Introduction
Polycystic ovary syndrome (PCOS) is the most common
and complex endocrine disorder, affecting up to 10% of women, and
may present with an irregular menstrual cycle (oligo-ovulation),
hyperandrogenism, hirsutism and polycystic ovaries (1,2).
Long-term risks conferred by PCOS include metabolic disorders such
as type 2 diabetes and cardiovascular disease (3). Additionally, PCOS has been confirmed
to be related to the quality of oocytes (4).
Cells with the granulosa support oocyte development.
There are two types of granulosa cells: Granulosa cells distributed
in and around the follicle wall (mural granulosa cells, mGCs) and
granulosa cells in the follicle wall (cumulus granulosa cells,
cGCs). Oocytes are enclosed within cGCs, providing necessary growth
factors, hormones and other nutrients for oocytes (5).
Mitochondria not only supply the energy needed for
the proliferation and metabolism of cGCs, but also regulate
important physiological processes including antioxidation,
apoptosis and autophagy. Mitochondria are the main source of
reactive oxygen species (ROS). Through the production of ROS,
mitochondria are the major determinant of numerous factors involved
in reproduction such as oocyte quality, follicular growth and
development, and granulosa cell proliferation (6).
Silent information regulator 1 (SIRT1), a
deacetylase enzyme that requires NAD+, regulates
mitochondrial biogenesis, oxidative stress defense and energy
homeostasis during folliculogenesis (7,8).
Moreover, SIRT1 deacetylates peroxisome proliferator-activated
receptor-γ co-activator 1α (PGC1α) and is involved in mitochondrial
biogenesis and activity. Among the proteins that mediate the
SIRT1-dependent action on mitochondria, a key role is played by
peroxisome proliferator-activated receptor PGC1α (9,10),
along with mitochondrial transcriptional factor A (TFAM), which is
a downstream target gene of SIRT1/PGC1α (11,12).
SIRT1 can deacetylate PGC1-α through the AMP-activated protein
kinase (AMPK) pathway to affect the function and quality of the
mitochondria of ovarian GCs (13).
However, there have been no studies on the impact of mitochondrial
biogenesis in granulosa cells of PCOS patients, and it is unknown
whether mitochondrial dysfunction in cGCs leads to a decrease in
oocyte quality in PCOS patients. As a result of the present study,
it was investigated whether regulation of a SIRT1/AMPK axis in
ovaries may be associated with mitochondrial biogenesis in patients
with PCOS.
Materials and methods
Patients
The participants were recruited from women who
attended the Reproductive Medicine Center of Hebei Reproductive
Health Hospital (Shijiazhuang, China) between June 2019 and June
2022. The diagnosis of PCOS was based on the Rotterdam criteria.
Patients who were included in the study presented at least two of
the following conditions: i) oligo-and/or anovulation; ii) androgen
excess; iii) polycystic ovaries. Patients with an ovarian cyst,
diminished ovarian reserve, hydrosalpinx, endometriosis, chromosome
abnormality, or other systemic diseases which could affect
granulosa cell gene expression were excluded. Infertile patients
who have common ovulation with tubal issues were recruited as
controls. The final study group included 66 PCOS group and 63
control (CON) group. The patient information statistics are shown
in Table I.
 | Table I.Comparison of general condition
between two groups (mean ± SD). |
Table I.
Comparison of general condition
between two groups (mean ± SD).
| General
condition | CON group
(n=66) | PCOS group
(n=63) | P-value |
|---|
| Age, years | 30.90±3.81 | 31.14±3.23 | 0.728 |
| BMI,
kg/m2 | 23.44±2.60 | 24.79±3.95 | 0.042 |
| Basal FSH,
IU/l | 7.27±2.13 | 7.38±3.19 | 0.826 |
| Basal LH,
IU/l, | 4.61±3.29 | 5.64±3.50 | 0.109 |
| FSH/LH | 2.02±1.14 | 1.70±0.96 | 0.114 |
| Basal E2,
pg/ml | 38.07±17.38 | 38.42±11.27 | 0.904 |
| Basal P, ng/ml | 0.70±0.60 | 0.62±0.50 | 0.413 |
| Basal PRL,
ng/ml | 15.46±11.41 | 12.53±5.85 | 0.109 |
| Basal T, ng/ml | 0.36±0.15 | 0.46±0.21 | 0.005 |
Ethical approval
The present study was approved by the Ethics
Committee of the Hebei Reproductive Health Hospital (approval no.
KYY-2021-LW-005; Shijiazhuang, China). Written informed consent was
obtained from each patient. The research followed the Helsinki
Declaration standards (1964).
Isolation of GCs
The oocyte-cumulus complexes (OCCs) were retrieved
36 h after human chorionic gonadotropin injection and washed with
phosphate-buffered saline (PBS) at 4°C to remove any remaining
mGCs, blood cells, or debris. The cumulus complex was selected
under a light microscope, and the cGCs around the oocyte were
carefully removed mechanically.
Transmission electron microscopy
(TEM)
The cGCs were collected and centrifuged at 400 × g
for 5 min at room temperature. Cells were obtained in 2.5%
glutaraldehyde solution at 4°C for 30 min and were then dehydrated
with a gradient of ethyl alcohol series, embedded in Araldite, then
sliced into 60-nm sections. Uranyl acetate and lead citrate were
used to stain the sections at room temperature for 15 min, after
which, the sections were examined under a transmission electron
microscope (Hitachi, Ltd.) and images were captured. Overall, the
TEM experiment involved 3 patients with CON and 3 patients with
PCOS.
Detection of intracellular ROS
levels
The 2,7-dichlorodihydrofluorescein diacetate
(DCFH-DA) fluorescent probe (Cayman Chemical Company) was employed
to detect the intracellular ROS levels. Briefly, cGCs were
resuspended using 10 µmol/l DCFH-DA, incubated for 30 min at 37°C
in the dark, and then washed thrice with PBS buffer (cat. no.
02-024-1ACS; Biological Industries; Sartorius AG). cGCs were
resuspended using 10 µg/ml Hoechst 3342 (Cayman Chemical Company)
and incubated for 10 min in the dark at 37°C, and then washed
thrice with PBS buffer. The cell fluorescence intensity was
detected with an ImageXpress confocal microscope (Molecular
Devices, LLC), and the ROS levels were expressed as the mean green
fluorescence intensity. This experiment involved 7 patients with
CON and 5 patients with PCOS.
Mito-Tracker Red CMXRos staining
cGCs were cultured with 10 µm/l Mito-Tracker Red
CMXRos staining solution for 30 min at 37°C in 5% CO2.
After washing thrice with PBS buffer. Then, cGCs were stained using
10 µg/ml Hoechst 3342, incubated for 10 min at 37°C in 5%
CO2, and then washed thrice with PBS buffer (Seville)
before placing in fresh medium. The fluorescence intensity of
mitochondrial Mito-Tracker Red CMXRos was detected immediately with
an ImageXpress confocal microscope (Molecular Devices, LLC). The
captured images were assessed, using ImageJ Imaging System software
(version 1.51; National Institutes of Health). Mito-Tracker Red
levels were assessed by calculating the mean red fluorescence
intensity. This experiment involved 3 patients with CON and 4
patients with PCOS.
Measurement of cellular ATP
levels
With the ATP Assay kit (cat. no. A095-1-1; Nanjing
Jiancheng Bioengineering Institute), it was possible to measure
cellular ATP levels. The ATP value of cGCs was assessed by
measuring the absorbance at a wavelength of 636 nm using a
microplate reader (Spectra Max iD3; Molecular Devices, LLC). To
avoid errors caused by differences in protein content, the
bicinchoninic acid (BCA) protein Assay kit (Beyotime Institute of
Biotechnology) was used to measure sample protein concentrations.
This experiment involved 11 patients with CON and 10 patients with
PCOS.
Mitochondrial membrane potential (MMP)
assay-JC-1
The MMP of cGCs was measured using the fluorescent
dye
5,6-dichloro-2-[3[(5,6-dichloro-1,3-diethyl-1,3-dihydro-2H-benzimidazole-2-ylidene)-1-propen-1-yl]-1,3-diethyl-1H
benzim-idazolium, monoiodide (JC-1; Cayman Chemical Company). cGCs
were collected and labeled with 10 µmol/l JC-1 fluorescent probe in
5% CO2 at 37°C for 30 min. Then were washed thrice with
PBS buffer. The aforementioned images were captured using
ImageXpress confocal microscope (Molecular Devices, LLC) to
demonstrate green fluorescence and red fluorescence. Then, captured
images were further analyzed using ImageJ Imaging System software
(version 1.51). The fluorescence was expressed as a ratio of red to
green. This experiment involved 4 patients with CON and 3 patients
with PCOS.
Reverse-transcription-quantitative PCR
(RT-qPCR)
Total RNA was purified with an RNA extract kit (cat.
no. TR254-2; Genstone Biotech) according to the manufacturer's
instructions, and cDNA was synthesized using the
Supersmart™ 6 min 1st Strand cDNA Synthesis kit (cat.
no. ZS-M14003M-50T; ZHONGSHI TONTRU) according to the
manufacturer's instructions. For qPCR, each 20 µl reaction system
contained 2 µl cDNA and 10 µl 2X SYBR® Green PCR Master
Mix (cat. no. RK21203; ABclonal Biotech Co., Ltd.), forward primer
0.4 µl, reverse primer 0.4 µl and ddH2O 7.2 µl. The qPCR
was carried out with the following conditions: Initial denaturation
at 95 °C for 3 min followed by 40 cycles at 95 °C for 5 sec and 60
°C for 30 sec for annealing, and extension at 72°C for 1 min. Each
sample was analyzed in triplicate, and the relative fold expression
of each gene was calculated using the comparative critical
threshold (Cq) method based on the 2-∆ΔCq calculation
(14), relative to the housekeeper
gene β-actin. Primer sequences of the related genes analyzed are
described in Table SI. This
experiment involved 11 patients with CON and 11 patients with
PCOS.
Western blotting
cGCs were homogenized in 50–100 µl RIPA buffer (50
µM Tris/HCl, pH 7.4; 150 µM NaCl; 1% NP-40; 0.5% deoxycholate; 0.1%
sodium dodecyl sulfate; 1% protease inhibitor cocktail; cat. no.
RW0001; Report Biotech). After centrifugation at 12,000 × g at 4°C
for 10 min, the supernatant was collected and the protein
concentration was determined using a BCA kit. Protein samples (5–8
µg) were separated by 12% SDS-PAGE at 120 V for 60 min.
Subsequently, the separated proteins were transferred onto a
polyvinylidene difluoride membrane (Sigma-Adrich; Merck KGaA) at
400 mA for 2 h (Bio-Rad Laboratories, Inc.). Afterward, a blocking
buffer containing 5% non-fat dry milk (Inner Mongolia Yili
Industrial Group Company Ltd.) was provided for 45 min to block the
PVDF membrane. The Membrane was then incubated with primary
antibodies for immunoblotting against SIRT1 (1:1,000; cat. no.
ab110304; Abcam), AMPK (1:1,000; cat. no. AF6423; Affinity
Biosciences, Ltd.), phosphorylated (P-)AMPK (1:1,000; cat. no.
JJ08-19; Huabio), PGC-1α (1:500; cat. no. sc-518038; Santa Cruz
Biotechnology, Inc.), optic nerve atrophy 1 (OPA1) (1:500; cat. no.
ET1705-9; Huabio), dynamin-related protein 1 (DRP1) (1:1,000; cat.
no. ab199722; Abcam), mitofusin 2 (MFN2) (1:1,000; cat. no. 9482S;
Cell Signaling Technology, Inc.), mitochondrial fission 1 (FIS1)
(1:500; cat. no. ET7109-17; Huabio) and β-actin (1:5,000; cat. no.
AC026; ABclonal Biotech Co., Ltd.) overnight at 4°C. Following the
primary incubation, the membrane was incubated with the HRP-goat
anti-rabbit IgG (H + L) (1:3,000; cat. no. RS0002; Immunoway) or
the HRP-goat anti-mouse IgG (H + L) (1:3,000; cat. no. S1001;
Report Biotech) for 1.5 h at room temperature. The protein bands
were detected using a Minichemi 320 chemiluminescence imaging
system (Beijing sage creation) and quantified with ImageJ 1.8.0
software. This experiment involved 27 patients with CON and 27
patients with PCOS.
Statistical analysis
Descriptive analyses were performed for the
variables studied. The results are presented as the mean ± SD for
continuous variables and absolute numbers and relative frequencies
for categorical variables. Comparisons of unpaired continuous data
were conducted using the unpaired Student's t-test or Mann-Whitney
U test, depending on how the data were distributed. Comparisons of
paired continuous data were conducted using the paired t-test or
Mann-Whitney U test, depending on the data distribution. the
chi-square test was used to analyze qualitative variables. Analysis
was conducted using SPSS 20.0 software (SPSS, Inc.; IBM Corp.).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Comparison of the general conditions
between two groups
In the present study, a total of 66 patients with
PCOS and 63 control patients were included. A comparative study of
clinical characteristics between PCOS patients and controls is
presented in Table I. Age
(30.90±3.81 vs. 31.14±3.23 years; P=0.728), basal follicular
stimulating hormone (FSH) level (7.27±2.13 vs. 7.38±3.19; P=0.826),
basal luteinizing hormone (LH) level (4.61±3.29 vs. 5.64±3.50;
P=0.109), FSH/LH ratio (2.02±1.14 vs. 1.70±0.96; P=0.114), basal
estradiol (E2) level (38.07±17.38 vs. 38.42±11.27; P=0.904), basal
progesterone (P) level (0.70±0.60 vs. 0.62±0.50; P=0.413) and basal
prolactin level (15.46±11.41 vs. 12.53±5.85, P=0.109) had no
significant difference between these two groups, while BMI
(23.44±2.60 vs. 24.79±3.95; P=0.042) and basal testosterone (T)
(0.36±0.15 vs. 0.46±0.21; P=0.005) were increased in the PCOS
group.
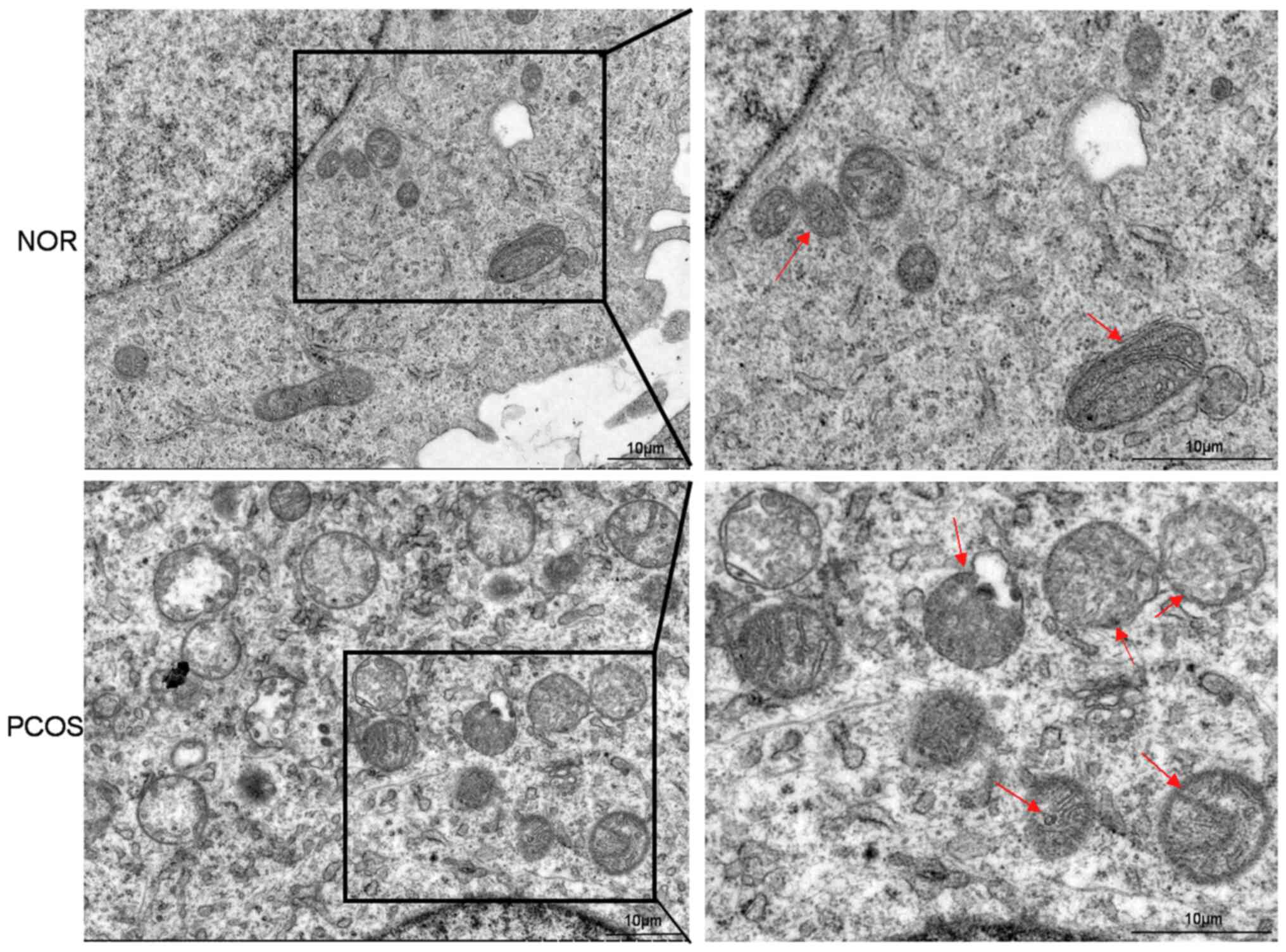
Injured mitochondria are observed in
the GCs of the patients with PCOS
To investigate the changes in mitochondrial
ultrastructure, TEM was used. In the control group, all the cGCs
had a normal nucleus with intact and continuous nuclear membranes,
and most round or rod-shaped mitochondria with a clear crista
around the nucleus. However, the cGCs of patients with PCOS
presented mitochondrial exhibited edema, reduced crista and
fracture, and mitochondrial vacuoles. It was suggested that the
mitochondrial structure of GCs in the ovary of PCOS patients was
abnormal and seriously damaged (Fig.
1).
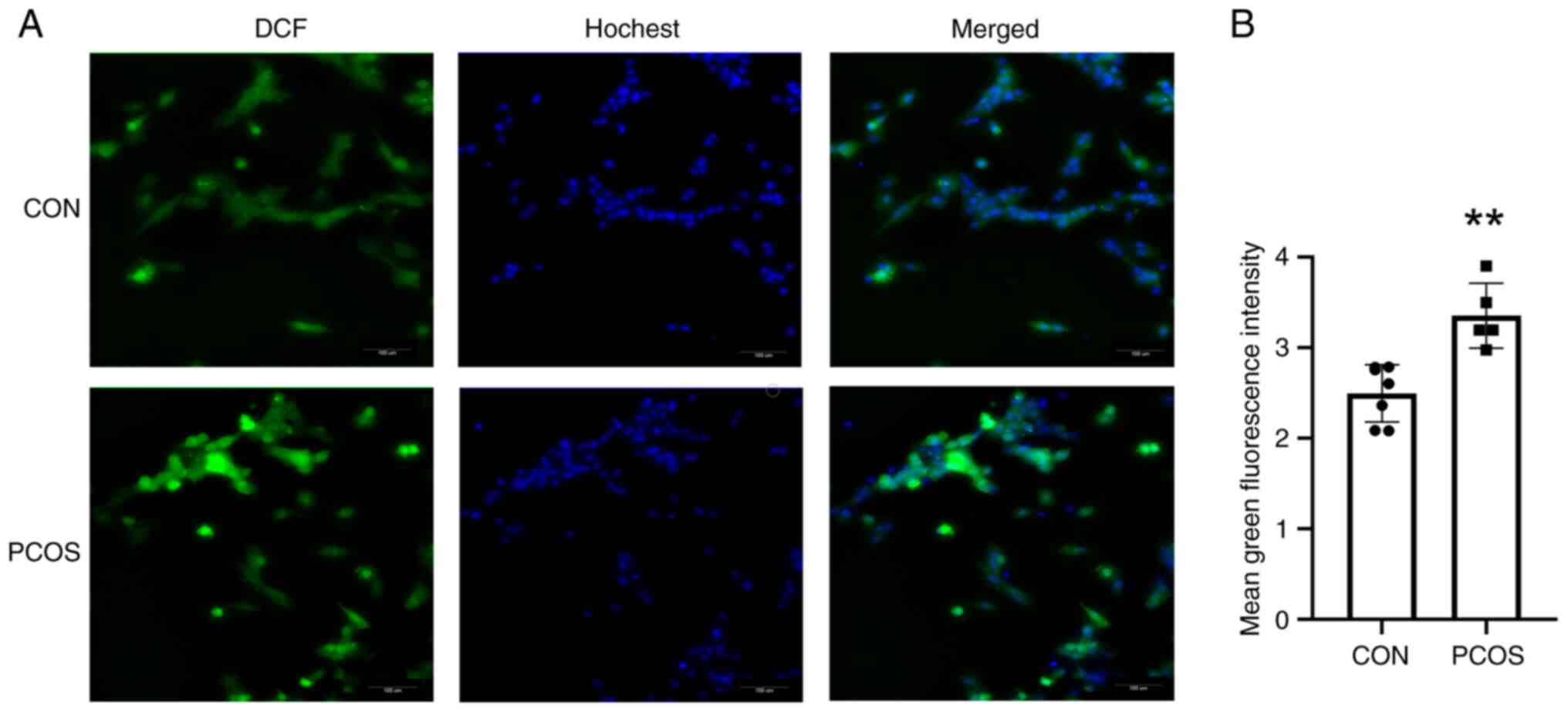
Detection of intracellular ROS
levels
When mitochondria produce excessive ROS, GCs become
dysfunctional and apoptotic. After incubating the probe, the cGCs
in the CON group showed weak fluorescence, while those in the PCOS
group demonstrated clearer fluorescence (Fig. 2A). As revealed in the bar graph,
mean green fluorescence intensity for cGCs from the patients of the
CON group was 2.51±0.46, which was significantly lower than that in
patients of the PCOS group (3.35±0.36) (P=0.001) (Fig. 2B).
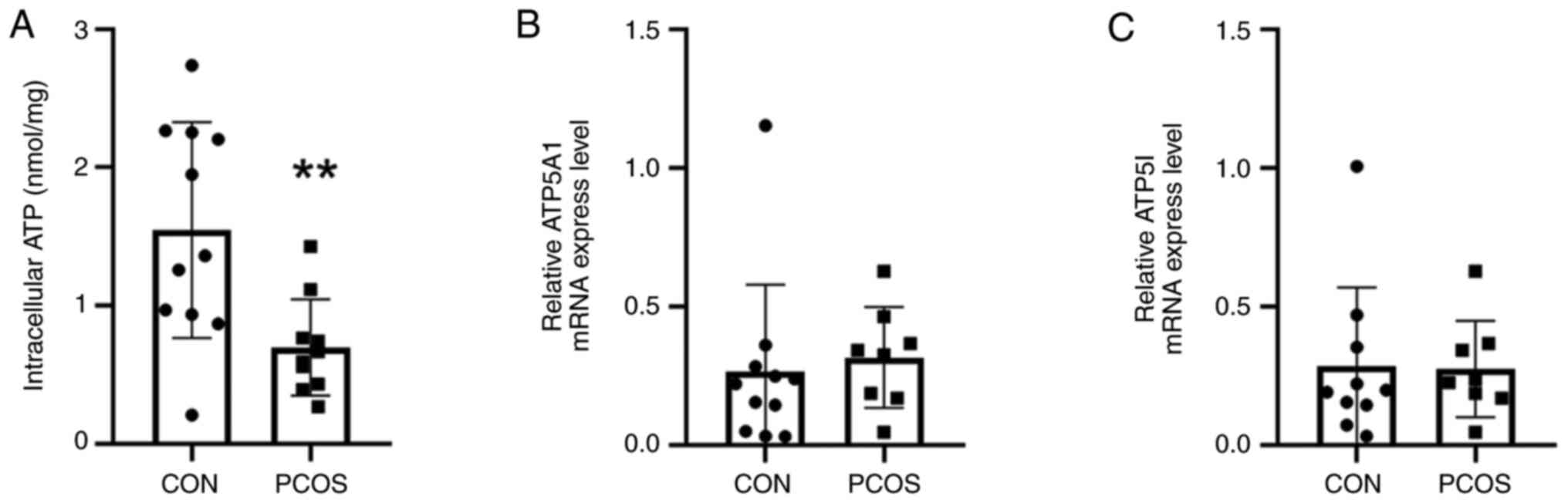
The ATP content of cGCs in patients
with PCOS is reduced
According to the analyses of the ATP bioluminescence
detection kit, the ATP content (0.70±0.35 mm) of cGCs in the PCOS
group was significantly reduced compared with the control group
(1.55±0.78 mm; P=0.006) (Fig. 3A).
To investigate the expression of ATP synthesizing genes, a RT-qPCR
was conducted. The mRNA transcription levels of ATP5A1 and ATP5I
were not significantly different between the two groups (P>0.05)
(Fig. 3B and C).
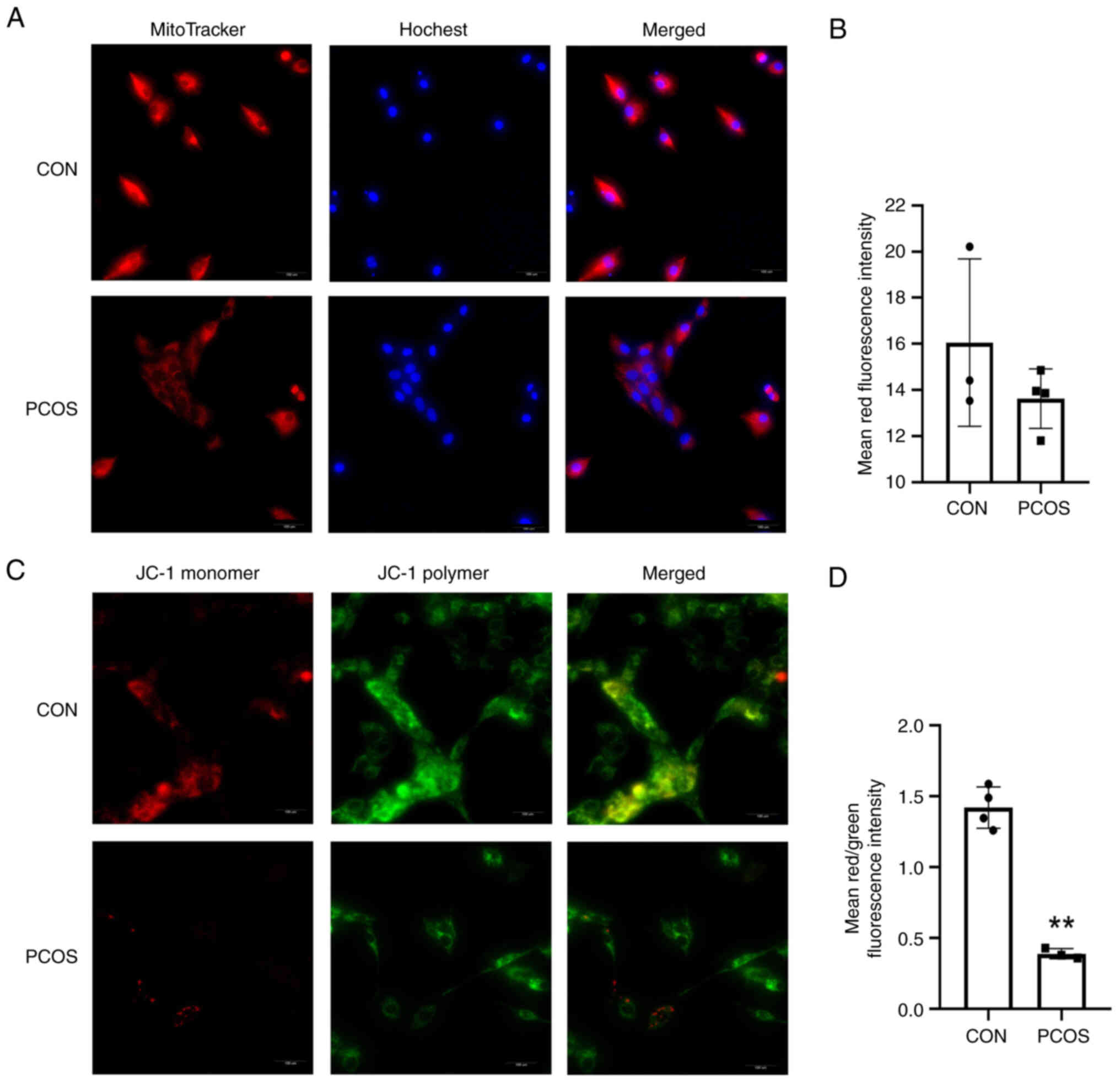
The numbers of mitochondria of cGCs in
patients with PCOS are reduced
Mito-Tracker Red CMXRos staining revealed the
presence of bioactive mitochondria with a stronger red-light
intensity, and compared with the CON group, the cells in the PCOS
group revealed weaker fluorescence (Fig. 4A). As shown in the bar graph, the
mean red fluorescence intensity of the cGCs from the patients of
the PCOS group was decreased, but no statistical significance was
observed (16.05±3.63 vs. 13.62±1.29; P=0.259) (Fig. 4B).
Increased mitochondrial membrane
damage in patients with PCOS
MMP was also monitored; the JC-1 staining reveals a
higher MMP when red fluorescence is present, and a lower MMP when
green fluorescence is present. A lower MMP was observed in GCs of
women with PCOS as compared with controls (Fig. 4C and D). The results showed that
the mean red/green fluorescence intensity ratio for cGCs from the
patients of the CON group was 1.47±0.12, which was significantly
higher than that (0.37±0.04) of patients of the PCOS group
(P=0.002) (Fig. 4D). These
observations indicated that mitochondrial dysfunction in GCs
appears to be accompanied by PCOS.
The mitochondrial dynamics are
unbalanced in cGCs of the patient with PCOS
Due to mitochondrial dysfunction, ROS are produced,
which in turn exacerbate mitochondrial damage. Therefore,
mitochondrial function was further investigated. It was found that
the protein levels of mitochondrial fusion-related genes MFN2
(1.49±0.81 vs. 2.69±1.69; P=0.002; Fig. 5A and E) and genes associated with
mitochondrial fission FIS1 (1.93±0.87 vs. 3.47±2.20; P=0.009;
Fig. 5A and C) were increased in
the PCOS group compared with the CON group. No statistical
significance were observed in the expression level of OPA1 and DRP1
between the PCOS and the CON groups (Fig. 5A, B and D). Repeated experiments
were conducted for numerous patients and the results revealed the
same trend. Results of repeated western blot experiments regarding
the proteins DRP1, FIS1, OPA1, MFN2 are included in Fig. S1, Fig. S2, Fig. S3, Fig. S4, respectively. The mRNA
transcription levels of genes including Opa1 (0.00081±0.00053 vs.
0.00064±0.00054; P=0.560; Fig.
5F), DRP1 (0.014±0.016 vs. 0.021±0.020; P=0.503; Fig. 5G), FIS1 (0.042±0.0080 vs.
0.070±0.049; P=0.002; Fig. 5H) and
MFN2 (0.00031±0.00035 vs. 0.0030±0.0042; P=0.188; Fig. 5I), did not demonstrate a
statistically significant difference between the two groups.
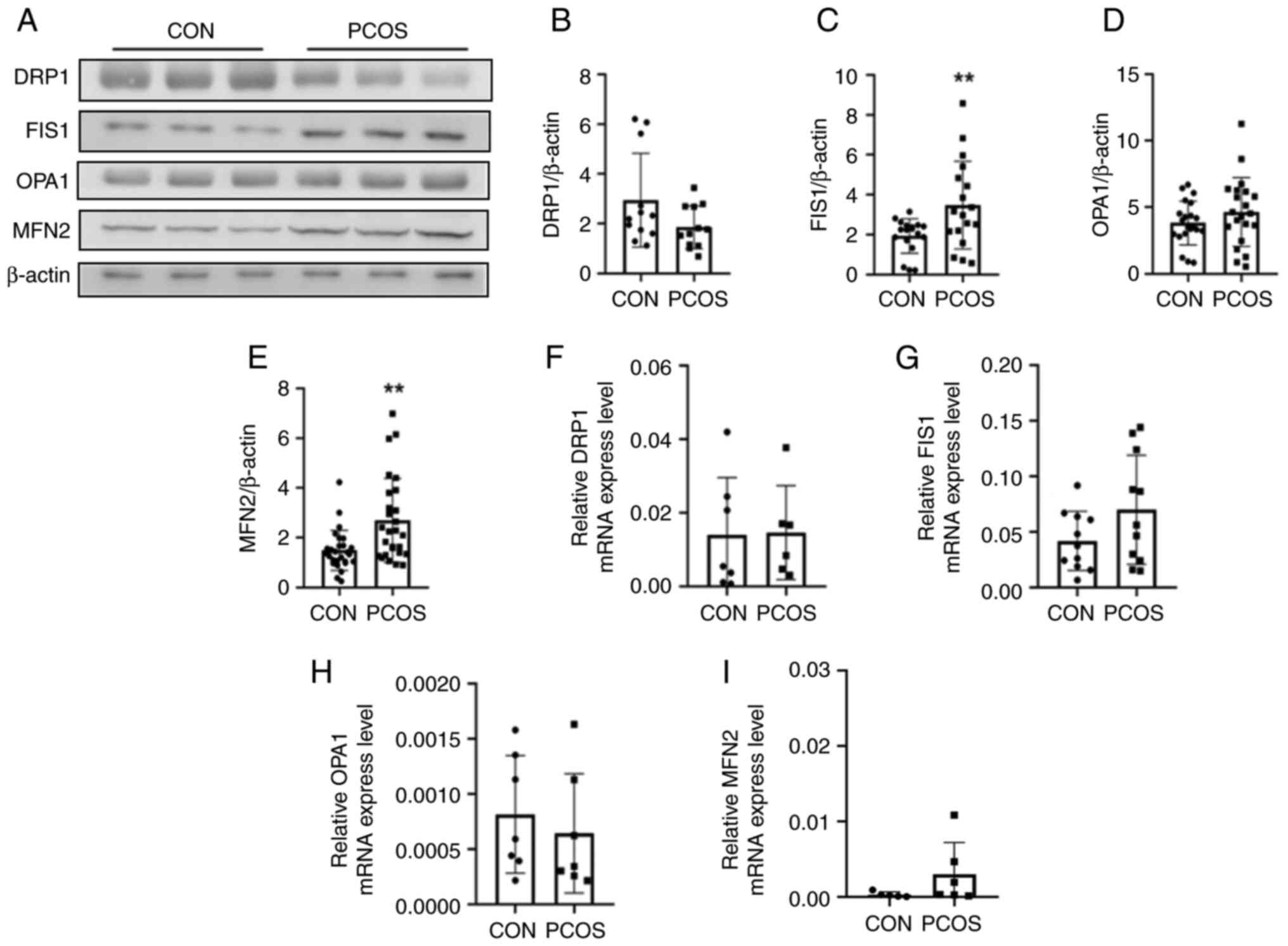 | Figure 5.Western blot analysis and RT-qPCR
comparison of DRP1, FIS1, OPA1 and MFN2 between the CON and PCOS
groups. (A) Mitochondrial protein expression, including DRP1, FIS1,
OPA1 and MFN2 was detected in the two groups by western blotting.
(B) Gray intensity of DRP1 expression relative to β-actin (n=3 vs.
n=3). No statistical significance was observed in the expression
levels of DRP1 between the PCOS and the CON groups. (C) The gray
intensity of FIS1 expression relative to β-actin (n=3 vs. n=3). The
protein expression of FIS1 in the cGCs of the PCOS group was
significantly increased compared with the CON group. (D) The gray
intensity of OPA1 expression relative to β-actin (n=3 vs. n=3). (E)
The gray intensity of MFN2 expression relative to β-actin (n=3 vs.
n=3). The protein expression of MFN2 in the cGCs of the PCOS group
was significantly increased compared with the CON group. (F) The
bar chart shows the mRNA transcription of DRP1 in the CON and PCOS
groups by Q-PCR (n=7 vs. n=6). (G) The bar chart shows the mRNA
transcription of FIS1 in the CON and PCOS groups by RT-qPCR (n=11
vs. n=11). (H) The bar graph shows the mRNA transcription of OPA1
in the CON and PCOS groups by RT-qPCR (n=7 vs. n=7). (I) The bar
chart shows the mRNA transcription of MFN2 in the CON and PCOS
groups by RT-qPCR (n=5 vs. n=6). The mRNA transcription levels of
the genes DRP1, FIS1, OPA1, MFN2 did not demonstrate statistically
significant difference between the two groups. **P<0.01.
RT-qPCR, reverse transcription-quantitative PCR; DRP1,
dynamin-related protein 1; FIS1, mitochondrial fission 1; OPA1,
optic nerve atrophy 1; MFN2, mitofusin 2; CON, normal ovarian
reserve; PCOS, polycystic ovary syndrome; cGCs, cumulus granulosa
cells. |
The expression of proteins regulating
mitochondrial biogenesis in patients with PCOS
Given the initial results of the present study, the
molecular regulatory mechanisms of mitochondrial function were then
explored. The mRNA transcription level of the mitochondrial
transcription factor A (TFAM) was detected, which is crucial for
the replication and transcription of mitochondrial DNA-encoded
genes. PGC-1α acts as a transcription coactivator by binding
directly to the transcription factor TFAM in the DNA promoter
region. SIRT1 and P-AMPK affect mitochondrial function by
activating PGC-1α.
The results of the statistical analysis revealed
that the mRNA transcription level of TFAM in the cells of the PCOS
group was significantly lower than that in the cells of the CON
group (0.0090±0.0046 vs. 0.0063±0.0073; P=0.045; Fig. 6H). Moreover, the levels of energy
depletion signaling-related factors proteins in the CON group were
higher than those in the PCOS group, including SIRT1 (2.29±0.43 vs.
0.86±0.20; P<0.0001; Fig. 6A and
B), AMPK (5.78±2.72 vs. 3.69±0.98; P=0.020), P-AMPKα
(10.18±3.97 vs. 2.60±1.53, P=0.006) and PGC-1α (0.77±0.29 vs.
0.34±0.11; P=0.015; Fig. 6A and
D). The protein ratio of P-AMPKα to AMPK in PCOS group was
significantly lower than in the CON group (2.42±1.21 vs. 0.71±0.47;
P=0.015; Fig. 6C). Repeated
experiments were performed in numerous patients, and the results
revealed the same trend. Results of repeated western blot
experiments regarding the proteins SIRT1, AMPK, PGC-1 and P-AMPKα
are included in Fig. S5, Fig. S6, Fig. S7, Fig. S8, respectively. There was no
statistically significant difference in SIRT1 (0.0035±0.0022 vs.
0.0057±0.0030; P=0.159; Fig. 6E),
AMPK (0.0038±0.0027 vs. 0.0045±0.0025; P=0.657; Fig. 6F) and PGC-1α (0.015±0.012 vs.
0.0065±0.0070; P=0.162; Fig. 6G)
mRNA transcription levels.
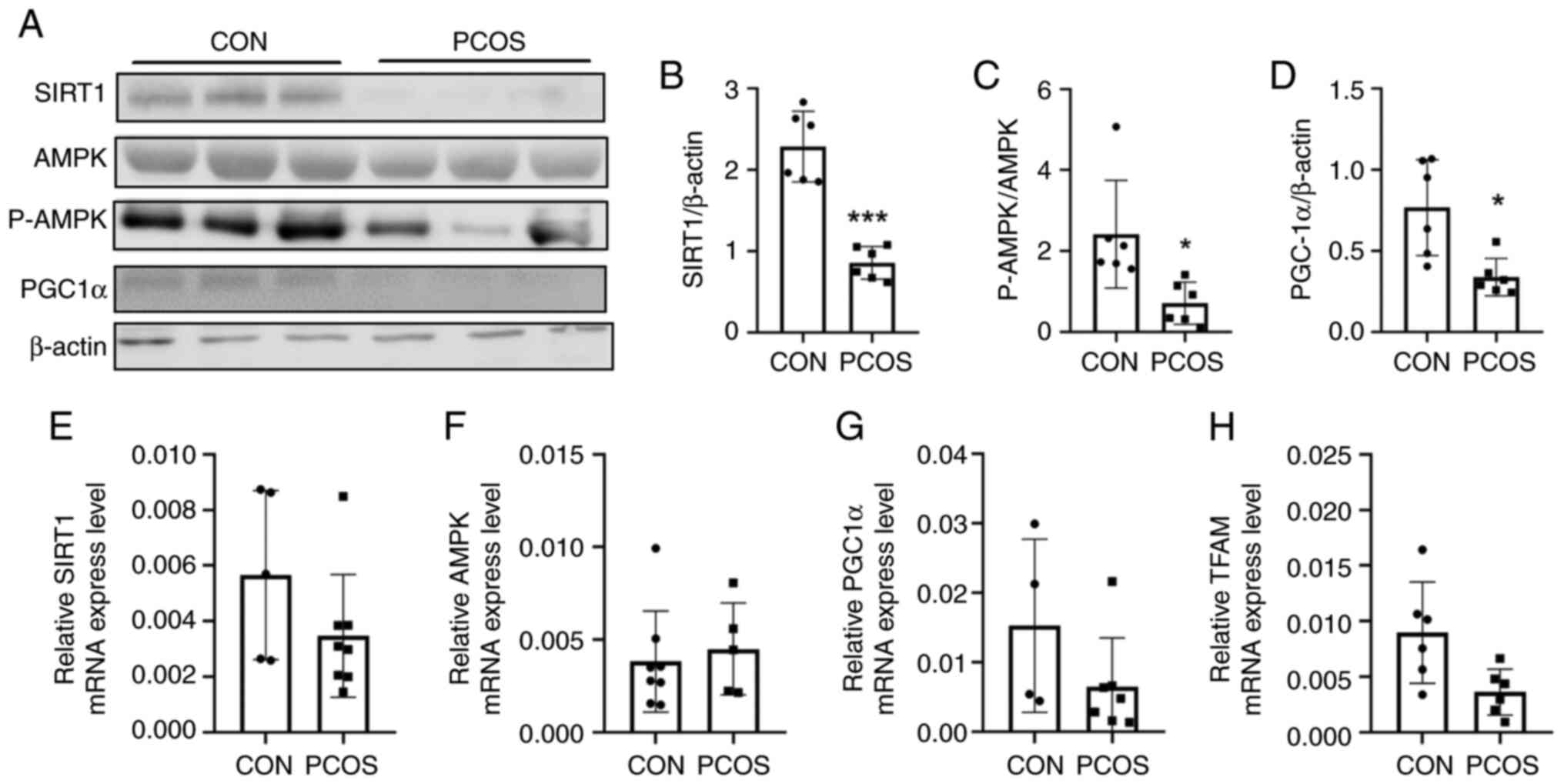 | Figure 6.Western blot analysis and RT-qPCR
comparison of pathway proteins between the CON and PCOS groups. (A)
Mitochondrial protein expression of the pathway protein was
detected by western blotting. (B) The gray intensity of SIRT1
expression relative to β-actin (n=3 vs. n=3). (C) The ratio of
P-AMPKα to AMPK expression (n=3 vs. n=3). The protein ratio of
P-AMPK to AMPK in the cGCs of the PCOS group was significantly
lower than in NOR group. (D) The gray intensity of PGC1α expression
relative to β-actin (n=3 vs. n=3). The expression protein levels of
the energy depletion signaling-related factors SIRT1, AMPK, p-AMPK
and PGC-1α were decreased in the cGCs of the PCOS group. (E) mRNA
transcription level of SIRT1 by RT-qPCR (n=5 vs. n=8). (F) mRNA
transcription level of AMPK by RT-qPCR (n=8 vs. n=5). (G) mRNA
transcription level of PGC-1α by RT-qPCR (n=4 vs. n=7). (H) mRNA
transcription level of TFAM detected by RT-qPCR (n=6 vs. n=6).
There was no statistically significant difference observed in
SIRT1, AMPK and PGC-1α mRNA transcription level. *P<0.05 and
***P<0.001. RT-qPCR, reverse transcription-quantitative PCR;
CON, normal ovarian reserve; PCOS, polycystic ovary syndrome;
SIRT1, silent information regulator 1; AMPK, AMP-activated protein
kinase; P-AMPKα, phosphorylated AMPKα; cGCs, cumulus granulosa
cells; PGC-1α, peroxisome proliferator-activated receptor-γ
coactivator 1α; TFAM, mitochondrial transcription factor A. |
Discussion
PCOS is one of the most common hormonal disorders of
women during the reproductive years. In patients with PCOS, the
mitochondrial state of GCs is altered, the spindle assembly
disordered, and the quality of oocytes decreases (15–17).
Any problems of GCs in growth differentiation, metabolism and
apoptosis will directly or indirectly affect the quality of
follicular (18–20).
In the present study, the mitochondrial structure
and function of cGCs were investigated. As the TEM revealed
increased abnormal mitochondrial in cGCs of the patients with PCOS,
which was in accordance with the study by Zhang et al
(21), MMP decreased, and
mitochondrial swelling was found in the patients with PCOS.
Mitochondria play a major role in intracellular
redox metabolism and are producers of ROS, which mediate oxidative
stress, affect cell development and maturation and aggravate cell
apoptosis (6,22,23).
The present results revealed that the level of ROS in the PCOS
group was significantly higher than in the CON group, suggesting
that the cells had oxidative damage in the PCOS group.
Intracellular ATP levels are often used to assess
mitochondrial function. Studies have shown that ATP deficiency
increases aneuploidy during oocyte development and embryonic
development, which is a common cause of chromosome segregation
(24,25). The decrease in mitochondrial ATP
levels in GCs ultimately limits oocyte development and maturation.
The ATP level in cGCs cells of the CON group was 2.21 times higher
than that of the PCOS group. ATP production is a complex process
involving four protein complexes and ATP synthase; the ATP5A1 gene
encodes the α-subunit of ATP synthase F1 complex, and the ATP5I
gene encodes the e-subunit of ATP synthase F0 complex (26); notably, the present results
revealed that the mRNA transcription levels of ATP5A1 and ATP5I
exhibited no difference between the two groups, thus indicating
that some other enzyme may have a role in ATP synthesis.
Recent studies suggested that there is a crucial
role for mitochondrial dynamics by fission and fusion in the
maintenance of healthy follicular development. Mitochondrial
fission is mediated by DRP1 and FIS1, while mitochondrial fusion is
promoted by MFN2 and OPA1. Inner membrane fusion requires OPA1
(27), whereas outer membrane
fusion requires MFN2 (28).
However, MFN2 and OPA1 showed an obvious imbalance in the
expression and interaction according to the results of the present
study. In the present study, the expression of MFN2 and OPA1 in the
control group was lower. The balance of inner and outer membrane
fusion maintains the normal function of mitochondria (29). DRP1 is recruited from the cytosol
to the outer membrane by mitochondrial dynamics proteins 49 and 51
(Mid49 and 51), FIS1 and MFF (30). Drp1 downregulation was also
associated with mitochondrial elongation. FIS1 protein expression
value of the PCOS group was significantly higher than that of the
CON group in the aforementioned experiment.
However, the molecular mechanisms controlling
mitochondrial function remain unclear. A mitochondrial protein
encoded by nuclear genetic material, PGC-1α belongs to the family
of nuclear-derived proteins. PGC1α tunes the activity of a range of
nuclear transcription factors, including nuclear respiratory factor
1 (NRF1) and NRF2, to control expression of nuclear-encoded OXPHOS
subunits and genes associated with mitochondrial replication,
transcription and translation, including mitoribosomal proteins.
PGC-1α regulates mitochondrial biogenesis and function through
mitochondrial transcription factor A (TFAM), downstream
transcription factors (31). In
the present study, it was revealed that the expression of PGC1-α
decreased in the PCOS ovaries when compared with normal ovaries.
One of these main control factors is TFAM which binds mitochondrial
DNA and regulates mitochondrial transcription and mitochondrial DNA
maintenance (32). It was revealed
that the expression of TFAM decreased in the granulosa cells of
PCOS ovaries when compared with healthy normal ovaries.
Recent studies showed that alterations in the level
of mitochondrial biogenesis can give rise to hyperandrogenism, and
an increase in mitochondrial biogenesis can ameliorate PCOS by
reducing ROS in granulosa cells, thus highlighting the potential
role of mitochondria in PCOS (33,34).
A large number of mitochondrial proteins are acylated, allowing
sirtuins to act as NAD+-dependent deacetylase enzymes,
thus regulating, including deacetylation, demalonylation and
desuccinylation. Recent studies demonstrated that activation of
SIRT1 alleviated mitochondrial damage and enhanced mitochondrial
recovery, to attenuate ischemic liver injury and diabetic
neuropathy (35,36), suggesting that SIRT1 plays an
important role in mitochondrial quality control. In the present
study, it was found that the excessively increased mitochondrial
damage may be caused by decreased expression of SIRT1 in the GCs of
PCOS.
Chronic activation of AMPK is associated with
increased mitochondrial biogenesis, probably mediated by increased
expression of PGC-1a (37). AMPK
phosphorylates DNA methyltransferase 1 (DNMT1) and RbAp46 in human
umbilical vein endothelial cells (HUVECs). This triggers the
inhibition of DNMT1 and leads to important nucleosomal remodeling.
This results in the upregulation of the nuclear genes that encode
the proteins involved in mitochondrial biogenesis and function,
including TFAM and PGC-1a (38).
PGC-1α has the only target of Sirt1 (39). In the present study, the following
explanations were provided for the inconsistent expression levels
of proteins AMPK, DRP1, OPA1 and mRNA: First, the expression of
genes is divided into two levels: Transcription and translation,
namely mRNA level and protein level. There is a spatiotemporal
interval between the time and location of transcription and
translation of eukaryotic gene expression; Second, after
transcription, there are several levels of post-transcriptional
processing, degradation of transcriptional products, translation,
post-translation processing and modification. Translation occurs by
a universally conserved set of stages that can be divided into i)
initiation, whereby the mRNA is loaded onto the ribosome and the
start codon is selected; ii) elongation of the polypeptide by
selective addition of amino acids; and iii) termination of
translation, followed by recycling of the ribosome. Each step is
mediated by nuclear-encoded translation factors that operate in
conjunction with the mitoribosome. Therefore, the level of
transcription and the level of translation are not exactly
consistent; Third, the half-life of mRNA is different from that of
protein. mRNA is highly degradable and exists for a short time in
tissues, while its properties are stable after being translated
into protein; and fourth, the timing of the detection may vary, as
the mRNA has already degraded when the protein reached its peak
value, or the protein amount is still increasing when the mRNA
reached its peak value.
In the present study, it was identified that the low
expression of SIRT1/P-AMPK-PGC-1α pathway protein may lead to
decreased expression of mitochondrial transcription factors in GCs,
which in turn affects mitochondrial function and ultimately affects
the quality of oocytes (Fig. S9).
It is possible to explore ways to improve the function of patients
with PCOS by studying the mechanism of this pathway.
However, the present study has some limitations.
Firstly, the phosphorylation of the DRP1 protein on S616 was not
detected; this may be due to the small number of cumulus granulosa
cells, low protein content or the relatively low proportion of
phosphorylation. Secondly, no in-depth study was performed on the
structure and function of mitochondria in oocytes from patients
with PCOS. Thirdly, further research should be carried out on how
to improve the quality of cGCs in patients with PCOS and thus
improve the quality of oocytes. In conclusion, the present study
revealed that the mitochondria of ovarian GCs in patients with PCOS
exhibit disordered structure and dysfunction; it was deduced that
the decrease in quality of GCs in patients with PCOS may be caused
by the SIRT1-P-AMPK-PGC-1α pathway. Research has suggested that
improving GC mitochondrial function will improve the quality of
oocytes in patients with PCOS, thereby improving clinical
outcomes.
Supplementary Material
Supporting Data
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the Hebei Medical Science
Research Project (grant nos. 20231196 and 20190144) and the
Government clinical medical talent training program (grant no.
ZF2023176).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
CX, HL, XZ, ZA, TC, WY, SW, DS and XW contributed to
the study conception and design. CX performed experiments. XZ, WY,
TC and ZA are responsible for the collection of cumulus granulosa
cells. HL and XW performed data collection. CX, SW, DS and XW
analyzed data. CX wrote the first draft of the manuscript, and all
authors commented on previous versions of the manuscript. CX and XW
confirm the authenticity of all the raw data. All authors read and
approved the final version of the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of the Hebei Reproductive Health Hospital (approval no.
KYY-2021-LW-005; Shijiazhuang, China). Written consent was obtained
from each patient or subject after full explanation of the purpose
and nature of all procedures used. The present study has followed
procedures that were in accordance with the ethical standards as
formulated in the Helsinki Declaration (1964).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Aversa A, La Vignera S, Rago R, Gambineri
A, Nappi RE, Calogero AE and Ferlin A: Fundamental concepts and
novel aspects of polycystic ovarian syndrome: Expert consensus
resolutions. Front Endocrinol (Lausanne). 11:5162020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gleicher N, Darmon S, Patrizio P and Barad
DH: Reconsidering the polycystic ovary syndrome (PCOS).
Biomedicines. 10:15052022. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Liu X, Wang L, Zuo X, Li C and Teng Y:
Women with PCOS with a history of early pregnancy loss show a
higher risk of gestational diabetes mellitus. Int J Gen Med.
14:6409–6416. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sayutti N, Abu MA and Ahmad MF: PCOS and
role of cumulus gene expression in assessing oocytes quality. Front
Endocrinol (Lausanne). 13:8438672022. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Richani D, Dunning KR, Thompson JG and
Gilchrist RB: Metabolic co-dependence of the oocyte and cumulus
cells: essential role in determining oocyte developmental
competence. Hum Reprod Update. 27:27–47. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kirillova A, Smitz JEJ, Sukhikh GT and
Mazunin I: The role of mitochondria in oocyte maturation. Cells.
10:24842021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li W, Cao J, Wang X, Zhang Y, Sun Q, Jiang
Y, Yao J, Li C, Wang Y and Wang W: Ferruginol restores
SIRT1-PGC-1α-mediated mitochondrial biogenesis and fatty acid
oxidation for the treatment of DOX-induced cardiotoxicity. Front
Pharmacol. 12:7738342021. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tian F, Li Q, Shi L, Li J, Shi M, Zhu Y,
Li H and Ge RS: In utero bisphenol AF exposure causes fetal Leydig
cell dysfunction and induces multinucleated gonocytes by generating
oxidative stress and reducing the SIRT1/PGC1α signals. Toxicol Appl
Pharmacol. 447:1160692022. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhao Y, Zhang J, Zheng Y, Zhang Y, Zhang
XJ, Wang H, Du Y, Guan J, Wang X and Fu J: NAD+ improves
cognitive function and reduces neuroinflammation by ameliorating
mitochondrial damage and decreasing ROS production in chronic
cerebral hypoperfusion models through Sirt1/PGC-1α pathway. J
Neuroinflammation. 18:2072021. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tian L, Cao W, Yue R, Yuan Y, Guo X, Qin
D, Xing J and Wang X: Pretreatment with Tilianin improves
mitochondrial energy metabolism and oxidative stress in rats with
myocardial ischemia/reperfusion injury via AMPK/SIRT1/PGC-1 alpha
signaling pathway. J Pharmacol Sci. 139:352–360. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chandrasekaran K, Anjaneyulu M, Choi J,
Kumar P, Salimian M, Ho CY and Russell JW: Role of mitochondria in
diabetic peripheral neuropathy: Influencing the
NAD+-dependent SIRT1-PGC-1α-TFAM pathway. Int Rev
Neurobiol. 145:177–209. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kalliora C, Kyriazis ID, Oka SI, Lieu MJ,
Yue Y, Area-Gomez E, Pol CJ, Tian Y, Mizushima W, Chin A, et al:
Dual peroxisome-proliferator-activated-receptor-α/γ activation
inhibits SIRT1-PGC1α axis and causes cardiac dysfunction. JCI
Insight. 5:e1295562019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Emidio GD, Placidi M, Rea F, Rossi G,
Falone S, Cristiano L, Nottola S, D'Alessandro AM, Amicarelli F,
Palmerini MG and Tatone C: Methylglyoxal-dependent glycative stress
and deregulation of SIRT1 functional network in the ovary of PCOS
mice. Cells. 9:2092020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
He Y, Lu Y, Zhu Q, Wang Y, Lindheim SR, Qi
J, Li X, Ding Y, Shi Y, Wei D, et al: Influence of metabolic
syndrome on female fertility and in vitro fertilization outcomes in
PCOS women. Am J Obstet Gynecol. 221:138.e1–138.e12. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gong Y, Luo S, Fan P, Zhu H, Li Y and
Huang W: Growth hormone activates PI3K/Akt signaling and inhibits
ROS accumulation and apoptosis in granulosa cells of patients with
polycystic ovary syndrome. Reprod Biol Endocrinol. 18:1212020.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Guijarro LG, Sanmartin-Salinas P,
Pérez-Cuevas E, Toledo-Lobo MV, Monserrat J, Zoullas S, Sáez MA,
Álvarez-Mon MA, Bujan J, Noguerales-Fraguas F, et al: Actinomycin D
arrests cell cycle of hepatocellular carcinoma cell lines and
induces p53-dependent cell death: A study of the molecular
mechanism involved in the protective effect of IRS-4.
Pharmaceuticals (Basel). 14:8452021. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Patil K, Shinde G, Hinduja I and Mukherjee
S: Compromised cumulus-oocyte complex matrix organization and
expansion in women with PCOS. Reprod Sci. 29:836–848. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yefimova MG, Lefevre C, Bashamboo A,
Eozenou C, Burel A, Lavault MT, Meunier AC, Pimentel C, Veau S,
Neyroud AS, et al: Granulosa cells provide elimination of apoptotic
oocytes through unconventional autophagy-assisted phagocytosis. Hum
Reprod. 35:1346–1362. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lai Q, Xiang W, Li Q, Zhang H, Li Y, Zhu
G, Xiong C and Jin L: Oxidative stress in granulosa cells
contributes to poor oocyte quality and IVF-ET outcomes in women
with polycystic ovary syndrome. Front Med. 12:518–524. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang Q, Ren J, Wang F, Pan M, Cui L, Li M
and Qu F: Mitochondrial and glucose metabolic dysfunctions in
granulosa cells induce impaired oocytes of polycystic ovary
syndrome through Sirtuin 3. Free Radic Biol Med. 187:1–16. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chiang JL, Shukla P, Pagidas K, Ahmed NS,
Karri S, Gunn DD, Hurd WW and Singh KK: Mitochondria in ovarian
aging and reproductive longevity. Ageing Res Rev. 63:1011682020.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Guijarro LG, Cano-Martínez D, Toledo-Lobo
MV, Ruiz-Llorente L, Chaparro M, Guerra I, Iborra M, Cabriada JL,
Bujanda L, Taxonera C, et al: Evaluation of AIF-1 (allograft
inflammatory factor-1) as a biomarker of Crohn's disease severity.
Biomedicines. 10:7272022. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Adhikari D, Lee IW, Yuen WS and Carroll J:
Oocyte mitochondria-key regulators of oocyte function and potential
therapeutic targets for improving fertility. Biol Reprod.
106:366–377. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhao S, Heng N, Wang H, Wang H, Zhang H,
Gong J, Hu Z and Zhu H: Mitofusins: From mitochondria to fertility.
Cell Mol Life Sci. 79:3702022. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu Y, Han M, Li X, Wang H, Ma M, Zhang S,
Guo Y, Wang S, Wang Y, Duan N, et al: Age-related changes in the
mitochondria of human mural granulosa cells. Hum Reprod.
32:2465–2473. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Maremanda KP, Sundar IK and Rahman I: Role
of inner mitochondrial protein OPA1 in mitochondrial dysfunction by
tobacco smoking and in the pathogenesis of COPD. Redox Biol.
45:1020552021. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen L, Liu B, Qin Y, Li A, Gao M, Liu H
and Gong G: Mitochondrial fusion protein Mfn2 and its role in heart
failure. Front Mol Biosci. 8:6812372021. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yapa NMB, Lisnyak V, Reljic B and Ryan MT:
Mitochondrial dynamics in health and disease. FEBS Lett.
595:1184–1204. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kalia R, Wang RY, Yusuf A, Thomas PV,
Agard DA, Shaw JM and Frost A: Structural basis of mitochondrial
receptor binding and constriction by DRP1. Nature. 558:401–405.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kim HK, Jeon J, Song IS, Heo HJ, Jeong SH,
Long LT, Thu VT, Ko TH, Kim M, Kim N, et al: Tetrahydrobiopterin
enhances mitochondrial biogenesis and cardiac contractility via
stimulation of PGC1α signaling. Biochim Biophys Acta Mol Basis Dis.
1865:1655242019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hao L, Zhong W, Dong H, Guo W, Sun X,
Zhang W, Yue R, Li T, Griffiths A, Ahmadi AR, et al: ATF4
activation promotes hepatic mitochondrial dysfunction by repressing
NRF1-TFAM signalling in alcoholic steatohepatitis. Gut.
70:1933–1945. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sun L, Tian H, Xue S, Ye H, Xue X, Wang R,
Liu Y, Zhang C, Chen Q and Gao S: Circadian clock genes REV-ERBs
inhibits granulosa cells apoptosis by regulating mitochondrial
biogenesis and autophagy in polycystic ovary syndrome. Front Cell
Dev Biol. 9:6581122021. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Safaei Z, Bakhshalizadeh S, Nasr-Esfahani
MH, Akbari Sene A, Najafzadeh V, Soleimani M and Shirazi R: Vitamin
D3 affects mitochondrial biogenesis through mitogen-activated
protein kinase in polycystic ovary syndrome mouse model. J Cell
Physiol. 235:6113–6126. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang C, Liu T, Tong Y, Cui R, Qu K, Liu C
and Zhang J: Ulinastatin protects against acetaminophen-induced
liver injury by alleviating ferroptosis via the SIRT1/NRF2/HO-1
pathway. Am J Transl Res. 13:6031–6042. 2021.PubMed/NCBI
|
|
36
|
Jalgaonkar MP, Parmar UM, Kulkarni YA and
Oza MJ: SIRT1-FOXOs activity regulates diabetic complications.
Pharmacol Res. 175:1060142022. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Guo A, Li K and Xiao Q: Fibroblast growth
factor 19 alleviates palmitic acid-induced mitochondrial
dysfunction and oxidative stress via the AMPK/PGC-1α pathway in
skeletal muscle. Biochem Biophys Res Commun. 526:1069–1076. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ortega MA, De Leon-Oliva D, Garcia-Montero
C, Fraile-Martinez O, Boaru DL, Del Val Toledo Lobo M, García-Tuñón
I, Royuela M, García-Honduvilla N, Bujan J, et al: Understanding
HAT1: A comprehensive review of noncanonical roles and connection
with disease. Genes (Basel). 14:9152023. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang ZH, Bao XG, Hu JJ, Shen SB, Xu GH and
Wu YL: Nicotinamide riboside enhances endothelial precursor cell
function to promote refractory wound healing through mediating the
Sirt1/AMPK pathway. Front Pharmacol. 12:6715632021. View Article : Google Scholar : PubMed/NCBI
|