Introduction
Chronic pancreatitis (CP), considered as a main
cause of pancreatic cancer responsible for high morbidity, is
associated with an increased incidence and prevalence worldwide
(1,2). CP is a pancreatic inflammatory
disease characterized by histological changes, including fibrosis,
acinar cell loss and immune cell infiltration and eventually leads
to damage of the pancreas, resulting in numerous downstream effects
(3). CP not only reduces the
quality of life of patients due to pain, weight loss and
malnutrition but is also a putative risk factor for pancreatic
cancer and diabetes and their consequent complications (4–9).
Recent research has suggested that in South Korea, the incidence of
cancer and the resultant mortality rate are 1.2 times higher in
patients with CP than those in the individuals without CP (10). Frequent alcohol consumption is
known to be the main cause of CP, accounting for ~80% of cases of
CP development. However, the non-alcohol-dependent incidence has
also been increasing recently (2,11,12).
Therefore, it is imperative to develop effective CP treatments
beyond lifestyle improvements.
Plants and their active ingredients which are found
in food and tea, are consumed daily and have been attracting
attention as valuable targets in drug development because they
reportedly, prevent diseases and improve treatment effects beyond
simple nutrition (13). Tea, in
particular, is the most consumed beverage in the world, with the
exception of water, and has beneficial effects, including
anti-fibrotic, anti-inflammatory and anti-cancer effects, mainly
caused by polyphenols (14–17).
Catechins, which are polyphenolic flavonoids, are abundant in
commonly consumed foods and herbs such as apples and tea (18). Among them,
(−)-epigallocatechin-3-gallate (EGCG), a major catechin in green
tea, has been reported to alleviate pancreas-related diseases,
including fibrosis, inflammation and cancer (18–21).
Initially, low-dose intake of catechins was considered safe, but
recent studies of hepatotoxicity in catechins have raised safety
concerns related to their consumption (22,23).
Catechin and catechin hydrate (CH) are natural
flavon-3-ol phytochemicals and are relatively less toxic than other
flavonoids including EGCG (24–26).
CH is found in Tamarindus indica fruit pulp, the fruit peel
of Areca catechu Linné and in green tea (24,27,28).
Beneficial effects of catechin on various diseases have been
previously reported. CH has protective efficacy against
benzo(a)pyrene induced lung injury by regulating apoptosis,
inflammation and oxidative stress (29). It has also demonstrated anticancer
effects through inhibition of the proliferation of MCF-7 cells and
inducing P53 and caspase-medicated apoptosis of human breast cancer
cells (26). In addition to
antioxidant and apoptosis activities, the efficacy of (+)-catechin
on hepatic fibrosis has been reported. Bragança de Moraes et
al (30) reported that it
reversed activated hepatic stellate cells (HSCs) to the quiescent
state by regulating intracellular lipid droplets and HSC quiescent
markers.
Previous studies have primarily focused on the
medicinal effects of green tea extract, which were limited to
assessing the effects of EGCG. Therefore, despite the
aforementioned results, studies on other catechins are still
lacking. In our previous study, it was demonstrated that the water
extract of fruit peel of Areca catechu Linné contained
catechin which showed remarkable inhibitory effects on pancreatic
fibrosis (27). Despite this,
little research has been reported regarding the effect of CH on
pancreatic fibrosis. Therefore, the present study assessed the
potentially beneficial effects of (+)-CH, another promising single
compound, on CP.
Materials and methods
Preparations of (+)-CH
(+)-CH was purchased from MilliporeSigma (cat. no.
1251G; purity ≥98%). The powder was dissolved in ethanol (cat no.
1.00983.1011; Merck Life Science UK, Ltd.) at a concentration of
200 mg/ml to prepare the stock solution and was further diluted in
saline or Dulbecco's Modified Eagle Medium (DMEM; cat. no.
11995-065; Thermo Fisher Scientific, Inc.) to obtain the desired
concentrations.
Animals
All experiments were performed with the approval of
The Animal Care Committee of Wonkwang University (approval no.
WKU22-43; Iksan, Republic of Korea). A total of 196 C57BL/6 mice
(6–8 weeks old, female, weighing 15–20 g) were purchased from
Samtako Biokorea Co., Ltd. Mice were housed in standard shoebox
cages in a climate-controlled room maintained at an ambient
temperature of 23±2°C and a 12 h light-dark cycle for 7 days. They
were fed standard laboratory chow, offered water ad libitum
and were arbitrarily assigned to the control and experimental
groups. Isoflurane (induction, 4.5%; maintenance, 1.5%) in 95%
O2 and 5% CO2 was used for anesthesia.
CO2 inhalation was used for euthanasia with a flow rate
which displaced 50% of the cage vol/min and cervical dislocation
was also performed to ensure death following CO2
asphyxiation.
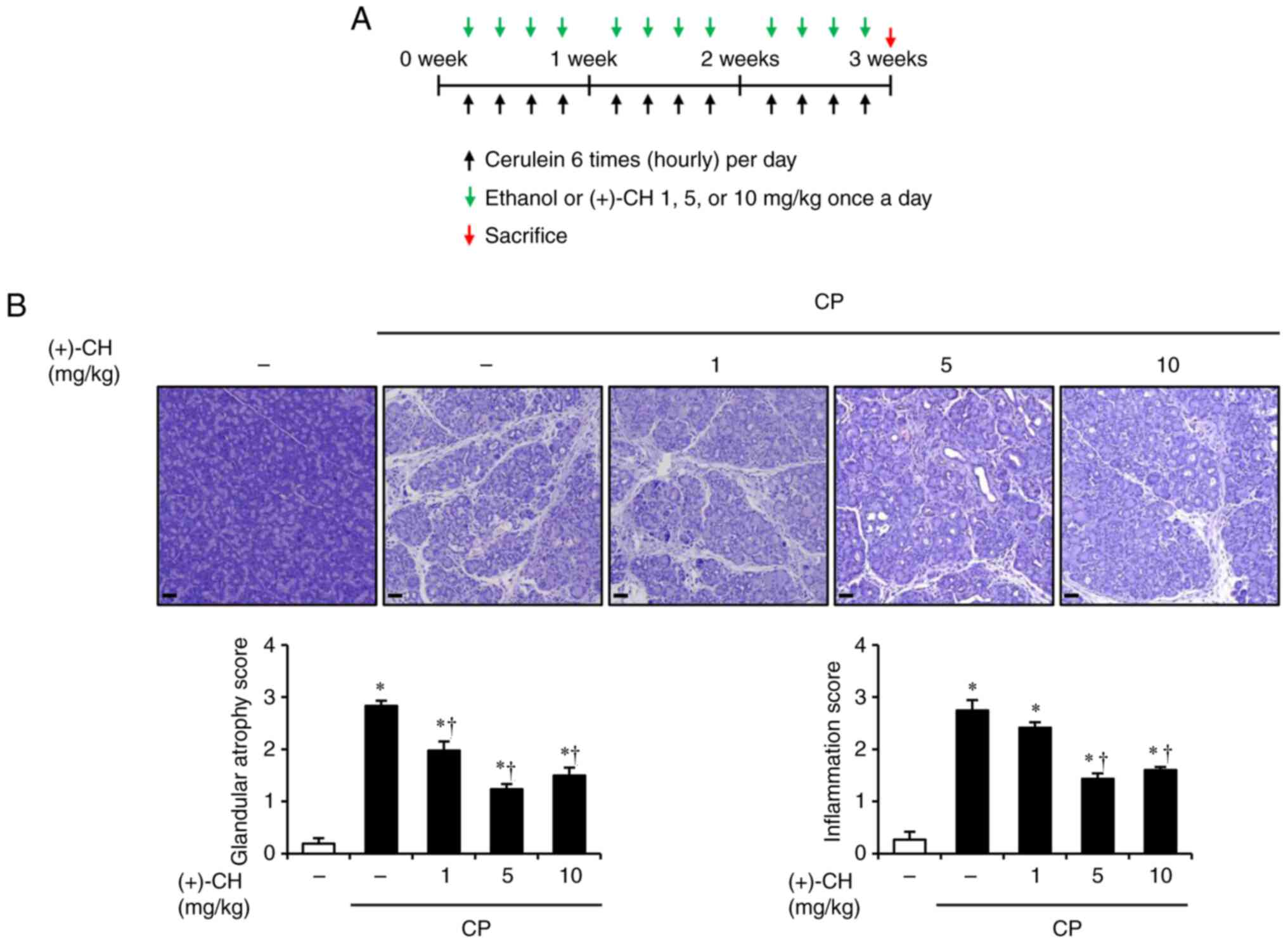
Experimental design
Mice were intraperitoneally injected with 50 µg/kg
cerulein (cat. no. H-3220; Bachem AG) six times/day at 1 h
intervals, four times/week, for 3 weeks. In the prophylactic
treatment groups, mice were intraperitoneally injected with (+)-CH
(1, 5 or 10 mg/kg) or ethanol (control group) 1 h before the first
daily injection four times/week (Fig.
1A). In the curative group, (+)-CH (5 mg/kg) was administered
for 2 weeks or 1 week after the first cerulein injection. After 3
weeks, the mice were sacrificed using CO2 asphyxiation
and the pancreas was collected and stored at −80°C for further
analysis.
Biochemical analysis
(+)-CH (1, 10, 20 or 50 mg/kg) or ethanol (control
group) were intraperitoneally administered to mice (n=3/group). At
24 h following administration, the serum was collected by cardiac
puncture and separated by centrifugation at 2,339 × g for 5 min at
4°C. Fresh serum was analyzed using biochemical analyzer (cat. no.
NX700i; FUJIFILM Wako Pure Chemical Corporation) according to the
manufacturer's instructions. The levels of the following molecules
were examined: Alkaline phosphatase (ALP), alanine transaminase
(ALT), aspartate transaminase (AST), blood urea nitrogen (BUN) and
creatinine (CREA).
Tissue processing
After obtaining fresh pancreas, tissues were washed
with cold phosphate buffered saline (PBS; cat. no. CNP010-1000;
Isell Co., Ltd.) and the specimens were fixed in 10%
neutral-buffered-formalin solution (cat. no. HT501128;
MilliporeSigma) for 24 h at room temperature. Tissues were then
dehydrated using an ascending alcohol series. Subsequently, ethanol
was cleared using xylene. The tissues were infiltrated with
paraffin wax and embedded and sectioned into 4 µm thick sections.
Formalin-fixed paraffin-embedded (FFPE) samples were deparaffinized
by placing slides in xylene twice for 10 min each and rehydrated in
100% ethanol twice for 5 min each, 95% ethanol for 2 min, 70%
ethanol for 2 min at room temperature. The samples were then
stained using hematoxylin for 8 min and eosin for 2 min at room
temperature. The experiment was performed independently, three
times. The sections were imaged using NIS-Elements Viewer 4.2
(Nikon instruments). The samples were scored on a scale from 0 to 3
based on the presence of glandular atrophy and inflammation (0,
normal, no glandular atrophy and inflammation; 1=mild, found in
less than 25% of the pancreas. 2=moderate, found in less than 25 to
75% of the pancreas. 3=found in more than 75 % of the
pancreas).
Immunohistochemical (IHC)
staining
Prior to IHC staining, FFPE pancreatic tissues were
deparaffinized and rehydrated. The slide was placed in 10-fold
diluted citrate buffer Antigen retriever (pH 6.0, cat. no. C9999;
MilliporeSigma) and heated for 20 min at 100°C. The slides were
blocked with 5% normal goat serum (Jackson ImmunoResearch
Laboratories, Inc.) at RT for 1 h and incubated with a primary
antibodies against α-smooth muscle actin (α-SMA; 1:100; cat. no.
sc-32251; Santa Cruz Biotechnology, Inc.) overnight at 4°C.
Following incubation with HRP-conjugated goat anti-rabbit secondary
antibody (1:1,000; cat. no. SA002-500; GenDEPOT, LLC) for 1 h at
RT, the slides were covered with diluted 3,3′-diaminobenzidine
(DAB) solution; Dako Liquid DAB+ Substrate Chromogen
System (1:50; cat. no. K3467; Dako; Agilent Technologies, Inc.) and
incubated for 3 min. Hematoxylin was used as the counterstain for 3
min at RT. The experiment was performed independently, three times.
Samples were imaged using a light microscope Nikon Ti2-U (Nikon
instruments) and the relative intensity was quantified using the
microscopy software NIS-Elements Viewer 4.2 (Nikon
instruments).
Immunofluorescence (IF) staining
IF analysis of collagen I was performed using
formalin-fixed pancreatic samples preserved in 25% sucrose.
Unfrozen fresh pancreases were collected and fixed in 10%
neutral-buffered-formalin solution (cat. no. HT501128;
MilliporeSigma) for 24 h at RT. Following fixation, the specimens
were embedded in FSC22 Frozen Section Media (cat. no. 3801480;
Leica Microsystems GmbH) and cut into 9 µm thick sections. The
tissues were blocked with 5% normal goat serum (Jackson
ImmunoResearch Laboratories, INC.) at RT for 1 h and incubated with
primary antibodies against collagen I (1:100; cat. no. ab34710;
Abcam) diluted in 5% normal goat serum overnight at 4°C.
Subsequently, the tissues were incubated with fluorescence-labeled,
Alexa Fluor® 488 goat anti-rabbit secondary antibodies
(1:2,000; cat. no. A27034; Invitrogen; Thermo Fisher Scientific,
Inc.) at RT for 2 h. Nuclei were counterstained using
4′,6-diamidino-2-phenylindole (DAPI; 5 ng/ml; cat. no. D1306;
Invitrogen; Thermo Fisher Scientific, Inc.) at RT for 5 min. The
experiment was performed independently three times. The stained
tissues were imaged using a FV1000 confocal laser scanning
biological microscope (Olympus Corporation) at the Core Facility
for Supporting Analysis and Imaging of Biomedical Materials at
Wonkwang University (Iksan, Republic of Korea), supported by the
National Research Facilities and Equipment Center (Daejeon,
Republic of Korea). The samples were visualized using FV10-ASW 4.2
viewer (Olympus).
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was isolated from the pancreas or
pancreatic stellate cells (PSCs) using an Easy-Blue™ RNA extraction
kit (cat. no. 17061; Intron Biotechnology, Inc.). RNA purity was
verified using a Gene Quant Pro RNA calculator (Biochrom, Ltd.).
RNA was reverse transcribed to cDNA using an ABI cDNA synthesis kit
(cat. no. 4387406; Applied Biosystems; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. Subsequently, the
cDNA was amplified using Real-Time PCR SYBR Master Mix (cat. no.
4367659; Applied Biosystems; Thermo Fisher Scientific, Inc.) in a
Step One Plus Real Time PCR system (Applied Biosystems; Thermo
Fisher Scientific, Inc.). The primer sequences used in the present
study were as follows: Acta-2 forward (F),
5′-GTCCCAGACATCAGGGAGTAA-3′ and reverse (R),
5′-TCGGATACTTCAGCGTCAGGA-3′; fibronectin 1 (FN1) F
5′-GATGTCCGAACAGCTATTTACCA-3′ and R 5′-CCTTGCGACTTCAGCCACT-3′;
Collagen I F 5′-GTGGTGACAAGGGTGAGACA-3′ and R
5′-GAGAACCAGGAGAACCAGGA-3′; Collagen III F
5′-TACACCTGCTCCTGTGCTTC-3′ and R 5′-CATTCCTCCCACTCCAGACT-3′;
Collagen IV F 5′-CAAAGGCATCAGGGGAATAACT-3′ and R
(5′-ACCCTTAGATCCGTTGCATCC-3′; and glyceraldehyde 3-phosphate
dehydrogenase (GAPDH) F 5′-TCCCACTCTTCCACCTTCGA-3′ and R
5′-AGTTGGGATAGGGCCTCTCTTG-3′. Amplification was achieved by a
series of steps as follows: 95°C for 10 min, followed by 40 cycles
at 95°C for 15 sec and 60°C for 1 min each, followed by
dissociation for 15 s at 95°C and 1 min at 60°C, followed by 15 sec
at 95°C on ABI Step One Plus. Step One software v2.3 (Applied
Biosystems; Thermo Fisher Scientific, Inc.) was used for data
analysis. Relative gene expression (with target gene expression
normalized to GAPDH) was calculated using the 2−ΔΔCq
method (31). The analysis was
independently conducted thrice.
Isolation of PSCs
PSCs were isolated from C57BL/6 mice using the
method previously described by Zang et al (32). Briefly, the pancreases of C57BL/6
mice were removed, minced with scissors and digested using
collagenase-containing Gey's balanced salt solution (GBSS with
sodium chloride; cat. no G9779; Sigma-Aldrich) for 20 min at 37°C
in a shaking water bath. Following collagenase digestion, the cell
suspension was filtered through a 100 µm nylon mesh (cat. no.
352360; Falcon; Corning Life Sciences) and subjected to isopycnic
separation with Nycodenz solution (Histonez; cat. no D2158;
Sigma-Aldrich; Merck KGaA). Subsequently, the cells were collected
from the top of the gradient, washed twice, resuspended in DMEM
supplemented with 10% fetal bovine serum (cat. no. 16000-044;
Thermo Fisher Scientific, Inc.) and 1% penicillin/streptomycin (1
mg/5 ml; cat. no. 15140-122; Thermo Fisher Scientific, Inc.) and
incubated in an environment with 95% O2 and 5%
CO2. All experiments were performed by culturing cells
between passages 0 and 3. PSCs were seeded at a density of
2×105 cell/well in 6 well and 5×105 cell/well
in 6 cm dish. After confirming that (+)-CH did not exhibit any
cytotoxic effects on PSCs at the specified concentrations, the
following experiments were performed.
Cell treatment
To measure changes in the protein expression levels
of Smad in TGF-β induced signaling, PSCs were pretreated with 250
µM (+)-CH for 1 h at 37°C and then exposed to 0.5 ng/ml TGF-β1
(cat. no. 7666-MB; R&D Systems, Inc.) for 15 min at 37°C. Whole
cell lysates were harvested for further experiments. To provide
information regarding the anti-fibrotic effect of (+)-CH at the
mRNA level, cells were pretreated with (+)-CH at various
concentrations (150, 200 and 250 µM) for 1 h at 37°C and then
stimulated with 0.5 ng/ml TGF-β1 or 25 ng/ml platelet-derived
growth factor (PDGF)-BB (cat. no. 315-18; PeproTech, Inc.) for 24 h
at 37°C.
Western blotting
To acquire proteins from PSCs, the medium in 6 cm
dishes with cells was discarded and cells were washed with ice-cold
PBS. Cells were lysed with radioimmunoprecipitation assay lysis
buffer (cat. no. IBS-BR004; iNtRON Biotechnology, Co.) containing
1% EZ block protease inhibitor cocktail (cat. no. K272-1;
BioVision, Inc.) and 1% phosphatase inhibitor cocktail (cat. no.
p5726; MilliporeSigma). The contents were agitated on ice for 1 h.
The supernatants were boiled for 5 min at 99°C in 62.5 mM Tris-HCl
buffer (pH 6.8) containing 2% sodium dodecyl sulfate (SDS; cat. no.
S1377.0500; Duchefa Biochemie B.V.), 20% glycerol (cat. no. G6279;
MilliporeSigma) and 10% 2-mercaptoethanol (cat. no. 21985-023;
Thermo Fisher Scientific, Inc.). The BCA method was used for
protein quantification. 20 µg proteins were separated on 8%
SDS-polyacrylamide gels and transferred onto nitrocellulose
membranes. The membranes were then rinsed and blocked with 5% skim
milk (cat. no. 232100; Becton, Dickinson and Company) in PBS
containing 1% Tween 20 (cat. no. T9100-010; GenDEPOT, LLC) for 2 h
at RT. The membranes were then incubated with diluted primary
antibodies over night at 4°C with gentle shaking. Antibodies
against phosphorylated Smad2/3 (1:500; cat. no. 8828S; Cell
Signaling Technology, Inc.), Smad2/3 (1:500; cat. no. 3102S; Cell
Signaling Technology, Inc.) and GAPDH (1:1,000; cat. no. 2118S;
Cell Signaling Technology, Inc.) were used as the primary
antibodies. After washing four times, the membranes were incubated
in a diluted solution of goat anti-rabbit secondary antibodies
(cat. no. SA002-500; GenDEPOT, LLC) for 1 h at RT. Antibodies were
diluted with an immunoreaction enhancer solution (cat. no. NKB-101;
Toyobo Life Science). The proteins were visualized using an
enhanced chemiluminescence detection system (Amersham; Cytiva)
according to the manufacturer's protocol. Samples were assessed in
triplicate. Densitometric analysis of western blot was used by
ImageJ version 1.53k (National Institutes of Health).
Statistical analysis
Data were analyzed using SPSS software (version 15;
IBM, SPSS). Results are presented as the mean ± standard error of
the mean. Significance was evaluated using a two-way analysis of
variance (ANOVA) with time and dose parameters followed by post hoc
Tukey's tests for multiple comparisons among groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
Effect of (+)-CH on biochemical
parameters in mice
Toxicity testing is imperative in drug development.
To assess the toxicity dosage of (+)-CH, biochemical parameters,
ALP, ALT, AST, BUN and CREA, were assessed under ordinary
conditions in mice. Either (+)-CH or ethanol (control) was
administered intraperitoneally to the mice and serum samples were
collected after 24 h to evaluate biochemical factors. Compared with
the control group, neither (+)-CH nor ethanol demonstrated
toxicity, even at 50 mg/kg (Table
I). According to preliminary experiments, the effect of
improving CP was better at a lower concentration (10 mg/kg) without
toxicity (data not shown). Therefore, concentrations of (+)-CH
<50 mg/kg were used in the subsequent experiments to determine
its medicinal effects on CP.
 | Table I.Biochemical parameters of mice
treated with (+)-CH in acute toxicity test. |
Table I.
Biochemical parameters of mice
treated with (+)-CH in acute toxicity test.
| Group | ALP, IU/l | ALT, IU/l | AST, IU/l | BUN, mg/dl | CREA, mg/dl |
|---|
| Normal | 293.67±46.37 | 26.33±1.53 | 144.33±46.50 | 21.33±3.51 | 0.11±0.02 |
| Ethanol | 277.67±18.77 | 20.67±3.21 | 130.00±33.42 | 21.00±1.00 | 0.12±0.01 |
| (+)-CH, mg/kg |
|
|
|
|
|
| 1 | 253.33±18.77 | 20.67±0.58 | 160.67±6.66 | 23.33±3.21 | 0.11±0.00 |
| 10 | 246.67±9.24 | 23.00±1.00 | 157.00±6.56 | 16.33±2.31 | 0.08±0.01 |
| 20 | 292.33±37.21 | 26.33±4.16 | 152.67±14.74 | 19.00±1.73 | 0.08±0.01 |
| 50 | 235.00±15.87 | 25.33±2.31 | 147.33±8.96 | 16.00±2.65 | 0.07±0.02 |
Effect of (+)-CH on histological
damage in cerulein-induced CP
CP leads to the destruction of the acini structure,
which is replaced by fibrotic tissue (33,34).
To determine whether (+)-CH could protect pancreatic acini from
cerulein-induced destruction, pancreatic histological changes were
examined. In response to repetitive inflammation, morphological
features of the pancreas in CP demonstrated deformation of
glandular tissues, characterized by loss of acinar cells and
transition to acinar-to-duct metaplasia (Fig. 1B). In contrast, the group
administered (+)-CH at concentrations of 1, 5 and 10 mg/kg revealed
slight edema and acinar cell transformation, which suggested mild
glandular atrophy. The infiltration of immune cells in the (+)-CH
groups was markedly lower than that in the CP group (Fig. 1B).
Effect of (+)-CH on the activation of
PSCs and production of ECM components during CP
α-SMA and extracellular matrix (ECM) proteins are
highly expressed in peri-acinar fibrotic areas during CP
development (35). α-SMA is
regarded as a marker of the activated PSCs (36). α-SMA was stained and the mRNA
expression level of Acta-2 was assessed to investigate the
PSC activation regulatory ability of (+)-CH in CP. In CP tissue,
α-SMA was stained strongly with DAB in the vicinity of the damaged
acini but was expressed at significantly lower levels in tissues in
the (+)-CH-administered groups (Fig.
2A). The mRNA expression of Acta-2 in the CP group was
considerably higher than that in the control group and it was
suppressed in the (+)-CH group (Fig.
2B). During fibrogenesis, ECM protein levels increase
substantially in the fibrotic region and are often accompanied by
morphological alterations (35,36).
To assess the preventive effect of (+)-CH on the production of ECM
components, IF staining and RT-qPCR were performed. Unlike in the
normal pancreas, the structure of type I collagen (Col I) in the CP
tissue was observed to be irregular and thicker and the staining
(green) intensity was also stronger than that of samples from the
normal pancreas. However, overall alterations and Col I deposition
decreased significantly in the pancreas of mice pretreated with
(+)-CH at concentrations of 5 and 10 mg/kg (Fig. 2C). To clarify these results, the
mRNA expression levels of FN1 and different types of
collagens were evaluated. In particular, (+)-CH concentrations of 5
and 10 mg/kg significantly reduced the mRNA expression levels of
FN1 and Col I compared with the control. In contrast,
the decrease in type III (Col III) and type IV (Col
IV) collagens, compared with the control, was significant only
at a concentration of 5 mg/kg (Fig. 2D
and E). As 5 mg/kg (+)-CH demonstrated greater effect than the
other concentrations used for the determination of prophylactic
effects of (+)-CH on CP, 5 mg/kg (+)-CH was considered the optimal
concentration and further experiments were performed using this
concentration.
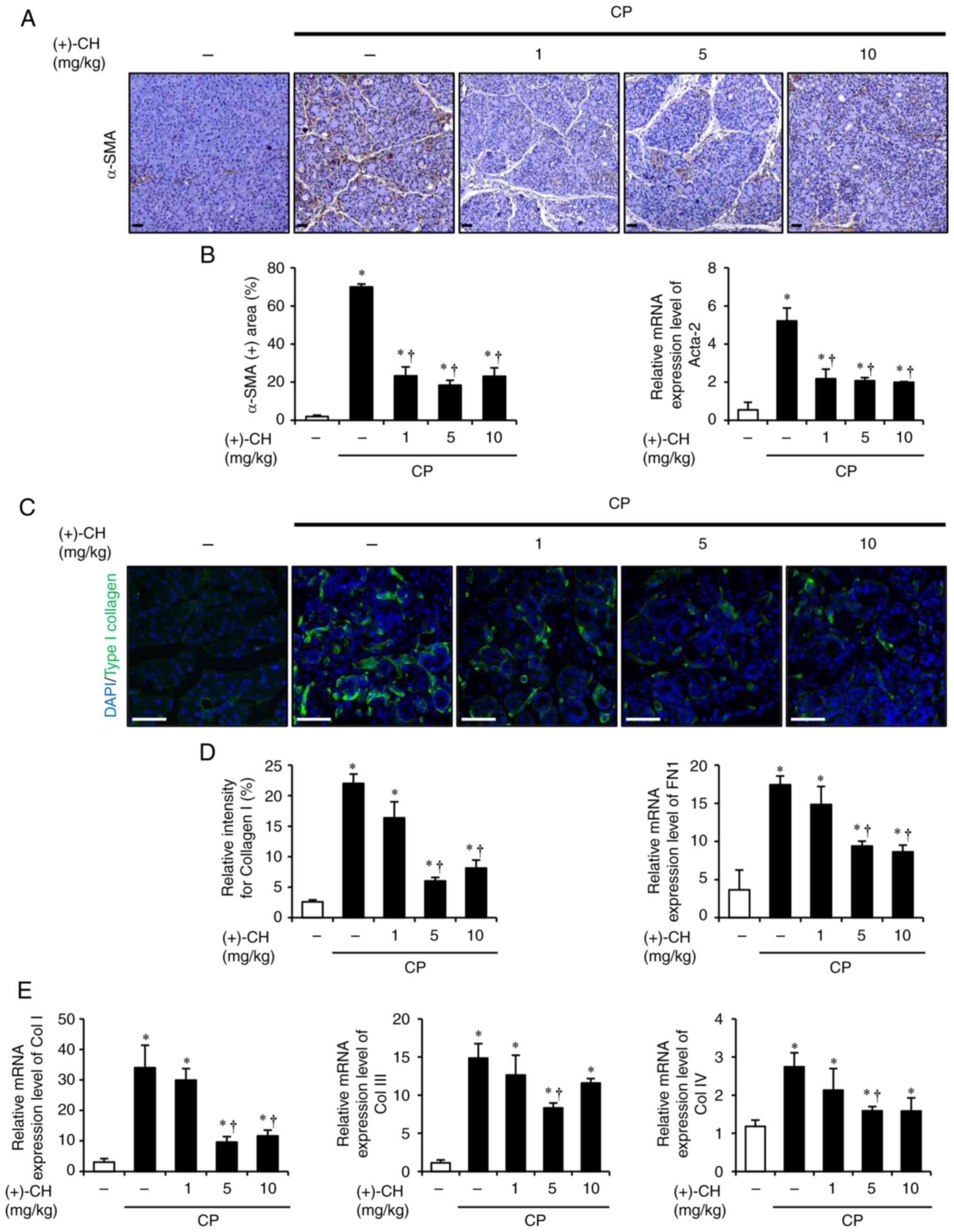 | Figure 2.Effect of (+)-CH on PSC activation
and production of ECM components during CP. (A) Images of
3,3′-diaminobenzidinestaining for α-SMA in the ethanol, CP and
(+)-CH (1, 5 and 10 mg/kg) groups; brown color indicates
α-SMA+ cells. (B) The mRNA expression level of
Acta-2 in the pancreas was assessed using RT-qPCR. (C)
Confocal images of immunofluorescence staining for collagen I in
the ethanol, CP and (+)-CH (1, 5, 10 mg/kg) groups; green color
indicates collagen I+ cells and blue color indicates
DAPI+ cells. (D) mRNA expression of FN 1 in the
pancreas was assessed using RT-qPCR. (E) The mRNA expression levels
of col I, III and IV in the pancreas were assessed
using RT-qPCR. Data are presented as mean ± SEM (n=3/group).
Results are representative of three experiments. *P<0.05 vs.
ethanol alone; †P<0.05 vs. CP alone. Scale bar, 50
µm. CP, chronic pancreatitis; CH, catechin hydrate; PSC, pancreatic
stellate cell; ECM, extracellular matrix; α-SMA/Acta-2, α-smooth
muscle actin; DAPI, 4′,6-diamidino-2-phenylindole; RT-qPCR, reverse
transcription-quantitative PCR; FN1, fibronectin 1; Col,
collagen. |
Effect of (+)-CH on the activation of
isolated PSCs
Activated PSCs are involved in pancreatic
fibrogenesis and are the principal source of α-SMA and ECM proteins
(36,37). Adjusting the activation of cultured
PSCs parallels the regulation of persistent activation of PSCs
in vivo, which is a critical event in fibrosis (37). Transforming growth factor-β (TGF-β)
has been reported to mediate continuous activation of PSCs, and
Smad is part of one of the TGF-β signal transduction pathways
(37,38). As TGF-β drives PSC activation
signal through a Smad2-dependent pathway (39), the protein expression of
phosphorylated Smad2 and total Smad2 was assessed to evaluate
whether (+)-CH regulated TGF-β signaling. It was demonstrated that
(+)-CH significantly inhibited the activation of Smad2 after TGF-β1
treatment in cultured PSCs (Fig.
3A). Furthermore, the regulatory effects of fibrotic factors
that appear as a result of the TGF-β cascade were assessed.
Treatment with TGF-β1 significantly decreased Acta-2, FN1, Col
III and Col IV mRNA expression levels at (+)-CH
concentrations of 150, 200 and 250 µM compared with the control
(Fig. 3B). Moreover, the mRNA
expression level of Col I was significantly reduced only at
a concentration of 250 µM (+)-CH. Changes in the mRNA expression
levels of Acta-2 and ECM components in the PSCs exposed to
platelet derived growth factor (PDGF), contribute to the mitogenic
effect of PSCs (40). Treatment
with (+)-CH significantly decrease the mRNA expression levels of
Acta-2, FN1, Col I, Col III and Col IV compared with
the PDGF-BB-only treatment group (Fig.
3C).
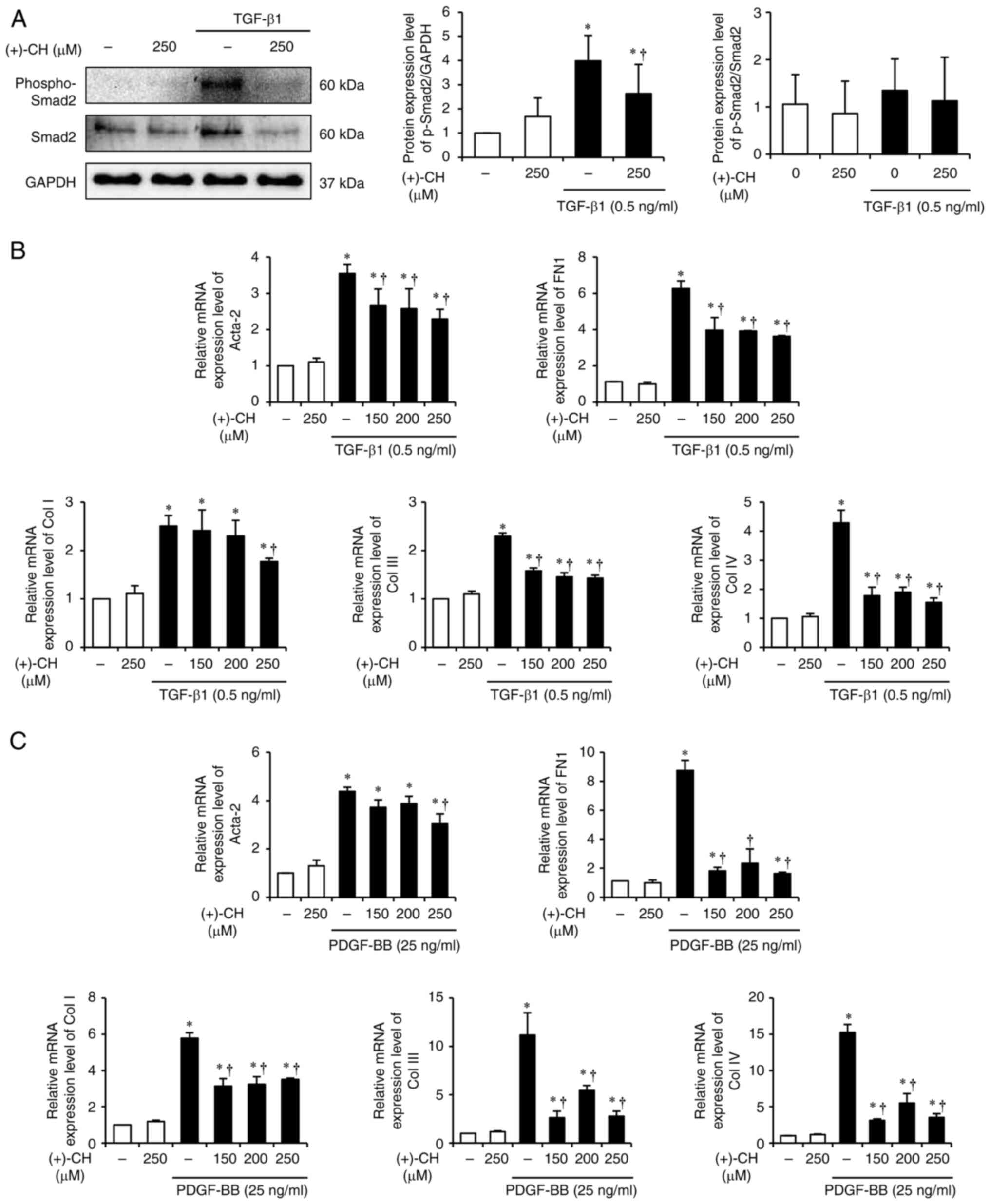 | Figure 3.Effect of (+)-CH on PSC activation in
isolated PSCs. (A) Protein expression levels of p-Smad2
(Ser465/467) and Smad2 were assessed using western blotting; GAPDH
was used as the loading control. (B) The mRNA expression levels of
TGF-β1 induced Acta-2, FN1 and col, III and IV
in isolated PSCs were assessed using RT-qPCR. (C) The mRNA
expression levels of PDGF-BB induced Acta-2, FN 1 and col
I, III and IV in isolated PSCs were assessed using
RT-qPCR. Data are presented as mean ± SEM. Results are
representative of three experiments. *P<0.05 vs. ethanol alone;
†P<0.05 vs. TGF-β1 alone or PDGF-BB alone. CP,
chronic pancreatitis; CH, catechin hydrate; phospho-Smad2/pSmad2,
phosphorylated Smad2; Col, collagen; Acta-2, α-smooth muscle actin;
FN1, fibronectin 1; PDGF-BB, platelet-derived growth factor; p,
phosphorylated; RT-qPCR, reverse transcription-quantitative PCR;
FN1, fibronectin 1; Col, collagen. |
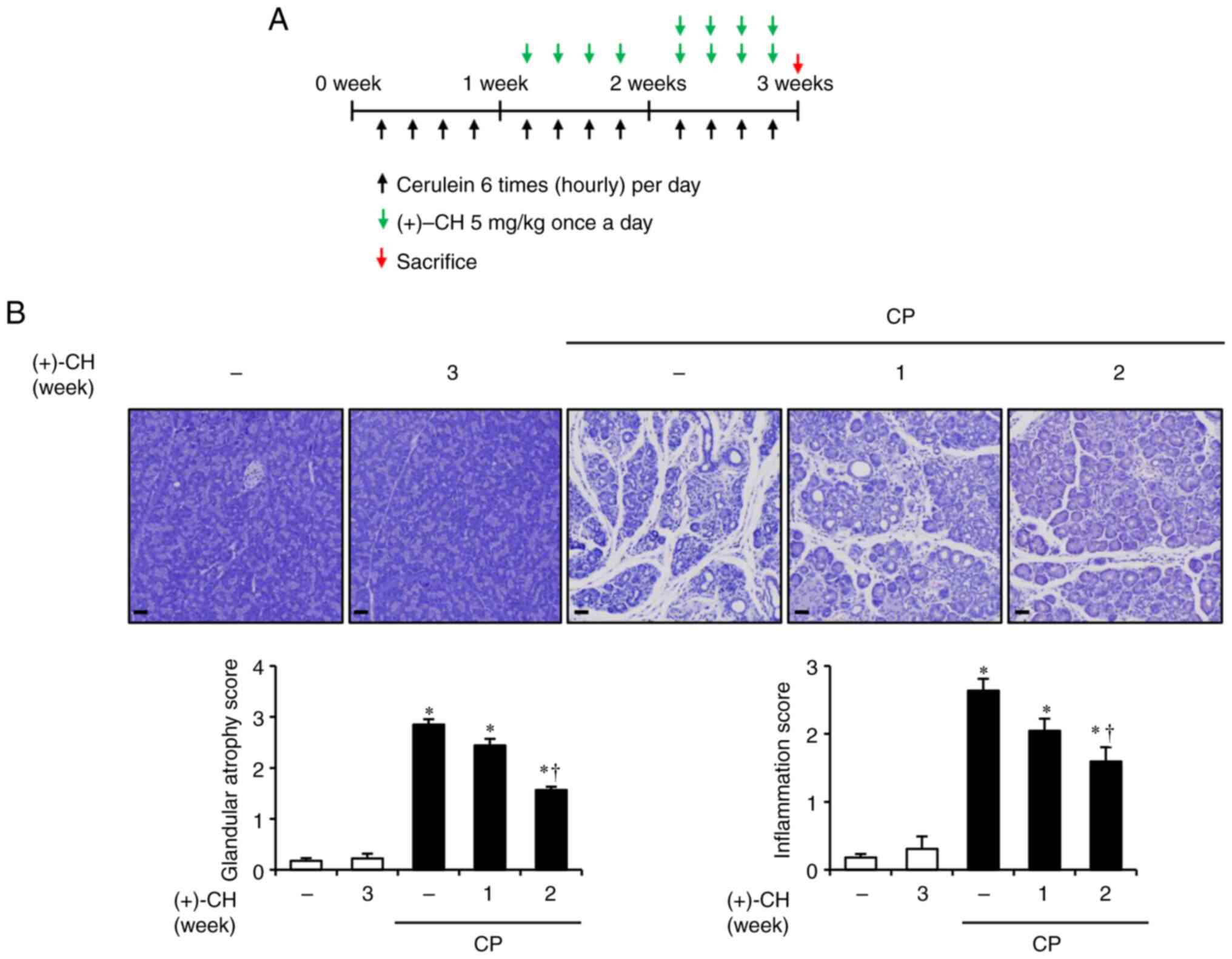
Therapeutic effect of (+)-CH on
CP
Based on earlier findings, whether (+)-CH
demonstrated a therapeutic effect against CP was evaluated.
Treatment with (+)-CH for 1 or 2 weeks alleviated glandular atrophy
and inflammation during CP (Fig.
4). Treatment with (+)-CH for 2 weeks resulted in faint
staining for α-SMA. However, treatment with (+)-CH for 1 week
resulted in a staining as strong as that observed for the CP group
(Fig. 5A). The mRNA expression
level of Acta-2 decreased markedly when (+)-CH was
administered for 1 week and was significantly decreased in the
group with (+)-CH administered for 2 weeks, compared with the
control (Fig. 5B). As for α-SMA,
the deposition of Col I and mRNA expression levels of FN1, Col
I, Col III and Col IV were all significantly reduced in
the group injected with (+)-CH for 2 weeks, compared with the
control (Fig. 5C-E).
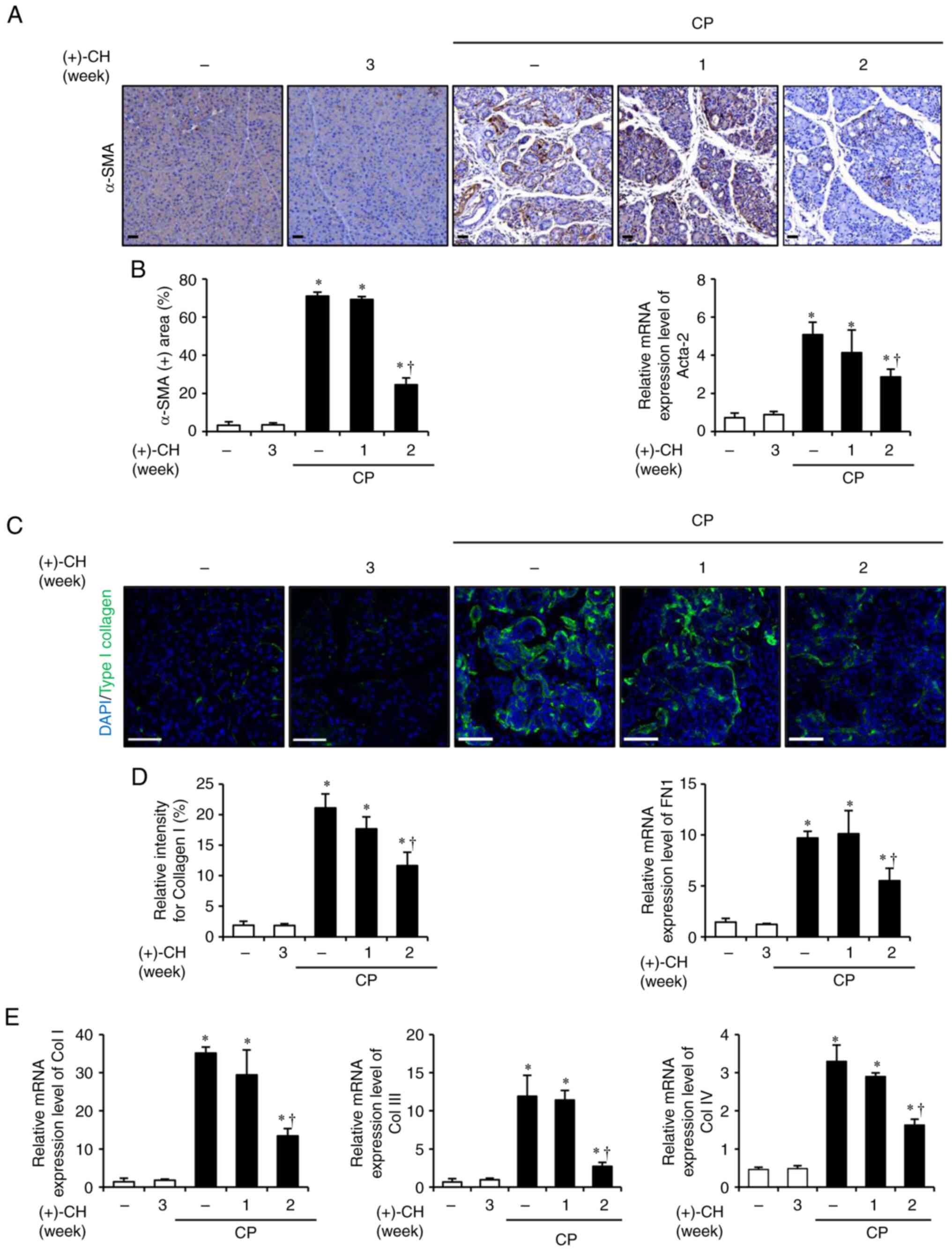 | Figure 5.Therapeutic effect of (+)-CH on
pancreatic fibrosis. (A) Images of 3,3′-diaminobenzidine staining
for α-SMA in the ethanol, (+)-CH alone, CP and 5 mg/kg (+)-CH
groups (1 and 2 weeks); brown color indicates α-SMA+
cells. (B) mRNA expression level of Acta-2 in the pancreas
was assessed using RT-qPCR. (C) Confocal images of
immunofluorescence staining for collagen I in the ethanol, (+)-CH
alone, CP and 5 mg/kg (+)-CH groups (1 and 2 weeks); green color
indicates collagen I+ cells and blue color indicates
DAPI+ cells. (D) mRNA expression level of FN1 in
the pancreas was assessed using RT-qPCR. (E) mRNA expression levels
of col I, III and IV in the pancreas were assessed
using RT-qPCR. Data are presented as mean ± SEM (n=3/group).
Results are representative of three experiments. *P<0.05 vs.
ethanol alone; †P<0.05 vs. CP alone. Scale bar, 50
µm. CP, chronic pancreatitis; CH, catechin hydrate; α-SMA/Acta-2,
α-smooth muscle actin; FN1, fibronectin 1; RT-qPCR, reverse
transcription-quantitative PCR; col, collagen. |
Discussion
CP is a pancreatic inflammatory disease accompanied
by fibrosis and can develop at any life stage from children to
adults, thus greater attention should be paid to it (41). CP is difficult to diagnose in the
early stages of the disease due to ambiguous symptoms such as
weight loss, pain and steatorrhea, something which further impedes
the quality of life of patients (3,42).
Therefore, understanding the process of pancreatic fibrogenesis and
finding a way to treat it are considered primary objectives
(3).
The progression of CP includes irregular distortion
of the glandular parenchyma owing to the formation of a duct-like
structure following pancreatic inflammation, which results in
pancreatic exocrine and endocrine loss (43). Destruction of the pancreatic acini
leads to impairment of pancreatic function and pain, which are
clinical features of CP (44).
Inflammatory infiltrates, which are increased in the pancreas of
patients with CP, affect the proliferation and activation of PSCs
by releasing cytokines and stimulating collagen synthesis to
promote pancreatic fibrosis (45,46).
As glandular tissue destruction and necrosis are essential
prerequisites of fibrotic pancreatic tissues (34), the histological changes, including
acini atrophy and immune cell infiltration in pancreatic tissues,
after (+)-CH injection were assessed and it was demonstrated that
(+)-CH alleviated parenchymal destruction and inflammation at all
tested concentrations.
The main pathological feature of CP is pancreatic
fibrosis and PSCs, resident cells in the inter-acinar regions,
contribute to the pathogenesis of CP in an activated state by
acting as key effector cells in fibrogenesis (37,47,48).
Activated PSCs exhibit morphological changes, including α-SMA
expression and production of ECM proteins such as collagen and FN
(37). α-SMA, which is abundantly
observed in pancreatic fibrotic areas, is the first to be expressed
in the process of fibrosis and is considered a marker of activated
PSCs (38). During fibrogenesis,
acinar cells are destroyed and replaced with connective tissue,
which arises from increased or disorganization of deposited ECM
proteins (35). FN, which is an
omnipresent non-collagen ECM component, participates in a variety
of cellular processes, including proliferation and migration;
excessive deposition of FN leads to organ impairment, which results
in fibrotic diseases (49,50). Overexpression of FN1, which belongs
to the FN family, precedes collagen deposition and is often
considered an indicator of collagen accumulation (46,51).
Col I and Col III, which are the main collagens constituents of the
interstitial matrix, are upregulated in a disorganized state with
FN in CP tissue and are formlessly deposited in thick bundles
(36). Col IV, which is
categorized as a network-forming collagen, is located in the
basement membrane (BM), which binds to numerous factors and is
responsible for forming the backbone of the BM (52). During acinar cell atrophy, Col IV
accumulation varies and is discontinuous in fibrotic areas adjacent
to the BM (50). Given that
collagen production increases in activated PSCs during fibrogenesis
(38), the inhibitory effect of
(+)-CH on the activation of PSCs and synthesis of ECM proteins was
demonstrated in the present study. The upregulated expression of
α-SMA (also known as Acta2), collagens and FN1 and (+)-CH was
demonstrated to have significantly prevented the activation of PSCs
and collagen deposition compared with the CP group. These results
indicated that (+)-CH has a prophylactic effect on CP.
Following stimulation, the acinar cells are injured,
which results in pancreatic inflammation. As a consequence of
pancreatic injury, paracrine factors such as cytokines, growth
factors and intracellular signaling molecules are secreted by the
injured cells (37,53). In response to the paracrine
factors, PSCs are persistently activated and subsequently regulate
the proliferation, migration and differentiation of PSCs, thereby
inducing fibrogenesis (37,53).
Among autocrine and paracrine factors, the growth factors PDGF and
TGF-β are the major factors responsible for the development of
pancreatic fibrosis (38). TGF-β
is secreted mainly in damaged acinar cells adjacent to fibrotic
areas and is a potent promoter of PSC activation (38). Smads, which have been implicated in
the fibrotic effect of TGF-β, can be directly phosphorylated by
TGF-β receptors (54).
Phosphorylated receptor-regulated Smads form a complex with the
common mediator Smad and translocate into the nucleus, thereby
regulating the transcription of target genes (54). TGF-β not only increases α-SMA
expression and collagen synthesis but also enhances PDGF receptor
expression, thereby increasing the responsiveness of PSCs to PDGF,
resulting in continuous PSC activation and permanent fibrosis in
the pancreas (40). PDGF-BB has
been reported to have shown a proliferative effect demonstrated by
increased cell numbers and DNA synthesis in PDGF-BB-exposed PSCs
(40). The cell proliferative
effect of PDGF is also affected by upregulated cell surface PDGF
receptor in fibrotic areas (40).
In cultured PSCs, (+)-CH inhibited Smad2 activity caused by TGF-β
and suppressed production of fibrosis factors. Likewise, the levels
of fibrosis factors acting through PDGF were significantly reduced
in PSCs treated with (+)-CH.
According to the European Food Safety Authority
(Parma, Italy), green tea catechins, particularly EGCG, are
generally considered safe if the amount adheres to the dosage
recommended by European member states (23). In addition, (+) catechin is
classified as having the least toxicity compared with other
catechins, including EGCG and epicatechin gallate (ECG), in normal
HGF-2 fibroblasts (25). To date,
no specific toxicity has been reported for (+) catechin or (+) CH.
An inhibitory effect of EGCG on PSC activation has been reported
previously (19). However, there
are few studies on the protective effects of (+) catechin or (+) CH
against pancreatic fibrosis. Because existing CP treatment, such as
pregabalin can result in adverse side effects include dizziness,
somnolence, blurred vision, even fainting (55,56),
(+) catechin or (+)-CH was anticipated to be an improved
therapeutic agent. Therefore, the therapeutic effect of (+)-CH
following its prophylactic effect against CP was assessed. The
severity of pancreatic tissue damage and fibrosis exhibited in CP
improved when (+)-CH was administered for 2 weeks. These results
provided evidence to support the hypothesis that (+)-CH has a
therapeutic effect on CP, nonetheless, further studies pertaining
to the analysis of safety and efficacy of (+)-CH are needed.
In conclusion, the findings of the present study
demonstrated that (+)-CH inhibited immune cell infiltration and
histological injury of the pancreas. Moreover, the administration
of (+)-CH contributed to a reduction in the levels of
fibrosis-related factors, including Acta-2, FN1, Col I, Col III and
Col IV both in TGF-β1 and PDGF-BB treated PSCs, and in CP. These
results demonstrated that (+)-CH had prophylactic and therapeutic
effects on CP. (+)-CH protected against the progression of
pancreatic fibrosis by inactivating the TGF-β/Smad2 signaling
pathway. Therefore, the results of the present study suggest that
(+)-CH itself or (+)-CH containing products may be a novel
therapeutic strategy for the treatment of CP. However, as efficacy
was confirmed only in cerulein induced CP and the safety issue of
catechins, further studies are required.
Acknowledgements
Not applicable.
Funding
This research was supported by a National Research Foundation of
Korea grant funded by the Korean government (grant no.
NRF-2017R1A5A2015805/2021R1I1A2053285).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
BK and DUK validated the data, performed the
experiments, wrote the original draft of the manuscript. JYO
performed the experiments. GSB and SJP conceptualized and
supervised the study and performed project administration. BK, DUK,
JYO, GSB and SJP confirm the authenticity of all the raw data. All
authors have read and approved the final manuscript.
Ethics approval and consent to
participate
All experiments were performed in accordance with
the animal care regulations set forth and approved by The Wonkwang
University Animal Ethics Committee (Iksan, Republic of Korea;
approval no. WKU22-43).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Yang HY, Wang D, Liu Y, Han C, Xin L, Li
ZS, Liao Z and Hu LH: Global status in chronic pancreatitis
research. Pancreas. 49:1283–1289. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Olesen SS, Mortensen LH, Zinck E, Becker
U, Drewes AM, Nøjgaard C, Novovic S, Yadav D and Tolstrup JS: Time
trends in incidence and prevalence of chronic pancreatitis: A
25-year population-based nationwide study. United European
Gastroenterol J. 9:82–90. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Etemad B and Whitcomb DC: Chronic
pancreatitis: Diagnosis, classification, and new genetic
developments. Gastroenterology. 120:682–707. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Olesen SS, Frandsen LK, Poulsen JL,
Vestergaard P, Rasmussen HH and Drewes AM: The prevalence of
underweight is increased in chronic pancreatitis outpatients and
associates with reduced life quality. Nutrition. 43–44. 1–7.
2017.
|
|
5
|
Lin CH, Yeh NC, Wang JJ, Ho CH, Her SH,
Tsay WI and Chien CC: Effect of chronic pancreatitis on
complications and mortality in DM patients: A 10-year nationwide
cohort study. J Clin Endocrinol Metab. 105:e739–e745. 2020.
View Article : Google Scholar
|
|
6
|
Drewes AM, Krarup AL, Detlefsen S,
Malmstrøm ML, Dimcevski G and Funch-Jensen P: Pain in chronic
pancreatitis: The role of neuropathic pain mechanisms. Gut.
57:1616–1627. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Madro A: Malnutrition in chronic
pancreatitis: Causes, assessment methods, and therapeutic
management. Can J Gastroenterol Hepatol.
2020:88754872020.PubMed/NCBI
|
|
8
|
Kirkegård J, Mortensen FV and
Cronin-Fenton D: Chronic pancreatitis and pancreatic cancer risk: A
systematic review and meta-analysis. Am J Gastroenterol.
112:1366–1372. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Malka D, Hammel P, Sauvanet A, Rufat P,
O'Toole D, Bardet P, Belghiti J, Bernades P, Ruszniewski P and Lévy
P: Risk factors for diabetes mellitus in chronic pancreatitis.
Gastroenterology. 119:1324–1332. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Han M, Tran TPT and Oh JK: Chronic
pancreatitis and cancer risk in a matched cohort study using
national claims data in South Korea. Sci Rep. 12:55452022.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Muniraj T, Aslanian HR, Farrell J and
Jamidar PA: Chronic pancreatitis, a comprehensive review and
update. Part I: Epidemiology, etiology, risk factors, genetics,
pathophysiology, and clinical features. Dis Mon. 60:530–550. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dufour MC and Adamson MD: The epidemiology
of alcohol-induced pancreatitis. Pancreas. 27:286–290. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Koehn FE and Carter GT: The evolving role
of natural products in drug discovery. Nat Rev Drug Discov.
4:206–220. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chacko SM, Thambi PT, Kuttan R and
Nishigaki I: Beneficial effects of green tea: A literature review.
Chin Med. 5:132010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Khan N and Mukhtar H: Tea polyphenols for
health promotion. Life Sci. 81:519–533. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kim HK, Yang TH and Cho HY: Antifibrotic
effects of green tea on in vitro and in vivo models of liver
fibrosis. World J Gastroenterol. 15:5200–5205. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Higdon JV and Frei B: Tea catechins and
polyphenols: Health effects, metabolism, and antioxidant functions.
Crit Rev Food Sci Nutr. 43:89–143. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Isemura M: Catechin in human health and
disease. Molecules. 24:5282019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Asaumi H, Watanabe S, Taguchi M, Tashiro
M, Nagashio Y, Nomiyama Y, Nakamura H and Otsuki M: Green tea
polyphenol (−)-epigallocatechin-3-gallate inhibits ethanol-induced
activation of pancreatic stellate cells. Eur J Clin Invest.
36:113–122. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cao Y, Bao S, Yang W, Zhang J, Li L, Shan
Z and Teng W: Epigallocatechin gallate prevents inflammation by
reducing macrophage infiltration and inhibiting tumor necrosis
factor-α signaling in the pancreas of rats on a high-fat diet. Nutr
Res. 34:1066–1074. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wei R, Cortez Penso NE, Hackman RM, Wang Y
and Mackenzie GG: Epigallocatechin-3-gallate (EGCG) suppresses
pancreatic cancer cell growth, invasion, and migration partly
through the inhibition of Akt pathway and epithelial-mesenchymal
transition: Enhanced efficacy when combined with gemcitabine.
Nutrients. 11:18562019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kim JM and Heo HJ: The roles of catechins
in regulation of systemic inflammation. Food Sci Biotechnol.
31:957–970. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
EFSA Panel on Food Additives and Nutrient
Sources added to Food (ANS), . Younes M, Aggett P, Aguilar F,
Crebelli R, Dusemund B, Filipič M, Frutos MJ, Galtier P, Gott D, et
al: Scientific opinion on the safety of green tea catechins. EFSA
J. 16:e052392018.PubMed/NCBI
|
|
24
|
Alshatwi AA, Hasan TN, Alqahtani AM, Syed
NA, Shafi G, Al-Assaf AH and Al-Khalifa AS: Delineating the
anti-cytotoxic and anti-genotoxic potentials of catechin hydrate
against cadmium toxicity in human peripheral blood lymphocytes.
Environ Toxicol Pharmacol. 38:653–662. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Babich H, Krupka M, Nissim HA and
Zuckerbraun HL: Differential in vitro cytotoxicity of
(−)-epicatechin gallate (ECG) to cancer and normal cells from the
human oral cavity. Toxicol In Vitro. 19:231–242. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Alshatwi AA: Catechin hydrate suppresses
MCF-7 proliferation through TP53/Caspase-mediated apoptosis. J Exp
Clin Cancer Res. 29:1672010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kweon B, Kim DU, Oh JY, Oh H, Kim YC, Mun
YJ, Bae GS and Park SJ: Arecae pericarpium water extract alleviates
chronic pancreatitis by deactivating pancreatic stellate cells.
Front Pharmacol. 13:9419552022. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Rai A, Das S, Chamallamudi MR, Nandakumar
K, Shetty R, Gill M, Sumalatha S, Devkar R, Gourishetti K and Kumar
N: Evaluation of the aphrodisiac potential of a chemically
characterized aqueous extract of Tamarindus indica pulp. J
Ethnopharmacol. 210:118–124. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shahid A, Ali R, Ali N, Hasan SK, Bernwal
P, Afzal SM, Vafa A and Sultana S: Modulatory effects of catechin
hydrate against genotoxicity, oxidative stress, inflammation and
apoptosis induced by benzo(a)pyrene in mice. Food Chem Toxicol.
92:64–74. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bragança de Moraes CM, Bitencourt S, de
Mesquita FC, Mello D, de Oliveira LP, da Silva GV, Lorini V,
Caberlon E, de Souza Basso B, Schmid J, et al: (+)-Catechin
attenuates activation of hepatic stellate cells. Cell Biol Int.
38:526–530. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zang G, Sandberg M, Carlsson PO, Welsh N,
Jansson L and Barbu A: Activated pancreatic stellate cells can
impair pancreatic islet function in mice. Ups J Med Sci.
120:169–180. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Steer ML, Waxman I and Freedman S: Chronic
pancreatitis. N Engl J Med. 332:1482–1490. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zimmermann A, Gloor B, Kappeler A, Uhl W,
Friess H and Büchler MW: Pancreatic stellate cells contribute to
regeneration early after acute necrotising pancreatitis in humans.
Gut. 51:574–578. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gress T, Menke A, Bachem M,
Müller-Pillasch F, Ellenrieder V, Weidenbach H, Wagner M and Adler
G: Role of extracellular matrix in pancreatic diseases. Digestion.
59:625–637. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kennedy RH, Bockman DE, Uscanga L, Choux
R, Grimaud JA and Sarles H: Pancreatic extracellular matrix
alterations in chronic pancreatitis. Pancreas. 2:61–72. 1987.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Omary MB, Lugea A, Lowe AW and Pandol SJ:
The pancreatic stellate cell: A star on the rise in pancreatic
diseases. J Clin Invest. 117:50–59. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Haber PS, Keogh GW, Apte MV, Moran CS,
Stewart NL, Crawford DH, Pirola RC, McCaughan GW, Ramm GA and
Wilson JS: Activation of pancreatic stellate cells in human and
experimental pancreatic fibrosis. Am J Pathol. 155:1087–1095. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ohnishi H, Miyata T, Yasuda H, Satoh Y,
Hanatsuka K, Kita H, Ohashi A, Tamada K, Makita N, Iiri T, et al:
Distinct roles of Smad2-, Smad3-, and ERK-dependent pathways in
transforming growth factor-beta1 regulation of pancreatic stellate
cellular functions. J Biol Chem. 279:8873–8878. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Apte MV, Haber PS, Darby SJ, Rodgers SC,
McCaughan GW, Korsten MA, Pirola RC and Wilson JS: Pancreatic
stellate cells are activated by proinflammatory cytokines:
Implications for pancreatic fibrogenesis. Gut. 44:534–541. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Schwarzenberg SJ and Whitcomb DC: Causes
and contributing risk factors for chronic pancreatitis in children
and adolescents. UpToDate in Medicine.
|
|
42
|
Duggan SN, Ni Chonchubhair HM, Lawal O,
O'Connor DB and Conlon KC: Chronic pancreatitis: A diagnostic
dilemma. World J Gastroenterol. 22:2304–2313. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Klöppel G and Maillet B: Pathology of
acute and chronic pancreatitis. Pancreas. 8:659–670. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Aghdassi AA, Mayerle J, Christochowitz S,
Weiss FU, Sendler M and Lerch MM: Animal models for investigating
chronic pancreatitis. Fibrogenesis Tissue Repair. 4:262011.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zheng L, Xue J, Jaffee EM and Habtezion A:
Role of immune cells and immune-based therapies in pancreatitis and
pancreatic ductal adenocarcinoma. Gastroenterology. 144:1230–1240.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Saotome T, Inoue H, Fujimiya M, Fujiyama Y
and Bamba T: Morphological and immunocytochemical identification of
periacinar fibroblast-like cells derived from human pancreatic
acini. Pancreas. 14:373–382. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Bachem MG, Schneider E, Gross H,
Weidenbach H, Schmid RM, Menke A, Siech M, Beger H, Grünert A and
Adler G: Identification, culture, and characterization of
pancreatic stellate cells in rats and humans. Gastroenterology.
115:421–432. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Apte MV, Haber PS, Applegate TL, Norton
ID, McCaughan GW, Korsten MA, Pirola RC and Wilson JS: Periacinar
stellate shaped cells in rat pancreas: Identification, isolation,
and culture. Gut. 43:128–133. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
To WS and Midwood KS: Plasma and cellular
fibronectin: Distinct and independent functions during tissue
repair. Fibrogenesis Tissue Repair. 4:212011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Yamaguchi T, Kihara Y, Taguchi M, Nagashio
Y, Tashiro M, Nakamura H and Otsuki M: Persistent destruction of
the basement membrane of the pancreatic duct contributes to
progressive acinar atrophy in rats with experimentally induced
pancreatitis. Pancreas. 31:365–372. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Liu B, Ding Y, Li P, Wang T, He S, Jia Z
and Yang J: MicroRNA-219c-5p regulates bladder fibrosis by
targeting FN1. BMC Urol. 20:1932020. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Sand J, Genovese F and Karsdal M: Type IV
collagen. Biochemistry of Collagens, Laminins and Elastin.
Elsevier; Amsterdam: pp. 31–41. 2016, View Article : Google Scholar
|
|
53
|
Hu LH, Ji JT and Li ZS: Potential
application of miRNAs as diagnostic and therapeutic tools in
chronic pancreatitis. J Cell Mol Med. 19:2049–2057. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Attisano L and Wrana JL: Signal
transduction by the TGF-beta superfamily. Science. 296:1646–1647.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Gurusamy KS, Lusuku C and Davidson BR:
Pregabalin for decreasing pancreatic pain in chronic pancreatitis.
Cochrane Database Syst Rev. 2:CD0115222016.PubMed/NCBI
|
|
56
|
Kwon CI, Cho JH, Choi SH, Ko KH, Tirkes T,
Gromski MA and Lehman GA: Recent advances in the diagnosis and
management of chronic pancreatitis. Korean J Intern Med.
34:242–260. 2019. View Article : Google Scholar : PubMed/NCBI
|