 50 and 100 µg/ml) at 37°C for
1 day. In order to compare the inhibitory effect of MFR and vitexin
on the expression of NFATc1 and c-Fos, 5×105 RAW 264.7
cells/well were seeded in a 60-mm dish and treated with RANKL (100
ng/ml) and MFR (50 and 100 µg/ml) or vitexin (0.0753 and 0.147
µg/ml) at 37°C for 1 day. To prepare whole-cell lysates, the cells
were washed with cold DPBS, and the total proteins were extracted
using RIPA lysis buffer (50 mM Tris-CI, 150 mM NaCl, 1% NP-40, 0.5%
Na-deoxycholate, 0.1% SDS, protease inhibitor cocktail, phosphatase
inhibitor cocktail). Protein concentration was determined using a
BCA assay. Proteins (30 µg) were separated by SDS-PAGE on 10% gels
and transferred to a nitrocellulose membrane (Whatman plc; GE
Healthcare Life Sciences) according to the manufacturer's protocol.
The membranes were blocked in 5% skimmed milk for 1 h at 37°C,
followed by overnight incubation at 4°C with primary antibodies
against NFATc1 (1:1,000), c-Fos (1:1,000), TRAF6 (1:1,000), MMP-9
(1:1,000), CTK (1:1,000) and β-actin (1:1,000). Subsequently,
membranes were incubated with secondary antibodies (1:10,000) for 1
h at room temperature. The membranes were visualized using enhanced
chemiluminescence reagent (cat no. RPN2106; GE Healthcare Life
Sciences) and protein expression levels were semi-quantified using
ImageJ (version 1.46; National Institutes of Health).
Semi-quantitative reverse
transcription-PCR (RT-PCR)
To examine the effect of MFR on osteoclast-related
markers, 2×105 cells/well seeded in a 6-well plate, and
treated with the RANKL (100 ng/ml) and MFR (12.5, 25, 50 and 100
µg/ml) at 37°C for 4 days. Total RNA of the treated cells was
extracted using RNAiso Plus (cat. no. 9109; Takara Bio, Inc.)
according to the manufacturer's protocol. A total of 2 µg RNA was
measured using a NanoDrop 2.0 spectrophotometer (NanoDrop; Thermo
Fisher Scientific, Inc.). cDNA was synthesized using a RT kit
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. The synthesized cDNA was amplified by PCR
using Taq polymerase (Kapa Biosystems; Roche Diagnostics). The PCR
analysis conditions were as follows: 26–40 cycles of 30 sec at 94°C
(denaturation), 30 sec at 53–58°C (annealing) and 30 sec at 72°C
(extension). The primer sequences are listed in Table I. β-actin was used as a loading
control. The products of qPCR were assessed on a 2% agarose gel
stained with SYBR-Green (Invitrogen; Thermo Fisher Scientific,
Inc.). The expression levels of mRNA were semi-quantified using
ImageJ version 1.46.
 | Table I.Primer sequences for reverse
transcription-PCR analysis. |
Table I.
Primer sequences for reverse
transcription-PCR analysis.
| Gene name | Sequence
(5′-3′) | Temperature
(°C) | Cycle | Accession no. |
|---|
| Nfatc1 | F:
TGCTCCTCCTCCTGCTGCTC | 58 | 32 | NM_198429.2m |
|
| R:
CGTCTTCCACCTCCACGTCG |
|
|
|
| Fos | F:
ATGGGCTCTCCTGTCAACAC | 58 | 40 | NM_010234.3 |
|
| R:
GGCTGCCAAAATAAACTCCA |
|
|
|
| Ctsk | F:
AGGCGGCTATATGACCACTG | 58 | 26 | NM_007802.4 |
|
| R:
CCGAGCCAAGAGAGCATATC |
|
|
|
| Mmp9 | F:
CGACTTTTGTGGTCTTCCCC | 58 | 30 | NM_013599.4 |
|
| R:
TGAAGGTTTGGAATCGACCC |
|
|
|
| Ca2 | F:
CTCTCAGGACAATGCAGTGCTGA | 58 | 32 | NM_001357334.1 |
|
| R:
ATCCAGGTCACACATTCCAGCA |
|
|
|
| Acp5 | F:
ACTTCCCCAGCCCTTACTACCG | 58 | 30 | NM_007388.3 |
|
| R:
TCAGCACATAGCCCACACCG |
|
|
|
|
Atp6v0d2 | F:
ATGGGGCCTTGCAAAAGAAATCTG | 58 | 30 | NM_175406.3 |
|
| R:
CGACAGCGTCAAACAAAGGCTTGTA |
|
|
|
| Dcstamp | F:
TGGAAGTTCACTTGAAACTACGTG | 63 | 30 | NM_001289506.1 |
|
| R:
CTCGGTTTCCCGTCAGCCTCTCTC |
|
|
|
| Oscar | F:
CTGCTGGTAACGGATCAGCTCCCCAGA | 53 | 35 | NM_001290377.1 |
|
| R:
CCAAGGAGCCAGAACCTTCGAAACT |
|
|
|
| Src | F:
TCCAGGCTGAGGAGTGGTACTTTGG | 64 | 40 | NM_001025395.2 |
|
| R:
ATACGGTAGTGAGGCGGTGACACAG |
|
|
|
| Prdm1 | F:
TTCTTGTGTGGTATTG | 50 | 40 | NM_007548.4 |
|
| R:
TTGGGGACACTCTTTG |
|
|
|
| Actb | F:
TTCTACAATGAGCTGCGTGT | 58 | 30 | NM_007393 |
|
| R:
CTCATAGCTCTTCTCCAGGG |
|
|
|
Animal experimental design
To further investigate the anti-osteoporotic effects
of MFR in vivo, an OVX rat model of menopausal osteoporosis
was used. A number of 48 female Sprague Dawley (SD)-rats (age, 12
weeks; weight, 230–250 g) were obtained from KOATECH. The rats were
housed in a standard environment with a controlled temperature of
22±2°C and humidity of 55±5%, under a 12-h light/dark cycle.
Animals were provided with ad libitum access to water and
food. The protocol for in vivo experiments was approved by
the Kyung Hee Medical Center Institutional Animal Care and Use
Committee (KHMC-IACUC; approval no. KHMC-IACUC 19–017). All animals
were allowed to acclimate for 1 week. For the postmenopausal
osteoporosis model, rats were deeply anesthetized using 5%
isoflurane (inhaled in 100% oxygen) and the bilateral ovaries of
the rats were removed under 2–2.5% isoflurane anesthesia. The sham
group underwent the same surgery to ensure they experienced the
same stress, but the ovaries were not removed. No animals died
during surgery. To prevent infection of the surgical site, the
wound was sutured and gentamicin (4 mg/kg) was injected
intraperitoneally for 3 days. After 1 week, the rats were randomly
divided into six groups (n=8/group): i) Sham group, in which rats
underwent the sham operation and were treated with vehicle (water);
ii) OVX group, in which OVX rats were treated with vehicle; iii)
E2 group, in which OVX rats were treated with
E2 (100 µg/kg); iv) ALN group, in which OVX rats were
treated with ALN (5 mg/kg); v) MFR-L group, in which OVX rats were
treated with a low dose of MFR (16.9 mg/kg); and vi) MFR-H group,
in which OVX rats were treated with a high dose of MFR (169 mg/kg)
for 8 weeks. E2 and ALN were used as the positive
controls. The humane endpoints used in the present study were:
Dirty hair and eye discharge; self-injury and anxiety; vomiting and
hemoptysis; inactivity; or anxiety and headache. None of the
animals exhibited abnormal behavior.
MFR dose was calculated as follows: In Korean
medicine, based on an average adult weight of 60 kg, a single dose
of 8 g is recommended. MFR (1.016 g; yield, 12.7%) was considered
equivalent to 8 g; thus, 16.9 mg MFR was required per 1 kg.
Therefore, the MFR-L group was treated with 16.9 mg/kg. The MFR-H
group was treated as follows: Rats are well-known to exhibit
6.4-fold faster metabolism than humans (32). In vitro experiments revealed
that MFR exhibited a higher inhibition of osteoclast
differentiation at high concentrations than at low concentrations.
Based on these findings, rats in the MFR-H groups were treated with
MFR at concentrations 10-fold higher than those in the MFR-L group
to induce higher pharmacological effects. Therefore, the MFR-H
group was treated with 169 mg/kg MFR (33–35).
Oral administration was performed every morning for 8 weeks. Body
weight was measured once a week and the dose was adjusted to
weight. After 8 weeks, all animals were anesthetized by 5%
isoflurane of inhaled anesthetics in 100% oxygen. Blood was
collected using a cardiac puncture following sacrifice by cervical
vertebrae dislocation. Subsequently, the uterus and femur were
collected and weighed. Femurs samples were collected and fixed in
10% neutral buffered formalin for 1 day at room temperature. Uterus
samples were collected and then stored at −80°C.
Serum biochemical analysis
Blood samples were incubated at room temperature for
30 min, and centrifuged at 29,739 × g for 10 min at 4°C. Serum
samples were stored at −80°C until required. Serum levels of
alkaline phosphatase (ALP), aspartate aminotransferase (AST) and
alanine aminotransferase (ALT) were measured by DKKorea.
C-telopeptide of collagen type 1 (CTX-1) levels were measured using
an ELISA kit (Elabscience; cat. no. E-EL-R1456) according to the
manufacturer's protocol. TRAP levels were measured using a TRAP kit
as aforementioned.
Micro-computed tomography (micro-CT)
analysis
The femoral head was scanned using micro-CT
(SkyScan1176; Bruker Corporation). Bone microarchitecture
parameters, including bone mineral density (BMD), trabecular
thickness (Tb.Th) and trabecular separation (Tb.sp) were analyzed
using NRecon software (SkyScan version 1.6.10.1; Bruker
Corporation).
Histological analysis
The fixed femur samples were washed using D.W at
room temperature for 1 day and decalcified using 10% EDTA at room
temperature for 3 weeks. Subsequently, the femur samples were
dehydrated at room temperature for 1 day and embedded in paraffin.
Paraffin-embedded tissues (5 µm) were sectioned on a rotary
microtome (Carl Zeiss AG) and tissue sections were mounted on
slides at room temperature for 1 day. The sections on the slides
were stained with hematoxylin-eosin (H&E); sections were
stained with hematoxylin for 10 min and with eosin for 10 sec at
room temperature. Subsequently, all slides were sealed using
mounting solution and the sections were viewed under a light
microscope (Olympus Corporation; magnifications, ×40 and ×100) for
histological evaluation.
Immunohistochemistry (IHC)
Paraffin-embedded tissues were deparaffinized in
xylene. Endogenous peroxidases were blocked in 0.3% hydrogen
peroxide at room temperature for 15 min and proteinase K (0.4
mg/ml) was used for antigen-retrieval at 37°C for 30 min. The
sectioned tissues were incubated with 10% normal serum (Gibco;
Thermo Fisher Scientific, Inc.) at room temperature for 1 h to
block nonspecific binding, then slides were washed with PBS and
incubated at 4°C for overnight with primary antibodies against
NFATc1 (1:100) and CTK (1:100). The following day, the slides were
washed with PBS and incubated with a secondary antibody (1:100;
rabbit; cat. no: BA-1000; Vector Laboratories, Inc.) at 4°C for 1
h. The signal was visualized using an ABC kit (Vector Laboratories,
Inc.) and 3, 3′-diaminobenzidine solution (Vector Laboratories,
Inc.). The stained tissues were imaged using a light microscope
(magnifications, ×100 and ×200).
MC3T3-E1 cell culture and cytotoxic
assay
MC3T3-E1 cells were purchased from American Type
Culture Collection. MC3T3-E1 cells were cultured in α-MEM without
ascorbic acid containing 10% FBS and 1% P/S in a cell incubator at
37°C and 5% CO2. To examine cell viability, the cells
were seeded in a 24-well plate at 1×104 cells/well and
incubated at 37°C for 24 h. Subsequently, the cells were treated
with α-MEM without ascorbic acid and MFR (12.5, 25, 50 and 100
µg/ml) at 37°C for 24 h. Subsequently, 20 µl/well MTS solution was
added to the cells and incubated at 37°C for 2 h. Cell viability
was evaluated using an ELISA reader at a wavelength of 490 nm and
was expressed as a percentage of the control.
Alizarin red S staining
MC3T3-E1 cells were seeded in a 24-well plate at
1×104 cells/well and incubated for 24 h. Subsequently,
osteoblast differentiation was induced using osteogenic medium
(α-MEM without ascorbic acid supplemented with 10 mM
β-glycerophosphate, 25 µg/ml ascorbic acid) and MFR (12.5, 25, 50
and 100 µg/ml) at 37°C for 14 days. The medium was replaced every 2
days. The cells were then washed with DPBS, fixed with 80% ethanol
at room temperature for 1 h and stained with Alizarin red S at room
temperature for 5 min. To quantify mineralization, the stained dye
was extracted using 10% (v/w) cetylpyridinium chloride in sodium
phosphate at room temperature for 15 min. The extracted dye was
measured using an ELISA reader at a wavelength of 570 nm.
Statistical analysis
Data are presented as the mean ± standard error of
the mean of three experiments. Statistical analyses were performed
using GraphPad Prism version 5.01 (GraphPad Software, Inc.). In
vitro and in vivo experiments were compared using a
one-way ANOVA followed by a Tukey's post hoc analysis. Animal body
weight was compared using a two-way ANOVA followed by Bonferroni
post hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
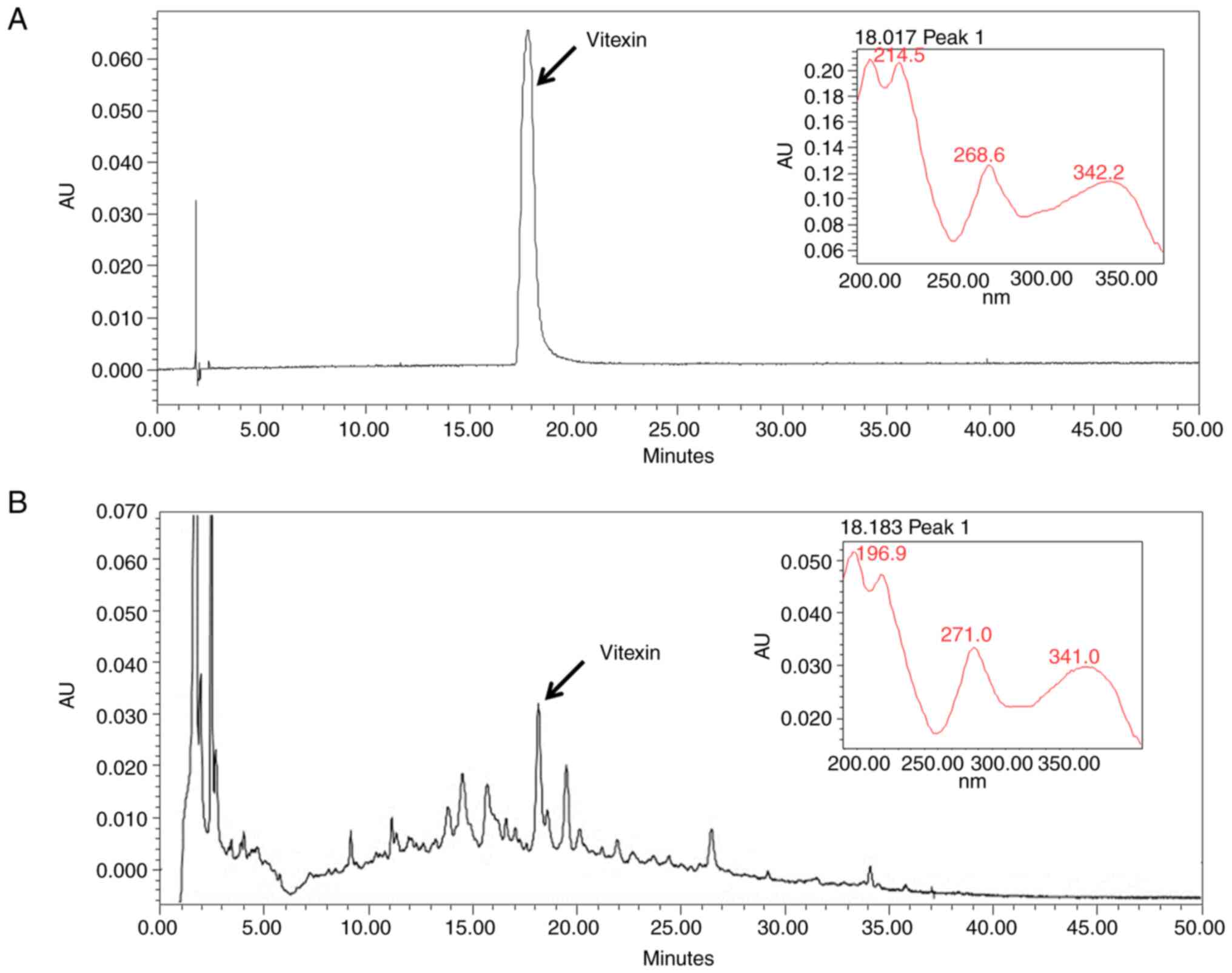
Quantitative analysis of the MFR
extract
Vitexin was used as a standard marker of MFR, as
described previously (29). The
chromatogram of the MFR water extract possessed several peaks at a
retention time of 0–30 min, and vitexin was observed at the same
retention time as the standard (Fig.
1).
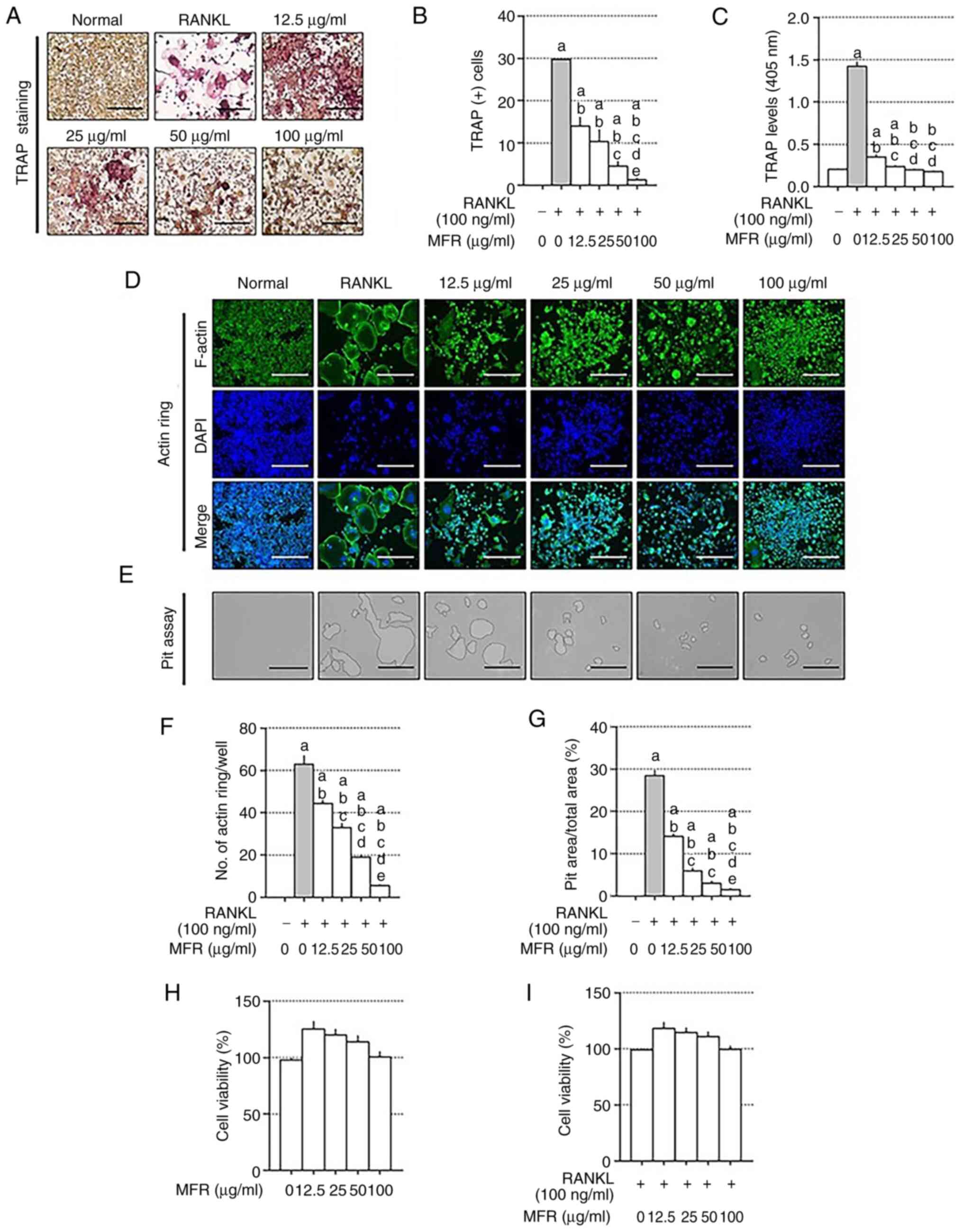
Effects of MFR on TRAP staining, and
F-actin ring and pit formation
To analyze the anti-osteoporotic effect of the MFR
extract in vitro, TRAP staining was performed using a TRAP
kit. Following treatment with RANKL, TRAP staining revealed that
the number of TRAP-positive cells was reduced by MFR treatment.
Consistent with the results of TRAP staining, MFR decreased TRAP
levels in the differentiation medium (Fig. 2A-C). As the actin ring is essential
in osteoclast differentiation (36), the effect of MFR on actin ring
formation was determined using immunocytochemistry. In addition,
the effect of MFR on pit formation was determined using
osteo-coated plates. The actin ring structures and the area of bone
resorption pits were increased in the RANKL-treated cells, but were
reduced following MFR treatment (Fig.
2D and E). Consistent with these results, MFR treatment reduced
the number of actin rings in a dose-dependent manner (Fig. 2F). In addition, the area of bone
resorption pits was significantly reduced following MFR treatment
(Fig. 2G). To confirm whether the
concentration of MFR used in the in vitro experiments
affected the viability of RAW 264.7 cells, the cells were treated
with 12.5–100 µg/ml MFR. MFR did not affect the viability of RAW
264.7 cells or mature osteoclasts (Fig.
2H and I). In addition, the present study compared and analyzed
the inhibitory effect of MFR and vitexin on osteoclast
differentiation to determine if the ability of MFR to inhibit
osteoclast differentiation was associated with vitexin. The content
of vitexin in MFR was 1.47 ppm (0.147%); 0.147 µg/ml vitexin in 100
µg/ml MFR and 0.0753 µg/ml vitexin in 50 µg/ml MFR. The effects of
the two substances (MFR and vitexin) on TRAP staining and TRAP
levels in the medium were subsequently assessed. To confirm whether
the concentration of vitexin contained in MFR affects the
osteoclast inhibitory effect, TRAP staining was performed. As shown
in Fig. S1A and B, TRAP-positive
cells and TRAP levels were increased in the RANKL-treated cells.
Conversely, the number of TRAP-positive cells was considerably
decreased by MFR (50 and 100 µg/ml) treatment compared with vitexin
(0.0753 and 0.147 µg/ml). Similarly, MFR (50 and 100 µg/ml) further
decreased TRAP levels compared with vitexin (0.0753 and 0.147
µg/ml) (Fig. S1B).
To determine whether the concentration of vitexin
contained in MFR affects the inhibitory effect on transcription
factors, NFATc1 and c-Fos were measured using western blotting. The
protein expression levels of NFATc1 and c-Fos were increased by
RANKL treatment. MFR (50 and 100 µg/ml) inhibited the expression of
NFATc1 and c-Fos compared with vitexin (0.0753 and 0.147 µg/ml)
(Fig. S1C and D). To confirm
whether the concentration of vitexin used in experiments affected
the viability of RAW 264.7 cells, the cells were treated with
vitexin (0.0753 and 0.147 µg/ml); vitexin not affect cell
cytotoxicity (Fig. S1E). Taken
together, it was revealed that the osteoclast inhibitory effect of
MFR was not mediated by vitexin.
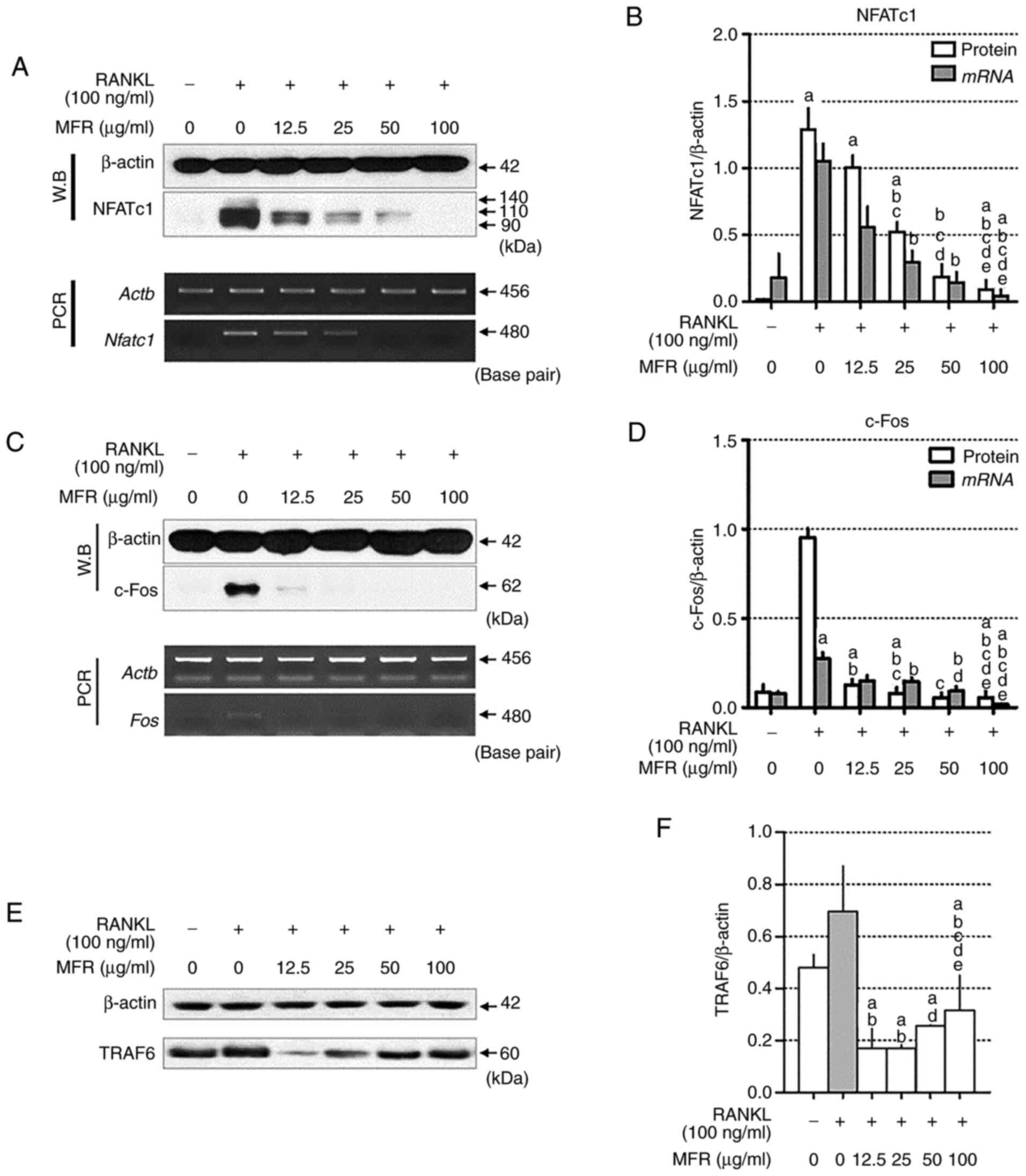
Effect of MFR on protein and mRNA
expression levels of NFATc1 and c-Fos
NFATc1 and c-Fos are important transcription factors
required for mature osteoclast differentiation in RAW 264.7 cells
(37,38). RANKL-induced expression of NFATc1
and c-Fos was measured using western blotting and RT-qPCR.
Treatment with RANKL significantly increased the expression levels
of NFATc1, whereas MFR decreased the protein and mRNA expression
levels of NFATc1 (Fig. 3A and B).
In addition, c-Fos expression was significantly increased by RANKL
treatment, whereas this increase was reversed by MFR treatment
(Fig. 3C and D). Furthermore, RANKL
stimulation increased TRAF6, but this finding was not significant,
and treatment of MFR attenuated the increased expression of TRAF6.
In particular, in cells treated with 25 µg/ml MFR, the expression
of TRAF6 was significantly decreased compared with that in
RANKL-only treated cells (Fig. 3E and
F).
Effect of MFR on RANKL-induced
expression of bone resorption and osteoclast-related markers
The inhibitory effects of the MFR extract on bone
resorption markers were examined using western blotting and
RT-qPCR. Treatment with RANKL significantly increased the
expression levels of MMP-9 and CTK, whereas MFR decreased the
protein expression levels of MMP-9 and CTK (Fig. 4A and B). Consistent with the results
of western blotting, MFR reduced the mRNA expression levels of
Mmp9, Ctsk and carbonic anhydrase 2 (CA2/Ca2)
(Fig. 4C and D). In addition, the
effects of MFR on RANKL-stimulated changes in osteoclast-specific
genes, including Acp5, Atp6v0d2, dendritic cell-specific
transmembrane protein (DC-STAMP/Dcstamp), Oscar,
c-Src (Src) and B lymphocyte-induced maturation protein-1
(Blimp-1/Prdm1), were determined, as they are crucial in
promoting osteoclastogenesis (39).
The mRNA expression levels of osteoclast-specific genes were
increased by RANKL treatment, whereas MFR attenuated the
RANKL-induced increase in the mRNA expression levels of
osteoclast-specific genes, including Acp5, Atp6v0d2, Dcstamp,
Oscar, Src and Prdm1, in a dose-dependent manner
(Fig. 4E-H).
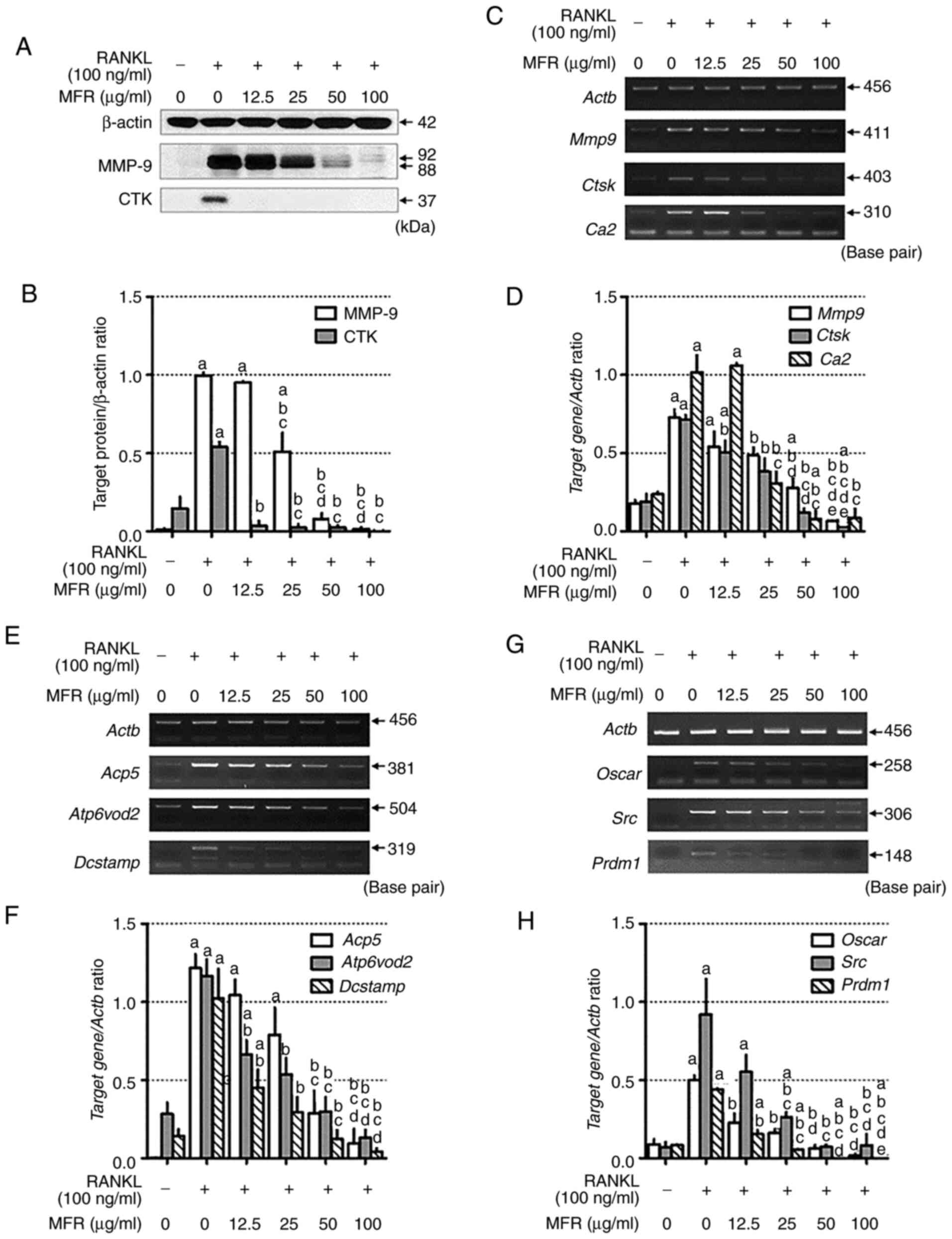 | Figure 4.Effect of MFR on the expression
levels of osteoclast-related genes. (A) Western blot analysis of
CTK and MMP-9. (B) Protein expression levels were normalized to
β-actin. (C) mRNA expression levels of Mmp9, Ctsk and
Ca2 were analyzed using RT-PCR. (D) mRNA expression levels
were normalized to Actb. (E) RT-PCR analysis was used to
determine the mRNA expression levels of Acp5, Atp6v0d2 and
Dcstamp, and (F) expression was normalized to Actb.
(G) RT-PCR analysis was performed to determine the mRNA expression
levels of Oscar, Src and Prdm1, and (H) expression
was normalized to Actb. All data are presented as the mean ±
standard error of the mean of three independent experiments. Data
were analyzed using one-way ANOVA followed by Tukey's post hoc
test. aP<0.05 vs. normal group (untreated cells);
bP<0.05 vs. RANKL treatment group;
cP<0.05 vs. MFR 12.5 µg/ml treatment group;
dP<0.05 vs. MFR 25 µg/ml treatment group;
eP<0.05 vs. MFR 50 µg/ml treatment group. MFR;
Melandrium firmum Rohrbach; RANKL, receptor activator of
nuclear factor-κB ligand; CTK/Ctsk, cathepsin k;
MMP-9/Mmp9, matrix metalloproteinase-9; CA2/Ca2,
carbonic anhydrase 2; RT-PCR, reverse transcription-PCR;
Acp5, tartrate-resistant acid phosphatase; Atp6v0d2,
ATPase H+ transporting V0 subunit D2; Dcstamp, dendritic
cell-specific transmembrane protein; Oscar,
osteoclast-associated receptor; Pdrm1, B lymphocyte-induced
maturation protein-1. |
Effect of MFR on body, uterus and
femur weights, and on the levels of hepatotoxicity, bone formation
and bone resorption markers
As shown in Fig. 5A,
the OVX group exhibited a significant increase in body weight
compared with that in the sham group from 4 weeks. The
E2 group had a significantly lower increase in body
weight after 3 weeks compared with the OVX group; however, MFR-L,
MFR-H and ALN treatment did not affect body weight. In addition,
the OVX group exhibited significantly decreased uterus weight
compared with that in the sham group, whereas E2
treatment prevented uterus weight loss. MFR-L, MFR-H and ALN groups
did not exhibit any protective effects on uterus weight loss
(Fig. 5B). While E2 and
ALN groups exhibited a preventive effect on the reduction of femur
weight compared with the OVX group, MFR-L and MFR-H groups did not
(Fig. 5C). To investigate the
extent of hepatotoxicity following treatment with MFR,
E2 and ALN, the serum levels of AST and ALT were
measured. Previous studies have shown that levels above 150 U/l for
AST and 40 U/l for ALT indicate hepatotoxicity in rats (40,41).
The levels of AST and ALT in the MFR-L, MFR-H, E2 and
ALN groups in the present study did not exceed these values;
therefore, it was indicated that MFR-L, MFR-H, E2 and
ALN did not induce hepatotoxicity (Fig.
5D and E). To determine the effect of MFR on bone formation,
the serum levels of ALP were measured. ALP was significantly
increased in the OVX group compared with that in the sham group;
moreover, the MFR-L group exhibited decreased serum levels of ALP,
whereas the MFR-H, E2 and ALN groups did not (Fig. 5F). To examine the effect of MFR on
bone resorption, serum levels of CTX-1 and TRAP were measured.
CTX-1 levels were increased in the OVX group compared with those in
the sham group, whereas they were significantly decreased in the
MFR-L, E2 and ALN groups compared with those in the OVX
group; there were no significant changes in the MFR-H group
(Fig. 5G). As shown in Fig. 5H, TRAP levels were increased due to
OVX, but the difference was not significant, whereas oral
administration of MFR-L, E2 and ALN reduced TRAP levels.
Furthermore, the TRAP levels were decreased in the MFR-H group;
however, this finding was not significant.
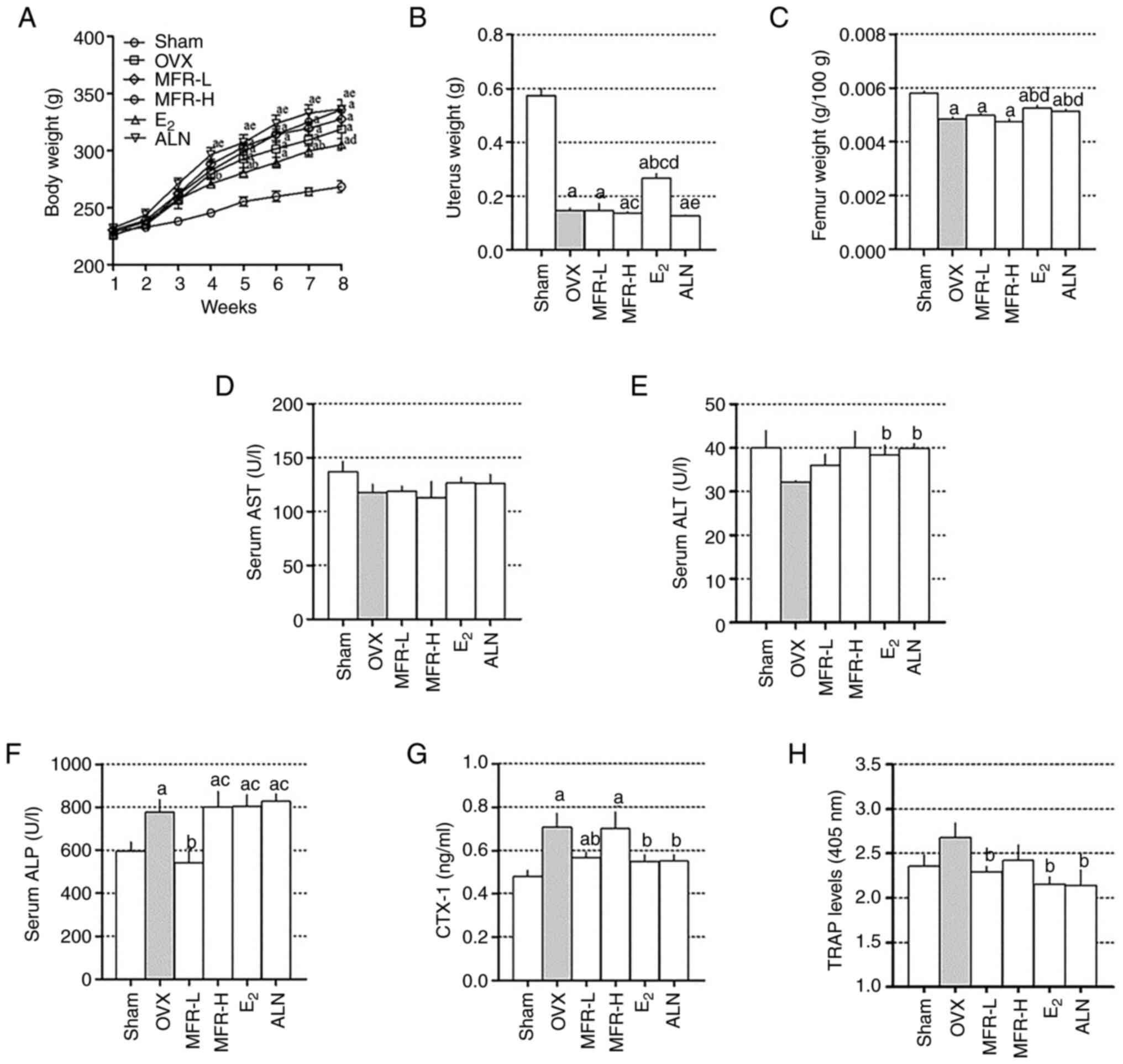 | Figure 5.Effect of MFR on body, uterus and
femur weights, and serum levels of ALP, AST and ALT. (A) Body
weight was measured weekly for 8 weeks. (B) Uterus and (C) femur
weights were measured after sacrificing the rats. The serum levels
of (D) AST, (E) ALT, (F) ALP, (G) CTX-1 and (H) TRAP were measured
using enzyme-linked immunosorbent assays. All data are presented as
the mean ± standard error of the mean (n=8 per group). (A) Data
were analyzed using two-way ANOVA followed by Bonferroni post hoc
test. (B-H) Data were analyzed using one-way ANOVA followed by
Tukey post hoc test. aP<0.05 vs. sham group;
bP<0.05 vs. OVX group; cP<0.05 vs.
MFR-L group; dP<0.05 vs. MFR-H group;
eP<0.05 vs. E2 group. MFR, Melandrium
firmum Rohrbach; OVX, ovariectomized; ALP, alkaline
phosphatase; AST, aspartate aminotransferase; ALT, alanine
aminotransferase; CTX-1, C-telopeptide of type I collagen; TRAP,
tartrate-resistant acid phosphatase; MFR-L, low dose of MFR; MFR-H,
high dose of MFR; E2, β-estradiol; ALN, alendronate. |
Effect of MFR on the bone
microarchitecture of ovariectomy-induced osteoporosis in rats
In order to investigate the protective effects of
MFR on ovariectomy-induced bone loss, micro-CT was used. As shown
in Fig. 6A, micro-CT images of the
femoral head indicated trabecular bone loss in OVX rats. BMD loss
was significantly inhibited in the MFR-L, E2 and ALN
groups compared with that in the OVX group (Fig. 6B). Furthermore, Tb.Th in the OVX
group was reduced compared with that in the sham group, whereas
Tb.Th was significantly increased in the ALN group. However, Tb.Th
in the MFR-L, MFR-H and E2 groups did not differ
significantly from that in the OVX group (Fig. 6C). As shown in Fig. 6D, Tb.sp was significantly increased
in the OVX group compared with that in the sham group, whereas
Tb.sp levels were decreased in the E2 and ALN groups.
However, Tb.sp levels were not affected in the MFR-L and MFR-H
groups.
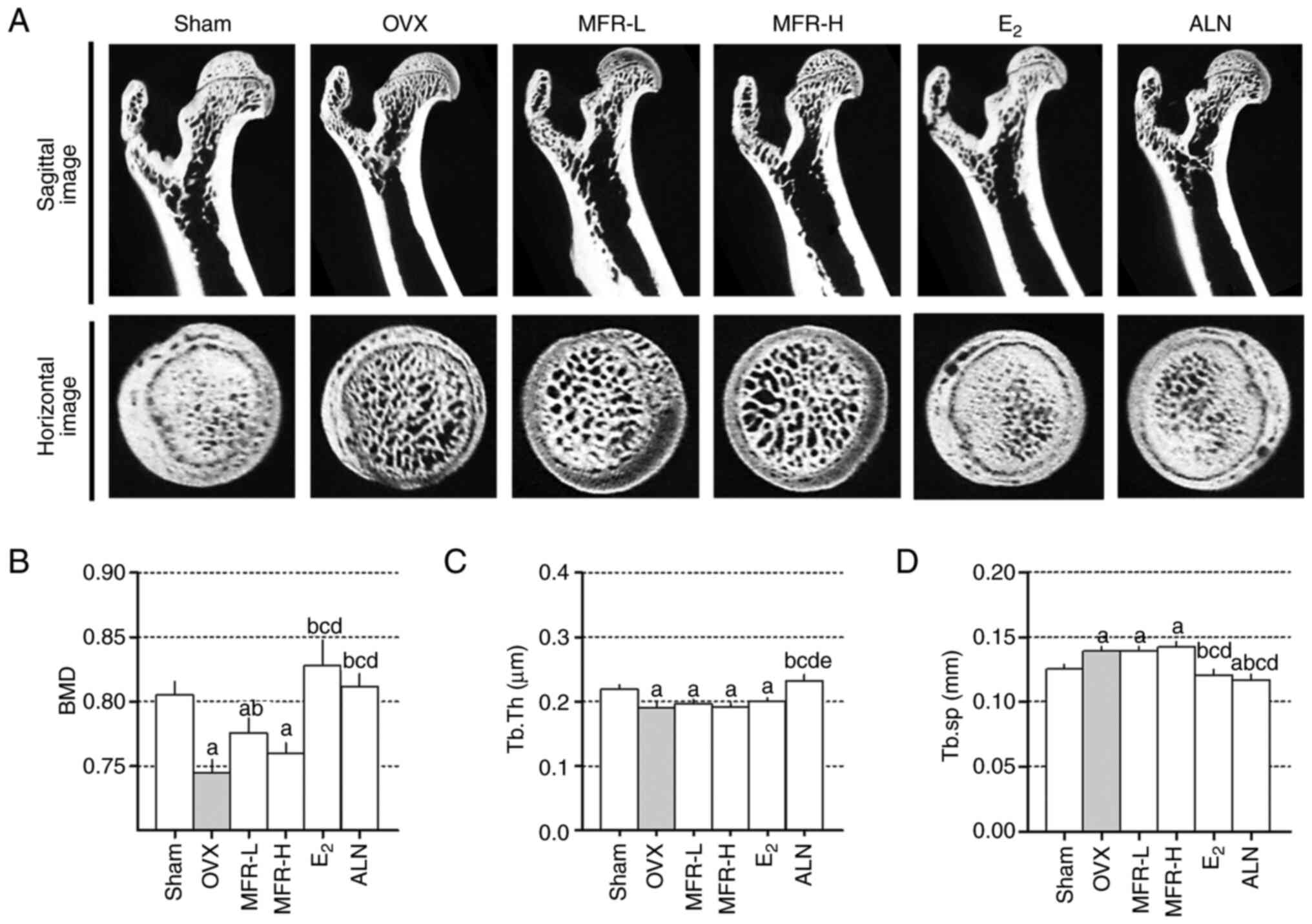 | Figure 6.Effects of MFR on bone
microarchitecture of OVX-induced osteoporosis rats. (A) Femurs were
analyzed using micro-CT imaging. (B) BMD, (C) Tb.Th and (D) Tb.sp
were analyzed using micro-CT image analysis. All data are presented
as the mean ± standard error of the mean (n=8/group). Statistical
analyses were performed using one-way ANOVA followed by Tukey post
hoc test. aP<0.05 vs. sham group;
bP<0.05 vs. OVX group; cP<0.05 vs.
MFR-L group; dP<0.05 vs. MFR-H group;
eP<0.05 vs. E2 group. MFR, Melandrium
firmum Rohrbach; OVX, ovariectomized; BMD, bone mineral
density; Tb.Th, trabecular thickness; Tb.sp, trabecular spacing;
micro-CT, micro computed tomography; MFR-L, low dose of MFR; MFR-H,
high dose of MFR; E2, β-estradiol; ALN, alendronate. |
Effect of MFR on bone loss in the OVX
rat model of osteoporosis
To investigate the histological changes in the
femoral head, H&E staining was performed on the bone tissue
(Fig. 7A). The trabecular area in
the femoral head of OVX rats was significantly decreased compared
with that in the sham group, whereas this reduction was prevented
by treatment with MFR-L, E2 and ALN (Fig. 7B).
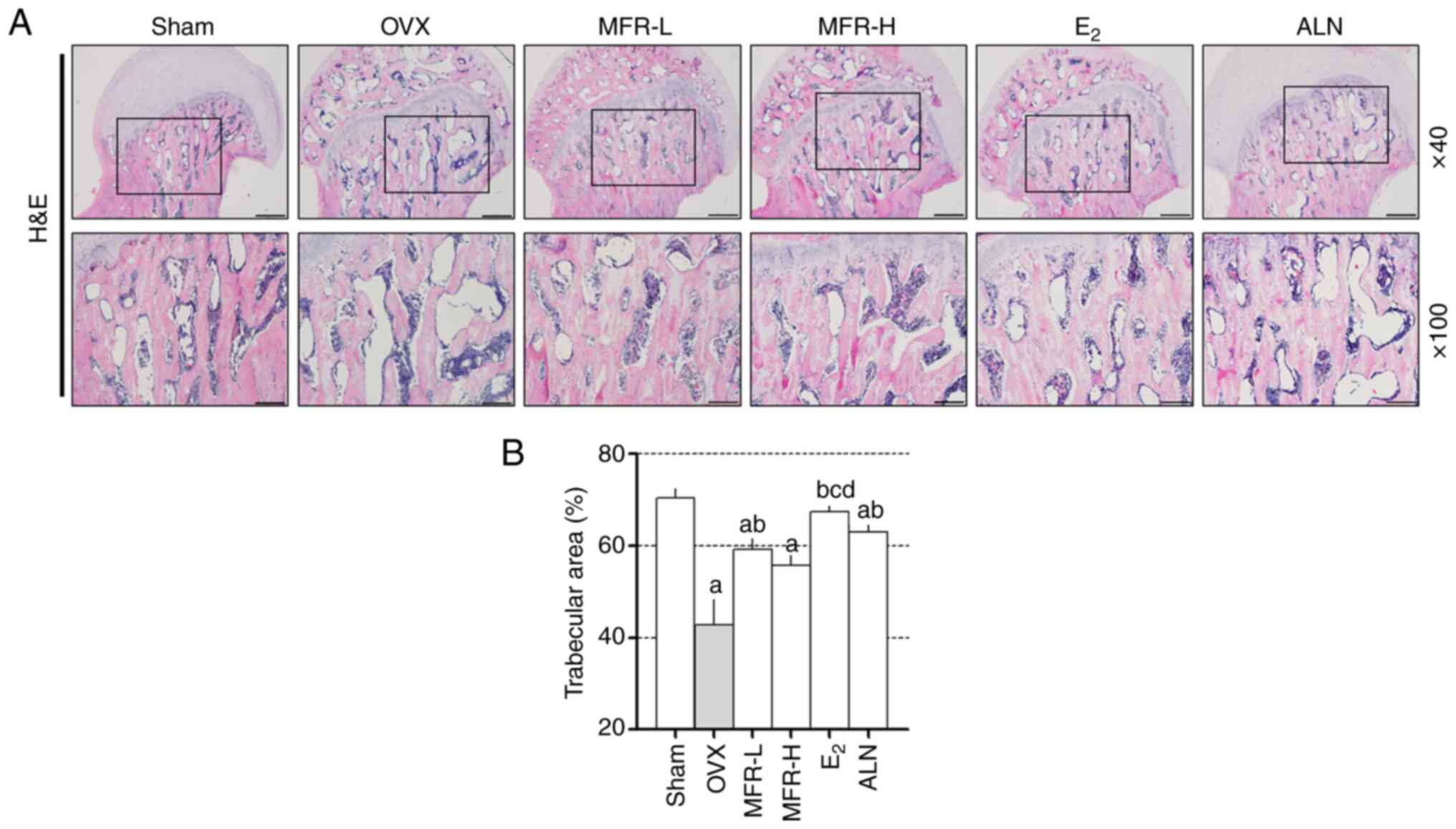 | Figure 7.Effect of MFR on trabecular area in
OVX-induced rats. (A) Bone tissues were stained with H&E.
Magnifications, ×40 and ×100; scale bar, 500 and 200 µm,
respectively. (B) Trabecular area was analyzed in the
H&E-stained sections. All data are presented as the mean ±
standard error of the mean (n=8/group). Statistical analyses are
performed using one-way ANOVA followed by Tukey post hoc test.
aP<0.05 vs. sham group; bP<0.05 vs. OVX
group; cP<0.05 vs. MFR-L group; dP<0.05
vs. MFR-H group. MFR, Melandrium firmum Rohrbach; OVX,
ovariectomized; H&E, hematoxylin and eosin; MFR-L, low dose of
MFR; MFR-H, high dose of MFR; E2, β-estradiol; ALN,
alendronate. |
Effect of MFR on the expression levels
of NFATc1 and CTK in femoral tissues in the in vivo osteoporosis
model
To determine the effect of MFR on the expression
levels of NFATc1 and CTK in the rat model of osteoporosis, IHC was
performed (Fig. 8A and B). The OVX
group exhibited increased expression levels of NFATc1 compared with
those in the sham group. By contrast, the MFR-L and E2
groups exhibited significantly reduced expression levels of NFATc1
expression; no changes were observed in the MFR-H and ALN groups
compared with the OVX group (Fig.
8C). In OVX rats, the expression levels of CTK were increased
compared with those in the sham group. Conversely, MFR-L, MFR-H,
E2 and ALN effectively reduced the expression levels of
CTK (Fig. 8D).
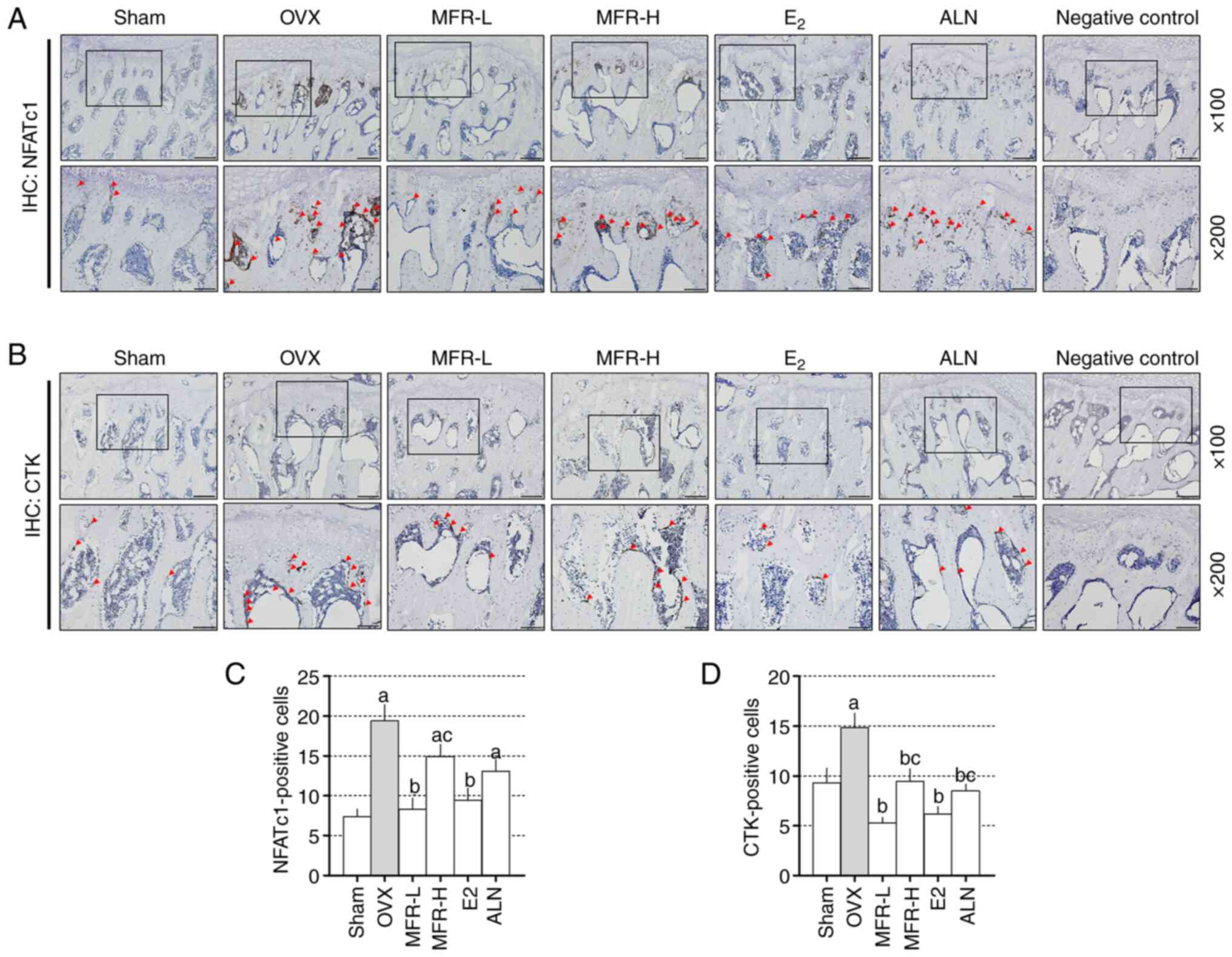 | Figure 8.Effects of MFR on the expression
levels of CTK and NFATc1 in OVX-induced rats. (A) NFATc1 and (B)
CTK-positive expression in bone tissue was assessed using IHC.
NFATc1 and CTK-positive expression is highlighted by the red
arrowheads. Mean number of (C) NFATc1 and (D) CTK-positive cells in
femur tissues was counted. Magnifications, ×100 and ×200; scale
bar, 200 and 100 µm, respectively. All data are presented as the
mean ± standard error of the mean (n=8/group). Statistical analyses
are performed using one-way ANOVA followed by Tukey post hoc test.
aP<0.05 vs. sham group; bP<0.05 vs. OVX
group; cP<0.05 vs. MFR-L group. MFR, Melandrium
firmum Rohrbach; CTK, cathepsin k; NFATc1, nuclear factor of
activated T-cells, cytoplasmic 1; OVX, ovariectomized; IHC,
immunohistochemistry; MFR-L, low dose of MFR; MFR-H, high dose of
MFR; E2, β-estradiol; ALN, alendronate. |
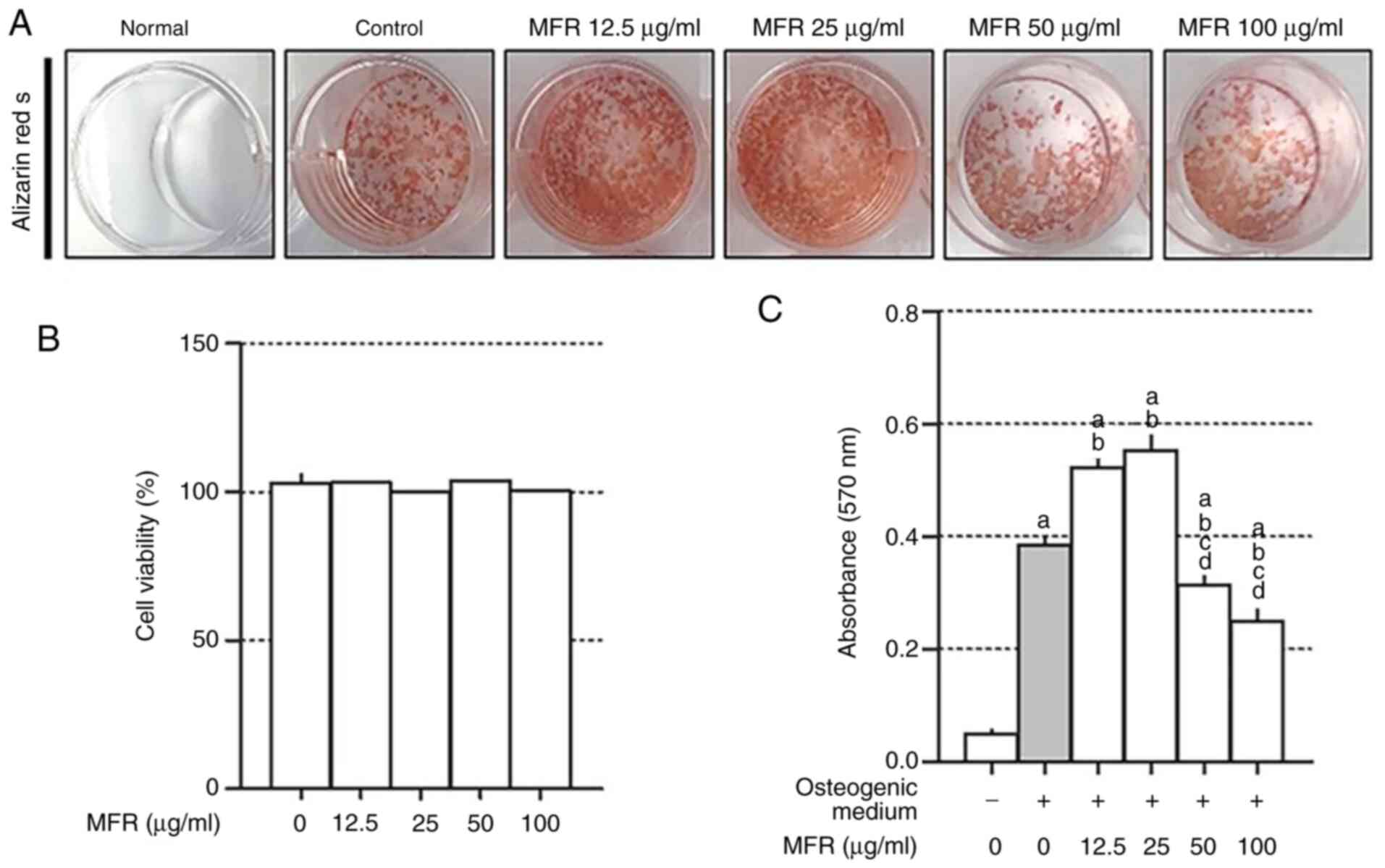
Effect of MFR on mineralization of
MC3T3-E1 cells
MC3T3-E1 cells derived from mouse calvaria are
characterized by proliferation, differentiation and mineralization
of osteoblasts, and are used in studies related to bone formation
(42). Alizarin red staining can be
used to detect the formation of mineralization (43). Therefore, to confirm the effect of
MFR on osteoblast differentiation and mineralization, Alizarin red
S staining was performed. The control cells exhibited significantly
increased mineralization compared with that in the normal cells.
Treatment with low concentrations of MFR (12.5 and 25 µg/ml)
promoted osteoblast differentiation, but high concentrations of MFR
(50 and 100 µg/ml) inhibited osteoblast differentiation (Fig. 9A). To examine the effects of the MFR
extract on MC3T3-E1 cell viability, cells were treated with
12.5–100 µg/ml MFR. Notably, MFR did not affect the viability of
MC3T3-E1 cells (Fig. 9B).
Quantification of Alizarin red staining revealed a significantly
reduced effect of MFR in a dose-dependent manner (Fig. 9C).
Discussion
The aim of the present study was to investigate the
inhibitory effect of MFR extract on osteoclast differentiation and
bone loss in ovariectomy-induced postmenopausal osteoporotic rats.
In vitro, MFR inhibited RANKL-induced osteoclast
differentiation, formation and function. In addition, MFR inhibited
the TRAF6 axis and inhibited the expression of NFATc1/c-Fos, which
is known as a key factor for osteoclast differentiation. Finally,
MFR suppressed the expression of osteoclast-related genes, such as
Acp5, Mmp9, CtsK, Atp6v0d2, Dcstamp and Src; this
potential mechanism of suppression is shown in Fig. 10. In vivo, MFR-L
considerably increased BMD, trabecular area, and decreased the
expression levels of NFATc1 and CTK in OVX-induced models.
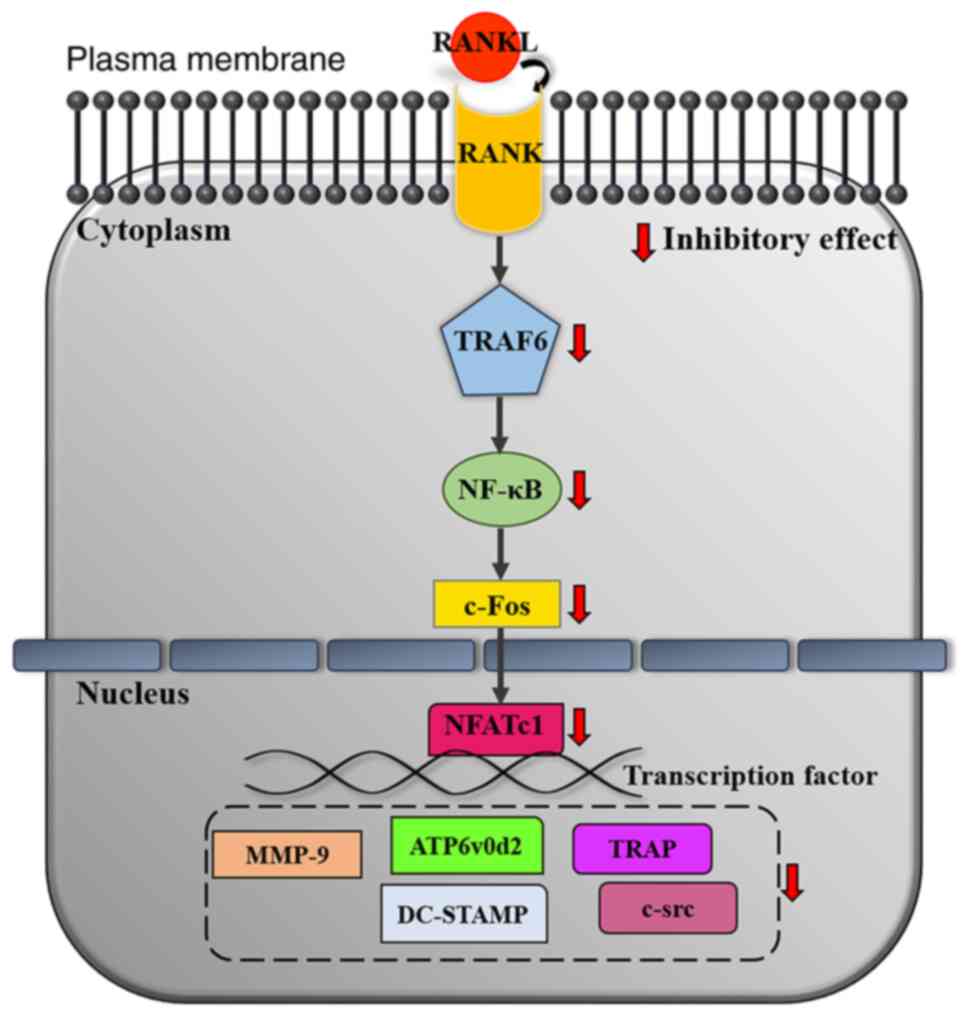 | Figure 10.Inhibitory mechanisms of MFR on
osteoclast differentiation. MFR, Melandrium firmum Rohrbach;
RANKL, receptor activator of nuclear factor-κB ligand; RANK,
receptor activator of nuclear factor-κB; TRAF6, tumor necrosis
factor receptor-associated factor 6; NF-κB, nuclear factor-κB;
NFATc1, nuclear factor of activated T cells, cytoplasmic 1; MMP-9,
matrix metalloproteinase-9; ATP6v0d2, ATPase H+ transporting V0
subunit D2; DC-STAMP, dendritic cell-specific transmembrane
protein; TRAP, tartrate-resistant acid phosphatase. |
Murine macrophage RAW 264.7 cells have a
monocyte/macrophage like cell lineage; they are derived from BALB/c
mice (44) and are suitable as a
cell experimental model in measuring osteoclast differentiation
(45). TRAP staining is the
standard method used to detect osteoclast formation and levels
(31,46,47).
In the present study, MFR suppressed RANKL-induced TRAP-positive
cells and TRAP levels. The F-actin ring is the cytoskeletal
structure that is required for formation of the sealing zone, and
permits the firm adhesion of mature osteoclasts to the bone surface
(48). The pit formation assay is
widely used to examine bone resorption and is an important
indicator of bone resorption from mature osteoclasts (48–50).
In the present study, MFR reduced the number and size of the
F-actin rings and the area of pit formation in RANKL-induced cells.
These results suggested that MFR may inhibit osteoclast formation
and bone resorption.
The RANK/RANKL signaling pathway has been reported
to induce the activation of TRAF6. TRAF6 deficiency in mice is
known to induce severe osteopetrosis and impairment in osteoclast
function (51–53). Activated TRAF6 can stimulate c-Fos,
which serves a crucial role in the induction of transcription
factors, including NFATc1 (54–56).
In a previous study, c-Fos-knockout mice developed osteopetrosis
due to attenuated osteoclast function (20). In addition, NFATc1-deficient adult
mice exhibited reduced bone loss in the absence of osteoclast
activity (17,19). In the present study, MFR was shown
to reduce the protein and mRNA expression levels of c-Fos and
NFATc1. These results suggested that MFR may decrease the
production of osteoclast differentiation, bone resorption and
F-actin ring formation by suppressing NFATc1 and c-Fos signaling
pathways. However, in contrast to the results regarding
NFATc1/c-Fos, the present study revealed that MFR significantly
inhibited TRAF6 at low concentrations but not at high
concentrations. The reason for the difference in the expression
levels of each factor can be inferred due to the following reasons.
RANKL binds to RANK, increases the expression and ubiquitination of
TRAF6, and leads to the accumulation of the TRAF6/transforming
growth factor-β-activated kinase 1 (TAK1) complex, which activates
sub-signaling molecules, such as phosphoinositide 3-kinases, MAPK
and NF-κB (57,58). It was hypothesized that treatment
with a high concentration of MFR in the present study could inhibit
the ubiquitination of TRAF6 or the accumulation of the TRAF6/TAK1
complex, rather than affecting the expression of TRAF6. However,
additional research needs to be conducted to assess this
hypothesis.
NFATc1 regulates several important
osteoclast-related genes, such as Mmp9, Ctsk and Ca2,
which are markers of bone resorption (17). Mmp9 and Ctsk are
well-known as bone resorption enzymes. Notably, Mmp9 is
expressed during the transformation of early osteoclasts to mature
osteoclasts (59) and deficiency of
Ctsk leads to impairments in bone resorption in mice and
humans (60). Furthermore,
Ca2 is an enzyme that is upregulated by c-Fos signaling, and
is expressed in the early stages of osteoclast differentiation and
influences the acidity of the bone surface (61). In the present study, MFR suppressed
the expression levels of mmp9, ctsk and ca2. These
results suggested that MFR may possess anti-osteoporotic activity
by regulating the expression of bone resorption markers.
Atp6vod2 and Dcstamp serve an important role in
cell-cell fusion during RANKL-stimulated osteoclast
differentiation. In addition, Atp6vod2 and Dcstamp
are important for F-actin ring formation (39,62).
Notably, gene-knockout mice of Atp6vod2 and Dcstamp
have been reported to develop osteopetrosis due to defects in
osteoclastogenesis (62,63). In the present study, MFR suppressed
the expression levels of Atp6vod2 and Dcstamp. These
results indicated that the inhibitory effect of MFR on F-actin ring
formation may be associated with the suppression of Atp6vod2
and Dcstamp. Oscar serves an important role in the
bone-specific regulation of osteoclast differentiation, and
activation of Oscar results in differentiation of early
osteoclasts to mature osteoclasts. In addition, Oscar can
induce calcium activation, resulting in increased expression of
NFATc1 (64). Src signaling
is important in regulating the osteoclast cytoskeleton (65). Prdm1 serves an essential role
in the differentiation and functions of macrophages and lymphocytes
(66). In addition, Prdm1
controls osteoclast development and bone homeostasis (67). In the present study, MFR decreased
the expression levels of Oscar, Src and Prdm1. These
results suggested that MFR may inhibit the expression of
osteoclast-related genes by suppressing NFATc1 signaling. Notably,
several genes were expressed in the form of two bands in the
present study; as a result of comparing the primer size using a DNA
ladder marker, the band at the top is considered the target, and
the bottom is inferred as the dimer remaining after the reaction
was completed.
Osteoporosis is the most common type of bone disease
worldwide, which is characterized by a low bone mass (68). Estrogen deficiency is the primary
cause of postmenopausal osteoporosis (69). Rats with ovariectomy-induced
osteoporosis have been used as a postmenopausal osteoporosis model
and share clinical characteristics with human osteoporosis
(3). It has been established that
an increase in body weight is a symptom observed in OVX rats. In
addition, reduction in uterus weight is indicative of successful
establishment (70). In the present
study, the body weight of animals was significantly increased 3
weeks after OVX and uterus weight was significantly decreased in
the OVX group compared with that in the sham group. These results
suggested that a postmenopausal osteoporosis model was successfully
established.
ALP is a marker commonly associated with bone
formation, which is produced during the early differentiation of
osteoblasts (71). Wu et al
(72) demonstrated that ALP
activity was increased in estrogen-deficient mice. Excessive
osteoclast activity causes an imbalance in bone metabolism,
resulting in increased osteoblast activity and ALP expression. In
the present study, the serum ALP levels were significantly
increased in the OVX group compared with those in the sham group,
whereas ALP levels were significantly decreased in the MFR-L group.
These results suggested that the MFR-L group reduced the levels of
ALP, which were increased due to excessive osteoclast activity. AST
and ALT are the most commonly used factors for assessment of
hepatotoxicity (73). The results
of the present study suggested that the levels of AST and ALT were
not affected in OVX rats. In addition, MFR-L, MFR-H, E2
and ALN did not exert hepatotoxic effects. CTX is the most widely
used indicator to measure bone resorption, and CTX-1 and TRAP are
bone resorption markers of osteoclasts that have been reported to
be increased in OVX models (74).
In the present study, CTX-1 levels were significantly increased in
the OVX group, whereas they were reduced in the MFR-L,
E2 and ALN groups; CTX-1 levels in the MFR-H group were
unchanged. In addition, TRAP levels were increased in the OVX
group; however, this difference was not significant. By contrast,
TRAP levels were reduced in the MFR-L, E2 and ALN
groups; TRAP levels were also reduced in the MFR-H group, but this
change was not significant.
Micro-CT has been used to study bone tissues and in
orthopedic studies (75). The
advantage of micro-CT is the lack of damage to the sample and ease
of reconstruction using image sections (76). BMD refers to the amount of mineral
content in bone tissue and is indicative of the strength of bones;
BMD is based on calcium content and is an important measure in the
evaluation of the OVX rat model (77). In the present study, the BMD in the
OVX group was significantly decreased compared with that in the
sham group, whereas BMD loss was reduced in the MFR-L,
E2 and ALN groups. Tb.Th and Tb.sp are indicators used
in assessment of the 3D image structure of cancellous bone
(78). In the present study, the
results revealed that the OVX group exhibited decreased Tb.Th and
increased Tb.sp compared with those in the sham group. Tb.Th was
increased following ALN treatment compared with that in the OVX
group, but no significant effect was observed on Tb.Th in the OVX
rats treated with MFR-L, MFR-H and E2. Tb.sp was
decreased in the E2 and ALN groups compared with that in
the OVX group, but no significant effect was observed in the MFR-L
and MFR-H groups. These findings indicated that oral administration
of MFR-L reduced bone loss.
Decreased trabecular area is a symptom commonly
observed in patients with osteoporosis and is the primary cause of
an increased risk of fracture; thus, the trabecular area has been
used as an important index of anti-osteoporotic activity in a
previous study (79). H&E
staining can also be used to assess the trabecular area in the
femur alongside micro-CT (80–84).
Bone loss of the femoral head was enhanced in the OVX rats, whereas
MFR-L, E2 and ALN reduced trabecular area loss in the
OVX-induced rat models. These results suggest that MFR-L,
E2 and ALN inhibited OVX-induced bone loss. IHC
demonstrated that the expression levels of NFATc1 were decreased
following treatment with MFR-L and E2. In addition,
administration of MFR-L, MFR-H, E2 and ALN reduced the
expression levels of CTK in the OVX rats.
The present study confirmed the effect of MFR on
osteoblast differentiation, which has an important role in bone
metabolism. MFR (12.5 and 25 µg/ml) promoted osteoblast
differentiation, whereas MFR (50 and 100 µg/ml) suppressed
osteoblast differentiation. These results indicated that high
concentrations of MFR can simultaneously inhibit osteoclast and
osteoblast differentiation. These results may be related to the
insignificant effect of the MFR-H group compared to the MFR-L in
bone loss in the OVX-induced rat model.
In conclusion, the effects of MFR on RANKL-induced
osteoclast differentiation and ovariectomy-induced bone loss in a
SD-rat model were determined. In vitro, MFR reduced
RANKL-induced osteoclast differentiation, function and formation.
MFR downregulated the expression levels of master transcription
factors, such as NFATc1 and c-Fos. In addition, MFR reduced the
expression levels of MMP-9, CTK, CA2, TRAP, ATP6v0d2, DC-STAMP,
OSCAR, c-Src and Blimp-1 through the downregulation of NFATc1 and
c-Fos signaling. In vivo, MFR-L increased BMD in the
OVX-induced bone loss model. MFR-H exhibited insignificant effects
on the OVX-induced bone loss model compared with MFR-L.
The present study had several limitations, as
follows: i) In vitro, MFR inhibited RANKL-induced expression
of NFATc1 and c-Fos signaling. MAPK and NF-κB signaling pathways
are also involved in NFATc1 and c-Fos signaling; however, the
effect of MFR was not determined on these factors. To further
assess the mechanisms underlying the inhibitory effects of MFR on
osteoclast differentiation via the NFATc1 and c-Fos pathway,
further studies regarding the involvement of MAPKs and NF-κB are
required. ii) When comparing the pharmacological effects of MFR in
the in vitro experiments and in vivo experiments, MFR
showed a relatively low pharmacological effect in in vivo
experiments compared with that in the in vitro experiments.
Therefore, further research on the effects of MFR on
osteoclast-mediated bone diseases, such as in the LPS-induced bone
loss model, senile osteoporosis and osteoporosis caused by steroid
side effects, are necessary. iii) In the micro-CT test results,
MFR-L increased BMD; however, it did not significantly affect the
bone micro-architecture. In general, an increase in BMD is
associated with changes in the bone micro-architecture; however,
this is not the case in this study. Further studies are required to
determine the reason behind this. iv) The present study focused on
the effects of MFR on inhibition of osteoclast differentiation;
According to previous studies, MFR contains a variety of components
(85), and constituents of MFR,
such as vitexin, linarin, ecdysterone and ursolic acid, were found
to be effective in inhibiting osteoclast differentiation. Vitexin
inhibited osteoclast differentiation and osteolysis (28), and Wang et al (84) revealed the inhibitory activity of
linarin on osteoclastogenesis through the RANKL-induced NF-κB
pathway. In addition, ecdysterone, another active compound in MFR,
prevented LPS-induced osteoclastogenesis (86). Ursolic acid has also been shown to
inhibit osteoclastogenesis and titanium particle-induced osteolysis
(87). However, to the best of our
knowledge, the effects of most of the components of MFR on
osteoclast differentiation have not been studied. Therefore,
analyzing the effect of each component of MFR on osteoclast
differentiation will help to understand the underlying
anti-osteoporotic mechanism of MFR.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This work was supported by the National Research
Foundation of Korea grant funded by the Korean government (grant
nos. 2019R1H1A2101224 and 2020R1A2C1007836).
Availability of data and materials
All data generated or analyzed during this study
are included in this published article.
Authors' contributions
YS designed the study. MK and BK prepared the
extract. MK, JHK and EYK performed the in vitro experiments
and analyzed the data. MK, JHK, SH and BK performed the in
vivo experiments and analyzed the data. MK, JHK and HSJ
contributed to the statistical analysis and helped interpret the
results. YS supervised the experiments in discussion with JHK and
MK. MK and JHK wrote the manuscript. MK and JHK confirm the
authenticity of all the raw data. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The protocols for this experiment were approved by
the Kyung Hee Medical Science Research Institute Animal Care and
Use Committee (approval no. KHMC-IACUC 19-017).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Matsuo K and Irie N: Osteoclast-osteoblast
communication. Arch Biochem Biophys. 473:201–209. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Riggs BL and Melton LJ III: The worldwide
problem of osteoporosis: Insights afforded by epidemiology. Bone.
17 (Suppl 5):505S–511S. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sözen T, Özışık L and Başaran NÇ: An
overview and management of osteoporosis. Eur J Rheumatol. 4:46–56.
2017. View Article : Google Scholar
|
|
4
|
Feng X and McDonald JM: Disorders of bone
remodeling. Annu Rev Pathol. 6:121–145. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tella SH and Gallagher JC: Prevention and
treatment of postmenopausal osteoporosis. J Steroid Biochem Mol
Biol. 142:155–170. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Khosla S, Burr D, Cauley J, Dempster DW,
Ebeling PR, Felsenberg D, Gagel RF, Gilsanz V, Guise T, Koka S, et
al: Bisphosphonate-associated osteonecrosis of the jaw: Report of a
task force of the American society for bone and mineral research. J
Bone Miner Res. 22:1479–1491. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wysowski DK: Reports of esophageal cancer
with oral bisphosphonate use. N Engl J Med. 360:89–90. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kennel KA and Drake MT: Adverse effects of
bisphosphonates: Implications for osteoporosis management. Mayo
Clin Proc. 84:632–638. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lenart BA, Lorich DG and Lane JM: Atypical
fractures of the femoral diaphysis in postmenopausal women taking
alendronate. N Engl J Med. 358:1304–1306. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Manson JE, Hsia J, Johnson KC, Rossouw JE,
Assaf AR, Lasser NL, Trevisan M, Black HR, Heckbert SR, Detrano R,
et al: Estrogen plus progestin and the risk of coronary heart
disease. N Engl J Med. 349:523–534. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Collaborative Group On Epidemiological
Studies Of Ovarian Cancer, ; Beral V, Gaitskell K, Hermon C, Moser
K, Reeves G and Peto R: Menopausal hormone use and ovarian cancer
risk: Individual participant meta-analysis of 52 epidemiological
studies. Lancet. 385:1835–1842. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Puhalla S, Bhattacharya S and Davidson NE:
Hormonal therapy in breast cancer: A model disease for the
personalization of cancer care. Mol Oncol. 6:222–236. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Collin-Osdoby P, Yu X, Zheng H and Osdoby
P: RANKL-mediated osteoclast formation from murine RAW 264.7 cells.
Methods Mol Biol. 80:153–166. 2003.PubMed/NCBI
|
|
14
|
Suda T, Takahashi N, Udagawa N, Jimi E,
Gillespie MT and Martin TJ: Modulation of osteoclast
differentiation and function by the new members of the tumor
necrosis factor receptor and ligand families. Endocr Rev.
20:345–357. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ye H, Arron JR, Lamothe B, Cirilli M,
Kobayashi T, Shevde NK, Segal D, Dzivenu OK, Vologodskaia M, Yim M,
et al: Distinct molecular mechanism for initiating TRAF6
signalling. Nature. 418:443–447. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Galibert L, Tometsko ME, Anderson DM,
Cosman D and Dougall WC: The involvement of multiple tumor necrosis
factor receptor (TNFR)-associated factors in the signaling
mechanisms of receptor activator of NF-kappaB, a member of the TNFR
superfamily. J Biol Chem. 273:34120–34127. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Roy A, Kim YB, Cho KH and Kim JH: Glucose
starvation-induced turnover of the yeast glucose transporter Hxt1.
Biochim Biophys Acta. 1840:2878–2885. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Boyle WJ, Simonet WS and Lacey DL:
Osteoclast differentiation and activation. Nature. 423:337–342.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Takayanagi H, Kim S, Koga T, Nishina H,
Isshiki M, Yoshida H, Saiura A, Isobe M, Yokochi T, Inoue J, et al:
Induction and activation of the transcription factor NFATc1 (NFAT2)
integrate RANKL signaling in terminal differentiation of
osteoclasts. Dev Cell. 3:889–901. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Arai A, Mizoguchi T, Harada S, Kobayashi
Y, Nakamichi Y, Yasuda H, Penninger JM, Yamada K, Udagawa N and
Takahashi N: Fos plays an essential role in the upregulation of
RANK expression in osteoclast precursors within the bone
microenvironment. J Cell Sci. 125:2910–2917. 2012.PubMed/NCBI
|
|
21
|
Perry LM and Metzger J: Medicinal plants
of east and southeast Asia: Attributed properties and uses.
Cambridge: MIT Press; 1980
|
|
22
|
Jeong YH, Oh YC, Cho WK, Lee B and Ma JY:
Anti-inflammatory effects of melandrii herba ethanol extract via
inhibition of NF-κB and MAPK signaling pathways and induction of
HO-1 in RAW 264.7 cells and mouse primary macrophages. Molecules.
21:8182016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Herbology editorial committee of Korean
medicine schools. Herbology. Seoul, . Yeonglimsa. 306–308.
2012.
|
|
24
|
Redlich K and Smolen JS: Inflammatory bone
loss: Pathogenesis and therapeutic intervention. Nat Rev Drug
Discov. 11:234–250. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lacativa PG and Farias ML: Osteoporosis
and inflammation. Arq Bras Endocrinol Metabol. 54:123–132. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
McLean RR: Proinflammatory cytokines and
osteoporosis. Curr Osteoporos Rep. 7:134–139. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ruscitti P, Cipriani P, Carubbi F,
Liakouli V, Zazzeroni F, Di Benedetto P, Berardicurti O, Alesse E
and Giacomelli R: The role of IL-1β in the bone loss during
rheumatic diseases. Mediators Inflamm. 2015:7823822015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jiang J, Jia Y, Lu X, Zhang T, Zhao K, Fu
Z, Pang C and Qian Y: Vitexin suppresses RANKL-induced
osteoclastogenesis and prevents lipopolysaccharide (LPS)-induced
osteolysis. J Cell Physiol. 234:17549–17560. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lee MY, Shin IS, Seo CS, Lee NH, Ha HK,
Son JK and Shin HK: Effects of Melandrium firmum methanolic
extract on testosterone-induced benign prostatic hyperplasia in
Wistar rats. Asian J Androl. 14:320–324. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Taciak B, Bialasek M, Braniewska A, Sas Z,
Sawicka P, Kiraga Ł, Rygiel T and Król M: Evaluation of phenotypic
and functional stability of RAW 264.7 cell line through serial
passages. PLoS One. 13:e01989432018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ballanti P, Minisola S, Pacitti MT,
Scarnecchia L, Rosso R, Mazzuoli GF and Bonucci E:
Tartrate-resistant acid phosphate activity as osteoclastic marker:
Sensitivity of cytochemical assessment and serum assay in
comparison with standardized osteoclast histomorphometry.
Osteoporos Int. 7:39–43. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tschöp MH, Speakman JR, Arch JR, Auwerx J,
Brüning JC, Chan L, Eckel RH, Farese RV Jr, Galgani JE, Hambly C,
et al: A guide to analysis of mouse energy metabolism. Nat Methods.
9:57–63. 2011. View Article : Google Scholar
|
|
33
|
Kim JH, Kim M, Jung HS and Sohn Y:
Leonurus sibiricus L. ethanol extract promotes osteoblast
differentiation and inhibits osteoclast formation. Int J Mol Med.
44:913–926. 2019.PubMed/NCBI
|
|
34
|
Kim M, Kim M, Kim JH, Hong S, Kim DH, Kim
S, Kim EY, Jung HS and Sohn Y: Crataegus pinnatifida bunge inhibits
RANKL-induced osteoclast differentiation in RAW 264.7 cells and
prevents bone loss in an ovariectomized rat model. Evid Based
Complement Alternat Med. 2021:55215622021. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Beconi MG, Howland D, Park L, Lyons K,
Giuliano J, Dominguez C, Munoz-Sanjuan I and Pacifici R:
Pharmacokinetics of memantine in rats and mice. PLoS Curr.
3:RRN12912011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jurdic P, Saltel F, Chabadel A and
Destaing O: Podosome and sealing zone: Specificity of the
osteoclast model. Eur J Cell Biol. 85:195–202. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhao Q, Wang X, Liu Y, He A and Jia R:
NFATc1: Functions in osteoclasts. Int J Biochem Cell Biol.
42:576–579. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Grigoriadis AE, Wang ZQ, Cecchini MG,
Hofstetter W, Felix R, Fleisch HA and Wagner EF: c-Fos: A key
regulator of osteoclast-macrophage lineage determination and bone
remodeling. Science. 266:443–448. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kim K, Lee SH, Ha Kim J, Choi Y and Kim N:
NFATc1 induces osteoclast fusion via up-regulation of Atp6v0d2 and
the dendritic cell-specific transmembrane protein (DC-STAMP). Mol
Endocrinol. 22:176–185. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Sharp PE, La Regina MC and Suckow MA: The
laboratory rat. Boca Raton: CRC Press; 1998
|
|
41
|
Hansan KMM, Tamanna N and Haque MA:
Biochemical and histopathological profiling of Wistar rat treated
with Brassica napus as a supplementary feed. Food Sci Hum Well.
7:77–82. 2018. View Article : Google Scholar
|
|
42
|
Sudo H, Kodama HA, Amagai Y, Yamamoto S
and Kasai S: In vitro differentiation and calcification in a new
clonal osteogenic cell line derived from newborn mouse calvaria. J
Cell Biol. 96:191–198. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Gregory CA, Gunn WG, Peister A and Prockop
DJ: An Alizarin red-based assay of mineralization by adherent cells
in culture: Comparison with cetylpyridinium chloride extraction.
Anal Biochem. 329:77–84. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hartley JW, Evans LH, Green KY, Naghashfar
Z, Macias AR, Zerfas PM and Ward JM: Expression of infectious
murine leukemia viruses by RAW264.7 cells, a potential complication
for studies with a widely used mouse macrophage cell line.
Retrovirology. 5:12008. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Collin-Osdoby P and Osdoby P:
RANKL-mediated osteoclast formation from murine RAW 264.7 cells.
Methods Mol Biol. 816:187–202. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kirstein B, Chambers TJ and Fuller K:
Secretion of tartrate-resistant acid phosphatase by osteoclasts
correlates with resorptive behavior. J Cell Biochem. 98:1085–1094.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hayman AR: Tartrate-resistant acid
phosphatase (TRAP) and the osteoclast/immune cell dichotomy.
Autoimmunity. 41:218–223. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Marchisio PC, Cirillo D, Naldini L,
Primavera MV, Teti A and Zambonin-Zallone A: Cell-substratum
interaction of cultured avian osteoclasts is mediated by specific
adhesion structures. J Cell Biol. 99:1696–1705. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Hsu H, Lacey DL, Dunstan CR, Solovyev I,
Colombero A, Timms E, Tan HL, Elliott G, Kelley MJ, Sarosi I, et
al: Tumor necrosis factor receptor family member RANK mediates
osteoclast differentiation and activation induced by
osteoprotegerin ligand. Proc Natl Acad Sci USA. 96:3540–3545. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Jimi E, Akiyama S, Tsurukai T, Okahashi N,
Kobayashi K, Udagawa N, Nishihara T, Takahashi N and Suda T:
Osteoclast differentiation factor acts as a multifunctional
regulator in murine osteoclast differentiation and function. J
Immunol. 163:434–442. 1999.PubMed/NCBI
|
|
51
|
Naito A, Azuma S, Tanaka S, Miyazaki T,
Takaki S, Takatsu K, Nakao K, Nakamura K, Katsuki M, Yamamoto T and
Inoue J: Severe osteopetrosis, defective interleukin-1 signalling
and lymph node organogenesis in TRAF6-deficient mice. Genes Cells.
4:353–362. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Wong BR, Josien R, Lee SY, Vologodskaia M,
Steinman RM and Choi Y: The TRAF family of signal transducers
mediates NF-kappaB activation by the TRANCE receptor. J Biol Chem.
273:28355–28359. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Hayden MS and Ghosh S: Shared principles
in NF-kappaB signaling. Cell. 132:344–362. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Yamashita T, Yao Z, Li F, Zhang Q, Badell
IR, Schwarz EM, Takeshita S, Wagner EF, Noda M, Matsuo K, et al:
NF-kappaB p50 and p52 regulate receptor activator of NF-kappaB
ligand (RANKL) and tumor necrosis factor-induced osteoclast
precursor differentiation by activating c-Fos and NFATc1. J Biol
Chem. 282:18245–18253. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Ray N, Kuwahara M, Takada Y, Maruyama K,
Kawaguchi T, Tsubone H, Ishikawa H and Matsuo K: c-Fos suppresses
systemic inflammatory response to endotoxin. Int Immunol.
18:671–677. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Fujioka S, Niu J, Schmidt C, Sclabas GM,
Peng B, Uwagawa T, Li Z, Evans DB, Abbruzzese JL and Chiao PJ:
NF-kappaB and AP-1 connection: Mechanism of NF-kappaB-dependent
regulation of AP-1 activity. Mol Cell Biol. 24:7806–7819. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Landström M: The TAK1-TRAF6 signalling
pathway. Int J Biochem Cell Biol. 42:585–589. 2010. View Article : Google Scholar
|
|
58
|
Wei ZF, Tong B, Xia YF, Lu Q, Chou GX,
Wang ZT and Dai Y: Norisoboldine suppresses osteoclast
differentiation through preventing the accumulation of TRAF6-TAK1
complexes and activation of MAPKs/NF-κB/c-Fos/NFATc1 pathways. PLoS
One. 8:e591712013. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Sundaram K, Nishimura R, Senn J, Youssef
RF, London SD and Reddy SV: RANK ligand signaling modulates the
matrix metalloproteinase-9 gene expression during osteoclast
differentiation. Exp Cell Res. 313:168–178. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Troen BR: The role of cathepsin K in
normal bone resorption. Drug News Perspect. 17:19–28. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
David JP, Rincon M, Neff L, Horne WC and
Baron R: Carbonic anhydrase II is an AP-1 target gene in
osteoclasts. J Cell Physiol. 188:89–97. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Yagi M, Miyamoto T, Sawatani Y, Iwamoto K,
Hosogane N, Fujita N, Morita K, Ninomiya K, Suzuki T, Miyamoto K,
et al: DC-STAMP is essential for cell-cell fusion in osteoclasts
and foreign body giant cells. J Exp Med. 202:345–351. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Lee SH, Rho J, Jeong D, Sul JY, Kim T, Kim
N, Kang JS, Miyamoto T, Suda T, Lee SK, et al: v-ATPase V0 subunit
d2-deficient mice exhibit impaired osteoclast fusion and increased
bone formation. Nat Med. 12:1403–1409. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Kim JH, Kim K, Jin HM, Youn BU, Song I,
Choi HS and Kim N: Upstream stimulatory factors regulate OSCAR gene
expression in RANKL-mediated osteoclast differentiation. J Mol
Biol. 383:502–511. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Miyazaki T, Tanaka S, Sanjay A and Baron
R: The role of c-Src kinase in the regulation of osteoclast
function. Mod Rheumatol. 16:68–74. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Nishikawa K, Nakashima T, Hayashi M,
Fukunaga T, Kato S, Kodama T, Takahashi S, Calame K and Takayanagi
H: Blimp1-mediated repression of negative regulators is required
for osteoclast differentiation. Proc Natl Acad Sci USA.
107:3117–3122. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Miyauchi Y, Ninomiya K, Miyamoto H,
Sakamoto A, Iwasaki R, Hoshi H, Miyamoto K, Hao W, Yoshida S,
Morioka H, et al: The Blimp1-Bcl6 axis is critical to regulate
osteoclast differentiation and bone homeostasis. J Exp Med.
207:751–762. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Eastell R, O'Neill TW, Hofbauer LC,
Langdahl B, Reid IR, Gold DT and Cummings SR: Postmenopausal
osteoporosis. Nat Rev Dis Primers. 2:160692016. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Riggs BL, Khosla S and Melton LJ III: A
unitary model for involutional osteoporosis: Estrogen deficiency
causes both type I and II osteoporosis in postmenopausal women and
contributes to bone loss in aging men. J Bone Miner Res.
13:763–773. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Kalu DN: The ovariectomized rat model of
postmenopausal bone loss. Bone Miner. 15:175–191. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Kuo TR and Chen CH: Bone biomarker for the
clinical assessment of osteoporosis: Recent developments and future
perspectives. Biomark Res. 5(18)2017.PubMed/NCBI
|
|
72
|
Wu X, Xie CQ, Zhu QQ, Wang MY, Sun B,
Huang YP, Shen C, An MF, Zhao YL, Wang XJ and Sheng J: Green tea
(Camellia sinensis) aqueous extract alleviates
postmenopausal osteoporosis in ovariectomized rats and prevents
RANKL-induced osteoclastogenesis in vitro. Food Nutr Res. 62:2018.
View Article : Google Scholar
|
|
73
|
Ozer J, Ratner M, Shaw M, Bailey W and
Schomaker S: The current state of serum biomarkers of
hepatotoxicity. Toxicology. 245:194–205. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Wronski TJ, Cintrón M and Dann LM:
Temporal relationship between bone loss and increased bone turnover
in ovariectomized rats. Calcif Tissue Int. 43:179–183. 1988.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Ito M: Recent progress in bone imaging for
osteoporosis research. J Bone Miner Metab. 29:131–140. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Bouxsein ML, Boyd SK, Christiansen BA,
Guldberg RE, Jepsen KJ and Müller R: Guidelines for assessment of
bone microstructure in rodents using micro-computed tomography. J
Bone Miner Res. 25:1468–1486. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Park SB, Lee YJ and Chung CK: Bone mineral
density changes after ovariectomy in rats as an osteopenic model:
Stepwise description of double dorso-lateral approach. J Korean
Neurosurg Soc. 48:309–312. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Parkinson IH and Fazzalari NL:
Interrelationships between structural parameters of cancellous bone
reveal accelerated structural change at low bone volume. J Bone
Miner Res. 18:2200–2205. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Osterhoff G, Morgan EF, Shefelbine SJ,
Karim L, McNamara LM and Augat P: Bone mechanical properties and
changes with osteoporosis. Injury. 47 (Suppl 2):S11–S20. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Kim EY, Kim JH, Kim M, Park JH, Sohn Y and
Jung HS: Abeliophyllum distichum Nakai alleviates postmenopausal
osteoporosis in ovariectomized rats and prevents RANKL-induced
osteoclastogenesis in vitro. J Ethnopharmacol. 257:1128282020.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Lee KY, Kim JH, Kim EY, Yeom M, Jung HS
and Sohn Y: Water extract of Cnidii Rhizoma suppresses
RANKL-induced osteoclastogenesis in RAW 264.7 cell by inhibiting
NFATc1/c-Fos signaling and prevents ovariectomized bone loss in
SD-rat. BMC Complement Altern Med. 19:2072019. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Kim M, Kim HS, Kim JH, Kim EY, Lee B, Lee
SY, Jun JY, Kim MB, Sohn Y and Jung HS: Chaenomelis fructus
inhibits osteoclast differentiation by suppressing NFATc1
expression and prevents ovariectomy-induced osteoporosis. BMC
Complement Med Ther. 20:352020. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Kim JH, Kim EY, Lee B, Min JH, Song DU,
Lim JM, Eom JW, Yeom M, Jung HS and Sohn Y: The effects of lycii
radicis cortex on RANKL-induced osteoclast differentiation and
activation in RAW 264.7 cells. Int J Mol Med. 37:649–658. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Wang J, Fu B, Lu F, Hu X, Tang J and Huang
L: Inhibitory activity of linarin on osteoclastogenesis through
receptor activator of nuclear factor kappaB ligand-induced
NF-kappaB pathway. Biochem Biophys Res Commun. 495:2133–2138. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Seo CS and Shin HK: Simultaneous
determination of the five marker compounds in melandriumfirmum
using high-performance liquid chromatography with photodiode-array
detection. Nat Prod Commun. 11:1667–1669. 2016.PubMed/NCBI
|
|
86
|
Li Y, Zhang J, Yan C, Chen Q, Xiang C,
Zhang Q, Wang X and Jiang K: β-Ecdysterone prevents LPS-induced
osteoclastogenesis by regulating NF-κB pathway in vitro. Res
Square. 2020.
|
|
87
|
Jiang C, Xiao F, Gu X, Zhai Z, Liu X, Wang
W, Tang T, Wang Y, Zhu Z, Dai K, et al: Inhibitory effects of
ursolic acid on osteoclastogenesis and titanium particle-induced
osteolysis are mediated primarily via suppression of NF-κB
signaling. Biochimie. 111:107–118. 2015. View Article : Google Scholar : PubMed/NCBI
|