Introduction
Lipid rafts are specialized domains rich in
cholesterol and sphingolipids in the cell membranes that serve as
physical platforms for a range of molecules to adjust the processes
of various signal-transducing molecules (1,2).
Flotillin proteins (FLOTs) are the primary proteins isolated from
lipid rafts. They are highly conserved proteins associated with
cell membranes and are part of the protein family that includes a
Stomatin-Prohibitin-Flotillin-HflK/C (SPFH) domain. The Flotillin
proteins (also known as the Reggie family) consist of two
homologous isoforms, Flotillin-1(FlOT1)/Reggie-2 and
Flotillin-2(FLOT2)/Reggie-1, which share 50% of the same amino acid
sequences, and both of which physically interact with each other to
form oligomeric and/or heterodimeric complexes (3,4).
Additionally, both FLOTs have been used as markers of lipid rafts,
and initiate receptor kinase signaling (5). FLOTs are plasma membrane
(PM)-associated proteins in lymphocytes, neurons, and other cell
types, as well as serving as scaffold proteins in non-vacuolar
lipid raft microdomains (6).
FLOT1 is composed of 428 amino acids, and has a
flotillin domain and a prohibition homology domain (PHB domain)
(2). It is generally expressed in
nearly all cell types (2), and is
primarily localized at the PM, but is also present in Golgi,
lysosomes, phagosomes, nuclei, endocytic compartments (7,8), as
well as in early endosomes and extracellular vehicles (EVs)
(9,10). Of note, the palmitoylation and
phosphorylation of FLOT1 can alter its subcellular localization
(11,12), and aberrant modification of FLOT1
is involved in promoting the progression of cervical cancer (CC)
(13). Additionally, FLOT1 is a
membrane protein that can be endocytosed from the PM to the
intracellular compartments (14).
In lipid rafts, FLOT1 is related to the formation of discrete
planar microdomains (1).
Naturally, FLOT1 has several raft-related functions, such as
promoting the endocytosis of dopamine (DA) transporter (DAT)
(15), glial glutamate transporter
(EAAT2) (16), insulin-like growth
factor-1 (IGF-1) receptor (IGF-1R) (17), muscarinic type 3 receptor (M3R)
(7), PrPC (6), certain glycosylphosphatidylinositol
(GPI)-anchored proteins and certain proteins that participate in
signal transduction and intracellular transport (15,18).
Additionally, FLOT1 can also mediate clathrin-independent
endocytosis (CIE) and the formation of hippocampal synapses
(2,17). Moreover, FLOT1 is a marker of
exosomes (19), participating in
membrane trafficking. However, the abnormal expression of FLOT1 can
lead to abnormal endocytosis, which induces certain
neurodegenerative diseases, such as Parkinson's disease (PD)
(16), and transmissible
spongiform encephalopathy (TSEs) (6).
In the present review, the role of FLOT1 in human
diseases by summarizing its structure, localization, physiological
function, and mechanisms that contribute to human diseases
including cancers, neurological diseases, dilated cardiomyopathy,
pathogenic microbial infection, diabetes-related diseases,
gynecological diseases and other diseases.
Structure and localization of FLOT1
The FLOT1 gene, present in chromosome 6,
contains 13 exons and encodes a protein consisting of 427 residues.
The FLOT1 gene can be silenced by the system of clustered
regularly interspaced short palindromic repeats (CRISPR)-associated
sequence 9 (CRISPR/Cas9), and altered splicing products can also
produce abnormal protein products (20). FLOT1 and FLOT2 belong to the SPFH
protein superfamily; they have a common N-terminal SPFH domain
without clear understanding of its corresponding function. The
C-terminus of FLOT1 and FLOT2 are longer than other SPFH proteins
as well as being longer than the flotillin domain. The flotillin
domain is characterized by the presence of glutamate-rich and
alanine-rich repeat sequences, which are expected to form three
coiled-coil stretches (21). FLOT1
also has a highly conserved PHB domain, which spans amino acids
1–154 and the flotillin domain spans amino acids 190–363 (22). FLOT1 and FLOT2 can interact with
each other to form oligomeric and/or heterodimeric complexes
(4). It has been shown that
proteasome degradation occurs in FLOT1 in the absence of FLOT2;
therefore stable FLOT1 protein expression requires the presence of
FLOT2. However, the membrane association of FLOT1 is stronger than
that of FLOT2, likely given the second hydrophobic stretch in the
SPFH domain (21).
N-methyl-d-aspartate receptors (NMDARs) are
glutamate receptors that regulate the transmission of excitatory
synaptic potentials in the brain, which are primarily composed of
NR2A and NR2B subunits. NR2B can bind to both FLOT1 and FLOT2,
while NR2A interacts directly with FLOT1 only. NR2A and NR2B both
interact with FLOT1 or FLOT2 at different subcellular localizations
via the PHB domain. In addition, the interaction between NMDARs and
FLOT1 seems to be stronger than that with FLOT2 (22).
PTMs of FLOT1
Palmitoylation, sumoylation and phosphorylation of
proteins are reversible post-translational modifications (PTMs).
FLOT1 is palmitoylated at Cys34, and sumolylated by UBC9 at Lys15
or Lys195. In addition, FLOT1 can be phosphorylated by protein
kinase C (PKC) at Ser315 and by Fyn, and a type of Src kinase at
Tyr160 (5,12,15).
Palmitoylation and phosphorylation play important roles in protein
subcellular localization (Fig.
1).
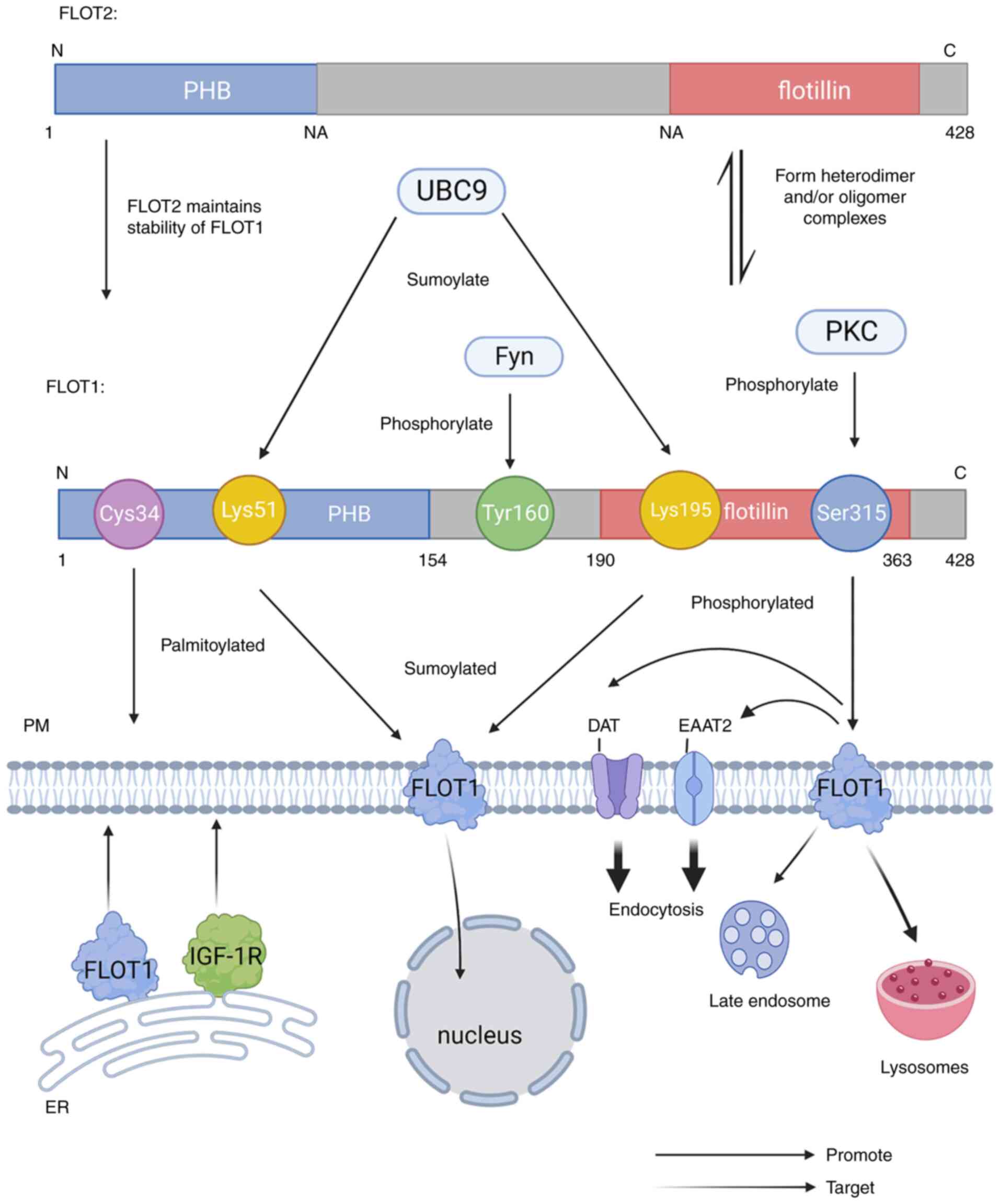 | Figure 1.The Structure of FLOT1 and FLOT2: The
C-terminus of FLOT1 and FLOT2 are longer than other SPFH proteins,
includes a flotillin domain, and the N-terminus also has a highly
conserved PHB domain. The PHB domain of FLOT1 spans amino acids
1–154 and the flotillin domain encompasses amino acids 190–363.
FLOT1 and FLOT2 interact with each other to form heterodimeric
and/or oligomeric complexes. In addition, FLOT1 requires the
presence of FLOT2 to be stable at the protein level. FLOT1 can be
sumolylated by UBC9 at Lys15 and Lys195, and phosphorylated by PKC
at Ser315 and by Fyn in Tyr160. FLOT, flotillin protein; PHB,
prohibition homology; PKC, protein kinase C; IGF-1R, insulin-like
growth factor-1 receptor; DAT, dopamine transporter; EAAT2, glial
glutamate transporter. |
Palmitoylation of FLOT1
FLOT1 is palmitoylated at Cys34, a conserved
cysteine residue in the PHB domain, and the palmitoylation of FLOT1
is indispensable for PKC-triggered endocytosis of DAT (15). In addition, the palmitoylation of
FLOT1 in the endoplasmic reticulum (ER) is indispensable for FLOT1
targeting to the PM with IGF-1R, which changes the subcellular
localization of FLOT1 (11). One
study showed that desmoglin 2 (Dsg2) function can be eliminated by
mutated forms (Dsg2cacs) that fail to be palmitoylated, resulting
in reduced subcellular localization of FLOT1 or other membrane raft
proteins, which is essential for the transport of early endosomal
and membrane raft proteins, thereby modulating the release of EVs
(10).
Phosphorylation of FLOT1
Ser315 of FLOT1 can be phosphorylated by activated
PKC, and this phosphorylation can promote the endocytosis of DAT
and EAAT2 (15). A study showed
that the redistribution of FLOT1 from the PM to lysosomes and late
endosomes was induced by the expression of an active form of Fyn,
which is relevant to the phosphorylation of FLOT1 by Src kinase.
The mutation of Tyr160 in FLOT1 to phenylalanine prevents
Fyn-induced FLOT1 internalization (12). In addition, Tyr160 of FLOT1 is
phosphorylated by Src kinase, and following this phosphorylation,
FLOT1 is endocytosed into late endosomes after stimulation by
epidermal growth factor (EGF) (23).
Sumoylation of FLOT1
As ubiquitin-associated proteins, small
ubiquitin-associated modifiers (SUMOs) can regulate protein
function by covalently conjugating to lysine residues in a large
number of proteins (24).
Upregulated E2 conjugating enzyme UBC9 can mediate the sumoylation
of FLOT1 at Lys51 and Lys195 with small ubiquitin-like modifier
(SUMO)-2/3 modification. The sumoylation of FLOT1 can trigger its
nuclear heterotopic, which is related to the occurrence of prostate
cancer (PCa) (5). In addition, one
study has shown that the sumoylation of FLOT1 participates in
modulating synaptic plasticity (25) (Table
I).
 | Table I.Regulators of FLOT1. |
Table I.
Regulators of FLOT1.
| First author/s,
year | Regulation | Regulator | Mode of
regulation | (Refs.) |
|---|
| Liu et al,
2019 | Translational
regulation | HOTAIR | The upregulation of
HOTAUR promotes the expression of FLOT1 in HCC. | (42) |
| Cai et al,
2021 |
| A1BG-AS1 | The upregulation of
A1BG-AS1 promotes the expression of FLOT1 in BC. | (44) |
| Lv et al,
2020 |
| TUG1 | The upregulation of
TUG1 promotes the expression of FLOT1 in ccRCC. | (45) |
| Wang et al,
2022 |
| FAM201A | The upregulation of
FAM201A promotes the expression of FLOT1 in CC. | (59) |
| Li et al,
2021 |
| SNHG6 | The upregulation of
SNHG6 promotes the expression of FLOT1 in Malignant glioma. | (60) |
| Liu et al,
2019, |
| miR-214-3p, | The upregulation of
miR-214-3p and miR-6809-5p | (42,61) |
| Yang et al,
2019 |
| miR-6809-5p | inhibits the
expression of FLOT1 in HCC. |
|
| Li et al,
2013, |
| miR-485-5p, | The upregulation of
miR-485-5p and miR-124 | (44,62) |
| Cai et al,
2021 |
| miR-124 | inhibits the
expression of FLOT1 in BC. |
|
| Yang et al,
2015, |
| miR-31-5p, | The upregulation of
miR-31-5p, miR-506 and | (45,56) |
| Lv et al,
2020 |
| miR-506,
miR-124-3p | miR-124-3p inhibits
the expression of FLOT1 in ccRCC. |
|
| Kan et al,
2020, |
| miR-1271-5p, | The upregulation of
miR-1271-5p and miR-1294 | (43,59) |
| Wang et al,
2022 |
| miR-1294 | inhibits the
expression of FLOT1 in CC. |
|
| Gong et al,
2013 |
| miR-138 | The upregulation of
miR-138 inhibits the expression of FLOT1 in ESCC. | (63) |
| Jang et al,
2019 | Post- | UBC9 | UBC9 sumolylates
FLOT1 at Lys15 and Lys195. | (5) |
| Cremona et
al, 2011 | translational
modification | PKC | PKC phosphorylates
FLOT1 at Ser315 | (15) |
| Riento et
al, 2009 |
| Fyn | Fyn phosphorylates
FLOT1 in Tyr160. | (12) |
Physiological effects of FLOT1
Lipid raft protein-dependent or
clathrin-independent endocytosis
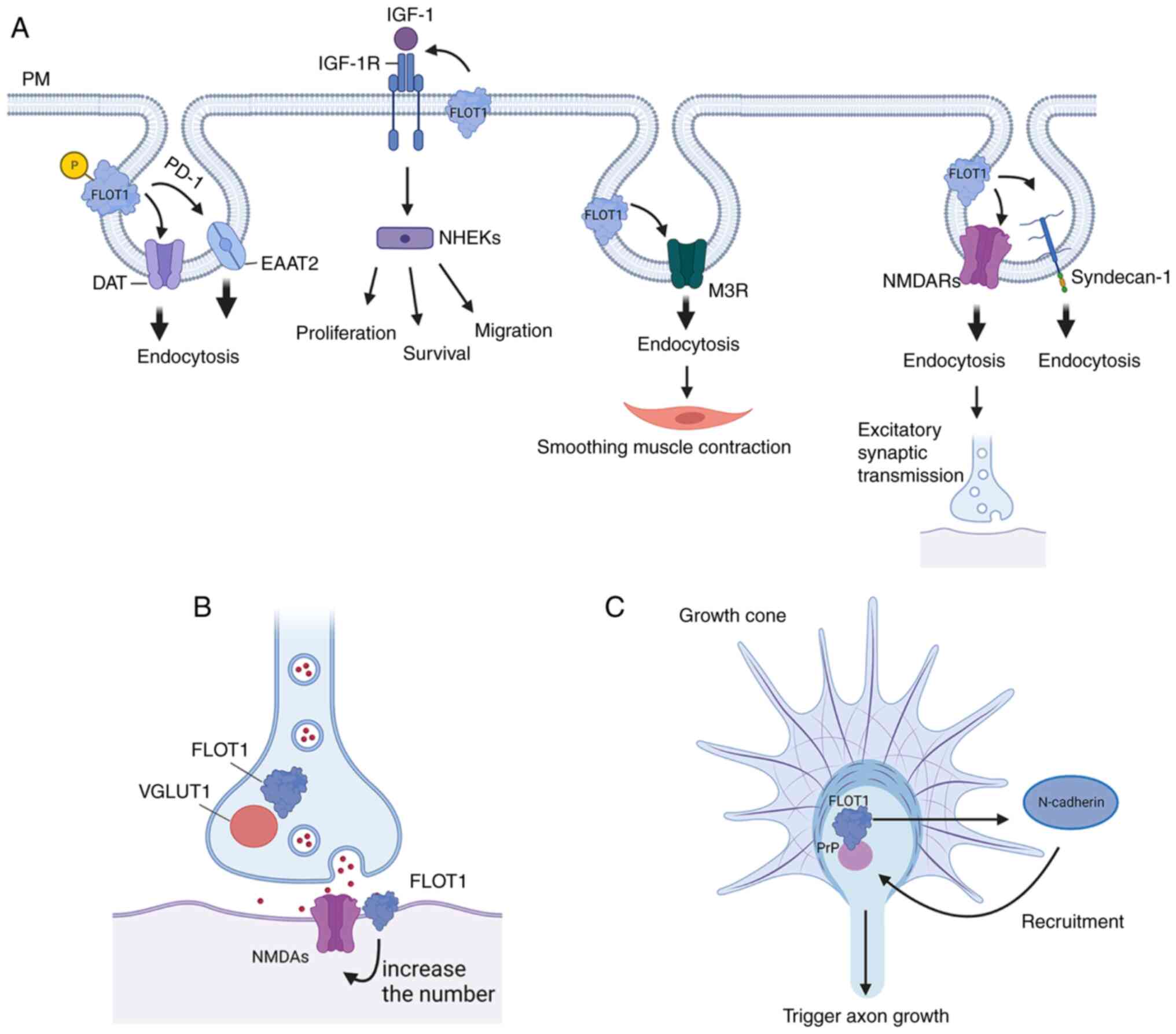
FLOT1 is a pivotal regulator of CIE pathways
(22). The cargoes of the
FLOT1-dependent pathway involved in CIE regulation are principally
certain GPI-anchored proteins, such as PrPC, cholera
toxin B, CD59, and Thy1, which positionally colocalize with FLOT1
at the PM microdomains (6,26). FLOT1 and FLOT2 can form homo- and
hetero-oligomers, and the latter is necessary for their endocytosis
(23). Endocytosis is a major
regulatory factor in the transmission of constitutive signals from
the cell surface to the cytoplasm and nucleus (17), maintaining cell homeostasis,
nutrient absorption, drug transport, and receptor signaling
regulation (7,27).
Firstly, FLOT1 is necessary for PKC-regulated
endocytosis of DAT and EAAT2. Mechanistically, activated PKC can
phosphorylate Ser315 of FLOT1 to promote endocytosis of DAT and
EAAT2, rather than directly phosphorylating the transporter.
Additionally, FLOT1 can maintain DAT in membrane rafts, and it is
required for the reverse transport of DA, although it does not
impact the DAT-mediated uptake of DA (15). In addition, the endocytosis of
EAAT2 induced by FLOT1 is promoted by Parkinson's disease protein 7
(DJ-1), which is an early-onset autosomal recessive gene associated
with PD (16). In addition, the
IGF-1R signaling pathway can promote the proliferation, migration,
and survival of keratinocytes (NHEKs). IGF-1 triggers endocytosis
by activating IGF-1R, which is involved in regulating continuous
signaling from the cell surface to the nucleus and cytoplasm. In
human embryonic kidney cell lines, IGF-1R colocalizes with FLOT1.
The endocytosis of IGF-1R is mediated by FLOT1 in lipid rafts and
the AP2A1/2 complex located in clathrin vesicles of inclusion
complexes. Notably, FLOT1-mediated endocytosis of IGF-1R is more
sensitive when IGF-1R presence is low compared with the classical
AP2A1/2 complex pathway, thus promoting rapid recovery of IGF-1R to
regulate IGF-1R signaling and stimulate a more durable ligand
response. Therefore, FLOT1-mediated endocytosis provides a novel
avenue for targeted therapy in diseases where IGF-1R signaling is
dysregulated (17). Similarly, as
a G-protein-coupled receptor (GPCR) located in the PM, M3R is
highly expressed in salivary glands and is related to physiological
activities such as smoothing the contraction of muscle and salivary
secretion. In addition, M3R can enter the cell by clathrin-mediated
endocytosis (CME), while FLOT1 and FLOT2 are internalized by CIE. A
study showed that FLOT1 and FLOT2 are partially related to the CME
of M3R by promoting the internalization of M3R. However, the
knockdown of FLOT1 or FLOT2 by siRNA reduces the CME of M3R.
Therefore, FLOT1 and FLOT2 of salivary gland epithelial cells may
play a role in the GPCR-mediated pathway (7). Additionally, PrPC forms a
complex with FLOT1 under the stimulation of Cu2+ in the
human neuroblastoma cells, and the PrPC-FLOT1 complex is
transferred from the cell membrane to the cytoplasm under the
treatment of Cu2+. However, the downregulation of FLOT1
in the human neuroblastoma cells notably eliminated the
Cu2+-stimulated endocytosis process of PrPC.
Therefore, the PrPC-FLOT1 complex may be involved in
PrPC transport and endocytosis (6). In addition, FLOT1 can promote the
degradation of anaplastic lymphoma kinase (ALK) in lysosomes
through CIE. A related study also showed that the overexpression of
FLOT1 promoted the endocytosis of ALK, while FLOT1 knockdown
disrupted the lysosomal marker LAMP2 to inhibit the degradation of
ALK, thus increasing the amount of ALK on the cell surface
(28). Moreover, FLOT1 can promote
the endocytosis of Syndecan-1, which is a receptor for C-TRLs
(residual apolipoprotein B rich in cholesterol and triglycerides)
(29). In addition, FLOT1 can
trigger the endocytosis of α-synuclein (α-SYN), and the
accumulation of α-SYN is a neuropathological hallmark of PD
(30). Finally, FLOT1 may promote
the internalization of NMDARs, which mediate excitatory synaptic
transmission in the brain (22)
(Fig. 2).
Formation of hippocampal synapses
In the nervous system, the development of
hippocampal neurons plays an important role in learning and memory
(31). Lipid rafts are a key
factor affecting synaptic formation, while synaptic malformations
are the basis of neurodevelopmental disease. Intact lipid rafts are
necessary to maintain synaptic stability (18). The lipid raft-associated protein
FLOT1 is directly related to synaptic plasticity (25). It plays an important role in
promoting hippocampal neuronal differentiation and neurite growth
in the early stages of neuronal development. FLOT1 colocalized with
the glutamatergic presynaptic marker vesicular glutamate
transporter 1 (VGLUT1) and synaptic NR1 (the obligatory subunit of
NMDA receptors). Of note, overexpression of FLOT1 resulted in an
increase in the presence of glutamatergic synapses, suggesting that
FLOT1 is associated with the formation and induction of
glutamatergic synapses. However, it should be noted that FLOT1 does
not affect GABAergic synapses, suggesting that FLOT1 is a molecular
target for regulating glutamatergic synaptogenesis (2). In addition, it has been shown that
the frequency of miniature excitatory postsynaptic currents
(mEPSCs) is increased by FLOT1 rather than miniature inhibitory
postsynaptic currents (mIPSCs) (2). Another study identified that a series
of synaptic adherence-like molecules (SALMs) promoted neurite
growth in certain brain regions in rats. Both SALMs and FLOT1 can
interact with NMDA receptors in glutamatergic synapses, and FLOT1
is found to be a molecular mediator of SALM4-induced neurite
branching. Moreover, FlOT1 alone can induce neurite formation and
branching, which is dependent on the presence of intact lipid
rafts. In conclusion, SALM4 can regulate the FLOT1-associated
pathway in hippocampal neurite branches (32). Another study showed that FLOT1
co-clustered with Prion protein (PrP) to transduce signals, causing
N-cadherin to aggregate into the PrPC-FLOT1 complex in
the growth cone, which triggered axonal growth (33). In addition, can induce filopodia
formation in mammalian cell lines, thus promoting hippocampal
neuronal differentiation and neurite outgrowth (2,33,34).
Other physiological effects
Gonadotropin-releasing hormone (GnRH) and a small
amount of glucocorticoid receptor (GR) colocalize with the lipid
raft protein FLOT1 at the PM, and colocalizes with FLOT1
independent of its ligands. GR and Gonadotropin-releasing hormone
receptor (GnRHR) crosstalk in lipid rafts that mediate
FLOT1-associated regulation of cell proliferation through the
activation of PKC and the upregulation of SGK-1 (35). FLOT1 is expressed during formation
of osteoclasts, where FLOT1-dominated rafts are converted to
CAV1-rich rafts (36). In
addition, FLOT1 plays a detectable role in the process of
CD8+ T cell-mediated host monitoring under physiological
conditions (37). FLOT1 can
maintain the membrane integrity of B and T lymphocytes, as well as
T-cell activation (38).
Additionally, FLOT1 co-localizes with the inclusion membrane
protein A (IncA) in the chlamydia pneumonia inclusion membranes and
promotes bacterial intracellular growth by directly interacting
with the chlamydia pathogen (39).
In addition, exosomes are small extracellular membrane vesicles
originating from late endosomes, that can mediate the intercellular
transfer of RNA and protein (40).
Exosomes are a subtype of EVs involved in breast cell-to-cell
communication and immune processes and capable of transferring
their materials to receptors (41). Meanwhile, exosome-mediated
intercellular communication is the basis of cell senescence.
Exercise, which promotes the release of exosomes, may be key to
promoting intercellular communication and facilitating the
adaptation of a system to exercise in aging or other diseases, such
as type 2 diabetes mellitus, cardiovascular disease, and
sarcopenia. As a significant marker of exosomes, FLOT1 may promote
exosome function (19). In
addition, FLOT1 is reported to participate in cell adhesion, and
elevated FLOT1 enhances cell spreading (38).
The role of FLOT1 in tumors
Studies have shown that FLOT1 is upregulated in
several types of cancer. For example, in the respiratory system,
FLOT1 is overexpressed in lung adenocarcinoma (LUAD) (1), small cell lung cancer (SCLC),
non-small cell lung cancer (NSCLC), and nasopharyngeal carcinoma.
As for the digestive system, FLOT1 expression is increased in
hepatocellular carcinoma (HCC), esophageal squamous cell carcinoma
(ESCC), nasopharyngeal carcinoma (NPC), squamous cell carcinoma of
the tongue, and colorectal cancer (CRC). In the urogenital system,
it is elevated in clear cell renal cell carcinoma (ccRCC), bladder
transitional cell carcinoma (BTCC), and PCa. In addition, FLOT1
expression is higher in gynecological cancers, such as CC, breast
cancer (BC), and human epithelial ovarian neoplasms (4,28,42–53).
However, FLOT1 is downregulated in neuroblastomas (28).
Upstream regulation of FLOT1 promotes
tumorigenesis
Emerging evidence has shown that the upstream
regulation of FLOT1 plays a pivotal role in maintaining the
homeostasis of normal physical processes; however, its expression
is dysregulated in several types of cancer, contributing to the
tumorigenesis (Table I).
lncRNAs and miRNAs in FLOT1-mediated
tumorigenesis
MicroRNAs (miRNAs/miRs) are non-coding RNAs
consisting of 17–24 nucleotides. miRNAs can interact with the
3′-untranslated regions (3′-UTRs) of target mRNAs, such as that of
the FLOT1 mRNA, forming a silencing target mRNA transcription
complex to inhibit and/or degrade the target mRNA (44), thus negatively regulating the
expression of genes post-transcriptionally (54). Studies have shown that long
intergenic non-coding RNAs (lncRNAs) and miRNAs are associated with
the occurrence and development of tumors by targeting FLOT1 mRNA
(44,54–56),
suggesting that FLOT1 is a potential therapeutic target for cancer
treatment (4,44,55).
Homeobox (HOX) transcript antisense RNA (HOTAIR) is a lncRNA
frequently reported to be involved in HCC tumorigenesis (57,58).
The upregulation of HOTAIR induces the increased expression of
FLOT1 by targeting miR-214-3p in hepatocytes. Therefore, the
HOTAIR/miR-214-3p/FLOT1 axis is involved in the proliferation,
invasion, and migration of HCC, and the downregulation of HOTAIR
produces the opposite result (42). In BC, A1BG-AS1 (a lncRNA) is
upregulated, resulting in the downregulation of miR-485-5p through
sponging. FLOT1 is the direct target of miR-485-5p; thus, elevated
A1BG-AS1 expression increases the expression of FLOT1, thus
promoting the tumorigenesis of BC (44). ccRCC is a common urinary tract
tumor in humans (56), with a high
rate of metastasis and poor survival (55). Taurine upregulated gene 1 (TUG1) is
a lncRNA that is significantly increased in the tissues and cells
of ccRCC to participate in tumor progression. TUG1 positively
modulated the expression of FLOT1 through sponging miR-31-5p. The
overexpression of FLOT1 attenuates the inhibition of cell
proliferation mediated by miR-31-5p and promotes apoptosis and
autophagy, promoting the progression of ccRCC (45). In addition, FAM201A (a lncRNA)
targets the Wnt/β-catenin pathway induced by the miR-1271-5p/FLOT1
axis. FAM201A is upregulated in CC, while miR-1271-5p is
downregulated. The overexpression of FAM201A increases FLOT1
expression and CC tumorigenesis, cell viability, migration, and
invasion in vivo, which may be reversed by the upregulation
of miR-1271-5p (59). In addition,
malignant glioma is the most common intracranial tumor in adults
and is often fatal. FLOT1 expression is upregulated in glioma
tissues and cells, where it serves as an oncogene. The upregulated
nuclear-cap-binding subunit 3 (NCBP3) in gliomas binds to SNHG6 (a
lncRNA) to stabilize the expression of SNHG6, inhibiting the
transcription of gastrulation brain homeobox 2 (GBX2) via histone
modification. GBX2 can reduce the promoter activity and
downregulate the expression of the FLOT1 oncogene; thus the
upregulation of NCBP3/SNHG6 and downregulation of GBX2 promotes the
expression of FLOT1, promoting the proliferation, migration,
invasion, and other malignant biological behaviors of glioma cells.
Conversely, the downregulation of NCBP3 and SNHG6 and the
upregulation of GBX2 can inhibit the malignant biological behaviors
of tumor cells; highlighting a novel avenue for the targeted
therapy of glioma (60).
A study showed that miR-6809-5p mediated HCC,
induced by luteolin (a natural flavonoid), via targeting of FLOT1.
miR-6809-5p is upregulated by luteolin, and miR-6809-5p directly
targets FLOT1 in hepatocytes to inhibit the growth of HCC cells.
However, knockdown of miR-6809-5p reversed the effect of luteolin
to restrain the development of HCC (61). In addition, FLOT1 is a direct
target of miR-124, and the ectopic expression of miR-124 can
significantly inhibit FLOT1, suppressing the growth and migration
of BC cells. Luciferase assays showed that miR-124 could directly
bind to the 3′-UTRs of FLOT1 and inhibit its translation. In BC,
the expression of miR-124 is downregulated, while FLOT1 is
extensively upregulated. miR-124 is also involved in tumor lymph
node metastasis (TNM) staging and the metastasis of lymph nodes
(62). In CC, the expression of
miR-1294 is decreased, and the overexpression of miR-1294 can block
EMT and inhibit the expression of β-catenin to inhibit the
viability and metastasis of CC cells. Notably, miR-1294 has been
shown to target FLOT1 directly, which inhibits cell viability,
migration, and invasion by inhibiting the expression of FLOT1.
Therefore, miR-1294 acts as a tumor inhibitor of CC by regulating
the expression of FLOT1 and blocking EMT (43). In ccRCC, FLOT1 is a potential
target gene of miR-506 and the target gene of miR-124, which are
associated with the genesis and development of ccRCC (55,56).
FLOT1 is negatively correlated with the expression of miR-506 and
is upregulated in ccRCC, and this upregulation of FLOT1 promotes
the growth and metastasis of ccRCC cells. miR-506 is an independent
prognostic marker of ccRCC patients, and its expression is
positively associated with advanced clinical stages (56). In addition, the miRNA target
network reveals that miR-124 is a key miRNA, and it leads to the
acquisition of aggressive behaviors in ccRCC by targeting CAV1 and
FLOT1. Patients with higher expression of FLOT1 and CAV1 exhibit
lower miR-124-3p levels and shorter overall survival. miR-124-3p,
miR-30a-5p, and miR-200c-3p are the most influential miRNAs in the
pathogenesis of ccRCC, and the recovery of these miRNAs reduces the
invasion, migration, and spread of ccRCC, which can be regarded as
a potential therapeutic strategy for ccRCC (55). ESCC is one of the most aggressive
tumors of the gastrointestinal tract. A related study showed that
the knockdown of miR-138 upregulated various lipid rafts
components, including FLOT1, FLOT2, and CAV1 to induce lipid raft
formation, thus promoting the invasion of ESCC via increased
expression of FLOT1. Increased miR-138 expression has the opposite
effect, which suggests that miR-138 plays a tumor-suppressive role
in ESCC by targeting FLOT1 (63).
Upregulation, sumoylation, and
palmitoylation of FLOT1 promote EMT
Epithelial-mesenchymal transition (EMT) is the
process by which epithelial cells acquire the characteristics of
mesenchymal cells (64). Numerous
studies have shown that EMT is involved in the occurrence,
invasion, and metastasis of tumors, as well as in the resistance to
therapy in several types of cancer (64,65).
Upregulation of FLOT1 promotes EMT by promoting TGF-β/Smad and AKT
signaling pathways (1,52). Sumoylation of FLOT1 can also
promote EMT and cancer metastasis by inhibiting Snail degradation,
which is a transcription factor regulating the expression of
EMT-related genes (5).
The EMT-related markers include N-cadherin, matrix
metalloproteinase (MMP)-2, MMP-9 (66), and E-cadherin (67). EMT is primarily dependent on the
TGF-β/Smad and AKT/mTOR pathways (68,69).
FLOT1 can regulate EMT to promote the proliferation and metastasis
of SCLC (50). In LUAD, FLOT1 can
downregulate the epithelial marker E-cadherin and upregulate the
mesenchymal markers β-catenin and MMP-2 to promote EMT. EMT
promotes the growth, migration, and invasion of cancer cells and
inhibits the apoptosis of cells. Therefore, targeting FLOT1 may be
a potential therapeutic strategy for the management of LUAD
(1). In addition, both the protein
and mRNA levels of MMP-2, MMP-9, N-cadherin, and the Wnt/β-catenin
signaling pathway are elevated following overexpression of FLOT1 in
CC cells (59).
Snail is a major transcription factor involved in
EMT, which mediates EMT gene expression and/or inhibits E-cadherin
expression (5,70). In metastatic PCa, upregulated E2
conjugating enzyme UBC9 sumolylates FLOT1 at Lys51 and Lys195
following SUMO2/3 modification. The sumoylation of FLOT1 promotes
EMT and cancer metastasis by interacting with Snail and inhibiting
the degradation of Snail through the proteasome pathway in a
sumoylation-dependent manner. Therefore, upregulation of UBC9 can
promote the above process, thus targeting UBC9 can be used to
regulate EMT in metastatic PCa, providing a novel therapeutic
direction (5). CRC is one of the
most common malignancies, the metastasis of which remains the
primary cause of death clinically amongst patients with CRC, and it
has been shown that FLOT1 can induce classical EMT, which is
mediated by the TGF-β/Smad pathway and increase the migratory
ability of CRC. FLOT1 is regulated by S100 calcium-binding protein
A11 (S100A11) as its downstream factors at the post-transcriptional
level instead of a transcriptional level, S100A11 can bind with LIM
and SH3 protein 1 (LASP1) to regulate EMT mediated by TGF-β/Smad
and the acquisition of a cell invasive phenotype (11). In CC, the upregulation and
palmitoylation of FLOT1 are positively correlated with the
induction of EMT genes, such as TIAM1 and GREM1, thus promoting the
progression and metastasis of CC (13).
Upregulation of FLOT1 promotes the
proliferation of cancer cells via regulation of the cell cycle
In LUAD, the overexpression of FLOT1 inhibits the
expression of cyclin-dependent kinase 2 (CDK2), Cyclin E, and
Cyclin D1 and elevates the expression of p16 to modulate the cell
cycle. In addition, FLOT1 regulates the cell cycle by activating
Erk/Akt signaling (1). A study
showed that the knockdown of FLOT1 increased the proportion of
cells in the G1 phase, suggesting that the suppression of FLOT1
could arrest SCLC cells at the G1 phase (50). In BC, the knockdown of FLOT1 could
upregulate the cyclin-dependent kinase inhibitors p21 and p27, and
reduce Cyclin D1 expression to inhibit the proliferation and
tumorigenicity of BC cells (9).
Another study showed that the knockdown of FLOT1 suppressed the
proliferation and induced G1-phase arrest in BCa cells, which is
related to AKT/forkhead box class O3a (FOXO3a) signaling (65).
FLOT1 affects signaling pathways
Gene expression profiling has shown that FLOT1
regulates the genes of AKT/FOXO3a, TGF-β-Smad2/3 (50), TNFR/NF-κB (63), and other signaling pathways. The
upregulation of FLOT1 promotes the AKT/FOXO3a signaling pathway to
promote the development of BC (9),
BCa (71), LUAD (1), and RCC (72), and promoted TGF-β-Smad2/3 signaling
to promote the development of SCLC (50) and NPC (52), and promoted TNFR/NF-κB signaling to
promote the development of ESCC (63) and HCC (61) (Fig.
3).
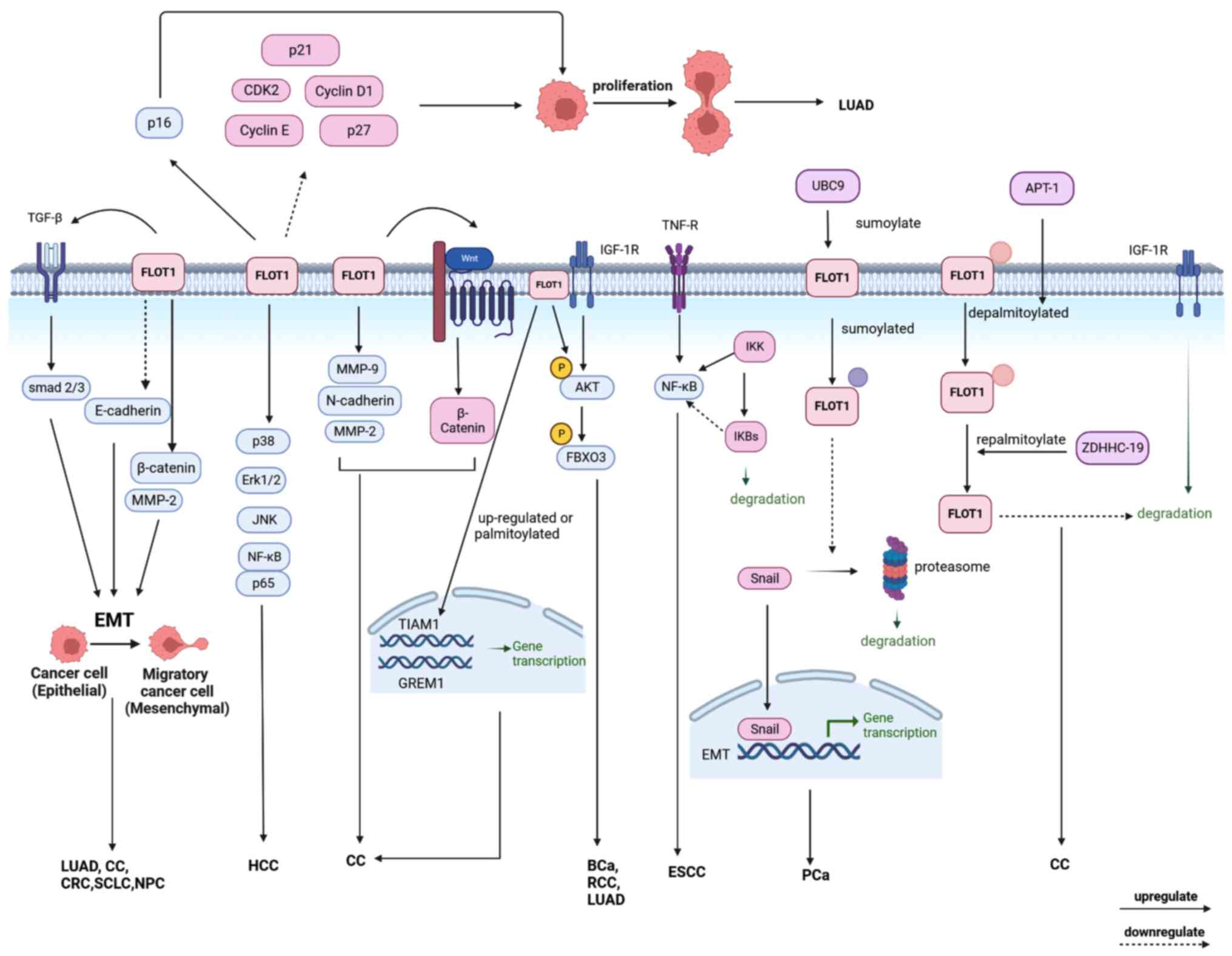 | Figure 3.The mechanisms of FLOT1 leading to
tumors: Upregulation, sumoylation, or palmitoylation of FLOT1
promotes EMT to promote the development of tumors. Upregulation of
FLOT1 promotes the proliferation of cancer cells by regulating the
cell cycle. In addition, FLOT1 is involved in the AKT/FOXO3a,
TGF-βsmad2/3, TNFR/NF-κB, Wnt, and IGF-1R signaling pathways to
mediate tumor proliferation, invasion, and metastasis in several
types of cancer. EMT, epithelial-mesenchymal transition. FLOT,
flotillin protein; FOXO3a, forkhead box class O3a; TGF-β,
transforming growth factor β; TNF, tumor necrosis factor; TNFR,
tumor necrosis factor-α receptor; IGF-1R, insulin-like growth
factor-1 receptor. |
Upregulation of FLOT1 promotes
AKT/FOXO3a signaling pathway
In BC, a study showed that the knockdown of FLOT1
was related to the inhibition of Akt activity and the enhanced
transcriptional activity of FOXO3a, which inhibits the
proliferation of BC cells (9). In
addition, miRNA-608 inhibits the proliferation and development of
BCa cells by significantly downregulating the levels of p-AKT and
p-FOXO3a to activate the AKT/FOXO3a signaling pathway, which is
opposed to FLOT1. However, the upregulation of FLOT1 can reverse
the inhibition of cell proliferation caused by miR-608 (71). In LUAD, the overexpression of FLOT1
upregulates the phosphorylation of Akt and downregulates the
expression of FOXO3a to induce EMT and modulate the cell cycle,
which promotes the growth, migration, and invasion of cancers cells
and inhibits cells apoptosis (1).
In RCC, the knockdown of FLOT1 decreased the phosphorylation of
both FOXO3a and AKT, resulting in the inhibition of AKT/FOXO3a
signaling (72).
Upregulation of FLOT1 promotes
TGF-βsmad2/3 signaling
FLOT1 is upregulated in SCLC, and its expression is
closely associated with the clinical stage, distant metastasis, and
a poor survival rate. FLOT1 promotes EMT in SCLC by increasing the
activities of TGF-β-smad2/3 and AKT signaling pathways. Therefore,
the knockdown of FLOT1 reduces the growth, migration, and invasion
of SCLC cells and reverses an EMT phenotype (50). NPC exhibits potent local invasion
and a high frequency of regional lymph node metastasis, and
patients with NPC often have a poor prognosis. A study showed that
upregulation of FLOT1 induced the expression of transforming growth
factor β1 (TGF-β1) and promoted EMT through activation of the
TGF-β/Smad3 signaling pathway, which accelerates the invasion and
metastasis of NPC. Therefore, FLOT1 is potentially important in the
prognosis of NPC (52).
Upregulation of FLOT1 promotes the
TNFR/NF-κB signaling pathway
The NF-κB pathway has been identified as a
carcinogenic signaling pathway that plays an important role in
inflammation and cancer (46). The
activation of the NF-κB signaling pathway plays a crucial role in
the occurrence and development of ESCC, and blocking the NF-κB
signaling pathway can inhibit the proliferation of ESCC. A study
showed that overexpression of FLOT1 could activate the NF-κB
signaling pathway and promote the invasion of ESCC, which is
inhibited by miR-138 (63). In
addition, FLOT1 promotes tumor necrosis factor-α receptor (TNFR)
signaling and NF-κB activation in ESCC. It was shown that FLOT1
promoted the recruitment of TNFR and IKK (NF-κB kinase)
signalosomes to lipid rafts and promoted K63-linked polyubiquitin
signaling. FLOT1 also promoted ubiquitin-coupled NF-κB signaling
and maintained NF-κB activation. The recruitment of TNF
receptor-associated factors (TRAFs) and receptor-interacting
proteins (RIPs) to the receptor are ubiquitinated by a K63-linked
polyubiquitin chain, facilitating the recruitment and activation of
inhibitors of TGF-β-activated kinase-1 (TAK1) and IKK complexes.
Activated IKK promotes the phosphorylation/proteasome degradation
of NF-κB suppressor proteins (IKBs), leading to the activation of
NF-κB (46). In HCC,
downregulation of FLOT1 inactivates Erk1/2, p38, JNK, and NF-κB/p65
signaling pathways, thus inhibiting the growth of HCC cells;
however, this effect can be reversed by upregulation of miR-6809-5p
(61).
Other FLOT1-regulated signaling
pathways in cancer
In CC, acyl protein thioesterases-1 (APT-1) promotes
the depalmitoylation of FLOT1, and zinc finger DHHC
domain-containing protein palmitoyltransferase-19 (ZDHHC-19)
repalmitoylated FLOT1, which is frequently depalmitoylated in CC
cells. The turnover of FLOT1 can prevent the desensitization of
IGF-1R through endocytosis and lysosomal degradation, thus
promoting the tumorigenesis of CC. Meanwhile, IGF-1 can promote
palmitoylation of FLOT1 following IGF-1R activation (13). FLOT1 is involved in the acquisition
of drug resistance in cancer. Multidrug resistance (MDR) of tumor
cells is the leading cause of failure of chemotherapy and other
anticancer drugs. A study showed that the knockdown of FLOT1
reduced drug resistance in CRC by downregulating the
phosphatidylinositol 3-kinase (PI3K)/AKT signaling pathway. After
disruption of lipid rafts, increased cell membrane permeability may
lead to increased drug accumulation in the cytoplasm, thus
reversing resistance. Therefore, FLOT1 can be used as a potential
therapeutic target in CC (73).
Other effects of FLOT1 in cancer
FLOT1 can induce the differentiation of certain cell
types. It has been reported that lovastatin (lova) has dual effects
on cancer cells. High levels of lova can induce apoptosis of
thyroid cancer ARO cells, while low concentrations can induce
differentiation of this cancer cell line; thus, lova may have
potential as an adjuvant for cancer treatment. FLOT1 levels were
increased in ARO cells following treatment with lova, and
overexpression of FLOT1 increased the expression of thyroid
differentiation markers, such as TG, TPO, TSHR, and SIS, suggesting
that FLOT1 transformed ARO cells from an undifferentiated state to
a differentiated state. These results suggest that FLOT may mediate
lova-induced differentiation at least to a certain extent (74). In addition, FLOT1 plays an
important role in cell proliferation, and its overexpression is
associated with adverse outcomes in BC patients with LUAD (75). It was found that vacuolar protein
sorting protein 33b (VPS33B) modulated exosome maturation and the
secretion of proteins, and the lack of VPS33B may lead to a delay
in leukemogenesis. Therefore, the study of FLOT1 and other exosome
markers is conducive to the development of improved cancer
treatment strategies (76).
In addition, FLOT1 is an independent prognostic
indicator of several types of cancer. For example, a study showed
that laryngeal cancer is a common type of cancer in men.
Researchers established gene models and found that ACE2, FLOT1, and
especially PRKD1 may have prognostic and biological significance.
Therefore, these genes can be used as independent prognostic
markers for postoperative recurrence of laryngeal squamous cell
carcinoma (77). The prognosis of
CRC patients after immunotherapy remains mixed; six immune-related
gene markers (CCL22, LIMK1, MAPKAPK3, FLOT1, GPRC5B, and IL20RB)
were found to be reliable prognostic indicators in CRC patients,
providing insights into personalized cancer therapy and improving
prognostic prediction in CRC patients (78).
The expression of FLOT1 in human ccRCC is involved
in the progression of ccRCC and is associated with poor survival.
Upregulated FLOT1 in ccRCC is involved in the tumorigenesis and
progression of ccRCC. Therefore, FLOT1 can be used as a therapeutic
target and an independent prognostic marker in patients with ccRCC
(79). The prevalence of cutaneous
squamous cell carcinoma (cSCC) was higher in patients who had
undergone immunosuppressive organ transplantation than in the
general population, and 16 T cell methylation domains (DMR) were
found to be different between patients with and without cSCC
following transplantation. An example of a gene annotated to DMR is
FLOT1, which encodes a protein related to the migration of T-cells
(80).
FLOT1-meidated inhibition of
tumorigenesis
Neuroblastoma is one of the most common solid tumors
in children, accounting for ~15% of childhood cancer-related deaths
(81). FLOT1 is downregulated in
neuroblastoma, and FLOT1 expression is inversely correlated with
clinical malignancy. Decreased expression of FLOT1 in neuroblasts
leads to dissociation of ALK from endosomes and membrane
accumulation of ALK, promoting the expression or phosphorylation of
ALK and the phosphorylation of the downstream mediators of ALK,
such as ERK1/2, AKT and STAT3 signaling, which enhances the
malignant features of neuroblastoma cells. Therefore, weakened
FLOT1-ALK binding activates ALK signaling, promoting the malignant
phenotype of neuroblastoma (28).
The role of FLOT1 in neurological
diseases
Raft disruption is an important cause of several
degenerative diseases including Alzheimer's disease (AD), PD, and
prion disease. FLOT1 may contribute to the total degenerative
process. Understanding the pathogenesis of lipid raft-related
structures in neurological diseases may be helpful for the
treatment of neurodegenerative diseases (8) (Fig.
4).
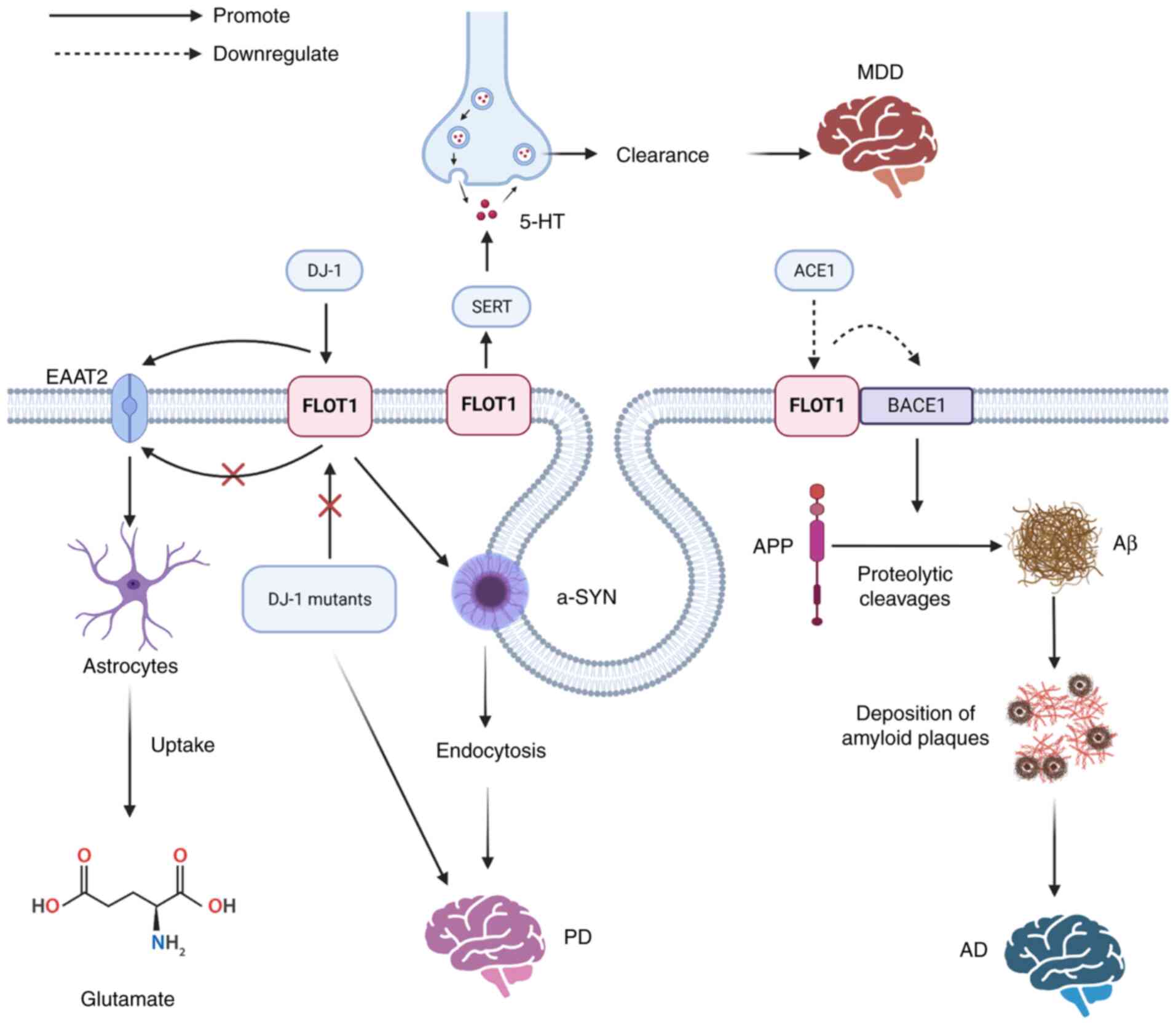 | Figure 4.The mechanisms of FLOT1 leading to
neurological diseases: Aβ is produced by proteolytic cleavages of
APP under the influence of BACE1, the accumulation of which can
lead to the development of AD, and the overexpression of FLOT1
suppresses the activity of BACE1. DJ-1 can regulate the protein
stability of FLOT1, but the PD-associated DJ-1 mutants fail to
regulate the FLOT1. DJ-1 promotes the expression of EAAT2 by
upregulating FLOT1, promoting the uptake of glutamate by
astrocytes. FLOT1 contributes to the microdomain localization and
regulation of SERT, and SERT can control the reuptake of released
5-HT from the synaptic cleft into the presynaptic terminal to
regulate 5-HT clearance, leading to the development of MDD. Aβ,
amyloid-β peptide; APP, amyloid precursor protein; BACE1, β-site
APP cleaving enzyme 1; AD, Alzheimer's disease; PD, Parkinson's
disease; FLOT, flotillin protein; DJ-1, PD protein 7; EAAT2, glial
glutamate transporter; SERT, presynaptic high-affinity 5-HT
transporter; MDD, major depressive disorder; EAAT2, glial glutamate
transporter; ACE1, angiotensin-converting enzyme 1; -SYN,
α-synuclein. |
FLOT1 in AD
AD is the most common neurodegenerative disorder,
accounting for ~80% of all dementia cases (82). The presence of amyloid plaques and
the deposition of phosphorylated Tau in the brain are the
neuropathological hallmarks of AD (82,83).
Plaques are primarily composed of amyloid-β peptide (Aβ), which is
produced by proteolytic cleavage of amyloid precursor protein (APP)
under the influence of β-and γ-secretase. β-site APP cleaving
enzyme 1 (BACE1) has been identified as a β-secretase. A study
showed that BACE1 can interact with FLOT1, and part of BACE1 is
recruited to lipid rafts in FLOT1-overexpressing cells. However,
overexpression of FLOT1 suppresses β-secretase activity. It was
speculated that the activity of β-secretase was inhibited as the
binding of BACE1 to FLOT1 may conceal its active site (83). In addition, the rennin-angiotensin
system (RAS) can enhance the expression of BACE1, which increases
the accumulation of Aβ, which in turn stimulates the development of
AD. Cleavage of APP into Aβ1-42 by BACE1 occurs in the lipid raft,
and the lipid raft colocalizes with several receptors and enzymes
involved in AD pathogenesis, such as estrogen receptor (Erα) and
BACE1. However, the brain-penetrating angiotensin-converting enzyme
1 (ACE1) inhibitor perindopril can inhibit the expression of Aβ1-42
and decrease the expression of FLOT1. A study showed that the
levels of FLOT1 in a hyperlipidemia AD model were significantly
increased. FLOT1 was closely related to AD, but the molecular
mechanism remains to be elucidated (82). In addition, a related study found
differences in the expression of ELA protein levels in
cerebrospinal fluid between AD and healthy controls, especially in
proteins involved in endocytosis (84).
FLOT1 in PD
PD is the second most common neurodegenerative
disorder (85), second to AD.
FLOT1 expression is significantly elevated in brains with PD at the
transcriptional and translational levels. The presence of
α-SYN-positive Lewy bodies (LBs) and loss of catecholaminergic
neurons are neuropathological features of PD; FLOT1 and DAT are the
components of α-LB. A study showed that extracellular α-SYN
promotes the binding of FLOT1-DAT and their accumulation at the
cell surface prior to endocytosis, facilitating the endocytosis of
DAT into dopaminergic neuron-like cells. Meanwhile, FLOT1 can
trigger the endocytosis of α-SYN (30). In addition, a study found that DJ-1
can regulate the stability of the FLOT1 protein; however,
PD-associated DJ-1 mutants fail to regulate FLOT1. DJ-1 promotes
the expression of EAAT2 by upregulating FLOT1, promoting the uptake
of glutamate by astrocytes. The overexpression of FLOT1 rescued the
decreased glutamate uptake and decreased expression of EAAT2 caused
by DJ-1 deficiency. These results suggest that the abnormal low
expression of FLOT1 may lead to neurodegeneration (16).
FLOT1 in cerebrovascular diseases
Cerebral small vessel diseases (CSVD) are the
primary leading cause of dementia and vascular cognitive
impairment. Hypertension (HTN) is common in the elderly population
and can lead to cerebral hemorrhage and other injuries. In the
spontaneous hypertensive stroke predisposition (SHR-SP) model of
HTN, the researchers found that FLOT1 is a key protein (86). In addition, FLOT1 is increased in
the spontaneously hypertensive rat (SHR) model. Therefore,
understanding the function of FLOT1 provides novel mechanistic
insights into the development of these different forms of CSVD
(87). Rupture of intracranial
aneurysms is the primary cause of subarachnoid hemorrhage (SAH), it
is important to study the specific gene expression profiles
associated with intracranial aneurysms. It was found that FLOT1 may
be a potential biomarker of SAH, which is more conducive to the
differential diagnosis of aneurysmal SAH so as to avoid
misdiagnosis and miss the optimal treatment opportunity (88).
FLOT1 in Major Depressive Disorder
(MDD)
Major depressive disorder (MDD) is the most common
psychiatric disorder (82,89). A study showed FLOT1 was
significantly upregulated in the brain tissues of MDD patients.
Several studies have identified FLOT1 as a novel MDD risk gene and
MDD-associated genetic variants may confer a risk of MDD by
affecting the expression of FLOT1 (90,91).
Serotonin (5-hydroxytryptamine, 5-HT) is a neurotransmitter
(92), the disruption of which is
associated with a variety of brain disorders, such as MDD. The
presynaptic high-affinity 5-HT transporter (SERT) can modulate the
reuptake of released 5-HT from the synaptic cleft into the
presynaptic terminal to regulate 5-HT clearance (92). SERT activity may be regulated by
SERT-interacting proteins (SIPs), FLOT1 is a SIP, which is
hypothesized to contribute to SERT microdomain localization and
regulation. Therefore, FLOT1 may promote the clearance of 5-HT.
Studying FLOT1 may help identify novel drug targets for the
treatment of 5-HT-related diseases such as depression (89). In chronic corticosterone response
(CORT), aberrant neurotransmission of 5-HT in the brain is
hypothesized to be the central mechanism in neuropsychiatric
disorders. As a SIP, FLOT1 is involved in the response to CORT
therapy, and gene deletion of FLOT1 promotes chronic CORT-induced
behavioral despair (92).
Other effects on the nervous
system
NMDAR dysfunction can lead to several neurological
disorders such as stroke or excitotoxic conditions. One potential
role of FLOT1 may be to recruit NMDARs to lipid rafts to initiate a
second messenger signaling system. The depletion of lipid rafts
protects neurons from NMDAR-induced excitotoxicity (22). Prions are a type of infectious
agent, which can cause a series of fatal neurodegenerative
diseases, also known as TSEs. The primary cause of prion-related
diseases is the conversion of the PrPC conformation
encoded by the normal host prion gene PRNP to the abnormal
conformation, PrPSc. However, the conversion of
PrPC to PrPSc may occur in lipid rafts or via
their associated intracellular processes. Moreover, in a
prion-infected cell, PrPSc has been reported to highly
colocalize with FLOT1 in the FLOT1-positive vesicles. However, the
specific mechanism by which FLOT1 affects the conversion of
PrPC into PrPSc has not been elaborated
(6).
The role of FLOT1 in DCM/idiopathic
(I)DCM
DCM is the primary cause of heart failure (HF),
which is attributed to systolic dysfunction and ventricular
dilatation. Currently, DCM is primarily treated by immunotherapy;
however, there are significant individual differences, and the
therapeutic effect requires further improvement. FLOT1 is
overexpressed in DCM with HF, and FLOT1 may affect the development
of DCM by activating T cells and accelerating cell dispersal,
highlighting potential therapeutic targets of DCM (38). IDCM can affect the vascularization
of myocardial tissues. Stromal cell-derived factor
(SDF-1α)-mediated migration may affect endothelial recovery in
patients. Significant colocalization of SDF-1α and FLOT1-specific
markers have been observed in IDCM, and SDF-1α is also highly
expressed in IDCM lipid rafts. A study provided novel insights into
the function of lipid rafts in IDCM and hypothesized more effective
treatments, although the mechanisms by which FLOT1 interacted with
SDF-1α were not elucidated (93).
The role of FLOT1 in pathogenic microbial
infections
A study showed that the FLOT1 gene was
differentially expressed following parvovirus B19 infection and it
was associated with integrin signaling, cytoskeleton, and tumor
inhibition (94). Infection with
Anaplasma phagocytophilum pathogens, also known as human
granulocyte anaplasmosis (HGA) requires phosphatide protein
recruitment of LDL cholesterol, and FLOT1 and FLOT2 as membrane
proteins in heavy phagocytes bacillus infection and cholesterol
played vital roles. FLOTs may contribute to anaplasma replication
in host cells by aiding the blister transport of LDL-derived free
cholesterol to anaplasma inclusions (3). A study showed that atherosclerosis
was associated with chlamydia pneumoniae, an intracellular pathogen
of the respiratory tract that can infect bronchoalveolar
macrophages and can be transported to sites of vascular injury.
Gene expression profiling of U937 human macrophages exposed to
chlamydia pneumoniae and/or LDL revealed several interesting
transcripts involved in structural integrity with respect to
atherosclerosis, including FLOT1. The transcriptional alteration of
FLOT1 was involved in atherosclerosis caused by chlamydia
pneumoniae infection (95).
Crohn's disease (CD) is a chronic and progressive disorder, and the
etiology of CD may result from an abnormal interaction between
microbiota and the enteric immune system in patients. Among them,
invasive Escherichia coli (AIEC) is of great significance. Anti-TNF
agents can limit AIEC survival within macrophages, and anti-TNF
agents can induce the increase of FLOT1 and decrease mRNA levels of
CHI3L1, which can promote the clearance of AIEC. Moreover, the
levels of FLOT1 are negatively correlated with AIEC survival in CD
patients treated with anti-TNF agents (96). FLOT1 in lipid rafts is an important
part of the phagocytic lysosomal membrane of macrophages, thus
FLOT1 plays an important role in anti-fungal immunity (97).
The role of FLOT1 in diabetes-related
diseases
Diabetes mellitus (DM) is characterized by
hyperinsulinemia and hyperglycemia (98,99).
ANGPTL8, which has a unique characteristic in regulating lipid and
glucose metabolism, is upregulated in diabetes and is becoming
increasingly recognized as a potential drug target for the
treatment of diabetes and related metabolic disorders (100,101). A study showed that the FLOT1 gene
was co-expressed with ANGPTL8. It may be involved in the
pathogenesis of diabetes and insulin resistance (102). Low FLOT1 expression in the livers
of patients with Type 2 diabetes mellitus (T2DM) may lead to
metabolic lipoproteinemia, which is due to impaired liver
processing of C-TRLs. Syndecan-1 is a receptor for C-TRLs that
mediates endocytosis through rafts. The interaction of C-TRLs and
syndecan-1 enhances the association of syndecan-1/FLOT1 on liver
cells. FLOT1 mRNA and protein levels are reduced in a rat model of
T2DM. Knockdown of FLOT1 in cultured liver cells substantially
inhibited endocytosis of syndecan-1, resulting in the accumulation
of C-TRLs. FLOT1 is a relatively newer player in the treatment of
harmful C-TRLs via syndecan-1 (29). Glucose and amino acid metabolism
are altered during exercise and rehabilitation in patients with
T2DM. The therapeutic benefits of physical activity for the
prevention and treatment of T2DM are generally accepted. Pathway
analysis of differentially regulated genes during exercise showed
that FLOT1 and other genes were elevated in T2DM patients who
exercised, highlighting novel insights into the underlying
mechanisms that ameliorate the disturbances in glucose and amino
acid metabolism associated with T2DM (103).
The role of FLOT1 in other diseases
Fabry disease (FD) is a rare and serious disorder
caused by α-galactosidase a (GLA) enzyme deficiencies with often
painful clinical features. Pain-related ion channels are related to
the pain-like properties of FD, and their expression may also be
influenced by the recruitment of lipid raft components such as
several FLOT1-mediated receptors and channels from the nucleus to
the cell membrane (104). A study
showed that 15 miRNAs and 4 lncRNAs had potential functions as
diagnostic markers of pediatric sepsis. FLOT1 has been shown to
play a critical role in sepsis at the mRNA level (105). Moreover, the FLOT1 gene may be
involved in the formation of rheumatoid arthritis (RA) (106), and a study identified novel genes
in RA, in which FLOT1 expression differed significantly between RA
patients and healthy controls (107). End-stage renal disease (ESRD) is
the final stage of chronic kidney disease, and nocturnal
hemodialysis (NHD) is a more favorable treatment approach in
patients with ESRD. It is important to understand the expression of
genes related to immune function in NHD in ESRD patients and to
improve the immune response. Notably, it was found that the FLOT1
gene may be a potential target regulated by core transcription
factors, which is related to the immunoreaction in NHD in ESRD
patients (108).
FLOT1 is also related to certain gynecological
diseases. In placental transcytosis, the abundant co-expression of
FLOT1 and FLOT2 in cytotrophoblasts (CTs) and endothelial cells of
full-term villous placenta, and the flotillin-dependent endocytosis
may be important in the CT and endothelium. FLOT1 has potential
implications for placental transcytosis (26). Abnormal adhesion of embryos to the
endometrium leads to embryo implantation failure and infertility.
Therefore, proteins involved in regulating adhesion in endometrial
epithelial cells (HEECs) may be potential biomarkers or targets for
infertility treatment. IL-11 is involved in HEEC adhesion, and its
expression is dysregulated in infertile women. IL-11 can increase
the expression of FLOT1 in HEEC membranes to regulate endometrial
epithelial cell adhesion. Therefore, FLOT1 may serve as a marker of
infertility or a pharmacological target for regulating fertility
(14).
The bioinformatic analysis of FLOT1
Following a pan-cancer analysis of FLOT1 in TIMER
2.0 (http://timer.cistrome.org/)at the mRNA
level, the results indicated that the expression of FLOT1 in breast
invasive carcinoma (BRCA), cholangiocarcinoma (CHOL), esophageal
carcinoma (ESCA), head and neck squamous cell carcinoma (HNSC),
kidney chromophobe (KICH), kidney renal clear cell carcinoma
(KIRC), liver hepatocellular carcinoma (LIHC), LUAD,
pheochromocytoma and paraganglioma (PCPG), stomach cancer (STAD),
and thyroid cancer (THCA) differed from that in normal tissues
(Fig. 5A), among which, the
differences observed in LUAD and LIHC were significant. Therefore,
Gene Ontology (GO; http://geneontology.org/) enrichment analysis and
Kyoto Encyclopedia of Genes and Genomes (KEGG; http://www.genome.jp/kegg/) pathway enrichment
analysis in LIHC (Fig. 5B) and
LUAD (Fig. 5C) was performed.
First, RNAseq data were obtained from The Cancer Genome Atlas-LIHC
(HCC) project STAR process (https://portal.gdc.cancer.gov) in TPM format
alongside the clinical data. The FLOT1 data was extracted from the
data, and the DESeq2 package in R version 4.2.1 (http://www.R-project.org) (109) was used for differential analysis
on the original Counts matrix of the selected public data according
to a standard procedure (110).
FLOT1 expression was stratified based on the median expression into
low and high-expression groups. The high and low-expression groups
consisted of 187 patients each, respectively. Following
differential analysis, a total of 272 molecules were screened out
based on a |log2FC|≥2 criterion, after which ID conversion was
performed on the input molecule list, and enrichment analysis was
performed using the clusterProfiler (111) package in R. A similar approach is
used for LUAD. The results are shown as bubble diagrams (Fig. 5B and C). In LIHC, Gene
Ontology-Biological Process (GO-BP) analyses showed that it was
richer in ‘antimicrobial humoral immune response mediated by
antimicrobial peptide’, ‘glandular epithelial cell
differentiation’, ‘lung epithelium development’, ‘lung cell
differentiation’, and ‘lung epithelial cell differentiation’. KEGG
analyses showed that FLOT1 was associated with ‘neuroactive
ligand-receptor interaction’, ‘drug metabolism-other enzymes’,
‘metabolism of xenobiotics by cytochrome P450’ and ‘drug
metabolism-cytochrome P450’. In LUAD, GO-BP analyses showed that it
was richer in ‘mRNA trans-splicing, SL addition’, and ‘mRNA
trans-splicing via spliceosome’ and ‘formation of quadruple
SL/U4/US/U6 snRNP’. Gene Ontology-Cellular Component (GO-CC)
analyses showed that it was richer in ‘small nuclear
ribonucleoprotein complex’, ‘spliceosomal snRNP complex’, and
‘U4/U6 × U5 tri-snRNP complex’. Gene Ontology-Molecular Function
(GO-MF) analyses showed that it was closely related to ‘heparin
binding’, ‘snRNA binding’, and ‘U6 snRNA binding’. KEGG analyses
showed that it was enriched in the ‘spliceosome’, ‘alcoholism’, and
‘systemic lupus erythematosus’.
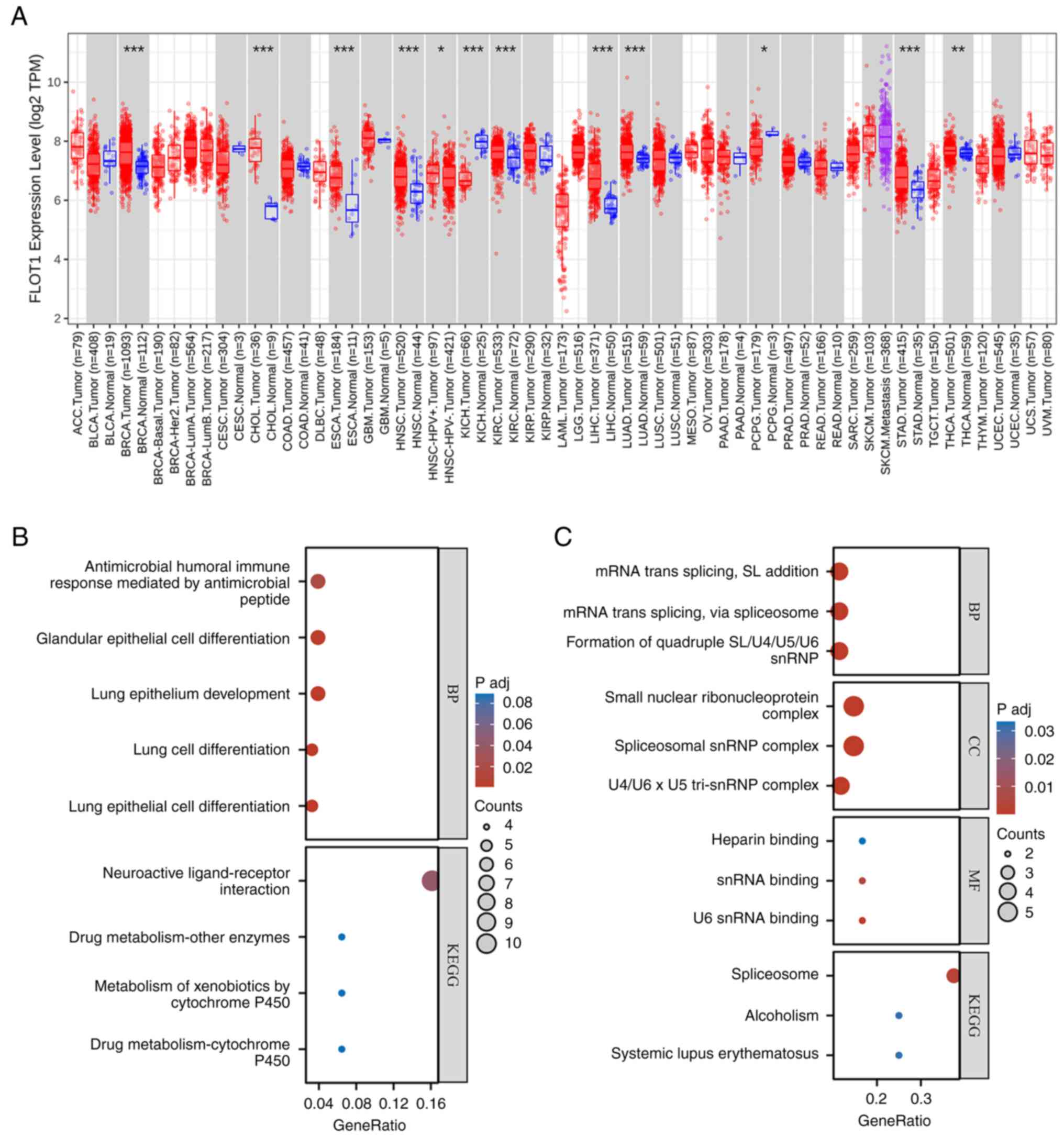 | Figure 5.Bioinformatic analysis of FLOT1: (A)
Pan-cancer analyses of FLOT1 in TIMER 2.0 showed that at the mRNA
level, the expression of FLOT1 in BRCA, CHOL, ESCA, HNSC, KICH,
KIRC, LIHC, LUAD, PCPG, STAD, and THCA differed from that in normal
tissues. *P<0.05, **P<0.01 and ***P<0.001. (B) In LIHC,
GO-BP analyses showed that it was enriched in antimicrobial humoral
immune response mediated by antimicrobial peptide, glandular
epithelial cell differentiation, lung epithelium development, lung
cell differentiation, and lung epithelial cell differentiation.
KEGG analyses showed that it was enriched in neuroactive
ligand-receptor interaction, drug metabolism - other enzymes,
metabolism of xenobiotics by cytochrome P450 and drug metabolism -
cytochrome P450. (C) In LUAD, GO-BP analyses showed that it was
enriched in mRNA trans-splicing, SL addition, mRNA trans-splicing,
via spliceosome, and formation of quadruple SL/U4/US/U6 snRNP.
GO-CC analyses showed that it was enriched in small nuclear
ribonucleoprotein complex, spliceosomal snRNP complex, and U4/U6 ×
U5 tri-snRNP complex. GO-MF analyses showed that it was closely
associated with heparin binding, snRNA binding, and U6 snRNA
binding. KEGG analyses showed that it was enriched in the
spliceosome, alcoholism, and systemic lupus erythematosus. BRCA,
breast invasive carcinoma; CHOL, cholangiocarcinoma; ESCA,
esophageal carcinoma; HNSC, head and neck squamous cell carcinoma;
KICH, kidney chromophobe; KIRC, kidney renal clear cell carcinoma;
LIHC, liver hepatocellular carcinoma; LUAD, lung adenocarcinoma;
PCPG, pheochromocytoma and paraganglioma; STAD, stomach cancer;
THCA, thyroid cancer; GO-BP, Gene Ontology Biological Process;
KEGG, Kyoto Encyclopedia of Genes and Genomes. |
Conclusions and future perspectives
The physiological effect and the role of FLOT1 in
human diseases have received significant attention. FLOT1 can
promote the internalization of DAT, α-SYN, and EAAT2. Abnormal
regulation of these can lead to the occurrence of nervous system
diseases, such as PD (16,30). In addition, FLOT1 can promote the
formation of hippocampal synapses, increase the number of
glutamatergic synapses, and trigger axonal growth through the
recruitment of N-cadherin (2,25,33).
Finally, FLOT1 plays a role in T-cell activation (38), regulating cell proliferation
(35), and participating in cell
adhesion (38), amongst other
processes. FLOT1 is also involved in several pathological
processes. The upregulation of FLOT1 promotes EMT to promote the
development of LUAD, PCa, and CC (1,5,59),
modulates the cell cycle in LUAD (1), and promotes the AKT/FOXO3a,
TGF-βsmad2/3, and TNFR/NF-κB signaling pathways to promote the
development of cancer (9,46,50),
while it plays an inhibitory role in neuroblastoma recurrence
(28). The reasons for this
difference may be due to the tumor microenvironment or tumor
heterogeneity.
However, the roles of FLOT1 in several tumors are
relatively limited. For example, a study showed that FLOT1 may be
an independent prognostic marker for laryngeal cancer patients
(77); however, the mechanism of
FLOT1 in laryngeal cancer requires further study. Additionally,
FLOT1 plays a role in T-cell activation, although the specific
process by which FLOT1 regulates T cells and whether it is related
to immunity have not been reported in detail, nor has it been
studied whether FLOT1 regulates tumor development by participating
in immune escape. Therefore, the relationship between FLOT1 and
immune responses requires further study.
FLOT1 can be upregulated by lncRNAs and
downregulated by miRNAs, thus promoting the occurrence of certain
types of tumors. Studying the mechanism of lncRNA/miRNA/FLOT1 axes
in the tumor is beneficial to developing tumor treatments. In
tumors in which FLOT1 acts as a tumor promoter, such as HCC and BC,
the pathogenic pathway of FLOT1 may be blocked by targeting certain
lncRNAs or upregulating associated miRNAs. For example, targeted
inhibition of HOTAIR in HCC, A1BG-AS1 in BC, TUG1 in ccRCC, and
FAM201A in CC may be considered, while in ccRCC, upregulation of
miR-506 and miR-124-3p may be considered. However, in tumors in
which FLOT1 acts as a tumor suppressor, such as neuroblastoma,
there are no reports of the IncRNAs and miRNAs associated with
FLOT1 in neuroblastoma. Therefore, additional studies into the
mechanism of FLOT1 in neuroblastoma are required. Notably, although
certain lncRNAs and miRNAs have been reported in tumors, there
remain several corresponding downstream miRNAs and upstream lncRNAs
that have not been mentioned. For example, SNHG6 a lncRNA can
promote the development of malignant glioma by upregulating FLOT1;
however, the downstream miRNA associated with SNHG6 has not been
studied. In addition, the upstream lncRNAs of miR-6809-5p, miR-124,
miR-1294, miR-138, miR-506, and miR-124-3p have not been studied as
of yet. lncRNAs/miRNAs/FLOT1 axes in tumors require further study.
Moreover, it was shown that FLOT1 played an important role in the
prognosis of tumors and is a potential prognostic target for
tumors, such as CRC (78), LUAD
(112), and ccRCC (79), although it is mostly confined to
mechanistic research, with clinical studies remaining limited, and
therapeutic targeting of FLOT1 for tumor management requires
further development and investigation.
FLOT1 also promotes the occurrence and development
of other diseases, such as PD (30), AD (82), and MDD (89). However, whether FLOT1 can be used
as a clinical drug target remains unknown, and further clinical
trials are required to verify its value in targeted therapy.
Additionally, although FLOT1 is associated with other diseases
apart from cancer, such as cerebrovascular diseases, RA, TSEs
(6,86,106), and DCM, its pathogenesis is not
clear (38). As FLOT1 is closely
associated with endocytosis, whether a disease involving
dysregulated FLOT1 function is caused by aberrant endocytosis
should be considered.
In general, the existing research provides support
for the physiological and pathological effects of FLOT1,
participating in the development of human diseases. As an important
regulatory factor in human diseases such as cancer, FLOT1 provides
a novel avenue for targeted therapy of diseases.
Acknowledgements
Not applicable.
Funding
This review was funded by The National Natural Science
Foundation of China (grant no. 32270821), The Natural Science
Foundation of Ningbo (grant no. 2021J065), The Fundamental Research
Funds for the Provincial Universities of Zhejiang (grant no.
SJLZ2022004), and The K.C. Wong Magna Fund from Ningbo University
(Ningbo, China).
Availability of data and materials
Not applicable.
Authors' contributions
ZZ conceived the subject of review, performed the
investigation, and wrote and edited the original draft. XJ and MY
wrote, reviewed, and edited the manuscript. All authors have read
and approved the final manuscript. Data authentication is not
applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ALK
|
anaplastic lymphoma kinase
|
|
APT-1
|
acyl protein thioesterases-1
|
|
AD
|
Alzheimer's disease
|
|
Aβ
|
amyloid-β peptide
|
|
APP
|
amyloid precursor protein
|
|
ACE1
|
angiotensin converting enzyme 1
|
|
AIEC
|
invasive Escherichia coli
|
|
ANGPTL8
|
angiopoietin-like protein 8
|
|
BTCC
|
bladder transitional cell
carcinoma
|
|
BC
|
breast cancer
|
|
BACE1
|
β-site APP cleaving enzyme 1
|
|
B19
|
parvovirus B19
|
|
CIE
|
clathrin-independent endocytosis
|
|
CC
|
cervical cancer
|
|
CRISPR
|
clustered regularly interspaced short
palindromic repeats
|
|
CRISPR/Cas9
|
clustered regularly interspaced short
palindromic repeats-associated sequence 9
|
|
CME
|
clathrin-mediated endocytosis
|
|
C-TRLs
|
residual apolipoprotein B rich in
cholesterol and triglycerides
|
|
CRC
|
colorectal cancer
|
|
ccRCC
|
clear cell renal cell carcinoma
|
|
CDK2
|
cyclin-dependent kinase 2
|
|
cSCC
|
cutaneous squamous cell carcinoma
|
|
CSVD
|
cerebral small vessel disease
|
|
CORT
|
chronic corticosterone response
|
|
CD
|
Crohn's disease
|
|
CT
|
cytotrophoblast
|
|
DCM
|
dilated cardiomyopathy
|
|
DAT
|
dopamine transporter
|
|
Dsg2
|
Desmoglein 2
|
|
Dsg2cacs
|
palmitoylated Dsg2
|
|
DA
|
transport of dopamine
|
|
DJ-1
|
Parkinson' disease protein 7
|
|
DMR
|
differential methylation regions
|
|
DM
|
Diabetes mellitus
|
|
DRG
|
dorsal root ganglion
|
|
EAAT2
|
glial glutamate transporter
|
|
EV
|
extracellular vesicle
|
|
EMT
|
epithelial-mesenchymal transition
|
|
ER
|
endoplasmic reticulum
|
|
EGF
|
epidermal growth factor
|
|
ESCC
|
esophageal squamous cell
carcinoma
|
|
Erα
|
estrogen receptor α
|
|
ESRD
|
end-stage renal disease
|
|
FLOT
|
Flotillin protein
|
|
FLOT1
|
FLOT-1/Reggie-2
|
|
FLOT2
|
FLOT-2/Reggie-1
|
|
FOXO3a
|
forkhead box class O3a
|
|
FD
|
Fabry's disease
|
|
GPI
|
glycosylphosphatidylinositol
|
|
GPCR
|
G-protein-coupled receptor
|
|
GnRH
|
gonadotropin-releasing hormone
|
|
GnRHR
|
GnRH receptor
|
|
GR
|
glucocorticoid receptor
|
|
GBX2
|
gastrulation brain homeobox 2
|
|
GLA
|
α-galactosidase A
|
|
Gb3
|
globulinyl ceramide
|
|
HCC
|
hepatocellular carcinoma
|
|
HOX
|
homeobox
|
|
HOTAIR
|
HOX transcript antisense RNA
|
|
HSC
|
hematopoietic stem cells
|
|
HTN
|
hypertension
|
|
HF
|
heart failure
|
|
HGA
|
human granulocyte form disease
|
|
HEECs
|
human endometrial epithelial
cells
|
|
IGF-1
|
insulin-like growth factor-1
|
|
IGF-1R
|
insulin-like growth factor-1
receptor
|
|
IncA
|
inclusion membrane protein A
|
|
IKK
|
NF-κB kinase
|
|
IKBs
|
NF-κB inhibitory protein
|
|
IDCM
|
idiopathic dilated cardiomyopathy
|
|
IL-11
|
Interleukin-11
|
|
lncRNAs
|
long intergenic non-coding RNAs
|
|
LUAD
|
lung adenocarcinoma
|
|
LASP1
|
LIM and SH3 protein 1
|
|
lova
|
lovastatin
|
|
LIC
|
leukemia initiating cell
|
|
LBs
|
α-SYN-positive Lewy bodies
|
|
M3R
|
muscarinic type 3 receptor, miRNA/miR
microRNA
|
|
mEPSC
|
miniature excitatory postsynaptic
current
|
|
mIPSC
|
miniature inhibitory postsynaptic
current
|
|
MMP-2
|
matrix metalloproteinase 2
|
|
MMP-9
|
metalloproteinase 9
|
|
MDR
|
multidrug resistance
|
|
MDD
|
major depressive disorder
|
|
NMDAR
|
N-methyl-d-aspartate receptor
|
|
NSCLC
|
non-small cell lung cancer
|
|
NPC
|
nasopharyngeal carcinoma
|
|
NCBP3
|
nuclear cap-binding subunit 3
|
|
NAFLD
|
non-alcoholic fatty liver disease
|
|
NHD
|
nocturnal hemodialysis
|
|
PHB
|
prohibition homology
|
|
PM
|
plasma membrane
|
|
PD
|
Parkinson's disease
|
|
PTM
|
post-translational modification
|
|
PKC
|
protein kinase C
|
|
PCa
|
prostate cancer
|
|
PrP
|
Prion protein
|
|
PBMC
|
peripheral blood mononuclear cell
|
|
RIP
|
receptor interacting protein
|
|
RAS
|
renin-angiotensin system
|
|
RA
|
rheumatoid arthritis
|
|
SUMO
|
small ubiquitin-related modifier
|
|
SALM
|
synaptic adherence-like molecule
|
|
SCLC
|
small cell lung cancer
|
|
S100A11
|
S100 calcium-binding protein A11
|
|
SHR-SP
|
spontaneous hypertensive stroke
predisposition
|
|
SHR
|
spontaneously hypertensive rats
|
|
SAH
|
subarachnoid hemorrhage
|
|
SERT
|
presynaptic high-affinity 5-HT
transporter
|
|
SIP
|
SERT-interacting protein
|
|
SDF-1α
|
stromal cell-derived factor
|
|
TSEs
|
transmissible spongiform
encephalopathy
|
|
TUG1
|
taurine upregulated gene 1
|
|
TNM
|
tumor lymph node metastasis
|
|
TGF-β1
|
transforming growth factor β1
|
|
TNF
|
tumor necrosis factor
|
|
TNFR
|
TNF-α receptor
|
|
TRAFs
|
TNF receptor-associated factors
|
|
TAK1
|
TGF-β-activated kinase-1
|
|
T2DM
|
type 2 diabetes mellitus
|
|
VGLUT1
|
vesicular glutamate transporter 1
|
|
VPS33B
|
vacuolar protein sorting protein
33b
|
|
ZDHHC-19
|
zinc finger DHHC domain-containing
protein palmitoyltransferase-19
|
|
3′-UTR
|
3′-untranslated region
|
|
αc5-HT
|
5-hydroxytryptamine
|
References
|
1
|
Zhang L, Mao Y, Mao Q, Fan W, Xu L, Chen
Y, Xu L and Wang J: FLOT1 promotes tumor development, induces
epithelial-mesenchymal transition, and modulates the cell cycle by
regulating the Erk/Akt signaling pathway in lung adenocarcinoma.
Thorac Cancer. 10:909–917. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Swanwick CC, Shapiro ME, Vicini S and
Wenthold RJ: Flotillin-1 promotes formation of glutamatergic
synapses in hippocampal neurons. Dev Neurobiol. 70:875–883.
2010.PubMed/NCBI
|
|
3
|
Xiong Q, Lin M, Huang W and Rikihisa Y:
Infection by anaplasma phagocytophilum requires recruitment of
low-density lipoprotein cholesterol by flotillins. mBio.
10:e02783–e02718. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li H, Zhang Y, Chen SW, Li FJ, Zhuang SM,
Wang LP, Zhang J and Song M: Prognostic significance of Flotillin1
expression in clinically N0 tongue squamous cell cancer. Int J Clin
Exp Pathol. 7:996–1003. 2014.PubMed/NCBI
|
|
5
|
Jang D, Kwon H, Choi M, Lee J and Pak Y:
Sumoylation of flotillin-1 promotes EMT in metastatic prostate
cancer by suppressing snail degradation. Oncogene. 38:3248–3260.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ren K, Gao C, Zhang J, Wang K, Xu Y, Wang
SB, Wang H, Tian C, Shi Q and Dong XP: Flotillin-1 mediates PrPc
endocytosis in the cultured cells during Cu(2)(+) stimulation
through molecular interaction. Mol Neurobiol. 48:631–646. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Park MY, Kim N, Wu LL, Yu GY and Park K:
Role of flotillins in the endocytosis of GPCR in salivary gland
epithelial cells. Biochem Biophys Res Commun. 476:237–244. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Persaud-Sawin DA, Banach L and Harry GJ:
Raft aggregation with specific receptor recruitment is required for
microglial phagocytosis of Abeta42. Glia. 57:320–335. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lin C, Wu Z, Lin X, Yu C, Shi T, Zeng Y,
Wang X, Li J and Song L: Knockdown of FLOT1 impairs cell
proliferation and tumorigenicity in breast cancer through
upregulation of FOXO3a. Clin Cancer Res. 17:3089–3099. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Flemming JP, Hill BL, Haque MW, Raad J,
Bonder CS, Harshyne LA, Rodeck U, Luginbuhl A, Wahl JK III, Tsai
KY, et al: miRNA- and cytokine-associated extracellular vesicles
mediate squamous cell carcinomas. J Extracell Vesicles.
9:17901592020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Niu Y, Shao Z, Wang H, Yang J, Zhang F,
Luo Y, Xu L, Ding Y and Zhao L: LASP1-S100A11 axis promotes
colorectal cancer aggressiveness by modulating TGFβ/smad signaling.
Sci Rep. 6:261122016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Riento K, Frick M, Schafer I and Nichols
BJ: Endocytosis of flotillin-1 and flotillin-2 is regulated by Fyn
kinase. J Cell Sci. 122:912–918. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kwon H, Choi M, Ahn Y, Jang D and Pak Y:
Flotillin-1 palmitoylation turnover by APT-1 and ZDHHC-19 promotes
cervical cancer progression by suppressing IGF-1 receptor
desensitization and proteostasis. Cancer Gene Ther. 30:302–312.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yap J, Foo CF, Lee MY, Stanton PG and
Dimitriadis E: Proteomic analysis identifies interleukin 11
regulated plasma membrane proteins in human endometrial epithelial
cells in vitro. Reprod Biol Endocrinol. 9:732011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cremona ML, Matthies HJ, Pau K, Bowton E,
Speed N, Lute BJ, Anderson M, Sen N, Robertson SD, Vaughan RA, et
al: Flotillin-1 is essential for PKC-triggered endocytosis and
membrane microdomain localization of DAT. Nat Neurosci. 14:469–477.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kim JM, Cha SH, Choi YR, Jou I, Joe EH and
Park SM: DJ-1 deficiency impairs glutamate uptake into astrocytes
via the regulation of flotillin-1 and caveolin-1 expression. Sci
Rep. 6:288232016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dam DHM, Jelsma SA, Yu JM, Liu H, Kong B
and Paller AS: Flotillin and AP2A1/2 promote IGF-1 receptor
association with clathrin and internalization in primary human
keratinocytes. J Invest Dermatol. 140:1743–1752. e17442020.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tsui-Pierchala BA, Encinas M, Milbrandt J
and Johnson EM Jr: Lipid rafts in neuronal signaling and function.
Trends Neurosci. 25:412–417. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Estebanez B, Visavadiya NP, de Paz JA,
Whitehurst M, Cuevas MJ, González-Gallego J and Huang CJ:
Resistance training diminishes the expression of exosome CD63
protein without modification of plasma miR-146a-5p and cfDNA in the
elderly. Nutrients. 13:6652021. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kapahnke M, Banning A and Tikkanen R:
Random splicing of several exons caused by a single base change in
the target exon of CRISPR/Cas9 mediated gene knockout. Cells.
5:452016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Solis GP, Hoegg M, Munderloh C, Schrock Y,
Malaga-Trillo E, Rivera-Milla E and Stuermer CAO: Reggie/flotillin
proteins are organized into stable tetramers in membrane
microdomains. Biochem J. 403:313–322. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Swanwick CC, Shapiro ME, Yi Z, Chang K and
Wenthold RJ: NMDA receptors interact with flotillin-1 and −2, lipid
raft-associated proteins. FEBS Lett. 583:1226–1230. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Babuke T, Ruonala M, Meister M, Amaddii M,
Genzler C, Esposito A and Tikkanen R: Hetero-oligomerization of
reggie-1/flotillin-2 and reggie-2/flotillin-1 is required for their
endocytosis. Cell Signal. 21:1287–1297. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yau TY, Sander W, Eidson C and Courey AJ:
SUMO interacting motifs: Structure and function. Cells.
10:28252021. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mohanraj N, Joshi NS, Poulose R, Patil RR,
Santhoshkumar R, Kumar A, Waghmare GP, Saha AK, Haider SZ,
Markandeya YS, et al: A proteomic study to unveil lead
toxicity-induced memory impairments invoked by synaptic
dysregulation. Toxicol Rep. 9:1501–1513. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Walton JR, Frey HA, Vandre DD, Kwiek JJ,
Ishikawa T, Takizawa T, Robinson JM and Ackerman WE IV: Expression
of flotillins in the human placenta: Potential implications for
placental transcytosis. Histochem Cell Biol. 139:487–500. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Polo S, Pece S and Di Fiore PP:
Endocytosis and cancer. Curr Opin Cell Biol. 16:156–161. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tomiyama A, Uekita T, Kamata R, Sasaki K,
Takita J, Ohira M, Nakagawara A, Kitanaka C, Mori K, Yamaguchi H
and Sakai R: Flotillin-1 regulates oncogenic signaling in
neuroblastoma cells by regulating ALK membrane association. Cancer
Res. 74:3790–3801. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen K, Wu Q, Hu K, Yang C, Wu X, Cheung P
and Williams KJ: Suppression of hepatic FLOT1 (Flotillin-1) by type
2 diabetes mellitus impairs the disposal of remnant lipoproteins
via syndecan-1. Arterioscler Thromb Vasc Biol. 38:102–113. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kobayashi J, Hasegawa T, Sugeno N, Yoshida
S, Akiyama T, Fujimori K, Hatakeyama H, Miki Y, Tomiyama A, Kawata
Y, et al: Extracellular α-synuclein enters dopaminergic cells by
modulating flotillin-1-assisted dopamine transporter endocytosis.
FASEB J. 33:10240–10256. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Craig AM and Banker G: Neuronal polarity.
Annu Rev Neurosci. 17:267–310. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Swanwick CC, Shapiro ME, Vicini S and
Wenthold RJ: Flotillin-1 mediates neurite branching induced by
synaptic adhesion-like molecule 4 in hippocampal neurons. Mol Cell
Neurosci. 45:213–225. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Stuermer CA: How reggies regulate
regeneration and axon growth. Cell Tissue Res. 349:71–77. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Stuermer CA: The reggie/flotillin
connection to growth. Trends Cell Biol. 20:6–13. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wehmeyer L, Du Toit A, Lang DM and Hapgood
JP: Lipid raft- and protein kinase C-mediated synergism between
glucocorticoid- and gonadotropin-releasing hormone signaling
results in decreased cell proliferation. J Biol Chem.
289:10235–10251. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hada N, Okayasu M, Ito J, Nakayachi M,
Hayashida C, Kaneda T, Uchida N, Muramatsu T, Koike C, Masuhara M,
et al: Receptor activator of NF-kappaB ligand-dependent expression
of caveolin-1 in osteoclast precursors, and high dependency of
osteoclastogenesis on exogenous lipoprotein. Bone. 50:226–236.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ficht X, Ruef N, Stolp B, Samson GPB,
Moalli F, Page N, Merkler D, Nichols BJ, Diz-Muñoz A, Legler DF, et
al: In vivo function of the lipid raft protein flotillin-1 during
CD8(+) T cell-mediated host surveillance. J Immunol. 203:2377–2387.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang X, Guan H, Liu W, Li H, Ding J, Feng
Y and Chen Z: Identification of immune markers in dilated
cardiomyopathies with heart failure by integrated weighted gene
coexpression network analysis. Genes (Basel). 13:3932022.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Korhonen JT, Puolakkainen M, Häivälä R,
Penttilä T, Haveri A, Markkula E and Lahesmaa R: Flotillin-1
(Reggie-2) contributes to Chlamydia pneumoniae growth and is
associated with bacterial inclusion. Infect Immun. 80:1072–1078.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ji H, Greening DW, Barnes TW, Lim JW,
Tauro BJ, Rai A, Xu R, Adda C, Mathivanan S, Zhao W, et al:
Proteome profiling of exosomes derived from human primary and
metastatic colorectal cancer cells reveal differential expression
of key metastatic factors and signal transduction components.
Proteomics. 13:1672–1686. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Reutgen H, Eckert H and Christ R: The
Kveim test in the diagnosis of sarcoidosis. Z Erkr Atmungsorgane.
170:306–313. 1988.(In German). PubMed/NCBI
|
|
42
|
Liu C, Shang Z, Ma Y, Ma J and Song J:
HOTAIR/miR-214-3p/FLOT1 axis plays an essential role in the
proliferation, migration, and invasion of hepatocellular carcinoma.
Int J Clin Exp Pathol. 12:50–63. 2019.PubMed/NCBI
|
|
43
|
Kan XQ, Li YB, He B, Cheng S, Wei Y and
Sun J: MiR-1294 acts as a tumor inhibitor in cervical cancer by
regulating FLOT1 expression. J Biol Regul Homeost Agents.
34:10.23812/20–10A. 2020.
|
|
44
|
Cai S, Zhou Y, Pan Y, Liu P, Yu K and Chen
S: Long non-coding RNA A1BG-AS1 promotes tumorigenesis in breast
cancer by sponging microRNA-485-5p and consequently increasing
expression of FLOT1 expression. Hum Cell. 34:1517–1531. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lv D, Xiang Y, Yang Q, Yao J and Dong Q:
Long non-coding RNA TUG1 promotes cell proliferation and inhibits
cell apoptosis, autophagy in clear cell renal cell carcinoma via
MiR-31-5p/FLOT1 axis. Onco Targets Ther. 13:5857–5868. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Song L, Gong H, Lin C, Wang C, Liu L, Wu
J, Li M and Li J: Flotillin-1 promotes tumor necrosis factor-alpha
receptor signaling and activation of NF-κB in esophageal squamous
cell carcinoma cells. Gastroenterology. 143:995–1005. e10122012.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Li H, Wang RM, Liu SG, Zhang JP, Luo JY,
Zhang BJ and Zhang XG: Abnormal expression of FLOT1 correlates with
tumor progression and poor survival in patients with non-small cell
lung cancer. Tumour Biol. 35:3311–3315. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Li J, Zuo X, Shi J, Zhang J, Duan X and Xu
G: Flotillin 1 is differentially expressed in human epithelial
ovarian tumors. Neoplasma. 65:561–571. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Guan Y, Song H, Zhang G and Ai X:
Overexpression of flotillin-1 is involved in proliferation and
recurrence of bladder transitional cell carcinoma. Oncol Rep.
32:748–754. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zhao L, Li J, Liu Y, Zhou W, Shan Y, Fan
X, Zhou X, Shan B, Song Y and Zhan Q: Flotillin1 promotes EMT of
human small cell lung cancer via TGF-β signaling pathway. Cancer
Biol Med. 15:400–414. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Baig N, Li Z, Lu J, Chen H, Yu S, Li T,
Niu Z and Niu J: Clinical significance and comparison of flotillin
1 expression in left and right colon cancer. Oncol Lett.
18:997–1004. 2019.PubMed/NCBI
|
|
52
|
Cao S, Cui Y, Xiao H, Mai M, Wang C, Xie
S, Yang J, Wu S, Li J, Song L, et al: Upregulation of flotillin-1
promotes invasion and metastasis by activating TGF-beta signaling
in nasopharyngeal carcinoma. Oncotarget. 7:4252–4264. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Wang R, Chen Z, Zhang Y, Ziao S, Zhang W,
Hu X, Xiao Q, Liu Q and Wang X: Flotillin-1 is a prognostic
biomarker for glioblastoma and promotes cancer development through
enhancing invasion and altering tumour microenvironment. J Cell Mol
Med. 27:392–402. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Iorio MV and Croce CM: microRNA
involvement in human cancer. Carcinogenesis. 33:1126–1133. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Butz H, Szabo PM, Khella HW, Nofech-Mozes
R, Patocs A and Yousef GM: miRNA-target network reveals miR-124as a
key miRNA contributing to clear cell renal cell carcinoma
aggressive behaviour by targeting CAV1 and FLOT1. Oncotarget.
6:12543–12557. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Yang FQ, Zhang HM, Chen SJ, Yan Y and
Zheng JH: MiR-506 is down-regulated in clear cell renal cell
carcinoma and inhibits cell growth and metastasis via targeting
FLOT1. PLoS One. 10:e01202582015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zhang B, Zhang M, Yang Y, Li Q, Yu J, Zhu
S, Niu Y and Shang Z: Targeting KDM4A-AS1 represses AR/AR-Vs
deubiquitination and enhances enzalutamide response in CRPC.
Oncogene. 41:387–399. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Zhang Y, Cheng X, Liang H and Jin Z: Long
non-coding RNA HOTAIR and STAT3 synergistically regulate the
cervical cancer cell migration and invasion. Chem Biol Interact.
286:106–110. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Wang Y, Wang Z, Cheng K and Hao Q: FAM201A
promotes cervical cancer progression and metastasis through
miR-1271-5p/Flotillin-1 axis targeting-induced Wnt/β-catenin
pathway. J Oncol. 2022:11238392022. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Li X, Zhang F, Ma J, Ruan X, Liu X, Zheng
J, Liu Y, Cao S, Shen S, Shao L, et al: NCBP3/SNHG6 inhibits GBX2
transcription in a histone modification manner to facilitate the
malignant biological behaviour of glioma cells. RNA Biol. 18:47–63.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Yang PW, Lu ZY, Pan Q, Chen TT, Feng XJ,
Wang SM, Pan YC, Zhu MH and Zhang SH: MicroRNA-6809-5p mediates
luteolin-induced anticancer effects against hepatoma by targeting
flotillin 1. Phytomedicine. 57:18–29. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Li L, Luo J, Wang B, Wang D, Xie X, Yuan
L, Guo J, Xi S, Gao J, Lin X, et al: Microrna-124 targets
flotillin-1 to regulate proliferation and migration in breast
cancer. Mol Cancer. 12:1632013. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Gong H, Song L, Lin C, Liu A, Lin X, Wu J,
Li M and Li J: Downregulation of miR-138 sustains NF-κB activation
and promotes lipid raft formation in esophageal squamous cell
carcinoma. Clin Cancer Res. 19:1083–1093. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Pastushenko I and Blanpain C: EMT
transition states during tumor progression and metastasis. Trends
Cell Biol. 29:212–226. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Nachiyappan A, Gupta N and Taneja R:
EHMT1/EHMT2 in EMT, cancer stemness and drug resistance: Emerging
evidence and mechanisms. FEBS J. 289:1329–1351. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Peng L, Wen L, Shi QF, Gao F, Huang B,
Meng J, Hu CP and Wang CM: Scutellarin ameliorates pulmonary
fibrosis through inhibiting NF-κB/NLRP3-mediated
epithelial-mesenchymal transition and inflammation. Cell Death Dis.
11:9782020. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Wang W, Dong L, Zhao B, Lu J and Zhao Y:
E-cadherin is downregulated by microenvironmental changes in
pancreatic cancer and induces EMT. Oncol Rep. 40:1641–1649.
2018.PubMed/NCBI
|
|
68
|
Miyazono K: Transforming growth
factor-beta signaling in epithelial-mesenchymal transition and
progression of cancer. Proc Jpn Acad Ser B Phys Biol Sci.
85:314–323. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Roshan MK, Soltani A, Soleimani A,
Kahkhaie KR, Afshari AR and Soukhtanloo M: Role of AKT and mTOR
signaling pathways in the induction of epithelial-mesenchymal
transition (EMT) process. Biochimie. 165:229–234. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Wang Y, Shi J, Chai K, Ying X and Zhou BP:
The role of snail in EMT and tumorigenesis. Curr Cancer Drug
Targets. 13:963–972. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Liang Z, Wang X, Xu X, Xie B, Ji A, Meng
S, Li S, Zhu Y, Wu J, Hu Z, et al: MicroRNA-608 inhibits
proliferation of bladder cancer via AKT/FOXO3a signaling pathway.
Mol Cancer. 16:962017. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Xu X, Wu J, Li S, Hu Z, Xu X, Zhu Y, Liang
Z, Wang X, Lin Y, Mao Y, et al: Downregulation of microRNA-182-5p
contributes to renal cell carcinoma proliferation via activating
the AKT/FOXO3a signaling pathway. Mol Cancer. 13:1092014.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Ye DM, Ye SC, Yu SQ, Shu FF, Xu SS, Chen
QQ, Wang YL, Tang ZT and Pan C: Drug-resistance reversal in
colorectal cancer cells by destruction of flotillins, the key lipid
rafts proteins. Neoplasma. 66:576–583. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Shui HA, Hsia CW, Chen HM, Chang TC and
Wang CY: Proteomics and bioinformatics analysis of
lovastatin-induced differentiation in ARO cells. J Proteomics.
75:1170–1180. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Arkhipova KA, Sheyderman AN, Laktionov KK,
Mochalnikova VV and Zborovskaya IB: Simultaneous expression of
flotillin-1, flotillin-2, stomatin and caveolin-1 in non-small cell
lung cancer and soft tissue sarcomas. BMC Cancer. 14:1002014.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Gu H, Chen C, Hao X, Wang C, Zhang X, Li
Z, Shao H, Zeng H, Yu Z, Xie L, et al: Sorting protein VPS33B
regulates exosomal autocrine signaling to mediate hematopoiesis and
leukemogenesis. J Clin Invest. 126:4537–4553. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Fountzilas E, Kotoula V, Angouridakis N,
Karasmanis I, Wirtz RM, Eleftheraki AG, Veltrup E, Markou K,
Nikolaou A, Pectasides D and Fountzilas G: Identification and
validation of a multigene predictor of recurrence in primary
laryngeal cancer. PLoS One. 8:e704292013. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Dai S, Xu S, Ye Y and Ding K:
Identification of an immune-related gene signature to improve
prognosis prediction in colorectal cancer patients. Front Genet.
11:6070092020. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Zhang Y, Li J, Song Y, Chen F, Pei Y and
Yao F: Flotillin-1 expression in human clear-cell renal cell
carcinoma is associated with cancer progression and poor patient
survival. Mol Med Rep. 10:860–866. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Peters FS, Peeters AMA, Mandaviya PR, van
Meurs JBJ, Hofland LJ, van de Wetering J, Betjes MGH, Baan CC and
Boer K: Differentially methylated regions in T cells identify
kidney transplant patients at risk for de novo skin cancer. Clin
Epigenetics. 10:812018. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Zafar A, Wang W, Liu G, Wang X, Xian W,
McKeon F, Foster J, Zhou J and Zhang R: Molecular targeting
therapies for neuroblastoma: Progress and challenges. Med Res Rev.
41:961–1021. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Messiha BAS, Ali MRA, Khattab MM and
Abo-Youssef AM: Perindopril ameliorates experimental Alzheimer's
disease progression: Role of amyloid β degradation, central
estrogen receptor and hyperlipidemic-lipid raft signaling.
Inflammopharmacology. 28:1343–1364. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Hattori C, Asai M, Onishi H, Sasagawa N,
Hashimoto Y, Saido TC, Maruyama K, Mizutani S and Ishiura S: BACE1
interacts with lipid raft proteins. J Neurosci Res. 84:912–917.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Krance SH, Wu CY, Chan ACY, Kwong S, Song
BX, Xiong LY, Ouk M, Chen MH, Zhang J, Yung A, et al:
Endosomal-lysosomal and autophagy pathway in alzheimer's disease: A
systematic review and meta-analysis. J Alzheimers Dis.
88:1279–1292. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Cabreira V and Massano J: Parkinson's
disease: Clinical review and update. Acta Med Port. 32:661–670.
2019.(In Portuguese). View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Schrader JM, Stanisavljevic A, Xu F and
Van Nostrand WE: Distinct brain proteomic signatures in cerebral
small vessel disease rat models of hypertension and cerebral
amyloid angiopathy. J Neuropathol Exp Neurol. 81:731–745. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Cheng Y, Sun D, Zhu B, Zhou W, Lv C, Kou F
and Wei H: Integrative metabolic and proteomic profiling of the
brainstem in spontaneously hypertensive rats. J Proteome Res.
19:4114–4124. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Zhao H, Li ST, Zhu J, Hua XM and Wan L:
Analysis of peripheral blood cells' transcriptome in patients with
subarachnoid hemorrhage from ruptured aneurysm reveals potential
biomarkers. World Neurosurg. 129:e16–e22. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Quinlan MA, Robson MJ, Ye R, Rose KL,
Schey KL and Blakely RD: Ex vivo quantitative proteomic analysis of
serotonin transporter interactome: Network impact of the SERT Ala56
coding variant. Front Mol Neurosci. 13:892020. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Zhong J, Li S, Zeng W, Li X, Gu C, Liu J
and Luo XJ: Integration of GWAS and brain eQTL identifies FLOT1 as
a risk gene for major depressive disorder. Neuropsychopharmacology.
44:1542–1551. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Liu W, Li W, Cai X, Yang Z, Li H, Su X,
Song M, Zhou DS, Li X, Zhang C, et al: Identification of a
functional human-unique 351-bp Alu insertion polymorphism
associated with major depressive disorder in the 1p31.1 GWAS risk
loci. Neuropsychopharmacology. 45:1196–1206. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Reisinger SN, Kong E, Molz B, Humberg T,
Sideromenos S, Cicvaric A, Steinkellner T, Yang JW, Cabatic M,
Monje FJ, et al: Flotillin-1 interacts with the serotonin
transporter and modulates chronic corticosterone response. Genes
Brain Behav. 18:e124822019. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Roura S, Galvez-Monton C, Pujal JM, Casani
L, Fernández MA, Astier L, Gastelurrutia P, Domingo M, Prat-Vidal
C, Soler-Botija C, et al: New insights into lipid raft function
regulating myocardial vascularization competency in human
idiopathic dilated cardiomyopathy. Atherosclerosis. 230:354–364.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Kerr JR, Kaushik N, Fear D, Baldwin DA,
Nuwaysir EF and Adcock IM: Single-nucleotide polymorphisms
associated with symptomatic infection and differential human gene
expression in healthy seropositive persons each implicate the
cytoskeleton, integrin signaling, and oncosuppression in the
pathogenesis of human parvovirus B19 infection. J Infect Dis.
192:276–286. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
95
|
Lim WC and Chow VT: Gene expression
profiles of U937 human macrophages exposed to Chlamydophila
pneumoniae and/or low density lipoprotein in five study models
using differential display and real-time RT-PCR. Biochimie.
88:367–377. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Douadi C, Vazeille E, Chambon C, Hébraud
M, Fargeas M, Dodel M, Coban D, Pereira B, Birer A, Sauvanet P, et
al: Anti-TNF agents restrict adherent-invasive escherichia coli
replication within macrophages through modulation of chitinase
3-like 1 in patients with Crohn's Disease. J Crohns Colitis.
16:1140–1150. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Schmidt F, Thywissen A, Goldmann M, Cunha
C, Cseresnyés Z, Schmidt H, Rafiq M, Galiani S, Gräler MH, Chamilos
G, et al: Flotillin-dependent membrane microdomains are required
for functional phagolysosomes against fungal infections. Cell Rep.
32:1080172020. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Salazar J, Angarita L, Morillo V, Navarro
C, Martínez MS, Chacín M, Torres W, Rajotia A, Rojas M, Cano C, et
al: Microbiota and diabetes mellitus: Role of lipid mediators.
Nutrients. 12:30392020. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Rojas-Carranza CA, Bustos-Cruz RH,
Pino-Pinzon CJ, Ariza-Marquez YV, Gomez-Bello RM and Canadas-Garre
M: Diabetes-related neurological implications and pharmacogenomics.
Curr Pharm Des. 24:1695–1710. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Abu-Farha M, Abubaker J and Tuomilehto J:
ANGPTL8 (betatrophin) role in diabetes and metabolic diseases.
Diabetes Metab Res Rev. 33:10.1002/dmrr.29. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Zhao Z, Deng X, Jia J, Zhao L, Wang C, Cai
Z, Guo C, Yang L, Wang D, Ma S, et al: Angiopoietin-like protein 8
(betatrophin) inhibits hepatic gluconeogenesis through PI3K/Akt
signaling pathway in diabetic mice. Metabolism. 126:1549212022.
View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Siddiqa A, Cirillo E, Tareen SHK, Ali A,
Kutmon M, Eijssen LMT, Ahmad J, Evelo CT and Coort SL: Biological
pathways leading from ANGPTL8 to diabetes mellitus-a co-expression
network based analysis. Front Physiol. 9:18412018. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Hansen JS, Zhao X, Irmler M, Liu X, Hoene
M, Scheler M, Li Y, Beckers J, de Angelis MH, Häring HU, et al:
Type 2 diabetes alters metabolic and transcriptional signatures of
glucose and amino acid metabolism during exercise and recovery.
Diabetologia. 58:1845–1854. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Spitzel M, Wagner E, Breyer M, Henniger D,
Bayin M, Hofmann L, Mauceri D, Sommer C and Üçeyler N:
Dysregulation of immune response mediators and pain-related ion
channels is associated with pain-like behavior in the GLA KO mouse
model of fabry disease. Cells. 11:17302022. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Zhang X, Cui Y, Ding X, Liu S, Han B, Duan
X, Zhang H and Sun T: Analysis of mRNAlncRNA and mRNAlncRNA-pathway
coexpression networks based on WGCNA in developing pediatric
sepsis. Bioengineered. 12:1457–1470. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Mo XB, Sun YH, Zhang YH and Lei SF:
Integrative analysis highlighted susceptibility genes for
rheumatoid arthritis. Int Immunopharmacol. 86:1067162020.
View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Zhu H, Xia W, Mo XB, Lin X, Qiu YH, Yi NJ,
Zhang YH, Deng FY and Lei SF: Gene-based genome-wide association
analysis in european and asian populations identified novel genes
for rheumatoid arthritis. PLoS One. 11:e01672122016. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Dai H, Zhou J and Zhu B: Gene
co-expression network analysis identifies the hub genes associated
with immune functions for nocturnal hemodialysis in patients with
end-stage renal disease. Medicine (Baltimore). 97:e120182018.
View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Prates L, Lemes RB, Hunemeier T and
Leonardi F: Population-based change-point detection for the
identification of homozygosity islands. Bioinformatics.
39:btad1702023. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
McDermaid A, Monier B, Zhao J, Liu B and
Ma Q: Interpretation of differential gene expression results of
RNA-seq data: Review and integration. Brief Bioinform.
20:2044–2054. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Yu G, Wang LG, Han Y and He QY:
clusterProfiler: An R package for comparing biological themes among
gene clusters. OMICS. 16:284–287. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Guo AY, Liang XJ, Liu RJ, Li XX, Bi W,
Zhou LY, Tang CE, Yan A, Chen ZC and Zhang PF: Flotilin-1 promotes
the tumorigenicity and progression of malignant phenotype in human
lung adenocarcinoma. Cancer Biol Ther. 18:715–722. 2017. View Article : Google Scholar : PubMed/NCBI
|