The tumor microenvironment consists of several types
of stromal cells, including adipocytes, fibroblasts, endothelial
cells and macrophages (1). Imaging
evaluation has revealed that adipose tissue occupies 56% of
non-lactating breast tissue and 35% of lactating breast tissue
(2,3), suggesting that adipocytes account for
a major part of the breast tumor microenvironment. Adipocytes,
preadipocytes, fibroblasts, immune cells, endothelial cells and
extracellular matrix (ECM) are the main components of adipose
tissue, which is divided into white, brown and beige adipose tissue
(4,5). White adipocytes are the most abundant
fat cells in the human body, and previous findings have associated
white adipose tissue (WAT) with an increased risk of breast cancer
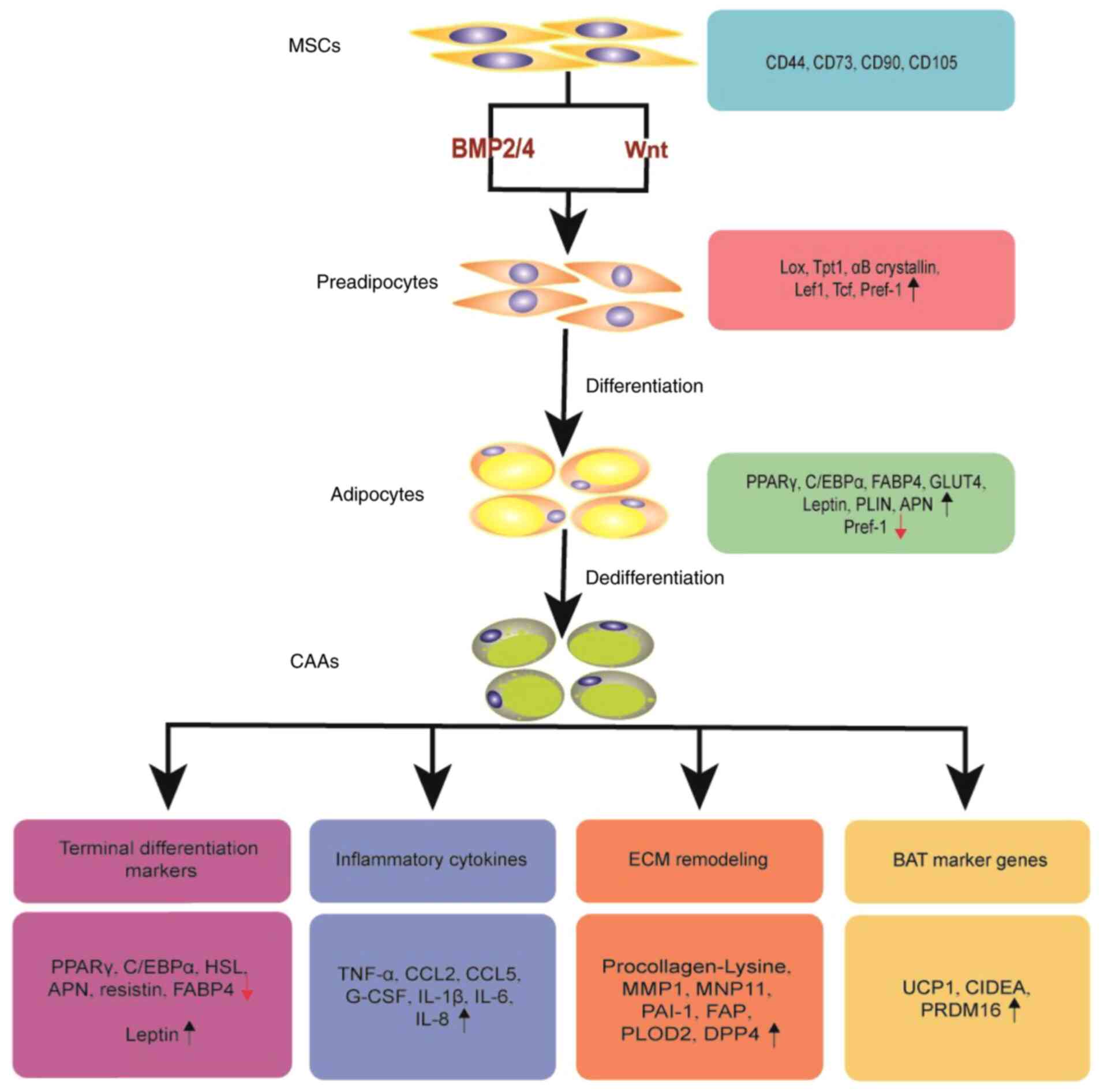
(6). In addition to serving as an
energy reservoir for triglycerides, adipose tissue is an active
endocrine organ that secretes hormones, adipokines, cytokines,
chemokines and proinflammatory molecules (7). Existing studies have confirmed the
hypothesis that adipocytes and cancer cells interact dynamically
(8–10), as opposed to adipocytes being
previously considered as static cells neighboring cancer cells.
Previous studies have mostly focused on the role of adipocytes in
the proliferation and migration of cancer cells, with little
attention to the changes in adipocytes (11). A particular class of adipocytes,
referred to as cancer-associated adipocytes (CAAs), have been
identified in the matrix surrounding invasive breast cancer
(12). Therefore, in addition to
summarizing the biological characteristics of CAAs, the aim of the
present review was to further focus on the underlying mechanisms
that contribute to CAA development, in the hope of providing new
perspectives for breast cancer treatment.
Cancer-adjacent adipocytes can be transformed into
CAAs after communicating with cancer cells, adopting a
dedifferentiation phenotype and secreting a significant number of
proinflammatory cytokines, thereby promoting malignant tumor
progression (12,33). The main characteristics of CAAs are
described below:
The expression of PPARγ and C/EBPα is significantly
inhibited, which results in a substantial reduction in the mRNA
levels of adipocyte-specific genes, including FABP4 and
hormone-sensitive lipase (HSL), resulting in dedifferentiation of
mature adipocytes (12,34). The reduction of lipid content and
cell size triggers the release of metabolites by CAAs, including
free fatty acids and ketone bodies, to facilitate tumor progression
(34–36). This effect was shown to be
significantly enhanced in a cell culture model of obesity (36). Furthermore, the expression and
activity of LPL and FAS in adipose tissue adjacent to the tumor
invasion front were found to be significantly decreased, and the
adipogenesis and fat storage capacity in patients with colorectal
cancer were impaired (37). These
changes may cause lipolysis in adipocytes, which then assume a
fibroblast-like morphology.
In CAAs, the expression of pro-tumorigenic
adipokines, such as leptin and resistin, is increased, while the
expression of the anti-tumorigenic adipokine, APN, is markedly
decreased (12). CAAs also exhibit
increased expression and secretion of proinflammatory cytokines,
such as IL-6, IL-1β and TNF-α (12,38),
and chemokines, such as C-C motif chemokine ligand (CCL)5, CCL2 and
IL-8 (also known as CXCL8), causing an increase in cancer invasive
and metastatic ability (39–41).
Our transcriptome sequencing data revealed that CAAs exhibited a
higher expression of granulocyte colony-stimulating factor, which
can promote the malignant progression of breast cancer via the
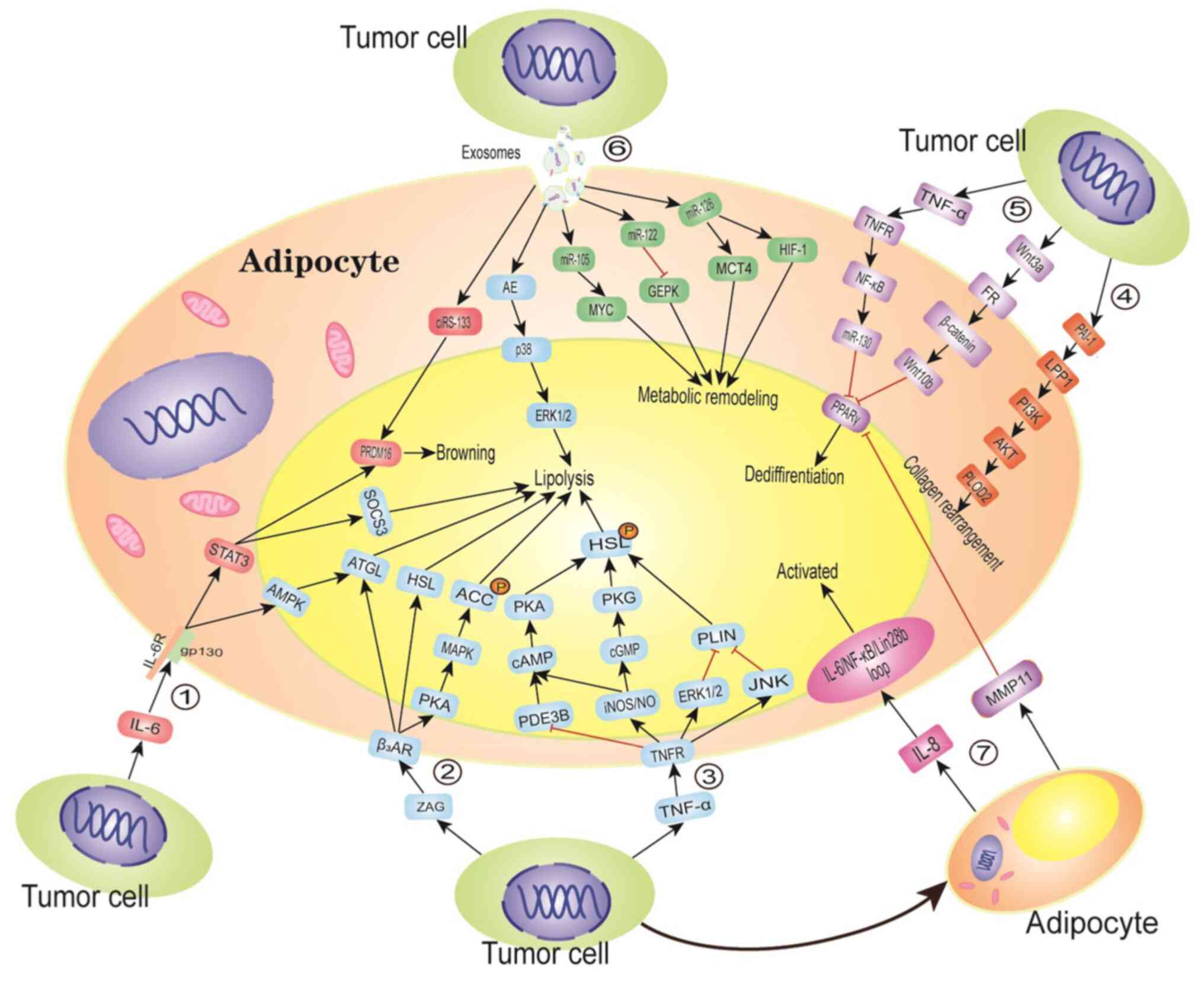
STAT3 pathway (Fig. 1) (42).
Tumors secrete numerous types of factors that induce
the transformation of adipocytes into CAAs. The factors secreted by
CAAs, such as IL-8 and autotaxin (ATX), can also maintain their
activated state and provide favorable conditions for tumor growth
and metastasis.
Tumor-derived TNF-α can promote the formation of
CAAs by selectively downregulating the expression of PPARγ and
C/EBPα (52). Stephens and Pekala
(53) demonstrated that culturing
differentiated 3T3-L1 adipocytes with TNF-α significantly inhibited
C/EBPα, GLUT4 and FABP4 gene expression in adipocytes.
Interestingly, TNF-α was found to significantly reduce the
stability of these mRNAs, which further prevented the accumulation
of GLUT4 and C/EBPα proteins. The marked reduction in C/EBPα
protein levels may lead to the enhancement of transcriptional
inhibition of C/EBPα, GLUT4 and FABP4 genes by TNF-α. In addition,
treatment with TNF-α activates the NF-κB pathway in adipocytes,
which further enhances the binding of p65 to the miR-130 promoter.
The upregulation of miR-130 eventually leads to a decrease in PPARγ
mRNA to preadipocyte levels (54).
Furthermore, by decreasing the terminal differentiation markers,
TNF-α increases lipolysis in WAT. Treatment with TNF-α was shown to
increase the levels of inducible nitric oxide synthase (iNOS) and
nitric oxide (NO) in multiple tissues, including adipose tissue,
activating cAMP/cGMP and stimulating HSL phosphorylation, resulting
in a significant increase in lipolysis (55). PLIN is a key regulator of HSL that
is located on the surface of intracellular triglyceride lipid
droplets. Exposing adipocytes to a TNF-α-rich tumor
microenvironment can downregulate PLIN expression to mediate
lipolysis via the ERK1/2 and JNK pathways (56). In addition, in TNF-α-treated
adipocytes, the expression of the cyclic nucleotide
phosphodiesterase 3B (PDE3B), a key hydrolase of insulin-activated
cAMP, was shown to decrease by 50%, resulting in an increase in
intracellular cAMP content and enhanced lipolysis (57–59).
The loss of terminal differentiation markers and the synergistic
effect of enhanced lipolysis promote the transition of mature
adipocytes from the adipocyte phenotype to the spindled
fibroblast-like phenotype.
The crosstalk between inflammatory signals, such as
IL-6, from tumor tissue, adipose tissue and other organs, can cause
energy imbalance and promote fat mobilization and catabolism
(60,61). Petersen et al (62) showed that IL-6 can directly act on
adipose tissue to enhance lipolysis in vivo and in
vitro. Tumor-derived IL-6 binds to glycoprotein 130 (gp130) to
enhance the adipose triglyceride lipase (ATGL)-stimulated lipolysis
cascade via the AMP-activated protein kinase (AMPK) pathway,
resulting in free fatty acids for breast cancer cells (50,63),
which then induce the migration and invasion of breast cancer cells
(64). Tsoli et al (50) also discovered that IL-6 can enhance
insulin signaling interference by upregulating the expression of
suppressor of cytokine signaling 3, which phosphorylates and
activates the STAT3 pathway, reducing glucose uptake and the
synthesis other lipid substrates by WAT, inhibiting lipid synthesis
and promoting the atrophy of WAT (65). Interestingly, WAT browning is known
to involve paracrine and neuroendocrine signaling, which can be
used by the transcription factor PRDM16 to induce the expression of
genes related to lipid utilization and thermogenesis, such as UCP1
(66). The STAT3 signaling pathway
can activate the PRDM16 gene, promoting the browning of WAT
(50,66). IL-6 also enhances sympathetic
activity, causing further stress-mediated release of IL-6 from BAT
(67). Adipocytes co-cultured with
tumor cells can increase the level of IL-6 by upregulating
mmu-miR-5112 and downregulating the expression of cytoplasmic
polyadenylation element-binding protein 1, a negative regulator of
IL-6 (68). In addition,
tumor-secreted leukemia inhibitory factor, a member of the IL-6
cytokine family, can activate the Janus kinase (JAK)/STAT pathway
through gp130 and induce ATGL activation to enhance lipolysis
(69).
CAAs expressing high levels of inflammatory
cytokines, adipokines and proteases are considered to be adipocytes
at an active state, as mentioned above. Compared to adipocytes at a
distance from the tumor invasion front, CAAs are characterized by a
high proliferation rate and strong invasive and migratory
abilities. Al-Khalaf et al (70) isolated CAAs and their adjacent
tumor-counterpart adipocytes (TCAs) from 10 patients with invasive
breast cancer and found that IL-8 plays a key role in maintaining
the activation of CAAs. It was also previously reported that all
CAAs secrete high levels of IL-8, which helps to induce the
migration of ovarian cancer cells to adipocyte-rich areas (34). In addition, the basic and active
forms of the AKT and STAT3 proteins, which are downstream effectors
of IL-8 signaling, are higher in CAAs compared with those in TCAs.
The ectopic expression of IL-8 in TCAs may cause their
self-activation. According to Hendrayani et al (71), the activation state of CAFs is
maintained via an IL-6/NF-κB/Lin28b positive feedback loop.
Consistently, treatment of CAAs with anti-IL-8 antibody decreases
the mRNA levels of IL-6, RelA and Lin28b, and inhibits the
activated state of CAAs. These results suggest that IL-8 may
maintain the activity of CAAs by activating the IL-6/NF-κB/Lin28b
positive feedback loop (70).
Breast cancer cells secrete Wnt3a, which activates
the Wnt/β-catenin pathway and induces the dedifferentiation of
mouse and human adipocytes, acquiring an ADF-like phenotype
(72). In adipocytes co-cultured
with breast cancer cells, Wan3a upregulates the expression of Wnt
target genes, such as dishevelled-1, Wnt10b, endothelin-1 and MMP7,
via a β-catenin-dependent pathway (45,73).
Wnt10b is known to block adipocyte differentiation by suppressing
PPARγ and C/EBPα expression. Treatment with ICG-001, a unique small
molecule that selectively inhibits Tcf/β-catenin transcription,
significantly inhibits the increase of Wnt10b expression, partially
restores the lipid accumulation and reverses the ADF phenotype
(45). Zoico et al (70) and Kang et al (71) demonstrated that pancreatic cancer
can induce adipocyte dedifferentiation through a Wnt5a-dependent
signaling pathway. The JAK/STAT3 pathway is activated by
adipocyte-derived inflammatory factors when pancreatic cancer cells
are co-cultured with adipocytes, leading to an increase in Wnt5a
expression (74). The inhibition of
adipogenesis by Wnt5a expressed by immune cells, such as
macrophages, is well known (75).
However, the opposite results have also been observed in other
types of cancer (76,77), which may be explained by the
different expression levels of Wnt5a isotypes. Wnt5a-long (L) can
inhibit tumors, whereas Wnt5a-short (S) can promote tumors
(77). The expression of Wnt5a-L
was found to be more pronounced in adipocytes co-cultured with
pancreatic cancer cells (Fig. 2)
(74).
MMP11, which is also known as stromelysin-3, is a
connective tissue-derived factor that is usually associated with
tumor invasion and poor prognosis (78). The expression of MMP11 in adipocytes
can be significantly increased during the crosstalk between cancer
cells and adipocytes (43).
Andarawewa et al (43) found
that MMP11 was expressed in adipocytes located adjacent to the
invasive breast cancer, but not in distally located adipocytes.
Northern blotting analysis demonstrated that the expression levels
of PPARγ and FABP4 in MMP11-deficient mice were higher compared
with those in wild-type mice with stronger adipocyte
differentiation potential, suggesting that MMP11 is a negative
physiological regulator of adipogenesis. In addition to reducing
the differentiation of preadipocytes, MMP11 induces the
dedifferentiation of mature adipocytes. Treatment with MMP11
decreases the number and size of lipid droplets in adipocytes,
reduces the number of adipocytes exhibiting a round phenotype, and
increases the number of fibroblast-like cells (43). Compared with FABP4, the expression
of PPARγ in MMP11-deficient tissues was found to be unregulated,
suggesting that MMP11 may impede adipogenesis by downregulating the
expression of PPARγ (43).
According to previous studies, MMP11 is expressed in fibroblasts
surrounding invasive cancer cells (79,80).
Interestingly, these fibroblasts do not express α-SMA. Therefore,
MMP11-expressing fibroblasts around the tumor may partially
transdifferentiate from adipocytes or preadipocytes. Motrescu et
al (81) further demonstrated
that MMP11 can specifically degrade the natural α3 chain of
collagen VI and change the ECM of tumors, unlike resting
adipocytes, which are surrounded by thin strands of collagen VI.
These results suggest that cancer cells can induce MMP11 expression
in adipocytes by secreting soluble substances or through direct
contact, resulting in an increased CAA phenotype and a high
proportion of tumor matrix fibroblasts (43).
PAI-1 is a mesenchymal marker that has been linked
to the progression of a variety of cancers. Overexpression of PAI-1
has been associated wit a poor prognosis of breast cancer,
particularly triple-negative breast cancer. The expression of PAI-1
is upregulated in both adipocytes and breast cancer cells after
co-culture, but breast cancer cells secrete five-fold higher PAI-1
amounts than adipose cells (47).
It has been revealed that PAI-1 mediates signal transduction by
binding to cell membrane receptors, such as LDL receptor-related
protein 1 (LRP-1) (82). In
addition, available data indicate that LRP-1 is highly expressed in
adipocytes and CAAs, and a high concentration of PAI-1 (200 ng/ml)
can upregulate the expression of LRP-1 in adipocytes (47). The LRP-1-dependent simulation of the
PI3K/AKT signaling pathway promotes the activation of PLOD2 (a
lysine hydroxylase gene) in CAAs, whereas knocking down PAI-1 in
MDA-MB-231 cells blocks the upregulation of PLOD2 expression in
CAAs, suggesting that PAI-1 is an important regulatory factor
(47). The activation of PLOD2 can
stimulate the rearrangement of adipocyte-derived collagen into a
linear structure and promote the metastasis of cancer cells along
the recombinant linear collagen fibers (47).
ATX is a secreted enzyme that catalyzes the
hydrolysis of LPC, the most abundant phospholipid in the plasma, to
produce vast amounts of lysophosphatidic acid (LPA) (83). LPA signaling is mediated by at least
six G protein-coupled receptors (LPA1-6) (84,85),
and it promotes cell survival, proliferation and migration, while
also controlling a variety of physiological and pathological
processes (86–88). Popnikolov et al (89) demonstrated that ATX is mainly
present in stromal cells. Adipose tissue is the main source of ATX
in the circulation. Therefore, obesity and a high-fat diet may
increase the production of ATX in adipocytes (90). In comparison to the surrounding
breast fat pad, a syngeneic breast cancer model revealed that 4T1
cells express negligible amounts of ATX (91). The activity of ATX can be inhibited
by LPA or sphingosine 1-phosphate-related lipids. Moreover, Benesch
et al (92) found that
inflammatory cytokines, such as TNF-α and IL-1β, produced by tumor
cells can overcome the LPA-mediated inhibitory effect on ATX mRNA
expression, suggesting that LPA and ATX may coexist in the tumor
microenvironment. The levels of ATX mRNA and protein in the adipose
tissue surrounding tumor tissues increase as breast cancer develops
(91). The increased LPA signaling
further promotes the production of inflammatory mediators in
adipose tissue and tumors (93).
Although the molecules implicated in the ATX pathway remain
unclear, evidence indicates that tumor-derived cytokines can
stimulate increased production of ATX in peripheral adipocytes,
promoting tumor cell proliferation, migration, metastasis, and
resistance to radiotherapy and chemotherapy in a paracrine manner
(84,86,90).
ZAG is a 43-kDa glycoprotein encoded by the AZGP-1
gene. Previous studies have shown that ZAG is produced by certain
cancer cells and WAT, and is closely associated with the prognosis
of several types of cancer (94,95),
including breast (96), colorectal
(97), prostate (98), and other cancers. Further research
has demonstrated that tumor-derived ZAG can regulate lipid
metabolism and promote WAT browning (99). Overexpression of ZAG suppresses the
expression of FAS, diacylglycerol acyltransferase and acetyl-CoA
carboxylase (ACC) in mouse adipose tissue (100). On the contrary, ZAG binds to β3
adrenoceptor (β3AR) and upregulates the expression of ATGL, HSL and
phosphorylated ACC via the protein kinase A (PKA)/p38 MAPK
signaling pathway, promoting lipolysis and inhibiting adipogenesis
(101). In 3T3-L1 preadipocytes,
ZAG was shown to suppress the differentiation of preadipocytes by
inhibiting the expression of PPARγ, C/EBPα and the adipogenic
enzyme FAS. According to Elattar et al (99), ZAG enhances the transcriptional
translation of PRDM16 by stimulating the expression of PPARγ and
early B cytokine 2. In addition, ZAG stimulates the expression of
PPARγ and PPARγ coactivator 1α, and promotes the recruitment of
PPARγ into the UCP1 promoter, leading to increased UCP1 expression
(99). Similarly, the
ZAG/β3AR/PKA/p38 MAPK signaling pathway can enhance the expression
of specific BAT markers (101).
Exosomes are small extracellular vesicles, with a
diameter of 30–100 nm, derived from endosomal multivesicular bodies
(MVBs). Exosomes are released in bursts when MVBs fuse with the
cell membrane, and their contents, such as miRNAs, other non-coding
RNAs, transcription factors, proteins and lipids, are transported
to target cells to participate in cell-cell communication (102). The miRNA characteristics of
exosomes are parallel to the miRNA expression profile of the tumor
cells (103), suggesting that the
miRNA expression pattern in body fluids, such as urine or saliva,
can be analyzed to replace traditional needle biopsy for early
diagnosis of breast cancer.
Breast cancer cell-derived exosomes transport
miRNAs, such as miR-144, miR-126 and miR-155, to resident cells in
the breast cancer microenvironment, inducing the formation of CAAs
(35). Previous studies have shown
that cancer cell-derived miR-144 can target the MAP3K8 gene and
reduce the phosphorylation level of ERK1/2 (35), resulting in the inhibition of PPARγ
S273 phosphorylation in adipocytes, increased expression of UCP1
and browning of WAT (35,104). Tumor-derived miR-126 can
downregulate the expression of GLUT4 in adipocytes by directly
targeting the insulin receptor substrate 1 gene. Furthermore,
miR-126 can activate AMPK and increase the protein levels of
hypoxia-inducible factor-1α and monocarboxylate transporter 4,
resulting in metabolic remodeling of adipocytes, which is
characterized by decreased glucose uptake and increased glycolysis
and secretion of metabolites such as lactic acid and pyruvate
(35,105). Breast cancer cell-derived miR-155
can promote the beiging/browning and metabolic remodeling of
adipocytes by downregulating the expression of PPARγ (51). In addition, higher levels of
circulating miR-122 have been associated with breast cancer
metastasis. Cancer cell-secreted miR-122 inhibits glucose uptake of
premetastatic niche cells by reducing the activity of the
glycolytic enzyme pyruvate kinase, which promotes the progression
of the disease (106). Similarly,
the expression of breast cancer-secreted miR-105 activates MYC
signaling in CAFs and CAAs, which allows them to fuel neighboring
cancer cells by enhancing glucose and glutamine metabolism under
adequate nutrient conditions (107). In addition, the liver cancer
secretes exosomes, which can activate the NF-κB signaling pathway
in adipocytes to promote tumor growth and angiogenesis, and recruit
additional macrophages (108).
Pancreatic cancer cells can transfer exosomal AE and induce
lipolysis of CAAs by activating the p38 and ERK1/2 pathways
(109,110). Advanced Lewis lung cancer can also
secrete exosomal IL-6, leading to skeletal muscle atrophy and
adipose tissue browning via activation of the IL-6/STAT3 pathway
(111). Gastric cancer cells can
transfer exosomal ciRS-133 (circRNA sponge for miR-133) to
preadipocytes and induce brown adipocyte differentiation by
activating PRDM16 and inhibiting the expression of miR-133
(109,112). Furthermore, adipocyte-derived
exosomal miR-21 can be transferred to tumor cells to promote their
proliferation, invasion and migration, drug resistance and
angiogenesis (113–115). Lazar et al (116) found that adipocyte-derived
exosomes can stimulate melanoma invasion and metastasis by fatty
acid oxidation. Adipocyte-derived exosomal MMP3 can also promote
lung cancer metastasis by increasing the activity of MMP9, which
adversely affects the prognosis of patients with lung cancer
(Fig. 2) (117).
Taken together, the aforementioned tumor-derived
cytokines or miRNAs target CAAs via a paracrine mechanism, and one
or more biological changes occur during the transition from mature
adipocytes to CAAs through the activation of the NF-κB, ERK,
β-catenin and other signaling pathways. Initially, tumor-derived
cytokines or miRNAs suppress the expression of the adipocyte
differentiation markers PPARγ and C/EBPα, resulting in adipocyte
lipolysis, increased expression of pro-inflammatory factors, ECM
remodeling and browning, until the adipocytes are finally
transformed into the unique CAAs.
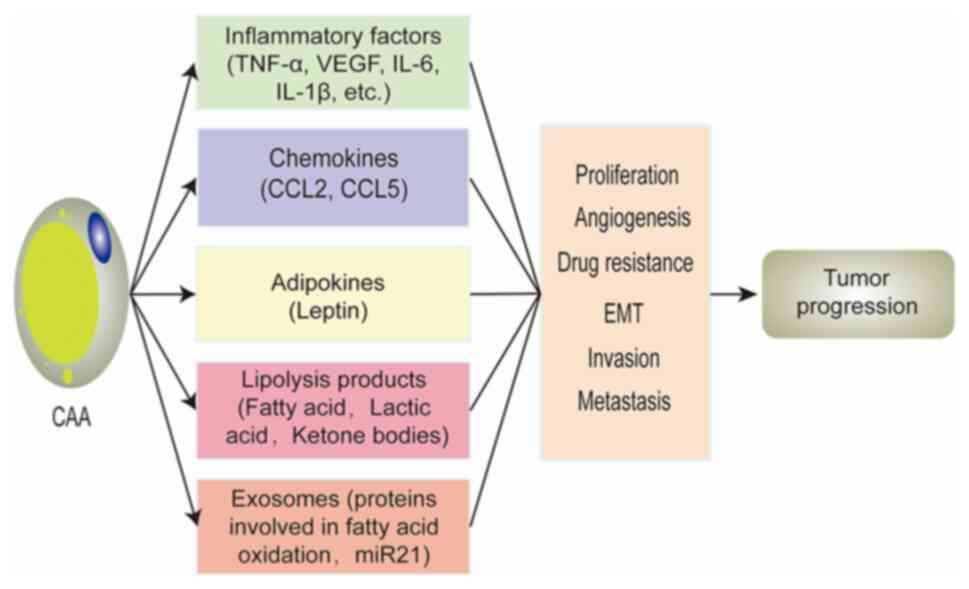
CAAs play a key role in breast cancer progression.
‘Activated’ CAAs can promote the progression of breast cancer
through regulating various aspects of tumor cell proliferation,
epithelial-to-mesenchymal transition (EMT), invasion, metastasis
and angiogenesis, by secreting chemokines (CCL2 and CCL5),
inflammatory factors (IL-6, IL-1β, TNF-α and VEGF), adipokines
(leptin), exosomes (proteins involved in fatty acid oxidation and
miR-21) and lipolysis products (fatty acids, lactic acid and ketone
bodies) (11). In a similar manner,
CAAs may contribute to the drug resistance of breast cancer.
Studies have reported that fatty acids can be used as a structural
unit for the synthesis of membrane phospholipids, which can change
the lateral and transverse membrane dynamics of breast cancer cells
to enhance their oxidative stress resistance and limit the drug
intake (118). Furthermore,
lipolysis products, such as fatty acids and lactate, profoundly
affect the homeostasis and differentiation of various immune cells,
promoting immune escape and tumor progression (119). Leptin can also activate NF-κB
signal transduction to reduce the toxicity of chemotherapeutics
in vitro (120). The
elevated expression of IL-6 in CAAs can induce the cancer stem
cell-positive phenotype by activating the STAT3/NF-κB pathway,
leading to chemotherapy resistance of HER2-positive breast cancer
(121). In CAAs, the upregulation
of the expression of matrix proteins, such as MMP11, MMP1 and
collagen VI, may also be involved in cisplatin resistance (Fig. 3) (38).
CAAs can create a favorable microenvironment for
tumor cell proliferation, invasion and migration, and communication
between CAAs and tumor cells can further affect the prognosis of
patients with cancer (11),
suggesting that CAAs may represent a potential therapeutic target
for breast cancer. However, the mechanisms underlying the formation
of CAAs have yet to be completely elucidated, and several problems
remain unresolved.
First, certain cytokines, such as TGF-β, strongly
reduce PPARγ and C/EBPα expression and induce fibrosis in
vivo (52). IL-11 and
angiopoietin-like 4 are highly expressed in tumor cells (122), inhibit the differentiation of
preadipocytes (123), or stimulate
lipolysis by increasing adipocyte cAMP levels (124). In addition to the miRNAs mentioned
above, Wu et al (35) also
discovered that several miRNAs, including miR-22, miR-210 and
miR-16, were found to be differentially expressed in MDA-MD-231
cell- and CAA-conditioned culture medium. However, whether those
cytokines and novel tumor-derived miRNAs are involved in the
formation of CAAs is unknown. HOX transcript antisense intergenic
RNA (HOTAIR), a long non-coding (lnc)RNA released by exosomes, is
highly expressed in a variety of tumor tissues (125,126). Interestingly, cancer cells
transfer exosomal HOTAIR to endothelial cells, resulting in
increased expression of VEGFA and angiogenesis (127). However, the role of lncRNAs
secreted by tumors in adipocytes requires further investigation in
the future.
Second, ADFs in the tumor center no longer contain
lipid droplets or express adipocyte markers, and CAFs have also
been found to overexpress UCP1 and MMP11 (80,128),
which makes ADFs indistinguishable from other CAFs, suggesting that
CAAs may constitute a part of the CAF population.
Third, adipocytes can be attracted to tumors and
some cancer cells preferentially migrate to sites rich in adipose
tissue, indicating the potential application of adipocytes as
cell-based delivery platforms for drugs (or prodrugs),
nanoparticles, or nucleic acids (129). However, in comparison, CAAs are
more aggressive and in an active state, exhibiting increased
expression of various inflammatory factors. CAAs can be cultured
and isolated in vitro, making them more suitable for
cell-based delivery platforms compared with normal mature
adipocytes.
Finally, CAAs can be transformed from an
‘accomplice’ to a ‘preventer’ of tumors, which is of higher
therapeutic value, by reversing their phenotype. However, due to
the lack of uniform standards for CAA protein markers and related
genes, it remains difficult to develop CAA-targeted treatments.
Therefore, an in-depth understanding of the formation of CAAs in
the breast cancer microenvironment should provide new insights into
the treatment of breast cancer.
Not applicable.
The present study was funded by grants from the
National Natural Science Foundation of China (grant no. 81760509)
and the Natural Science Foundation of Jiangxi Province of China
(grant no. 20181BAB205043).
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
XX, YT and WZ contributed to the concept and the
design of this review. YT, TS and WZ wrote the manuscript. XX, YT,
TS, WZ and XH helped draft the manuscript and drew the figures. XH
and XX provided significant suggestions for the study. YT, TS and
WZ searched the literature and collated important reference
information. WZ and XX critically reviewed the manuscript. All the
authors have read and approved the final manuscript and agree to be
accountable for all aspects of the work in ensuring that questions
related to the accuracy or integrity of any part of the work are
appropriately investigated and resolved.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Montel V, Mose ES and Tarin D:
Tumor-stromal interactions reciprocally modulate gene expression
patterns during carcinogenesis and metastasis. Int J Cancer.
119:251–263. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Vandeweyer E and Hertens D: Quantification
of glands and fat in breast tissue: An experimental determination.
Ann Anat. 184:181–184. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ramsay DT, Kent JC, Hartmann RA and
Hartmann PE: Anatomy of the lactating human breast redefined with
ultrasound imaging. J Anat. 206:525–534. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Luo L and Liu M: Adipose tissue in control
of metabolism. J Endocrinol. 231:R77–R99. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang YX, Zhu N, Zhang CJ, Wang YK, Wu HT,
Li Q, Du K, Liao DF and Qin L: Friend or foe: Multiple roles of
adipose tissue in cancer formation and progression. J Cell Physiol.
234:21436–21449. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Iyengar NM, Zhou XK, Mendieta H, Giri DD,
El-Hely O, Winston L, Falcone DJ, Wang H, Meng L, Landa J, et al:
Effects of adiposity and exercise on breast tissue and systemic
metabo-inflammatory factors in women at high risk or diagnosed with
breast cancer. Cancer Prev Res (Phila). 14:541–550. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nieman KM, Romero IL, Van Houten B and
Lengyel E: Adipose tissue and adipocytes support tumorigenesis and
metastasis. Biochim Biophys Acta. 1831:1533–1541. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhao C, Wu M, Zeng N, Xiong M, Hu W, Lv W,
Yi Y, Zhang Q and Wu Y: Cancer-associated adipocytes: Emerging
supporters in breast cancer. J Exp Clin Cancer Res. 39:1562020.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Uehara H, Kobayashi T, Matsumoto M,
Watanabe S, Yoneda A and Bando Y: Adipose tissue: Critical
contributor to the development of prostate cancer. J Med Invest.
65:9–17. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Picon-Ruiz M, Marchal JA and Slingerland
JM: Obtaining human breast adipose cells for breast cancer cell
co-culture studies. STAR Protoc. 1:1001972020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu Q, Li B, Li Z, Li J and Sun S and Sun
S: Cancer-associated adipocytes: Key players in breast cancer
progression. J Hematol Oncol. 12:952019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dirat B, Bochet L, Dabek M, Daviaud D,
Dauvillier S, Majed B, Wang YY, Meulle A, Salles B, Le Gonidec S,
et al: Cancer-associated adipocytes exhibit an activated phenotype
and contribute to breast cancer invasion. Cancer Res. 71:2455–2465.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kokabu S, Lowery JW and Jimi E: Cell fate
and differentiation of bone marrow mesenchymal stem cells. Stem
Cells Int. 2016:37535812016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ridge SM, Sullivan FJ and Glynn SA:
Mesenchymal stem cells: Key players in cancer progression. Mol
Cancer. 16:312017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Han Y, Li X, Zhang Y, Han Y, Chang F and
Ding J: Mesenchymal stem cells for regenerative medicine. Cells.
8:8862019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen Q, Shou P, Zheng C, Jiang M, Cao G,
Yang Q, Cao J, Xie N, Velletri T, Zhang X, et al: Fate decision of
mesenchymal stem cells: Adipocytes or osteoblasts? Cell Death
Differ. 23:1128–1139. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shepherd PR, Gnudi L, Tozzo E, Yang H,
Leach F and Kahn BB: Adipose cell hyperplasia and enhanced glucose
disposal in transgenic mice overexpressing GLUT4 selectively in
adipose tissue. J Biol Chem. 268:22243–22246. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Huang HY, Hu LL, Song TJ, Li X, He Q, Sun
X, Li YM, Lu HJ, Yang PY and Tang QQ: Involvement of
cytoskeleton-associated proteins in the commitment of C3H10T1/2
pluripotent stem cells to adipocyte lineage induced by BMP2/4. Mol
Cell Proteomics. 10:M110.002691. 2011. View Article : Google Scholar
|
|
19
|
de Winter TJ and Nusse R: Running against
the Wnt: How Wnt/β-catenin suppresses adipogenesis. Front Cell Dev
Biol. 9:6274292021. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Huang H, Song TJ, Li X, Hu L, He Q, Liu M,
Lane MD and Tang QQ: BMP signaling pathway is required for
commitment of C3H10T1/2 pluripotent stem cells to the adipocyte
lineage. Proc Natl Acad Sci USA. 106:12670–12675. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tang QQ and Lane MD: Adipogenesis: From
stem cell to adipocyte. Annu Rev Biochem. 81:715–736. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Farmer SR: Transcriptional control of
adipocyte formation. Cell Metab. 4:263–273. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nielsen R, Pedersen TA, Hagenbeek D,
Moulos P, Siersbaek R, Megens E, Denissov S, Børgesen M, Francoijs
KJ, Mandrup S and Stunnenberg HG: Genome-wide profiling of
PPARgamma:RXR and RNA polymerase II occupancy reveals temporal
activation of distinct metabolic pathways and changes in RXR dimer
composition during adipogenesis. Genes Dev. 22:2953–2967. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lefterova MI, Zhang Y, Steger DJ, Schupp
M, Schug J, Cristancho A, Feng D, Zhuo D, Stoeckert CJ Jr, Liu XS
and Lazar MA: PPARgamma and C/EBP factors orchestrate adipocyte
biology via adjacent binding on a genome-wide scale. Genes Dev.
22:2941–2952. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Rosen ED, Walkey CJ, Puigserver P and
Spiegelman BM: Transcriptional regulation of adipogenesis. Genes
Dev. 14:1293–1307. 2000.PubMed/NCBI
|
|
26
|
Kaestner KH, Christy RJ, McLenithan JC,
Braiterman LT, Cornelius P, Pekala PH and Lane MD: Sequence, tissue
distribution, and differential expression of mRNA for a putative
insulin-responsive glucose transporter in mouse 3T3-L1 adipocytes.
Proc Natl Acad Sci USA. 86:3150–3154. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hwang CS, Mandrup S, MacDougald OA, Geiman
DE and Lane MD: Transcriptional activation of the mouse obese (ob)
gene by CCAAT/enhancer binding protein alpha. Proc Natl Acad Sci
USA. 93:873–877. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hwang CS, Loftus TM, Mandrup S and Lane
MD: Adipocyte differentiation and leptin expression. Annu Rev Cell
Dev Biol. 13:231–259. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Soukas A, Socci ND, Saatkamp BD, Novelli S
and Friedman JM: Distinct transcriptional profiles of adipogenesis
in vivo and in vitro. J Biol Chem. 276:34167–34174. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kim KH, Lee K, Moon YS and Sul HS: A
cysteine-rich adipose tissue-specific secretory factor inhibits
adipocyte differentiation. J Biol Chem. 276:11252–11256. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chang E and Kim CY: Natural products and
obesity: A focus on the regulation of mitotic clonal expansion
during adipogenesis. Molecules. 24:11572019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Park A, Kim WK and Bae KH: Distinction of
white, beige and brown adipocytes derived from mesenchymal stem
cells. World J Stem Cells. 6:33–42. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lapeire L, Hendrix A, Lambein K, Van
Bockstal M, Braems G, Van Den Broecke R, Limame R, Mestdagh P,
Vandesompele J, Vanhove C, et al: Cancer-associated adipose tissue
promotes breast cancer progression by paracrine oncostatin M and
Jak/STAT3 signaling. Cancer Res. 74:6806–6819. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Nieman KM, Kenny HA, Penicka CV, Ladanyi
A, Buell-Gutbrod R, Zillhardt MR, Romero IL, Carey MS, Mills GB,
Hotamisligil GS, et al: Adipocytes promote ovarian cancer
metastasis and provide energy for rapid tumor growth. Nat Med.
17:1498–1503. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wu Q, Li J, Li Z, Sun S, Zhu S, Wang L, Wu
J, Yuan J, Zhang Y, Sun S and Wang C: Exosomes from the
tumour-adipocyte interplay stimulate beige/brown differentiation
and reprogram metabolism in stromal adipocytes to promote tumour
progression. J Exp Clin Cancer Res. 38:2232019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Balaban S, Shearer RF, Lee LS, van
Geldermalsen M, Schreuder M, Shtein HC, Cairns R, Thomas KC,
Fazakerley DJ, Grewal T, et al: Adipocyte lipolysis links obesity
to breast cancer growth: Adipocyte-derived fatty acids drive breast
cancer cell proliferation and migration. Cancer Metab. 5:12017.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Notarnicola M, Miccolis A, Tutino V,
Lorusso D and Caruso MG: Low levels of lipogenic enzymes in
peritumoral adipose tissue of colorectal cancer patients. Lipids.
47:59–63. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Choi J, Cha YJ and Koo JS: Adipocyte
biology in breast cancer: From silent bystander to active
facilitator. Prog Lipid Res. 69:11–20. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Fujisaki K, Fujimoto H, Sangai T,
Nagashima T, Sakakibara M, Shiina N, Kuroda M, Aoyagi Y and
Miyazaki M: Cancer-mediated adipose reversion promotes cancer cell
migration via IL-6 and MCP-1. Breast Cancer Res Treat. 150:255–263.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
D'Esposito V, Liguoro D, Ambrosio MR,
Collina F, Cantile M, Spinelli R, Raciti GA, Miele C, Valentino R,
Campiglia P, et al: Adipose microenvironment promotes triple
negative breast cancer cell invasiveness and dissemination by
producing CCL5. Oncotarget. 7:24495–24509. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kim EJ, Kim YK, Kim S, Kim JE, Tian YD,
Doh EJ, Lee DH and Chung JH: Adipochemokines induced by ultraviolet
irradiation contribute to impaired fat metabolism in subcutaneous
fat cells. Br J Dermatol. 178:492–501. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Liu L, Wu Y, Zhang C, Zhou C, Li Y, Zeng
Y, Zhang C, Li R, Luo D, Wang L, et al: Cancer-associated
adipocytes-derived G-CSF promotes breast cancer malignancy via
Stat3 signaling. J Mol Cell Biol. 12:723–737. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Andarawewa KL, Motrescu ER, Chenard MP,
Gansmuller A, Stoll I, Tomasetto C and Rio MC: Stromelysin-3 is a
potent negative regulator of adipogenesis participating to cancer
cell-adipocyte interaction/crosstalk at the tumor invasive front.
Cancer Res. 65:10862–10871. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Iyengar P, Espina V, Williams TW, Lin Y,
Berry D, Jelicks LA, Lee H, Temple K, Graves R, Pollard J, et al:
Adipocyte-derived collagen VI affects early mammary tumor
progression in vivo, demonstrating a critical interaction in the
tumor/stroma microenvironment. J Clin Invest. 115:1163–1176. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Bochet L, Lehuédé C, Dauvillier S, Wang
YY, Dirat B, Laurent V, Dray C, Guiet R, Maridonneau-Parini I, Le
Gonidec S, et al: Adipocyte-derived fibroblasts promote tumor
progression and contribute to the desmoplastic reaction in breast
cancer. Cancer Res. 73:5657–5668. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Côté JA, Guénard F, Lessard J, Lapointe M,
Biron S, Vohl MC and Tchernof A: Temporal changes in gene
expression profile during mature adipocyte dedifferentiation. Int J
Genomics. 2017:51493622017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wei X, Li S, He J, Du H, Liu Y, Yu W, Hu
H, Han L, Wang C, Li H, et al: Tumor-secreted PAI-1 promotes breast
cancer metastasis via the induction of adipocyte-derived collagen
remodeling. Cell Commun Signal. 17:582019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Master SR, Hartman JL, D'Cruz CM, Moody
SE, Keiper EA, Ha SI, Cox JD, Belka GK and Chodosh LA: Functional
microarray analysis of mammary organogenesis reveals a
developmental role in adaptive thermogenesis. Mol Endocrinol.
16:1185–1203. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wang F, Gao S, Chen F, Fu Z, Yin H, Lu X,
Yu J and Lu C: Mammary fat of breast cancer: Gene expression
profiling and functional characterization. PLoS One. 9:e1097422014.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Tsoli M, Schweiger M, Vanniasinghe AS,
Painter A, Zechner R, Clarke S and Robertson G: Depletion of white
adipose tissue in cancer cachexia syndrome is associated with
inflammatory signaling and disrupted circadian regulation. PLoS
One. 9:e929662014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wu Q, Sun S, Li Z, Yang Q, Li B, Zhu S,
Wang L, Wu J, Yuan J, Yang C, et al: Tumour-originated exosomal
miR-155 triggers cancer-associated cachexia to promote tumour
progression. Mol Cancer. 17:1552018. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Guerrero J, Tobar N, Cáceres M, Espinoza
L, Escobar P, Dotor J, Smith PC and Martinez J: Soluble factors
derived from tumor mammary cell lines induce a stromal mammary
adipose reversion in human and mice adipose cells. Possible role of
TGF-beta1 and TNF-alpha. Breast Cancer Res Treat. 119:497–508.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Stephens JM and Pekala PH: Transcriptional
repression of the C/EBP-alpha and GLUT4 genes in 3T3-L1 adipocytes
by tumor necrosis factor-alpha. Regulations is coordinate and
independent of protein synthesis. J Biol Chem. 267:13580–13584.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kim C, Lee H, Cho YM, Kwon OJ, Kim W and
Lee EK: TNFalpha-induced miR-130 resulted in adipocyte dysfunction
during obesity-related inflammation. FEBS Lett. 587:3853–3858.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Lien CC, Au LC, Tsai YL, Ho LT and Juan
CC: Short-term regulation of tumor necrosis factor-alpha-induced
lipolysis in 3T3-L1 adipocytes is mediated through the inducible
nitric oxide synthase/nitric oxide-dependent pathway.
Endocrinology. 150:4892–4900. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Ryden M, Dicker A, van Harmelen V, Hauner
H, Brunnberg M, Perbeck L, Lonnqvist F and Arner P: Mapping of
early signaling events in tumor necrosis factor-alpha -mediated
lipolysis in human fat cells. J Biol Chem. 277:1085–1091. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zhang HH, Halbleib M, Ahmad F, Manganiello
VC and Greenberg AS: Tumor necrosis factor-alpha stimulates
lipolysis in differentiated human adipocytes through activation of
extracellular signal-related kinase and elevation of intracellular
cAMP. Diabetes. 51:2929–2935. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Rydén M, Arvidsson E, Blomqvist L, Perbeck
L, Dicker A and Arner P: Targets for TNF-alpha-induced lipolysis in
human adipocytes. Biochem Biophys Res Commun. 318:168–175. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Souza SC, de Vargas LM, Yamamoto MT, Lien
P, Franciosa MD, Moss LG and Greenberg AS: Overexpression of
perilipin A and B blocks the ability of tumor necrosis factor alpha
to increase lipolysis in 3T3-L1 adipocytes. J Biol Chem.
273:24665–24669. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Tsoli M and Robertson G: Cancer cachexia:
Malignant inflammation, tumorkines, and metabolic mayhem. Trends
Endocrinol Metab. 24:174–183. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Arner P and Langin D: Lipolysis in lipid
turnover, cancer cachexia, and obesity-induced insulin resistance.
Trends Endocrinol Metab. 25:255–262. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Petersen EW, Carey AL, Sacchetti M,
Steinberg GR, Macaulay SL, Febbraio MA and Pedersen BK: Acute IL-6
treatment increases fatty acid turnover in elderly humans in vivo
and in tissue culture in vitro. Am J Physiol Endocrinol Metab.
288:E155–E162. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Ahmadian M, Abbott MJ, Tang T, Hudak CS,
Kim Y, Bruss M, Hellerstein MK, Lee HY, Samuel VT, Shulman GI, et
al: Desnutrin/ATGL is regulated by AMPK and is required for a brown
adipose phenotype. Cell Metab. 13:739–748. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Wang YY, Attané C, Milhas D, Dirat B,
Dauvillier S, Guerard A, Gilhodes J, Lazar I, Alet N, Laurent V, et
al: Mammary adipocytes stimulate breast cancer invasion through
metabolic remodeling of tumor cells. JCI Insight. 2:e874892017.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Ueki K, Kondo T and Kahn CR: Suppressor of
cytokine signaling 1 (SOCS-1) and SOCS-3 cause insulin resistance
through inhibition of tyrosine phosphorylation of insulin receptor
substrate proteins by discrete mechanisms. Mol Cell Biol.
24:5434–5446. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Seale P, Conroe HM, Estall J, Kajimura S,
Frontini A, Ishibashi J, Cohen P, Cinti S and Spiegelman BM: Prdm16
determines the thermogenic program of subcutaneous white adipose
tissue in mice. J Clin Invest. 121:96–105. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Cannon B and Nedergaard J: Brown adipose
tissue: Function and physiological significance. Physiol Rev.
84:277–359. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Lee J, Hong BS, Ryu HS, Lee HB, Lee M,
Park IA, Kim J, Han W, Noh DY and Moon HG: Transition into
inflammatory cancer-associated adipocytes in breast cancer
microenvironment requires microRNA regulatory mechanism. PLoS One.
12:e01741262017. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Arora GK, Gupta A, Narayanan S, Guo T,
Iyengar P and Infante RE: Cachexia-associated adipose loss induced
by tumor-secreted leukemia inhibitory factor is counterbalanced by
decreased leptin. JCI Insight. 3:e1212212018. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Zoico E, Darra E, Rizzatti V, Budui S,
Franceschetti G, Mazzali G, Rossi AP, Fantin F, Menegazzi M, et al:
Adipocytes WNT5a mediated dedifferentiation: a possible target in
pancreatic cancer microenvironment. Oncotarget. 7:20223–20235.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Kang MI, Baker AR, Dextras CR, Cabarcas
SM, Young MR and Colburn NH: (2012). Targeting of Noncanonical
Wnt5a Signaling by AP-1 Blocker Dominant-Negative Jun When It
Inhibits Skin Carcinogenesis. Genes cancer. 3:37–50. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Gustafson B and Smith U: Activation of
canonical wingless-type MMTV integration site family (Wnt)
signaling in mature adipocytes increases beta-catenin levels and
leads to cell dedifferentiation and insulin resistance. J Biol
Chem. 285:14031–14041. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Christodoulides C, Lagathu C, Sethi JK and
Vidal-Puig A: Adipogenesis and WNT signalling. Trends Endocrinol
Metab. 20:16–24. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Zoico E, Darra E, Rizzatti V, Budui S,
Franceschetti G, Mazzali G, Rossi AP, Fantin F, Menegazzi M, Cinti
S and Zamboni M: Adipocytes WNT5a mediated dedifferentiation: A
possible target in pancreatic cancer microenvironment. Oncotarget.
7:20223–20235. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Bilkovski R, Schulte DM, Oberhauser F,
Mauer J, Hampel B, Gutschow C, Krone W and Laudes M: Adipose tissue
macrophages inhibit adipogenesis of mesenchymal precursor cells via
wnt-5a in humans. Int J Obes (Lond). 35:1450–1454. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Klaus A and Birchmeier W: Wnt signalling
and its impact on development and cancer. Nat Rev Cancer.
8:387–398. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Bauer M, Bénard J, Gaasterland T, Willert
K and Cappellen D: WNT5A encodes two isoforms with distinct
functions in cancers. PLoS One. 8:e805262013. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Zhuang Y, Li X, Zhan P, Pi G and Wen G:
MMP11 promotes the proliferation and progression of breast cancer
through stabilizing Smad2 protein. Oncol Rep. 45:162021. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Rio MC: From a unique cell to metastasis
is a long way to go: Clues to stromelysin-3 participation.
Biochimie. 87:299–306. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Xu G, Zhang B, Ye J, Cao S, Shi J, Zhao Y,
Wang Y, Sang J, Yao Y, Guan W, et al: Exosomal miRNA-139 in
cancer-associated fibroblasts inhibits gastric cancer progression
by repressing MMP11 expression. Int J Biol Sci. 15:2320–2329. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Motrescu ER, Blaise S, Etique N, Messaddeq
N, Chenard MP, Stoll I, Tomasetto C and Rio MC: Matrix
metalloproteinase-11/stromelysin-3 exhibits collagenolytic function
against collagen VI under normal and malignant conditions.
Oncogene. 27:6347–6355. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Kozlova N, Jensen JK, Chi TF, Samoylenko A
and Kietzmann T: PAI-1 modulates cell migration in a LRP1-dependent
manner via β-catenin and ERK1/2. Thromb Haemost. 113:988–998. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Benesch MG, Ko YM, McMullen TP and
Brindley DN: Autotaxin in the crosshairs: Taking aim at cancer and
other inflammatory conditions. FEBS Lett. 588:2712–2727. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Brindley DN, Lin FT and Tigyi GJ: Role of
the autotaxin-lysophosphatidate axis in cancer resistance to
chemotherapy and radiotherapy. Biochim Biophys Acta. 1831:74–85.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Choi JW and Chun J: Lysophospholipids and
their receptors in the central nervous system. Biochim Biophys
Acta. 1831:20–32. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Samadi N, Bekele R, Capatos D, Venkatraman
G, Sariahmetoglu M and Brindley DN: Regulation of lysophosphatidate
signaling by autotaxin and lipid phosphate phosphatases with
respect to tumor progression, angiogenesis, metastasis and
chemo-resistance. Biochimie. 93:61–70. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
So J, Wang FQ, Navari J, Schreher J and
Fishman DA: LPA-induced epithelial ovarian cancer (EOC) in vitro
invasion and migration are mediated by VEGF receptor-2 (VEGF-R2).
Gynecol Oncol. 97:870–878. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Murph MM, Hurst-Kennedy J, Newton V,
Brindley DN and Radhakrishna H: Lysophosphatidic acid decreases the
nuclear localization and cellular abundance of the p53 tumor
suppressor in A549 lung carcinoma cells. Mol Cancer Res.
5:1201–1211. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Popnikolov NK, Dalwadi BH, Thomas JD,
Johannes GJ and Imagawa WT: Association of autotaxin and
lysophosphatidic acid receptor 3 with aggressiveness of human
breast carcinoma. Tumour Biol. 33:2237–2243. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Brindley DN, Tang X, Meng G and Benesch
MGK: Role of adipose tissue-derived autotaxin, lysophosphatidate
signaling, and inflammation in the progression and treatment of
breast cancer. Int J Mol Sci. 21:59382020. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Benesch MG, Tang X, Maeda T, Ohhata A,
Zhao YY, Kok BP, Dewald J, Hitt M, Curtis JM, McMullen TP and
Brindley DN: Inhibition of autotaxin delays breast tumor growth and
lung metastasis in mice. FASEB J. 28:2655–2666. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Benesch MG, Zhao YY, Curtis JM, McMullen
TP and Brindley DN: Regulation of autotaxin expression and
secretion by lysophosphatidate and sphingosine 1-phosphate. J Lipid
Res. 56:1134–1144. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Benesch MG, Tang X, Dewald J, Dong WF,
Mackey JR, Hemmings DG, McMullen TP and Brindley DN: Tumor-induced
inflammation in mammary adipose tissue stimulates a vicious cycle
of autotaxin expression and breast cancer progression. FASEB J.
29:3990–4000. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Russell ST, Zimmerman TP, Domin BA and
Tisdale MJ: Induction of lipolysis in vitro and loss of body fat in
vivo by zinc-alpha2-glycoprotein. Biochim Biophys Acta. 1636:59–68.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Bing C, Bao Y, Jenkins J, Sanders P,
Manieri M, Cinti S, Tisdale MJ and Trayhurn P:
Zinc-alpha2-glycoprotein, a lipid mobilizing factor, is expressed
in adipocytes and is up-regulated in mice with cancer cachexia.
Proc Natl Acad Sci USA. 101:2500–2505. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Delort L, Perrier S, Dubois V, Billard H,
Mracek T, Bing C, Vasson MP and Caldefie-Chézet F:
Zinc-α2-glycoprotein: A proliferative factor for breast cancer?
In vitro study and molecular mechanisms. Oncol Rep.
29:2025–2029. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Zhu H, Liu M, Zhang N, Pan H, Lin G, Li N,
Wang L, Yang H, Yan K and Gong F: Circulating and adipose tissue
mRNA levels of Zinc-α2-glycoprotein, leptin, high-molecular-weight
adiponectin, and tumor necrosis factor-alpha in colorectal cancer
patients with or without obesity. Front Endocrinol (Lausanne).
9:1902018. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Henshall SM, Horvath LG, Quinn DI,
Eggleton SA, Grygiel JJ, Stricker PD, Biankin AV, Kench JG and
Sutherland RL: Zinc-alpha2-glycoprotein expression as a predictor
of metastatic prostate cancer following radical prostatectomy. J
Natl Cancer Inst. 98:1420–1424. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Elattar S, Dimri M and Satyanarayana A:
The tumor secretory factor ZAG promotes white adipose tissue
browning and energy wasting. FASEB J. 32:4727–4743. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Gong FY, Zhang SJ, Deng JY, Zhu HJ, Pan H,
Li NS and Shi YF: Zinc-alpha2-glycoprotein is involved in
regulation of body weight through inhibition of lipogenic enzymes
in adipose tissue. Int J Obes (Lond). 33:1023–1030. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Xiao XH, Qi XY, Wang YD, Ran L, Yang J,
Zhang HL, Xu CX, Wen GB and Liu JH: Zinc alpha2 glycoprotein
promotes browning in adipocytes. Biochem Biophys Res Commun.
496:287–293. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Vlassov AV, Magdaleno S, Setterquist R and
Conrad R: Exosomes: Current knowledge of their composition,
biological functions, and diagnostic and therapeutic potentials.
Biochim Biophys Acta. 1820:940–948. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Minciacchi VR, Freeman MR and Di Vizio D:
Extracellular vesicles in cancer: Exosomes, microvesicles and the
emerging role of large oncosomes. Semin Cell Dev Biol. 40:41–51.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Wang H, Liu L, Lin JZ, Aprahamian TR and
Farmer SR: Browning of white adipose tissue with roscovitine
induces a distinct population of UCP1(+) adipocytes. Cell Metab.
24:835–847. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Tomasetti M, Nocchi L, Staffolani S,
Manzella N, Amati M, Goodwin J, Kluckova K, Nguyen M, Strafella E,
Bajzikova M, et al: MicroRNA-126 suppresses mesothelioma malignancy
by targeting IRS1 and interfering with the mitochondrial function.
Antioxid Redox Signal. 21:2109–2125. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Fong MY, Zhou W, Liu L, Alontaga AY,
Chandra M, Ashby J, Chow A, O'Connor ST, Li S, Chin AR, et al:
Breast-cancer-secreted miR-122 reprograms glucose metabolism in
premetastatic niche to promote metastasis. Nat Cell Biol.
17:183–194. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Yan W, Wu X, Zhou W, Fong MY, Cao M, Liu
J, Liu X, Chen CH, Fadare O, Pizzo DP, et al: Cancer-cell-secreted
exosomal miR-105 promotes tumour growth through the MYC-dependent
metabolic reprogramming of stromal cells. Nat Cell Biol.
20:597–609. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Wang S, Xu M, Li X, Su X, Xiao X, Keating
A and Zhao RC: Exosomes released by hepatocarcinoma cells endow
adipocytes with tumor-promoting properties. J Hematol Oncol.
11:822018. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Sagar G, Sah RP, Javeed N, Dutta SK, Smyrk
TC, Lau JS, Giorgadze N, Tchkonia T, Kirkland JL, Chari ST and
Mukhopadhyay D: Pathogenesis of pancreatic cancer exosome-induced
lipolysis in adipose tissue. Gut. 65:1165–1174. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Kong F, Li L, Du Y, Zhu H, Li Z and Kong
X: Exosomal adrenomedullin derived from cancer-associated
fibroblasts promotes lipolysis in adipose tissue. Gut.
67:2226–2227. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Hu W, Ru Z, Zhou Y, Xiao W, Sun R, Zhang
S, Gao Y, Li X, Zhang X and Yang H: Lung cancer-derived
extracellular vesicles induced myotube atrophy and adipocyte
lipolysis via the extracellular IL-6-mediated STAT3 pathway.
Biochim Biophys Acta Mol Cell Biol Lipids. 1864:1091–1102. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Zhang H, Zhu L, Bai M, Liu Y, Zhan Y, Deng
T, Yang H, Sun W, Wang X, Zhu K, et al: Exosomal circRNA derived
from gastric tumor promotes white adipose browning by targeting the
miR-133/PRDM16 pathway. Int J Cancer. 144:2501–2515. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Bhome R, Goh RW, Bullock MD, Pillar N,
Thirdborough SM, Mellone M, Mirnezami R, Galea D, Veselkov K, Gu Q,
et al: Exosomal microRNAs derived from colorectal cancer-associated
fibroblasts: Role in driving cancer progression. Aging (Albany NY).
9:2666–2694. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Au Yeung CL, Co NN, Tsuruga T, Yeung TL,
Kwan SY, Leung CS, Li Y, Lu ES, Kwan K, Wong KK, et al: Exosomal
transfer of stroma-derived miR21 confers paclitaxel resistance in
ovarian cancer cells through targeting APAF1. Nat Commun.
7:111502016. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Zheng Z, Liu L, Zhan Y, Yu S and Kang T:
Adipose-derived stem cell-derived microvesicle-released miR-210
promoted proliferation, migration and invasion of endothelial cells
by regulating RUNX3. Cell Cycle. 17:1026–1033. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Lazar I, Clement E, Dauvillier S, Milhas
D, Ducoux-Petit M, LeGonidec S, Moro C, Soldan V, Dalle S, Balor S,
et al: Adipocyte exosomes promote melanoma aggressiveness through
fatty acid oxidation: A novel mechanism linking obesity and cancer.
Cancer Res. 76:4051–4057. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Wang J, Wu Y, Guo J, Fei X, Yu L and Ma S:
Adipocyte-derived exosomes promote lung cancer metastasis by
increasing MMP9 activity via transferring MMP3 to lung cancer
cells. Oncotarget. 8:81880–81891. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Zaidi N, Lupien L, Kuemmerle NB, Kinlaw
WB, Swinnen JV and Smans K: Lipogenesis and lipolysis: The pathways
exploited by the cancer cells to acquire fatty acids. Prog Lipid
Res. 52:585–589. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Kedia-Mehta N and Finlay DK: Competition
for nutrients and its role in controlling immune responses. Nat
Commun. 10:21232019. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Gonzalez-Perez RR, Xu Y, Guo S, Watters A,
Zhou W and Leibovich SJ: Leptin upregulates VEGF in breast cancer
via canonic and non-canonical signalling pathways and
NFkappaB/HIF-1alpha activation. Cell Signal. 22:1350–1362. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Liu S, Lee JS, Jie C, Park MH, Iwakura Y,
Patel Y, Soni M, Reisman D and Chen H: HER2 overexpression triggers
an IL1α proinflammatory circuit to drive tumorigenesis and promote
chemotherapy resistance. Cancer Res. 78:2040–2051. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Zhao J, Liu J, Wu N, Zhang H, Zhang S, Li
L and Wang M: ANGPTL4 overexpression is associated with progression
and poor prognosis in breast cancer. Oncol Lett. 20:2499–2505.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Meng L, Zhou J, Sasano H, Suzuki T,
Zeitoun KM and Bulun SE: Tumor necrosis factor alpha and
interleukin 11 secreted by malignant breast epithelial cells
inhibit adipocyte differentiation by selectively down-regulating
CCAAT/enhancer binding protein alpha and peroxisome
proliferator-activated receptor gamma: Mechanism of desmoplastic
reaction. Cancer Res. 61:2250–2255. 2001.PubMed/NCBI
|
|
124
|
Gray NE, Lam LN, Yang K, Zhou AY, Koliwad
S and Wang JC: Angiopoietin-like 4 (Angptl4) protein is a
physiological mediator of intracellular lipolysis in murine
adipocytes. J Biol Chem. 287:8444–8456. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Kogo R, Shimamura T, Mimori K, Kawahara K,
Imoto S, Sudo T, Tanaka F, Shibata K, Suzuki A, Komune S, et al:
Long noncoding RNA HOTAIR regulates polycomb-dependent chromatin
modification and is associated with poor prognosis in colorectal
cancers. Cancer Res. 71:6320–6326. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Gupta RA, Shah N, Wang KC, Kim J, Horlings
HM, Wong DJ, Tsai MC, Hung T, Argani P, Rinn JL, et al: Long
non-coding RNA HOTAIR reprograms chromatin state to promote cancer
metastasis. Nature. 464:1071–1076. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Ma X, Li Z, Li T, Zhu L, Li Z and Tian N:
Long non-coding RNA HOTAIR enhances angiogenesis by induction of
VEGFA expression in glioma cells and transmission to endothelial
cells via glioma cell derived-extracellular vesicles. Am J Transl
Res. 9:5012–5021. 2017.PubMed/NCBI
|
|
128
|
Sanchez-Alvarez R, Martinez-Outschoorn UE,
Lamb R, Hulit J, Howell A, Gandara R, Sartini M, Rubin E, Lisanti
MP and Sotgia F: Mitochondrial dysfunction in breast cancer cells
prevents tumor growth: Understanding chemoprevention with
metformin. Cell Cycle. 12:172–182. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Munteanu R, Onaciu A, Moldovan C, Zimta
AA, Gulei D, Paradiso AV, Lazar V and Berindan-Neagoe I:
Adipocyte-based cell therapy in oncology: The role of
cancer-associated adipocytes and their reinterpretation as delivery
platforms. Pharmaceutics. 12:4022020. View Article : Google Scholar : PubMed/NCBI
|