Introduction
Cerebrovascular disease (CVD) has become a global
public health problem due to its high morbidity, association with
disability and recurrence rates. In China, the incidence of CVD in
the population is increasing annually and ischemic stroke is the
most common type of CVD; ~70% of patients have ischemic stroke and
the recurrence rate is as high as 17.7% (1,2).
There is currently a lack of effective treatment methods for
ischemic stroke, An effective approach after cerebrovascular
embolism in clinical practice is timely thrombolysis or
endovascular intervention, intended to restore blood perfusion in
the ischemic area; yet at the same time it can induce
ischemia-reperfusion injury, which leads to brain damage both in
the ischemic core and penumbra area. An ischemic stroke consists of
two related pathological injury processes: Primary ischemia-induced
brain injury and secondary ischemia reperfusion injury (3). A study has shown that neurons in the
ischemic penumbra may undergo apoptosis hours or days after
ischemia and alleviating ischemia reperfusion injury is an
achievable therapeutic goal in the early intervention of ischemic
stroke aimed at limiting the amount of infarction (4). When ischemic stroke occurs, cerebral
ischemia and hypoxia cause the release of excessive excitatory
amino acids, mainly glutamic acid and aspartic acid, which exert
excitotoxic effects on the central nervous system. These
excitotoxic effects play important roles in neuronal and
blood-brain barrier damage after cerebral ischemia (5,6).
Extracellular excitatory amino acids are mainly transported into
cells by excitatory amino acid transporters (EAATs) expressed on
astrocytes to avoid excessive excitation of neurons. EAAT2 accounts
for 80–90% of this activity of extracellular excitatory amino acid
uptake (7). Changes in EAAT2
function are closely related to excitotoxicity in the central
nervous system. Heat shock protein (HSP) 90 is abundant in cells,
mainly in the form of homodimers, including HSP90α and HSP90β. As a
molecular chaperone, HSP90 regulates the conformational maturation
and functional stability of many signaling proteins in cells,
serves important roles in cell growth, differentiation, apoptosis
and tumor development and is an important antitumor target
(8,9). A study found that HSP90α is not
necessary in mammals, but HSP90β is (10), suggesting that HSP90α and HSP90β
have different physiological roles. Significantly increased HSP90
expression levels has been observed in human patients with
hippocampal sclerosis and in a mouse model of epilepsy. The
increase in the HSP90β expression level was predominant, while the
increase in the HSP90α expression level was not significant
(11), further supporting the
different roles of HSP90α and HSP90β. At present, the role of
HSP90β in ischemic brain injury has not been reported. The purpose
of the present study was to clarify the changes in HSP90β
expression in a rat cerebral ischemia-reperfusion model and the
effects of these changes on the expression of EAAT2 and to further
explore the potential of HSP90β as a molecular target for reducing
brain ischemia-reperfusion injury.
Materials and methods
Laboratory animals
A total of 87 Healthy adult Sprague-Dawley (SD) male
rats weighing 280–300 g (6–8 weeks) were purchased from Changsha
Tianqin Biological Technology Company [license number: SCXK (Xiang)
2019–0004]. The animals were reared in separate cages with 12-h
light/dark cycle at a constant temperature (24±1°C) and 55±5%
humidity with free access to food and water during the experimental
period. The experimental groups included the control group,
sham-operated group, middle cerebral artery occlusion (MCAO) group,
empty virus group and lentivirus group. The rats were anaesthetized
with 1% pentobarbital sodium (40 mg/kg) before being sacrificed by
cervical dislocation. All efforts were made to minimize the
suffering of the rats. All procedures were approved by the Animal
Care and Use Committee of Zunyi Medical University, Zunyi, China
(approval number ZMU21-2203-487).
Establishment of a cerebral
ischemia-reperfusion model in rats through MCAO
The cerebral ischemia-reperfusion model was
established through MCAO (12,13).
The specific procedure was as follows: The rats were anesthetized
by administering an intraperitoneal injection of 1% pentobarbital
(40 mg/kg), a small incision in the skin to the right of the neck
was created and the carotid sheath was exposed. The common carotid
artery and external carotid artery were ligated. A small incision
was made at the common carotid artery ~4 mm from the bifurcation of
the internal and external carotid arteries and the MCAO suture was
inserted into the internal carotid artery. The length of the suture
was ~18–20 mm. The suture was fixed onto the internal carotid
artery. The incision was sterilized and sutured. After 2 h of
embolization, the suture was slowly removed to achieve reperfusion.
The sham-operated group underwent the same treatment, but the MCAO
suture was only inserted 10 mm into the internal carotid artery.
During the entire experimental operation, a 40 W light bulb
illuminated the surgical field and an electric blanket was used to
keep the rats warm. After the operation, rats were housed in
separate cages and they had access to a normal diet and drinking
water. Referring to a previous study (14), the modified neurological severity
score (mNSS) was determined 24 h after reperfusion and a higher
score indicated a more severe neurological deficit.
Reagents, constructs and
antibodies
The oligonucleotide sequence for HSP90β microRNA
interference (miRNAi) constructs was 5′-AACCGCATCTACCGCATGATT-3′.
Antibodies were purchased from Abcam [HSP90β, EAAT2, Bax, Bcl-2 and
glial fibrillary acidic protein (GFAP)], ZEN-BIO, Inc. (HSP90β and
EAAT2), Proteintech Group, Inc. (β-tubulin and β-actin) and
FUJIFILM Wako Pure Chemical Corporation ionized calcium-binding
adaptor molecule 1 (IBA1). CoraLite488-conjugated goat anti-rabbit
IgG (H+L) and CoraLite594-conjugated goat anti-mouse IgG (H+L) were
purchased from Proteintech Group, Inc. Hematoxylin and eosin
(H&E) and triphenyltetrazolium chloride (TTC) staining reagents
were purchased from Beijing Solarbio Science & Technology Co.,
Ltd.
Lentivirus construction and
injection
HSP90β lentivirus and empty virus were synthesized
and constructed by Shanghai Genechem Co., Ltd. HSPβ90 lentivirus
decreased expression of HSP90β. The three target sequence of HSPβ90
LV were designed before formal experiments (Table SI). The lentivirus was injected
into the right lateral ventricle. Using the fontanel as the origin,
a site 1.3 mm horizontally to the right and then 1.5 mm backward
was located and a hole was drilled at this site with an electric
miniature animal skull drill. Then, a 10 µl microinjector was used
to aspirate 10 µl of virus and the needle was slowly inserted
vertically 3.8 mm at the drilled hole. The empty virus group was
injected with empty virus as a control. Successful infection was
confirmed through immunofluorescence and western blotting which
screened highest transfection efficiency in decreasing HSP90β
(Fig. S1).
TTC staining
Following anesthesia, the rats were quickly
decapitated, the brain was removed and the brain tissue was placed
in a −20°C freezer (the olfactory bulb, cerebellum and lower brain
stem were removed) for 20 min. The brain tissue was divided into
five parts from front to back with a scalpel at the midpoint of the
connecting line between the anterior pole and the optic chiasm, the
optic chiasm, the infundibular stalk and between the infundibular
stalk and the caudal pole of the posterior lobe. The brain slices
were placed in a 1% TTC staining solution (37°C; 30 min) and
arranged in order of brain tissue structures from rostral to caudal
locations and then images were captured. The infarct volume ratio
was calculated using Image-Pro Plus software 6.0 (Media
Cybernetics, Inc.) to measure the area as follows: Infarct volume
ratio: (normal side volume-infarct side noninfarct volume)/normal
side volume
Immunofluorescence staining
Rat brain tissue was perfused with 4%
paraformaldehyde overnight at 4°C, and then rehydrated using a
graded ethanol series (70, 80, 90 and 100%) for 1.5 h at room
temperature, cleared in xylene twice and embedded in paraffin, then
sectioned to a thickness of 5 µm. Sections were dewaxed with
xylene, hydrated with a graded series of ethanol solutions and
placed in 3% H2O2 for 10 min at room
temperature to block endogenous peroxidase activity. Antigen
retrieval was performed by high-pressure repair for 2 min in 0.01
mol/l citrate (pH=6.0; ~120°C). Following blocking at room
temperature for 30 min using the goat serum (cat no. C0265;
Beyotime Institute of Biotechnology) blocking solution, the primary
antibody (HSP90β; cat no. R380807; ZEN-BIO, Inc.; GFAP, cat no.
ab4648; Abcam; IBA1; cat no. 019–19741; FUJIFILM Wako Pure Chemical
Corporation) was added and then incubated in the refrigerator at
4°C overnight. After thorough washing, a fluorescent dye-conjugated
secondary antibody (cat no. A0507; Beyotime Institute of
Biotechnology) was added and incubated for 30 min at room
temperature. After adding an anti-fluorescence quencher, sections
were mounted and images were captured for observation.
Western blotting
Rats were decapitated quickly following anesthesia
and brain tissue was collected. After removing the olfactory bulb,
cerebellum and lower brain stem, the brain tissue was quickly
placed in a −80°C freezer for later use. Radioimmunoprecipitation
assay (RIPA; cat no. R0010; Beijing Solarbio Science &
Technology Co., Ltd.) buffer containing protease inhibitors (PMSF;
cat no. P0100; Beijing Solarbio Science & Technology Co., Ltd.)
was added to the brain tissue, the tissue was fully disrupted with
a homogenizer and centrifuged for 30 min (4°C; 16,311 × g) and the
supernatant was collected for protein quantification. The
concentration of the supernatant was estimated using an Instant BCA
Protein Assay Kit (cat no. ZJ101; EpiZyme, Inc.). After adding 20
µl of the corresponding sample to SDS-PAGE gels (stacking, 5%;
separation, 10%), proteins were electrophoresed and transferred to
a polyvinylidene fluoride (PVDF) membrane. After blocking with 5%
non-fat milk powder at room temperature for 1 h, the membrane was
transferred into the diluted primary antibody (HSP90β; cat no.
ab203085; Abcam; EAAT2; cat no. ab205248; Abcam; HSP90α; cat no.
R380809; ZEN-BIO, Inc.; BAX, cat no. ab32503; Bcl-2, Abcam; cat no.
ab182858; Abcam; β-actin; cat no. 81115-1-RR; Proteintech Group,
Inc.; β-tubulin; cat no: 10094-1-AP; Proteintech Group, Inc.) and
placed in a 4°C refrigerator overnight. After washing, the
secondary antibody (HRP-conjugated Affinipure goat anti-rabbit IgG
(H+L); cat no. SA00001-2, Proteintech Group, Inc.) was added and
incubated for 1 h at room temperature before another washing step.
The PVDF membrane was incubated with the luminescent agent (cat no.
SQ202, EpiZyme, Inc.) for a sufficient period and then exposed to
determine the protein levels. Blots were imaged using ChemicDoc
Imaging System (Bio-Rad Laboratories, Inc.). ImageJ v1.52a
(National Institutes of Health) was used to assess the greyscale
values and SPSS 29.0 (IBM Corp.) was used to analyse the data.
Statistical methods
SPSS 29.0 (IBM Corp.) was used for statistical
analyses of experimental data, and the results are presented as the
means ± standard deviations (± s). The nonparametric Kruskal-Wallis
test was used to analyze the mNSS data. One-way ANOVA and Tukey's
post hoc test were used to compare differences between groups.
Statistical graphs were plotted using GraphPad Prism 6.0 software
(GraphPad; Dotmatics). P<0.05 was considered to indicate a
statistically significant difference.
Results
Assessment of the MCAO model
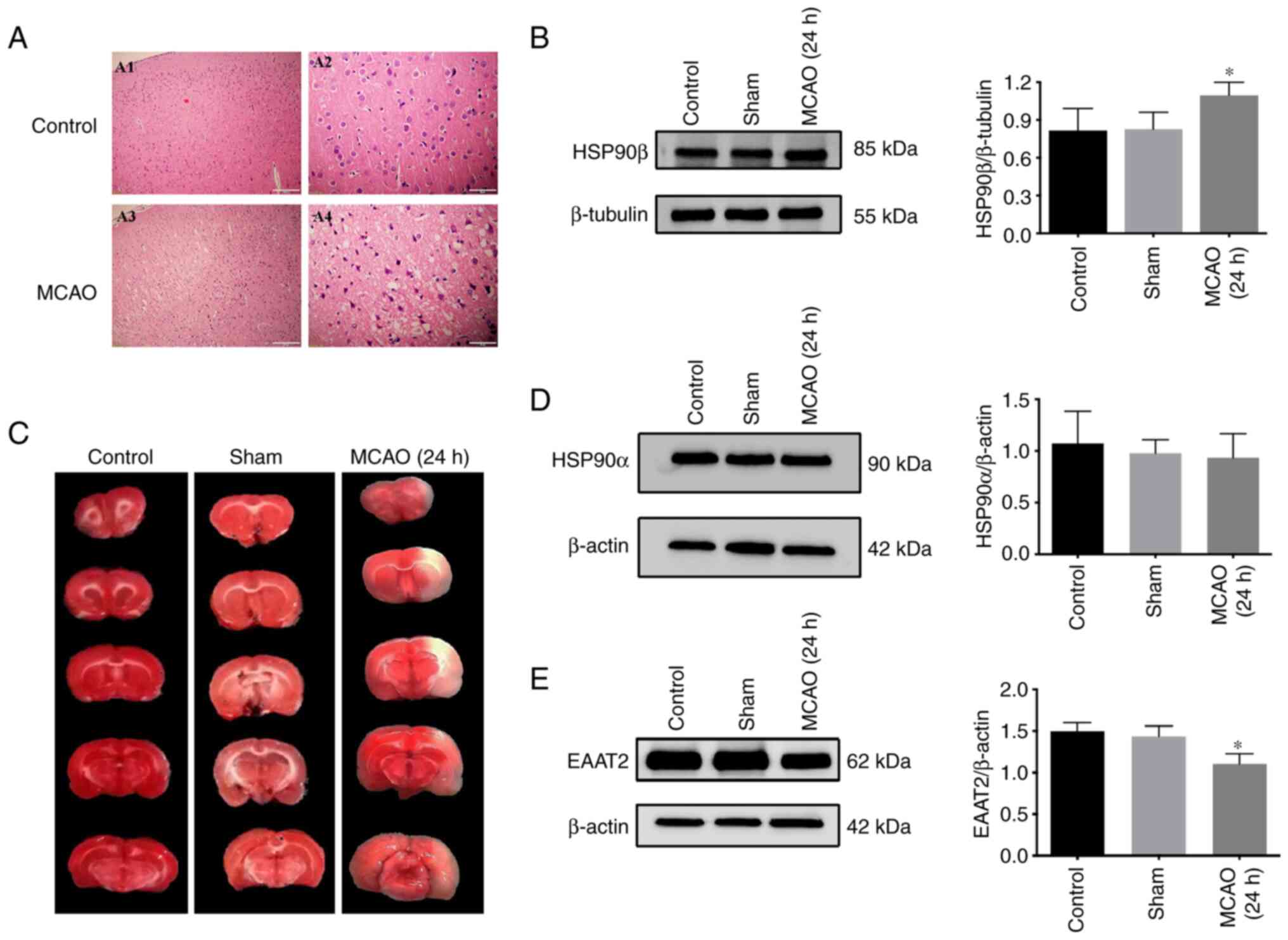
The MCAO model was first tested and H&E staining
performed to observe whether histopathological changes occurred in
rat brain tissue following ischemia and reperfusion. The neurons on
the noninfarcted side of the brain tissue were regularly arranged
and the nuclei were round, clear and uniform in size. In the
ischemic area of the infarcted side, most of the neurons in the
brain tissue were irregularly arranged and structurally disordered.
The nuclei were not clearly displayed and had various sizes; the
nuclei were also darker due to pyknosis (Fig. 1A). The results of TTC staining
showed red staining in the control group and the sham-operated
group and no infarction was detected. White infarcts were observed
in the brain tissue of rats after ischemia-reperfusion for 24 h and
the infarct volume ratio was significantly increased compared with
the control group and the sham-operated group (Fig. 1B).
Changes in the expression of HSP90β,
EAAT2 and HSP90α in the rat MCAO model
According to previous results from our research
group, the lowest EAAT2 expression was detected at 24 h of ischemia
and reperfusion. Therefore, 24 h of reperfusion was selected as the
observation point. Western botting was used to detect the changes
in protein expression in the brain tissue of rats from each group.
HSP90β, HSP90α and EAAT2 proteins were expressed in the brain
tissue of the control group, sham-operated group and MCAO group.
The expression level of HSP90β in the MCAO group was increased and
the difference was statistically significant compared with those in
the control group and the sham-operated group (Fig. 1C). The EAAT2 protein expression
level was significantly decreased compared with those in the
control group and sham-operated group (Fig. 1E). A significant change in HSP90α
expression was not observed (Fig.
1D).
Expression and localization of HSP90β
in brain tissue after ischemia-reperfusion in rats
Using immunohistofluorescence, glial cells were
simultaneously labeled with the astrocyte marker GFAP and the
microglial marker IBA1. The results did not reveal obvious
expression of HSP90 in microglia and astrocytes under normal
conditions. However, after cerebral ischemia-reperfusion in rats,
HSP90β expression coincided with astrocyte markers in the ischemic
penumbra area, while no expression was observed in microglia
(Fig. 2).
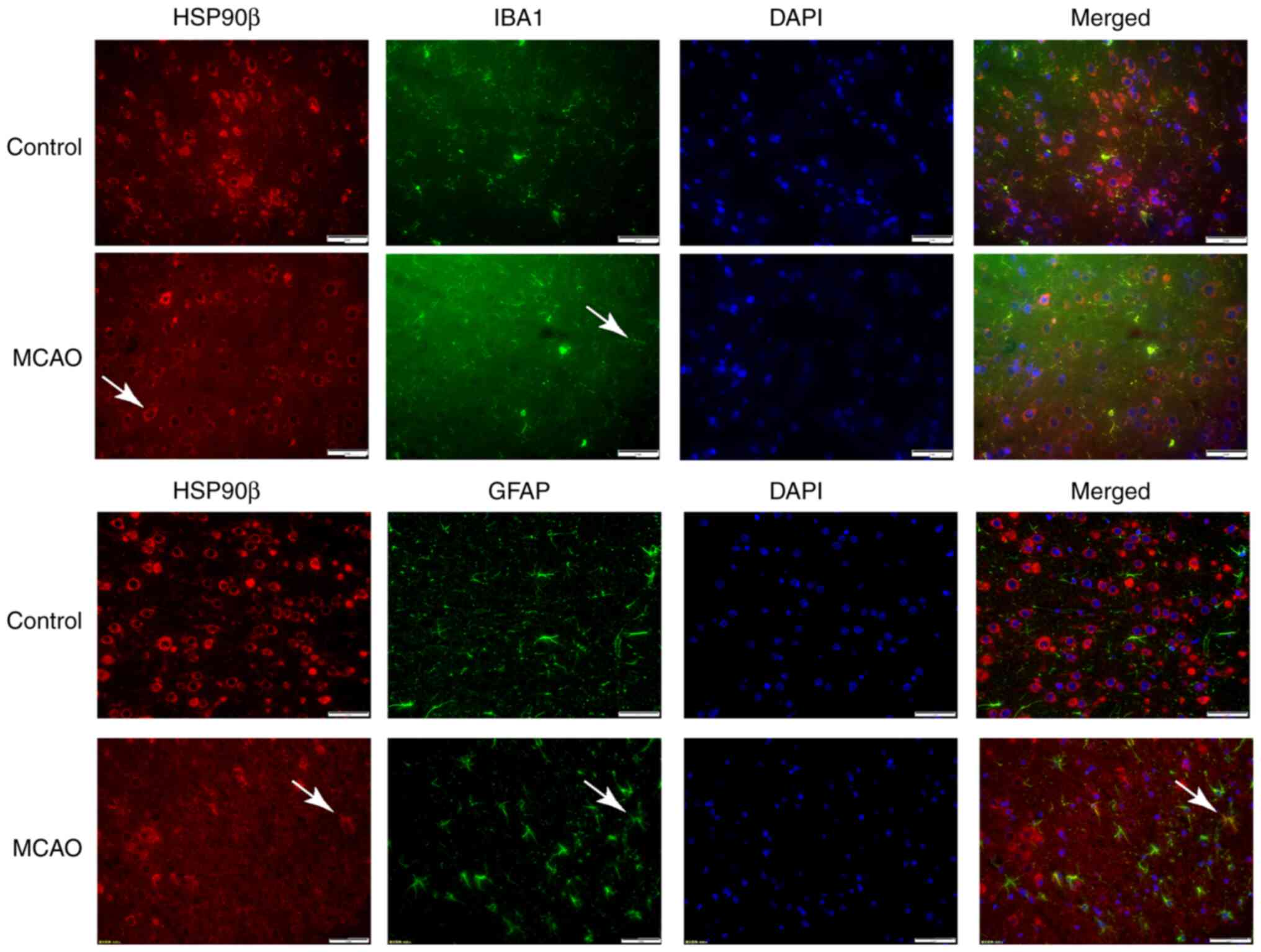 | Figure 2.Immunofluorescence detection of the
expression of HSP90β in the ischemic penumbra of MCAO rats. Arrows
indicate HSP90β-positive cells, GFAP-positive cells, IBA1-positive
cells and DAPI-positive cells; magnification, ×400, n=6 in each
group. HSP, heat shock protein; MCAO, middle cerebral artery
occlusion; GFAP, glial fibrillary acidic protein IBA1, ionized
calcium-binding adaptor molecule 1; DAPI,
4′,6-diamidino-2-phenylindole. |
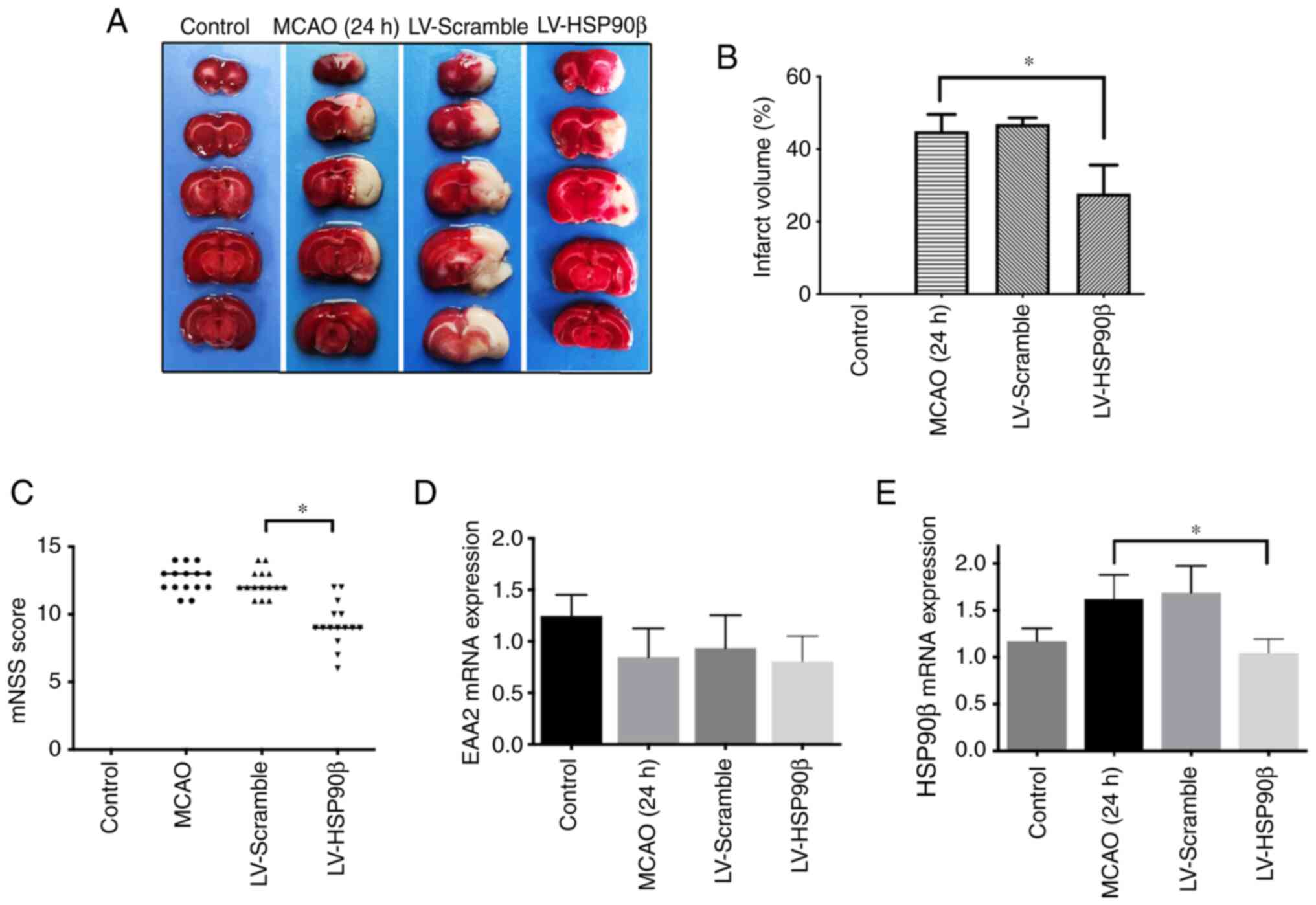
Inhibition of HSP90β expression is
neuroprotective in the rat MCAO model
Rats in each group were evaluated by determining the
mNSS score to judge the degree of neurological deficiency. The
control group and the sham-operated group had no symptoms of
neurological deficiency, while the MCAO group, the empty virus
group and the lentivirus group had different degrees of symptoms of
neurological deficiency. The mNSS score of the lentivirus group was
significantly lower than that of the MCAO group and the empty virus
group (Fig. 3C; P<0.05). TTC
staining was performed to analyze the volume of cerebral infarction
in each group. Except for the control group with no infarction, the
MCAO group, the empty virus group and the HSP90β lentivirus group
all had different degrees of infarction. After inhibiting HSP90β
expression, the infarct volume ratio was significantly reduced,
which was significantly different from the MCAO group and the empty
virus group (Fig. 3A and B).
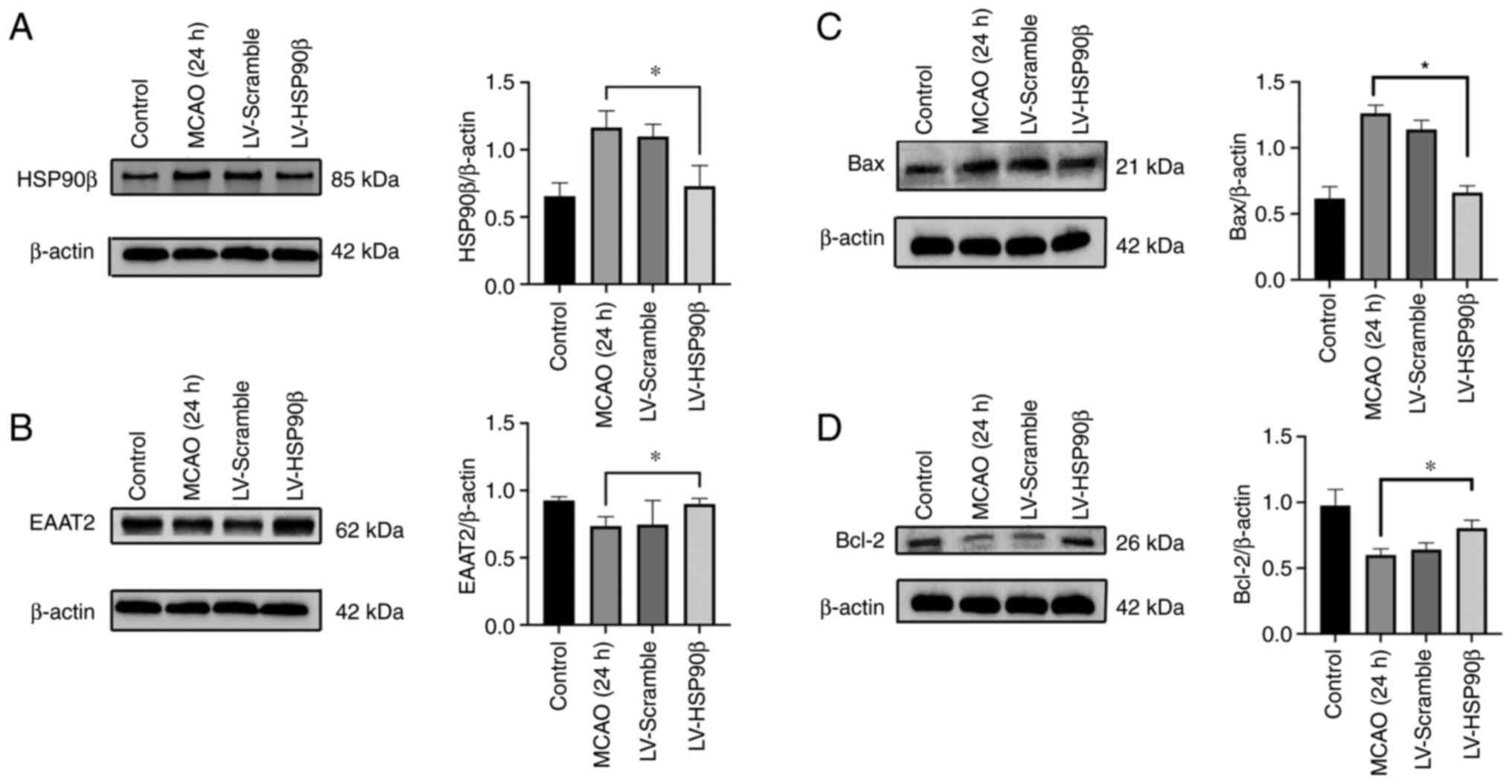
Inhibition of HSP90β expression
increases the EAAT2 protein expression level but not the EAAT2 mRNA
expression level
After inhibiting the expression of HSP90β, EAAT2
expression was detected using western blotting. The EAAT2
expression level was significantly increased and the HSP90β
expression level was significantly decreased (Fig. 4A and B). Additionally, the RT-PCR
results showed a decreased HSP90β mRNA expression level, but the
expression of EAAT2 mRNA was not affected (Fig. 3D and E). The expression of Bax in
the virus intervention group was significantly lower than that in
the MCAO group. The expression of Bcl-2 in the virus intervention
group was significantly higher than that in the MCAO group
(Fig. 4C and D; P<0.05). Based
on these results, inhibition of HSP90β exerted neuroprotective
effects.
Discussion
Stroke is one of the most common diseases in
neurology and is characterized by high morbidity, disability and
mortality rates and is the first cause of death and disability
among adults in China, seriously threatening human life and health
and imposing a huge economic burden on patients, their families and
society. Global burden of disease data show that stroke is the
number one cause of years of life lost in China (15,16).
Effective treatments are lacking for ischemic stroke. The use of
thrombolytic drugs and thrombus autolysis can further induce
ischemia-reperfusion injury and aggravate cellular dysfunction and
structural damage in the ischemic core and surrounding penumbra
area, which is the main mechanism by which neurological deficits
are aggravated (17–19). Therefore, reducing
ischemia-reperfusion injury is one of the key links in the early
treatment of ischemic stroke and is a current challenge in the
field of neurology. The MCAO model was introduced to simulate the
pathological process of ischemia-reperfusion injury, Thus, it is
widely used to investigated as an excellent animal model in
research field of ischemic stroke.
The relationship between HSP90 and stroke is
currently poorly studied. It has been shown that the use of the
HSP90 inhibitor 17-allylamino-demethoxygeldanamycin exerts a
neuroprotective effect on cultured neurons with oxygen-glucose
deprivation and the protective mechanism may be to activate the
PI3K/Akt and MAPK cytoprotective pathways, which inhibit the
cellular oxidative stress response and produce protective effects
(20). Rats are effectively
protected against ischemic brain injury after pretreatment with the
antioxidant N-acetylcysteine. The mechanism may be attributed to
the increased stability of HSP90 binding to hypoxia-inducible
factor 1 alpha, which provides neuroprotection through antioxidant
effects (21). The HSP90 inhibitor
geldanamycin exerts a dual inhibitory effect on JNK3 and has potent
neuroprotective effects (22).
Another study found that the HSP90 expression level was elevated
after cerebral infarction and that the use of inhibitors reduced
the cerebral infarction volume in mice; the mechanism may be
related to the inhibition of the inflammatory NF-κB signaling
pathway (23,24). Although some of these studies
suggested that HSP90 was involved in the process of cerebral
ischemic injury, the difference in the expression of HSP90α and
HSP90β was not clearly indicated. HSP90β serves an essential role
in maintaining cell viability, while HSP90α may mainly serve a role
in ensuring cellular adaptability (10,25),
suggesting the existence of different physiological functions of
HSP90α and HSP90β. The results of the present study further
confirmed that the increased expression of HSP90 after cerebral
ischemia-reperfusion in rats was mainly attributed to a change in
HSP90β expression, while HSP90α expression did not change
significantly, suggesting that HSP90β is involved in the process of
cerebral ischemia-reperfusion injury. Immunofluorescence staining
showed that HSP90β was not significantly expressed in glial cells,
including microglia and astrocytes, under normal conditions.
However, after ischemia-reperfusion in rat brain, HSP90β expression
overlapped with astrocyte markers in the ischemic penumbra area,
while no expression was detected in microglia, suggesting that the
increased HSP90β was mainly expressed on astrocytes. Astrocytes
serve a major role in glutamate uptake from the surrounding
neuronal synapsis and its posterior recycling into glutamine, which
can then be reused by neurons as a substrate for glutamate
synthesis (26). Microglia
activation is one of the first events that occurs after an insult
such as brain ischemia and some of microglia functions include an
increase in phagocytosis rate, release of anti- or pro-inflammatory
cytokines, proliferation and migration (27). The different role of astrocyte and
microglia in ischemia-reperfusion injury related to HSP90β
expression site selectivity.
The main function of EAAT2 is to transport
extracellular excitatory amino acids inside cells to avoid
excitotoxicity; EAAT2 is mainly expressed on astrocytes (5). Excitatory amino acid release, altered
transporter function, receptor expression and the activation of
downstream cell death signals caused by excitatory amino acids have
become the focus of research on the mechanism of cerebral ischemic
injury. EAAT2 dysfunction plays an important role in excitotoxicity
and its associated neurological damage and dysregulation at the
genetic, epistemic regulatory, transcriptional or translational
levels may lead to EAAT2 dysfunction and ultimately cause neuronal
cell death (28).
Studies have shown that regulating EAAT2 reduces the
extracellular glutamate level, reduces the stimulation of
excitatory receptors on the postsynaptic membrane by glutamate,
reduces excitatory damage and exerts a neuroprotective effect
(29–32). In the MCAO model, HSP90β and EAAT2
were mainly expressed on astrocytes. It was hypothesized that
HSP90β may be related to EAAT2. Studies in the field of epilepsy
have shown that increased HSP90β expression levels promote the
degradation of EAAT2 and EAAT2 degradation is significantly reduced
after its expression is inhibited. This mechanism allows the
function of EAAT2 to be retained and functional EAAT2 transports
extracellular excitatory amino acids, which reduces excitotoxicity
(11,33). It has not yet been determined
whether this change occurs in ischemic stroke. The present study
showed that after inhibiting the expression of HSP90β with a
lentivirus, the mNSS score and the infarct area of the brain tissue
in rats decreased, suggesting that inhibiting HSP90β expression has
a neuroprotective effect. Western blotting and PCR results showed
that inhibiting HSP90β expression increased the expression of EAAT2
protein but not EAAT2 mRNA, indicating that the effect of HSP90β on
EAAT2 may be related to its posttranslational regulatory function,
consistent with the present study Additionally, after inhibiting
the expression of HSP90β, Bax expression was decreased and Bcl-2
expression was increased, indicating that inhibiting the expression
of HSP90β reduced the death of neurons. Thus, inhibition of HSP90β
exerted neuroprotective effects; HSP90β has the potential to be a
molecular target of neuroprotection and inhibition of HSP90β may
alleviate cerebral ischemia-reperfusion injury. However, the
present study also has some limitations. On the one hand, it did
not conduct cell-based experiments to verify that HSP90β promoted
EAAT2 degradation or determine the degradation pathway. Sha et
al (32) verified that HSP90β
recruits EAAT2 into the 20S proteasome in human embryonic kidney
cells, thereby promoting EAAT2 degradation; thus, this part of the
work was not repeated. As the existing inhibitors do not have good
selectivity for HSP90α and HSP90β, only a lentivirus was used to
inhibit the expression of HSP90β to obtain the results, which is
different from clinical applications. It is planned to perform all
of these related experiments in the future.
An improved understanding of the exact mechanism of
HSP90β involvement in cerebral ischemia-reperfusion injury is
important for stroke treatment. It is to be hoped that a novel drug
in the field of neuroscience that targets HSP90β will soon be
available to provide a new approach for ischemic stroke
treatment.
Supplementary Material
Supporting Data
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by Guizhou Provincial Health
Commission Science and Technology Fund (grant nos. gzwkj2021-017
and gzwkj2023-005) and Guizhou Administration of Traditional
Chinese Medicine (grant no. QZYY-2021-006).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
PX, JZh and ZX conceived and designed the study. TL,
XH, JZe and LZ performed the experiments. TL and ZZ performed
statistical analysis. TL and PX wrote the manuscript. JZe, ZX and
PX reviewed and edited the manuscript. TL and PX confirm the
authenticity of all the raw data. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
All procedures were approved by the Animal Care and
Use Committee of Zunyi Medical University, Zunyi, China (approval
number ZMU21-2203-487).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wang W, Jiang B, Sun H, Ru X, Sun D, Wang
L, Wang L, Jiang Y, Li Y, Wang Y, et al: Prevalence, incidence, and
mortality of stroke in China: Results from a nationwide
population-based survey of 480 687 adults. Circulation.
135:759–771. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang D, Liu J, Liu M, Lu C, Brainin M and
Zhang J: Patterns of stroke between university hospitals and
nonuniversity hospitals in Mainland China: Prospective multicenter
hospital-based registry study. World Neurosurg. 98:258–265. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yuan Y, Tian Y, Jiang H, Cai LY, Song J,
Peng R and Zhang XM: Mechanism of PGC-1α-mediated mitochondrial
biogenesis in cerebral ischemia-reperfusion injury. Front Mol
Neurosci. 16:12249642023. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Uzdensky AB: Apoptosis regulation in the
penumbra after ischemic stroke: Expression of pro- and
antiapoptotic proteins. Apoptosis. 24:687–702. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chao XD, Fei F and Fei Z: The role of
excitatory amino acid transporters in cerebral ischemia. Neurochem
Res. 35:1224–1230. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chi OZ, Hunter C, Liu X and Weiss HR:
Effects of exogenous excitatory amino acid neurotransmitters on
blood-brain barrier disruption in focal cerebral ischemia.
Neurochem Res. 34:1249–1254. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Takahashi K, Foster JB and Lin CL:
Glutamate transporter EAAT2: Regulation, function, and potential as
a therapeutic target for neurological and psychiatric disease. Cell
Mol Life Sci. 72:3489–3506. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sankhala KK, Mita MM, Mita AC and Takimoto
CH: Heat shock proteins: A potential anticancer target. Curr Drug
Targets. 12:2001–2008. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Whitesell L and Lindquist SL: HSP90 and
the chaperoning of cancer. Nat Rev Cancer. 5:761–772. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Grad I, Cederroth CR, Walicki J, Grey C,
Barluenga S, Winssinger N, De Massy B, Nef S and Picard D: The
molecular chaperone Hsp90α is required for meiotic progression of
spermatocytes beyond pachytene in the mouse. PLoS One.
5:e157702010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang R, Liu C, Ji Y, Teng L and Guo Y:
Neuregulin-1β plays a neuroprotective role by inhibiting the Cdk5
signaling pathway after cerebral ischemia-reperfusion injury in
rats. J Mol Neurosci. 66:261–272. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Longa EZ, Weinstein PR, Carlson S and
Cummins R: Reversible middle cerebral artery occlusion without
craniectomy in rats. Stroke. 20:84–91. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yu X, Luo Y, Yang L, Chen P and Duan X:
P-hydroxybenzyl alcohol ameliorates neuronal cerebral
ischemia-reperfusion injury by activating mitochondrial autophagy
through SIRT1. Mol Med Rep. 27:682023. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhou M, Wang H, Zeng X, Yin P, Zhu J, Chen
W, Li X, Wang L, Wang L, Liu Y, et al: Mortality, morbidity, and
risk factors in China and its provinces, 1990–2017: A systematic
analysis for the global burden of disease study 2017. Lancet.
394:1145–1158. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
GBD 2017 Causes of Death Collaborators, .
Global, regional, and national age-sex-specific mortality for 282
causes of death in 195 countries and territories, 1980–2017: A
systematic analysis for the global burden of disease study 2017.
Lancet. 392:1736–1788. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bradley E, Zhao X, Wang R, Brann D,
Bieberich E and Wang G: Low dose Hsp90 inhibitor 17AAG protects
neural progenitor cells from ischemia induced death. J Cell Commun
Signal. 8:353–362. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chinese Society of Neurology and Chinese
Stroke Society, . Chinese guidelines for diagnosis and treatment of
acute ischemic stroke 2018. Chin J Neurol. 9:666–682. 2018.
|
|
18
|
Adelson JD, Barreto GE, Xu L, Kim T, Brott
BK, Ouyang YB, Naserke T, Djurisic M, Xiong X, Shatz CJ and Giffard
RG: Neuroprotection from stroke in the absence of MHCI or PirB.
Neuron. 73:1100–1107. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pundik S, Xu K and Sundararajan S:
Reperfusion brain injury: Focus on cellular bioenergetics.
Neurology. 79 (13 Suppl 1):S44–S51. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang Z, Yan J, Taheri S, Liu KJ and Shi H
and Shi H: Hypoxia-inducible factor 1 contributes to
N-acetylcysteine's protection in stroke. Free Radic Biol Med.
68:8–21. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wen XR, Li C, Zong YY, Yu CZ, Xu J, Han D
and Zhang GY: Dual inhibitory roles of geldanamycin on the c-Jun
NH2-terminal kinase 3 signal pathway through suppressing the
expression of mixed-lineage kinase 3 and attenuating the activation
of apoptosis signal-regulating kinase 1 via facilitating the
activation of Akt in ischemic brain injury. Neuroscience.
156:483–497. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Qi J, Liu Y, Yang P, Chen T, Liu XZ, Yin
Y, Zhang J and Wang F: Heat shock protein 90 inhibition by
17-dimethylaminoethylamino-17-demethoxygeldanamycin protects
blood-brain barrier integrity in cerebral ischemic stroke. Am J
Transl Res. 7:1826–1837. 2015.PubMed/NCBI
|
|
23
|
Qi J, Han X, Liu HT, Chen T, Zhang JL,
Yang P, Bo SH, Lu XT and Zhang J:
17-Dimethylaminoethylamino-17-demethoxygel-danamycin attenuates
inflammatory responses in experimental stroke. Biol Pharm Bull.
37:1713–1718. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Voss AK, Thomas T and Gruss P: Mice
lacking HSP90beta fail to develop a placental labyrinth.
Development. 127:1–11. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pajarillo E, Rizor A, Lee J, Aschner M and
Lee E: The role of astrocytic glutamate transporters GLT-1 and
GLAST in neurological disorders: Potential targets for
neurotherapeutics. Neuropharmacology. 161:1075592019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mahmoud S, Gharagozloo M, Simard C and
Gris D: Astrocytes maintain glutamate homeostasis in the CNS by
controlling the balance between glutamate uptake and release.
Cells. 8:1842019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang H, Li J, Zhang H, Wang M, Xiao L,
Wang Y and Cheng Q: Regulation of microglia polarization after
cerebral ischemia. Front Cell Neurosci. 17:11826212023. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Leung SW, Lai JH, Wu JC, Tsai YR, Chen YH,
Kang SJ, Chiang YH, Chang CF and Chen KY: Neuroprotective effects
of emodin against ischemia/reperfusion injury through activating
ERK-1/2 signaling pathway. Int J Mol Sci. 21:28992020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Peng M, Ling X, Song R, Gao X, Liang Z,
Fang F and Cang J: Upregulation of GLT-1 via PI3K/Akt pathway
contributes to neuroprotection induced by dexmedetomidine. Front
Neurol. 10:10412019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bacigaluppi M, Russo GL,
Peruzzotti-Jametti L, Rossi S, Sandrone S, Butti E, De Ceglia R,
Bergamaschi A, Motta C, Gallizioli M, et al: Neural stem cell
transplantation induces stroke recovery by upregulating glutamate
transporter GLT-1 in astrocytes. J Neurosci. 36:10529–10544. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Krzyżanowska W, Pomierny B, Bystrowska B,
Pomierny-Chamioło L, Filip M, Budziszewska B and Pera J:
Ceftriaxone- and N-acetylcysteine-induced brain tolerance to
ischemia: Influence on glutamate levels in focal cerebral ischemia.
PLoS One. 12:e01862432017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sha L, Wang X, Li J, Shi X, Wu L, Shen Y
and Xu Q: Pharmacologic inhibition of Hsp90 to prevent GLT-1
degradation as an effective therapy for epilepsy. J Exp Med.
214:547–563. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Peng YC, Wang S, Zhang Y, Huang LJ, Wang
XL and Peng Y: Hsp90β inhibitors prevent GLT-1 degradation but have
no beneficial efficacy on absence epilepsy. J Asian Nat Prod Res.
21:905–915. 2019. View Article : Google Scholar : PubMed/NCBI
|