Introduction
Myasthenia gravis (MG) is a rare chronic autoimmune
disease induced by the postsynaptic membrane receptors and
autoantibody attack at the neuromuscular junction (NMJ) (1,2). MG
manifests as fatigue and muscle weakness, and is associated with
diplopia, ptosis and systemic symptoms (3). Previous studies have reported that
there are several types of subgroups of MG and that the majority of
patients with MG are positive for acetylcholine receptor (AChR)
autoantibodies (4–6). At present, there are multiple
treatment options available for MG, such as cholinesterase
inhibitors, glucocorticoids, immunosuppressants, intravenous
immunoglobulins, plasmapheresis and thymectomy, which primarily
focus on immunoregulatory mechanisms (7); however, the efficacy of these
treatments is limited. Additionally, the numerous side effects of
treatments, and the economic burden of therapeutics and surgery
further reduce the quality of life of patients. A real-world study
showed that after long-term treatment, two-thirds of patients still
had moderate to severe symptoms (8). Furthermore, a study performed in
Sweden on 1,077 patients showed that ~50% of the patients were
‘dissatisfied’ with the control of their symptoms (9). MG is a heavy burden on patients
worldwide. Although research has previously been performed on MG
(10); its pathogenesis has not
been fully elucidated and there is an urgent need for novel
treatment options.
Mitochondria are the primary organelles that provide
energy for cell metabolism. Notably, mitochondrial dysfunction
affects human health (11), and an
increasing number of studies have revealed that mitochondria are
regarded as a therapeutic target for numerous diseases (12,13).
Furthermore, it has been identified that mitochondria serve key
roles in skeletal muscle function and mitochondrial dysfunction is
related to skeletal muscle damage (14). Previous studies have highlighted
the crucial role of normal mitochondrial function in preserving NMJ
structure and neurotransmission (15–18);
therefore, restoring mitochondrial function holds promise for
ameliorating NMJ damage and treating MG (19,20).
Previous studies have successfully established an animal model of
MG and have performed a series of experimental studies on
mitochondria, suggesting that improving mitochondrial function can
reduce the symptoms of experimental autoimmune (EA)MG in a rat
model (19,21,22).
Mitophagy and apoptosis are the primary means of
maintaining mitochondrial homeostasis. Mitophagy is a protective
phenomenon that requires the participation of lysosomes, whereas
apoptosis is a type of programmed cell death (23). Mitophagy can eliminate the impact
of mitochondrial dysfunction, including the excess reactive oxygen
species produced, inflammation and oxidative stress (24). Notably, in neurodegenerative
diseases typically characterized by the loss of neurons, in which
mitophagy and apoptosis are widely involved, abnormalities in
synaptic function and instability of the nervous system have been
observed (25). Multiple studies
have reported that mitophagy and apoptosis serve an important role
in diseases, such as spinal muscular atrophy and Duchenne muscular
dystrophy (26–29) and regulating mitophagy and
apoptosis may exert protective effects on skeletal muscles
(30,31). Nevertheless, to the best of our
knowledge, no previous studies have elucidated the mechanism
underlying aberrant mitophagy and apoptosis in MG. Our unpublished
study examined isolated peripheral blood mononuclear cells (PBMC)
from patients with MG and observed a decrease in the expression of
key signaling molecules involved in promoting mitophagy, along with
an increase in the expression of key signaling molecules involved
in promoting apoptosis, compared to healthy subjects. Thus,
exploring the underlying mechanisms of dysregulated mitophagy and
apoptosis in MG is important.
Astragaloside IV (AS-IV) is the primary bioactive
component derived from Astragalus membranaceus and serves a
protective role in various diseases (32). In our previous study, Qiangji
Jianli decoction (QJJLD), which is a clinically effective
traditional Chinese medicine used for the treatment of MG, was
explored and shown to significantly improve gastrocnemius muscle
injury in EAMG rats (19). In
addition, AS-IV has been identified as the primary component in
QJJLD using high-performance liquid chromatography analysis
(33); thus, whether AS-IV exerts
a similar therapeutic role in MG was assessed in the present study.
AS-IV has been reported to exert a potential neuroprotective
effect, particularly in neurodegenerative disorders, such as
Parkinson's disease (PD) and autoimmune encephalomyelitis (AE)
(34). Additionally, traditional
Chinese medicines with several multiple active ingredients, such as
ginsenoside Rg1 (Rg1), tanshinone IIA (TSA) and AS-IV, which have
been shown to exert notable advantages including anti-inflammatory,
antioxidant, and anti-apoptotic properties, contributing to
neuroprotective effects (35).
AS-IV has also been reported to regulate the immune response by
regulating the T helper 17/regulatory T cell ratio in vivo
(36). Furthermore, AS-IV exerts a
neuroprotective role by maintaining mitochondrial homeostasis via
the regulation of apoptosis and autophagy (37). Notably, AS-IV has been shown to not
only regulate mitochondrial dysfunction and inhibit apoptosis of
skeletal muscle cells, but also to improve the motor ability of
muscles and reverse skeletal muscle injury (38). However, a comprehensive account of
the effects of AS-IV on the treatment of MG is, to the best of our
knowledge, lacking.
Therefore, in the present study, the protective
mechanism of AS-IV in ameliorating EAMG was assessed, with a focus
on the regulation of mitophagy and apoptosis, in order to determine
the potential mechanism of action and highlight a novel potential
therapeutic option for the treatment of MG.
Materials and methods
Animals
A total of 36 female specific pathogen-free (SPF)
Lewis rats (weight, 120–140 g; age, 5–6 weeks) were purchased from
Charles River Laboratories, Inc. Rats were maintained in SPF
conditions at 23±2°C, 55±5% relative humidity, and under a 12-h
light/dark cycle with ad libitum access to food and drinking
water. Pentobarbital sodium (50 mg/kg) was injected
intraperitoneally for anesthesia prior to modeling and repetitive
nerve stimulation (RNS) detection to minimize animal pain. All
experimental procedures were approved by the Experimental Animal
Ethics Committee of Guangzhou University of Chinese Medicine
(approval no. 20200509001; Guangzhou, China) and adhered to the
National Institutes of Health Guide for the Care and Use of Animals
guidelines (39).
Chemicals and reagents
Rat 97–116 peptide of the AChR-α subunit (R97-116,
DGDFAIVKFTKVLLDYTGHI) was synthesized by GL Biochem (Shanghai) Ltd.
Complete Freund's adjuvant (CFA), incomplete Freund's adjuvant
(IFA) and sodium pentobarbital were purchased from MilliporeSigma
(cat. nos. F5881, F5506 and P3761, respectively). PBS (cat. no.
10010-023) was purchased from Gibco (Thermo Fisher Scientific,
Inc.). An AChR-antibody (AChR-Ab) assay kit (cat. no. ml003131) was
obtained from Shanghai Enzyme-linked Biotechnology Co., Ltd. AS-IV
(CAS no. 84687-43-4) was purchased from Chengdu Herbpurify Co.,
Ltd. A Hematoxylin and Eosin (H&E) Staining Kit (cat. no.
C0105S) was purchased from Beyotime Institute of Biotechnology.
ATPase and cytochrome c (Cyt-C) oxidase (COX) assay kits
were obtained from Leagene; Beijing Regan Biotechnology Co., Ltd.
(cat. nos. DE0045 and DE0031, respectively). Forward and reverse
primers were purchased from Generay Biotech Co., Ltd. Phosphatase
and tensin homolog-induced putative kinase 1 (PINK1), LC3I/II,
Bcl-2, caspase 9 and GAPDH primary antibodies were purchased from
Abcam (cat. nos. ab23707, ab128025, ab59348, ab52298 and ab8245,
respectively). Parkin, p62, Bax, Cyt-C and caspase 3 primary
antibodies were obtained from Cell Signaling Technology, Inc. (cat.
nos. 4211, 39749, 2772, 11940 and 9662, respectively).
Establishment of the EAMG animal
model
A total of 36 female Lewis rats were acclimated for
1 week in an SPF-level animal room, of which eight were randomly
selected as the control group and the remaining 28 rats as the
model group. The day of the first immunization was considered day
0. The modeling agent was prepared by mixing 100 µg R97-116
peptide, 0.1 ml CFA and 0.1 ml PBS evenly to form an emulsion.
After the preparation of the emulsion mixture, four sites were
selected for immunogen injection, including the back, abdomen, and
left and right hind foot pads. Each site was injected with 50 µl
immune emulsion and the injection site was washed with 75% ethanol.
Each rat received a total of 200 µl modeling agent during the first
immunization. The second and third immunization boosts were
performed on days 30 and 45, respectively, after the first
immunization. For these subsequent immunizations, the reagent was
prepared by mixing 50 µg R97-116 peptide, 0.1 ml IFA and 0.1 ml
PBS, which was injected subcutaneously at the same sites. During
the second and third immunization periods, each rat received a
total of 200 µl modeling agent each time. During each immunization
period, for rats in the control group, the rats were injected
subcutaneously with a mixed emulsion of 0.1 ml PBS and 0.1 ml CFA
or IFA without R97-116 peptide, and the injection sites and doses
were the same as those of the model group rats (40). During the modeling period, the
mental state, activity, appetite, stool characteristics and hair
luster of each group were recorded daily. The body weight of the
rats was measured twice a week, and the Lennon score was recorded.
The Lennon score was evaluated using the criteria described
previously (41,42), and the assigned Lennon scores were
as follows: Grade 0, no weakness; grade 1, fatigability or weakness
only observed after exercise; grade 2, clinical signs of weakness
present before exercise, including hunched posture or head down;
grade 3, severe muscle weakness with dyspnea/apnea or moribund
state; grade 4, death. Intermediate signs in rats were categorized
into grades 0.5, 1.5, 2.5 or 3.5, accordingly. The model was
evaluated 1 week after the third immunization. Rats in both groups
undergoing experimental treatment were euthanized at day 80 or when
they reached a humane endpoint, such as self-harming behavior,
severe weight loss of 20%, compromised breathing or lack of
response to external stimuli. Sodium pentobarbital (150 mg/kg) was
injected intraperitoneally to euthanize the rats. Death was
confirmed by monitoring the cessation of breathing and heartbeat,
and pupil dilation. Throughout the course of the experiment, only
one rat in the model group reached the predefined humane endpoints
at the day 49.
RNS
After anesthesia, the rats in the model group were
immobilized. Stimulating electrodes of the electrophysiological
recorder (cat. no. PL3516; PowerLab; ADInstruments, Ltd.) were
placed near the sciatic nerve in the gastrocnemius muscle,
recording electrodes were subcutaneously implanted in the medial
head of the gastrocnemius muscle and Achilles tendon on the same
side, and reference electrodes were subcutaneously implanted in the
abdomen (43). All electrodes
received 10 pulses of 5 Hz electrical stimulation, and the
percentage change of the first and fifth action potentials was
measured. A positive result was defined as an attenuation rate of
>10%.
Detection of AChR-Ab
On day 7 after modeling completion, rats were
anesthetized with an intraperitoneal injection of 30 mg/kg
pentobarbital sodium. Subsequently, a single blood sample of
0.2–0.4 ml was collected from the orbital venous plexus of each
rat. Additionally, upon completion of the entire experiment and
euthanasia of the rats, a blood sample of 0.8–1 ml was collected
once from the abdominal aorta. The collected blood samples were
placed in centrifuge tubes for 30 min and left to stand. The tubes
were then centrifuged at 1,500 × g for 10 min at 4°C, and the
supernatant was collected and stored at −80°C. The AChR-Ab levels
in the rats were measured in the serum using the aforementioned
specific ELISA kit.
Drug intervention
Following three separate immunizations, 24 of the 28
rats in the model group were successfully modeled, and the rate at
which the EAMG rat model was successfully established was 85.71%.
The model group was randomly divided into three subgroups: Model
group, low-dose AS-IV (L-ASIV) group and high-dose AS-IV (H-ASIV)
group (n=8 rats/group). The rats in the L-ASIV and H-ASIV groups
were treated with 40 and 80 mg/kg/day AS-IV by gavage, respectively
(44–46). The control and model groups were
administered equal volumes of saline by gavage, and the gavage
volume was calculated according to the principle of 1 ml/kg body
weight. During the four-week Drug intervention, rats in each group
were administered via gavage once a day, following the respective
treatment method. The rats were fasted for 24 h after the last
administration. Sodium pentobarbital (150 mg/kg) was injected
intraperitoneally for euthanasia, and death was confirmed by
monitoring the cessation of breathing and heartbeat, and pupil
dilation (47).
H&E staining
After the rats were sacrificed, the gastrocnemius
muscle tissue was immediately removed and fixed overnight at room
temperature in 4% paraformaldehyde and was then embedded in
conventional paraffin. The tissues were cut into 4-µm sections,
which were stained with 100% hematoxylin solution at room
temperature for 2 min and were then differentiated in 1%
hydrochloric acid ethanol for several seconds. After rinsing with
water and dehydrating in a series of increasing concentrations of
ethanol, the sections were stained with 100% eosin solution at room
temperature for 1 min. The sections were then placed in xylene
until they became transparent and were mounted with neutral gum.
Images of the sections from each group were captured under an
optical microscope at a magnification of ×400 (Olympus BX53;
Olympus Corporation).
Transmission electron microscopy
(TEM)
After 4 weeks of treatment, gastrocnemius muscle
samples were cut to a volume of 1 mm3 and fixed with
2.5% glutaraldehyde at 4°C for 2–6 h. The sections were then rinsed
in phosphoric acid buffer solution for 30 min (repeated six times),
fixed with 1% osmic acid at 4°C for 1.5–2 h, and rinsed again in
phosphoric acid buffer solution for 10 min (repeated three times).
After dehydration in a series of increasing concentrations of
ethanol, the tissue samples were embedded in epoxy resin monomer
overnight at 37°C. An ultramicrotome (RM2016; Leica Microsystems
GmbH) was used for sectioning (60 nm), and the samples were then
stained with uranium dioxide acetate at room temperature for 12 min
and with lead citrate at room temperature for 8 min. Finally, the
ultrastructure was observed using TEM (Jem-1400; JEOL, Ltd.).
ATPase staining
An ATPase staining kit was used for ATPase staining.
Fresh gastrocnemius muscle tissues were embedded in a frozen
section embedding agent and frozen at −20°C for further processing.
The embedded tissues were then cut into 6 µm frozen sections and
placed in distilled water. The sections were incubated in calcium
salt solution at 37°C for 5 min, then incubated with ATPase
incubation solution at 37°C for 40 min, followed by washing with
distilled water. Subsequently, the sections were immersed in cobalt
nitrate solution at 37°C for 5 min and then in distilled water.
They were then fixed with 4% neutral formalin at room temperature
for 2 min and rinsed with running water for 2 min. The sections
were placed in a prepared sulfide working solution and incubated
for 2 min. After washing with water, the sections were mounted with
glycerol gelatin onto slides. Images were captured under a
fluorescence microscope (Olympus Corporation) at ×50 magnification
and the average optical density values of the images were
calculated using ImagePro Plus (version 6.0; Media Cybernetics,
Inc.).
Assay of COX activity
The frozen sections of gastrocnemius muscle of rats
were dehydrated in an increasing concentration gradient of ethanol
(5 min at each concentration). The frozen sections were then soaked
in xylene solution twice (10 min each), the excess liquid on the
slide was removed with absorbent paper and the slides were sealed
with neutral gum. Subsequently, DAB solution was added to the
frozen sections and incubated at 37°C in the dark for 90 min,
before the sections were washed with distilled water three times (3
sec each). The staining of frozen sections was observed under an
optical microscope (EcliPse E100; Nikon Corporation).
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was obtained from the gastrocnemius muscle
tissue samples of rats using TRNzol-A+ (Tiangen Biotech
Co., Ltd.). cDNA was synthesized by RT using a PrimeScript™ RT
MasterMix Kit (Takara Bio, Inc.). The expression of target genes
was quantified by qPCR with TB Green™ Premix Ex Taq™ II (Takara
Bio, Inc.) using a CFX96 Real-Time PCR System (Bio-Rad
Laboratories, Inc.). The RT temperature protocol was 37°C for 15
min and 85°C for 5 sec. The thermocycling conditions for qPCR
amplification were 95°C for 30 sec for initial denaturation;
followed by 40 cycles of 95°C for 5 sec and 60°C for 30 sec and
extension at 72°C for 20 sec, followed by a final extension at 72°C
for 5 min. The primer sequences of all target genes are shown in
Table I. Relative quantification
was performed using the 2−ΔΔCq method (48), with GAPDH as the reference
gene.
 | Table I.Primers used for reverse
transcription-quantitative PCR. |
Table I.
Primers used for reverse
transcription-quantitative PCR.
| Gene | Forward primer,
5′-3′ | Reverse primer,
3′-5′ |
|---|
| PINK1 |
CATGGCTTTGGATGGAGAGT |
TGGGAGTTTGCTCTTCAAGG |
| Parkin |
CTGGCAGTCATTCTGGACAC |
CTCTCCACTCATCCGGTTTG |
| p62 |
CCAGCACAGGCACAGAAGATAAGAG |
TCCCACCGACTCCAAGGCTATC |
| LC3II |
TCGCCGACCGCTGTAAGGAG |
CGCCGGATGATCTTGACCAACTC |
| Caspase 3 |
TCTACCGCACCCGGTTACTA |
CGTACAGTTTCAGCATGGCG |
| Caspase 9 |
CTGAGCCAGATGCTGTCCCATA |
GACACCATCCAAGGTCTCGATGTA |
| Bcl-2 |
TCCTTCCAGCCTGAGAGCAACC |
CGACGGTAGCGACGAGAGAAGT |
| Bax |
CCCCCGAGAGGTCTTTTTCCG |
GGGCCTTGAGCACCAGTTTGC |
| Cyt-C |
GCTAAACACCAGGACGGAACT |
CCACTCCCAATCAGGCATGAAC |
| GAPDH |
TGATTCTACCCACGGCAAGTT |
TGATGGGTTTCCCATTGATGA |
Western blot analysis
Total protein was extracted from the gastrocnemius
muscle tissues with RIPA lysis buffer (cat. no. KGP2100; Nanjing
KeyGen Biotech Co., Ltd.) on ice, then centrifuged at 10,000 × g
for 15 min at 4°C to collect the supernatants. A bicinchoninic acid
assay (cat. no. KGP902; Nanjing KeyGen Biotech Co., Ltd.) was used
to measure the protein concentration. Equal amounts of protein (30
µg) were loaded on 10 and 12% SDS-gels, resolved using SDS-PAGE and
subsequently transferred to a PVDF membrane. Membranes were blocked
using 5% skimmed milk (cat. no. 232100; Difco; BD Biosciences) at
room temperature for 1 h and were then incubated overnight with
primary antibodies at 4°C with agitation. The following primary
antibodies were used: Anti-PINK1 (1:1,000), anti-LC3A/B (1:1,000),
anti-Bcl-2 (1:1,000), anti-caspase 9 (1:1,000), anti-GAPDH
(1:5,000), anti-Parkin (1:1,000), anti-SQSTM1/p62 (1:1,000),
anti-caspase 3 (1:1,000), anti-Bax (1:1,000) and anti-Cyt-C
(1:1,000). Subsequently, the membranes were washed and incubated
with the secondary HRP-conjugated Goat Anti-Rabbit IgG antibody
(1:3,000; cat. no. CW0103S; CW Biosciences) or Goat Anti-Mouse IgG
antibody (1:3,000; cat. no. CW0102S; CW Biosciences) at room
temperature for 1 h, and the protein bands were observed using a
Western ECL Substrate kit (cat. no. 170-5061; Bio-Rad Laboratories,
Inc.). Signals were visualized using a gel imaging system (Chemi
Doc XRS+; Bio-Rad Laboratories, Inc.), and Image Lab software
(version 6.0; Bio-Rad Laboratories, Inc.) was used for
densitometric analysis. All experiments were repeated independently
at least three times.
Statistical analysis
All data are presented as the mean ± standard
deviation or as median and IQR. Graphs were generated using
GraphPad Prism version 9 (Dotmatics). SPSS version 22.0 (IBM Corp.)
was used for data processing. Statistical differences between two
groups were determined using unpaired Student's t-test. Comparisons
among more than two groups were carried out using one-way analysis
of variance, followed by the Tukey's post hoc test. The
nonparametric Mann-Whitney U test or Kruskal-Wallis test followed
by Dunn's multiple comparisons test were used to analyze the Lennon
score data. P<0.05 was considered to indicate a statistically
significant difference.
Results
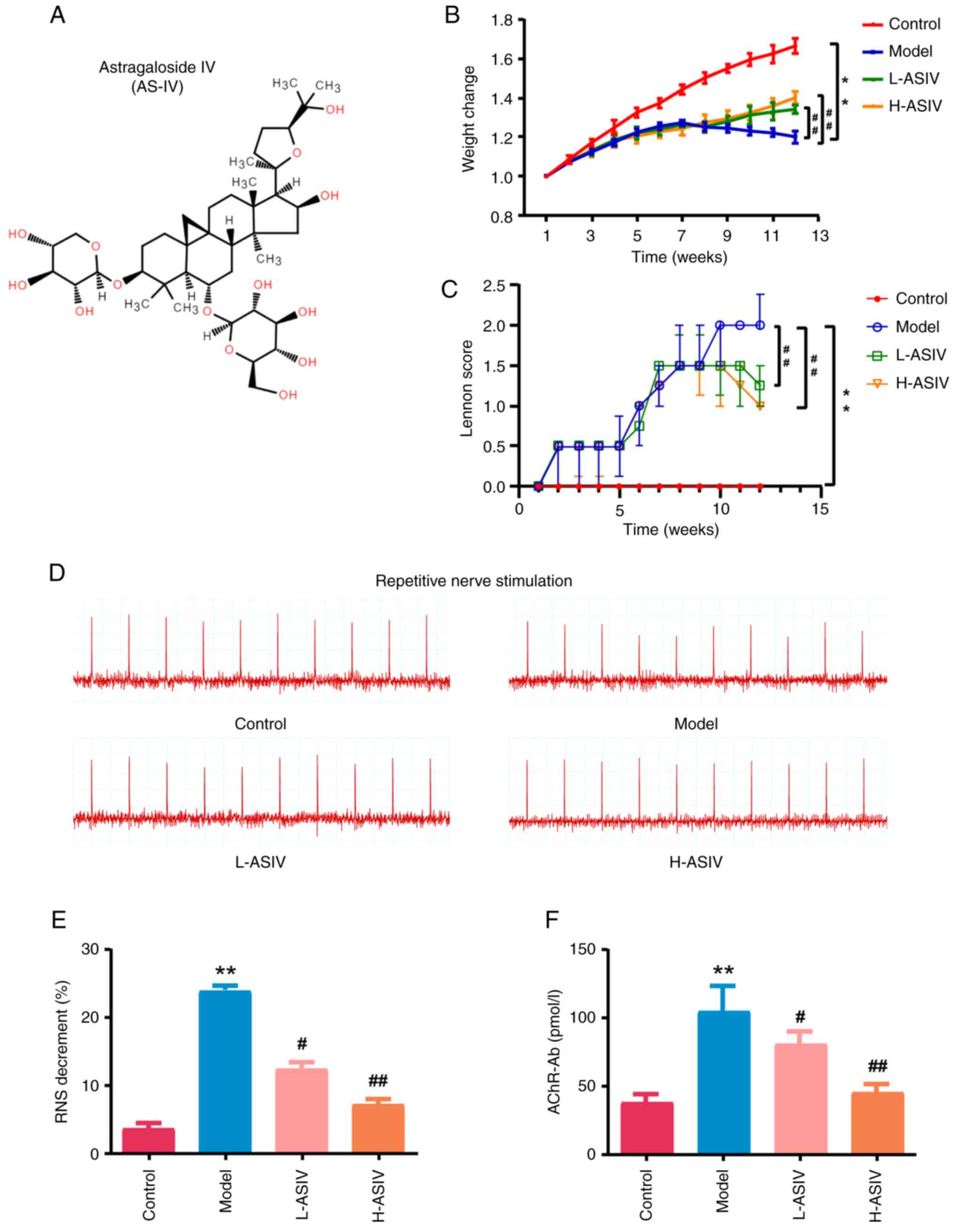
Construction of the EAMG rat
model
Lewis rats were immunized by subcutaneously
injecting peptide R97-116 as an antigen to induce EAMG (Fig. 1A); ~20 days after the first
immunization, the majority of the rats in the model group exhibited
a decrease in water and food consumption, weight loss and muscle
weakness (increased Lennon score), compared with the rats in the
control group (Fig. 1B and C).
After the third booster immunization, one rat was sacrificed due to
reaching humane endpoints, and the rest of the model group rats
exhibited decreased activity, low and weak cries, dull and yellow
fur, marked weight loss and muscle weakness (increased Lennon
score); the differences between the model group and the control
group were statistically significant (both P<0.01; Fig. 1B and C). After modeling, the rats
were anesthetized and the changes in the response to RNS in each
group were measured using electromyography. The attenuation of the
first action potential amplitude and the fifth action potential
amplitude of model rats was >10%, and the differences between
the model group and the control group was statistically significant
(P<0.01; Fig. 1D and E). ELISA
was used to detect the levels of AChR-Ab in the rats. Compared with
in the control group, the AChR-Ab content in the serum of rats in
the model group was significantly increased (P<0.01; Fig. 1F).
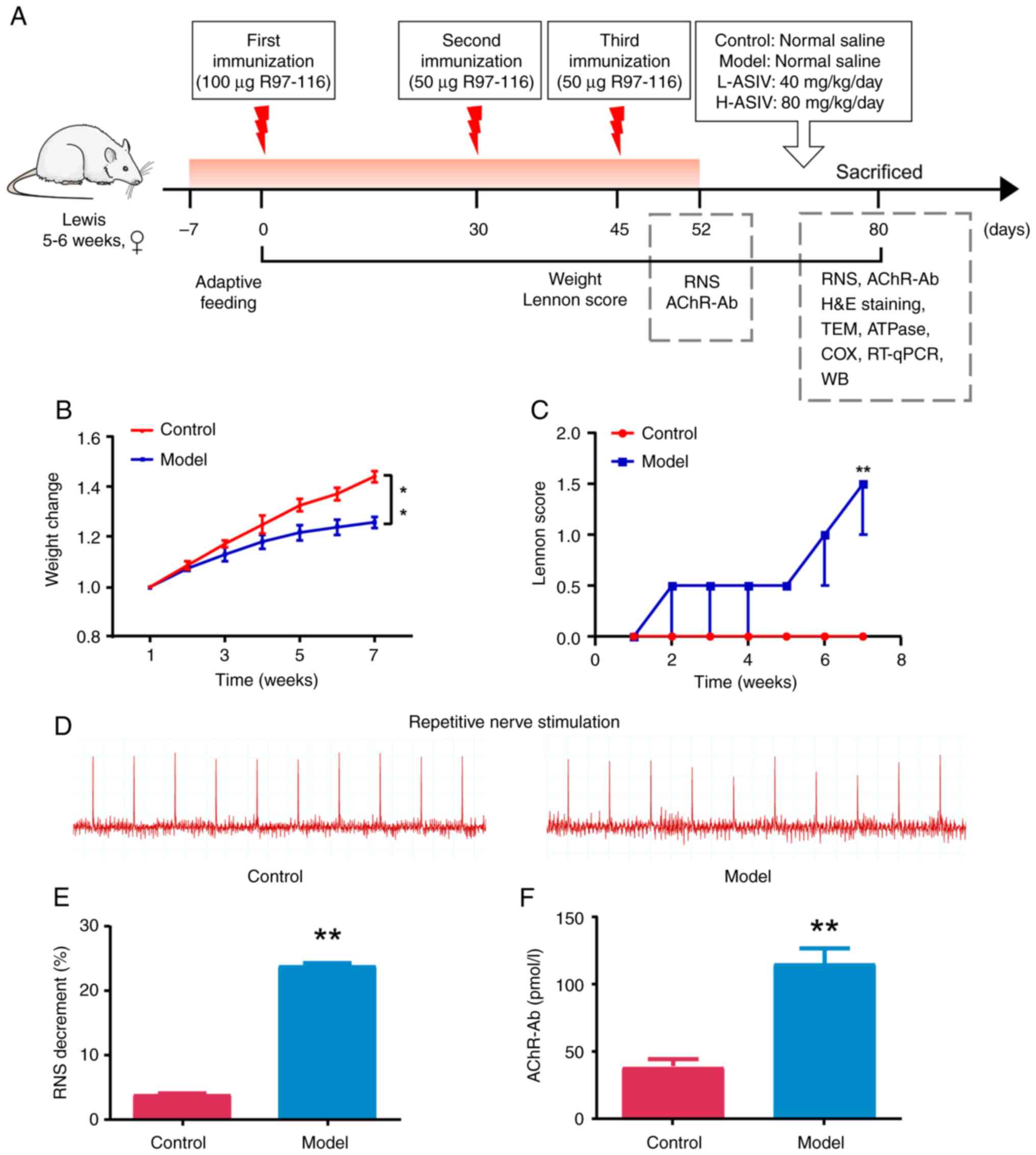 | Figure 1.Construction and evaluation of the
EAMG rat model. (A) Experimental protocol for treatment of EAMG
rats with AS-IV. (B) Weight change rate after modeling. (C) Changes
in Lennon score in different groups. (D) Degree of electrical
attenuation was detected by RNS in both the control and model
groups at 1 week after the last immunization. (E) Quantitative
diagram of the RNS analysis. The amplitude attenuation of the first
and fifth action potentials in the model group decreased by
>10%, which was considered positive. (F) Rat serum samples were
collected at the indicated time points after disease induction and
were analyzed by ELISA for rat AChR-Ab. **P<0.01 vs. control.
AChR-Ab, acetylcholine receptor antibody; AS-IV, astragaloside IV;
COX, cytochrome c oxidase; EAMG, experimental autoimmune
myasthenia gravis; H-ASIV, high-dose AS-IV; H&E, hematoxylin
and eosin; L-ASIV, low-dose AS-IV; RNS, repetitive nerve
stimulation; RT-qPCR, reverse transcription-quantitative PCR; TEM,
transmission electron microscopy; WB, western blotting. |
Effects of AS-IV on EAMG rats
After modeling, the different groups were treated
with normal saline or AS-IV (Fig.
2A) for 4 weeks. In the model group, the weight and muscle
strength of rats continued to decline after treatment with normal
saline. After 4 weeks of treatment, compared with in the model
group, the average weight of EAMG rats in the H-ASIV and L-ASIV
groups significantly increased (P<0.01; Fig. 2B). The Lennon score also
significantly decreased, indicating improved MG symptoms
(P<0.01; Fig. 2C).
Electromyography (EMG) was then used to observe the changes in the
response to RNS in each group, and the levels of AChR-Ab in the
blood samples from each group were measured. There was no
significant change in the EMG amplitude in the control group, but
the EMG amplitude attenuation rate in the model group was >10%
compared with that in the control group (P<0.01; Fig. 2D and E). By contrast, the RNS
amplitude attenuation of rats in the H-ASIV and L-ASIV groups
recovered and was significantly reduced compared with that in the
model group (P<0.01 and P<0.05, respectively; Fig. 2D and E). Compared with in the
control group, the AChR-Ab levels of rats in the model group were
significantly increased (P<0.01; Fig. 2F). By contrast, the AChR-Ab levels
in the blood samples from the H-ASIV and L-ASIV groups were
significantly reduced compared with those in the model group
(P<0.01 and P<0.05, respectively; Fig. 2F). Taken together, these results
suggested that AS-IV can improve the symptoms of EAMG in a rat
model.
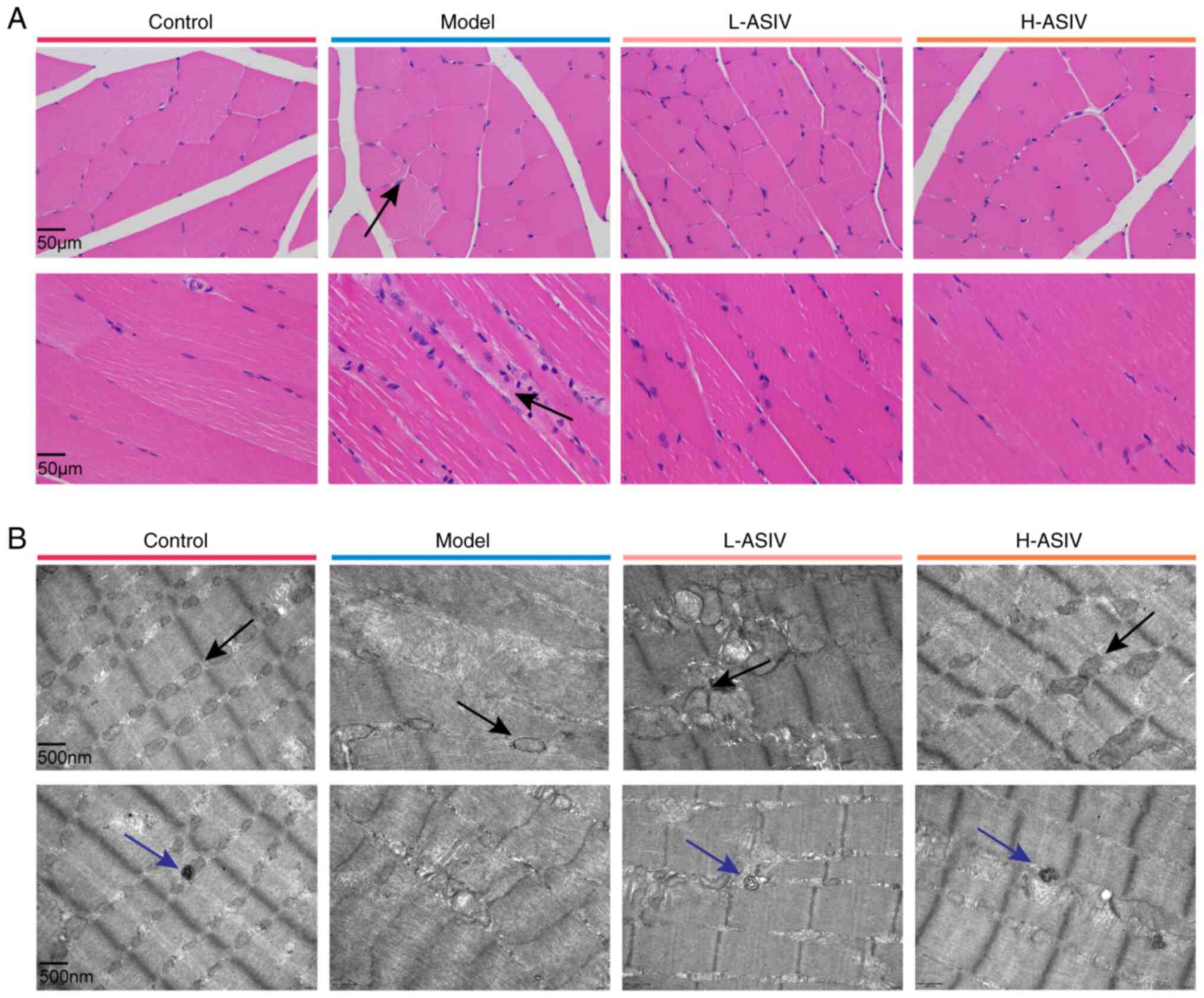
Effects of AS-IV on the morphology and
ultrastructure of gastrocnemius muscle
The muscle fibers of rats in the control group were
tight, normal and orderly, with nuclei evenly distributed at the
edge of the fibers (Fig. 3A). In
the model group, muscle fiber atrophy, widening of the spaces
between fibers, edema and degeneration, connective tissue formation
and mass inflammatory cell infiltration were observed. Following
treatment with AS-IV, both the low and high dose groups showed
varying degrees of improvement, especially at the higher dose, and
the muscle fiber morphology was similar to that observed in the
control group (Fig. 3A).
TEM was used to observe changes in the muscle fiber
arrangement and mitochondrial ultrastructure in the gastrocnemius
muscle tissue of rats (Fig. 3B).
In the control group, the gastrocnemius muscle tissue was tight,
the muscle fibers were arranged in an orderly manner, the
mitochondrial structure was complete and clear, and the cristae
were visible. In the model group, most muscle fibers appeared
broken, the number of mitochondria was notably reduced, and the
mitochondrial structure was incomplete. In the L-ASIV group, the
muscle fibers of rats appeared severely broken, some mitochondria
were swollen, vacuolated and showed aggregation, but the number of
mitochondria was higher than that in the model group. In the H-ASIV
group, fewer broken muscle fibers were observed compared with in
the model group, with visible cristae and complete mitochondrial
structure. In addition, autophagosomes in the gastrocnemius muscle
tissue were observed using TEM. In the model group, no
autophagosomes were observed under TEM, whereas AS-IV treatment
markedly increased the number of autophagosomes (Fig. 3B). Briefly, these results indicated
that AS-IV notably improved the morphology of the gastrocnemius
muscle, the ultrastructure of mitochondria and the dysregulated
mitophagy in rats.
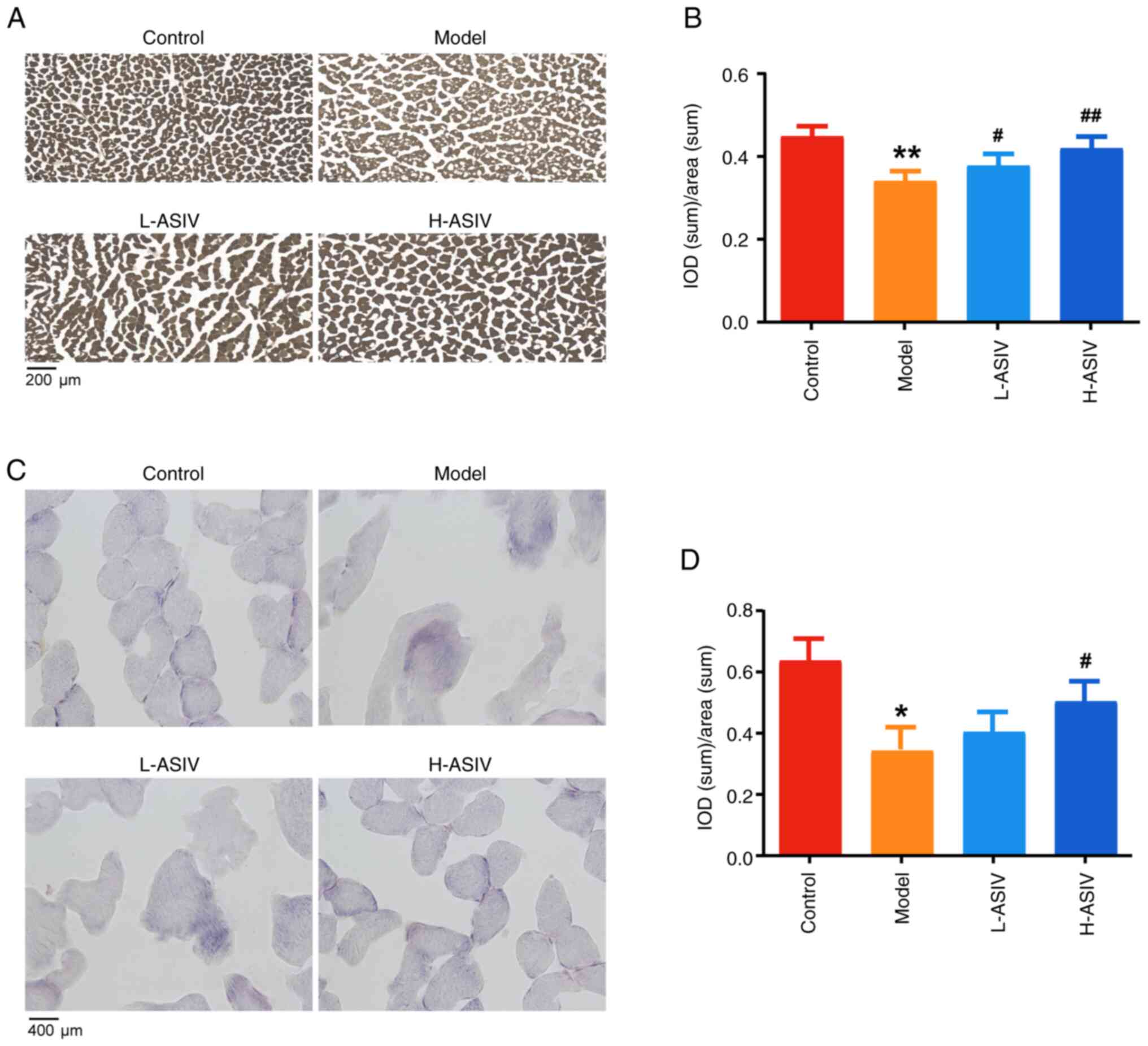
Effects of AS-IV on ATPase activity in
gastrocnemius muscle
ATPase hydrolyzes ATP into ADP and phosphate, and
releases energy for almost all essential cellular processes
(49). In the gastrocnemius muscle
of control rats, active site of ATPase staining is brown and black,
and positive expression of ATPase was detected (Fig. 4A and B). In the model group, the
intensity of staining was low and ATPase expression was
significantly reduced compared with that in the control group,
suggesting that ATPase activity was decreased (P<0.01). After
treatment with AS-IV, the intensity of staining was higher,
expression increased and ATPase activity was significantly
increased in H-ASIV and L-ASIV groups compared with that in the
model group (P<0.01 and P<0.05, respectively; Fig. 4A and B). In summary, these results
suggested that AS-IV can improve ATPase activity and restore
mitochondrial function.
Effect of AS-IV on COX activity in the
skeletal muscle of EAMG rats
COX is an enzyme present in mitochondria, which
serves a crucial role in maintaining the normal structure and
function of mitochondria (50). To
evaluate COX activity, an optical microscope was used to observe
the differences in COX staining in the gastrocnemius muscle tissues
after intragastric treatment for 4 weeks. COX staining of frozen
sections of gastrocnemius tissue in the control group showed that
the myofibrils were polygonal and of equal size; the color at
active part of COX staining was indicated by brown grain
precipitation. Compared with in the control group, COX activity in
the model group was lower with lighter brown grain precipitation in
most positively stained areas, and irregularities in the size and
shape of myofibrils was observed (Fig.
4C). In the L-ASIV group, COX activity was low and the
positively stained area showed a lighter brown grain precipitation,
accompanied with the myofibrils exhibited irregular shapes. There
was a notable difference in the H-ASIV group when compared with the
model group, which presented regularly shaped myofibrils and more
intense staining (Fig. 4C).
Compared with in the control group, the intensity of distribution
(IOD)/area of COX activity in the model group was significantly
reduced (P<0.05; Fig. 4D). By
contrast, compared with in the model group, in the H-ASIV group,
the IOD/area of COX activity was significantly increased
(P<0.05; Fig. 4D). Therefore,
these results suggested that AS-IV improved COX activity to
preserve mitochondrial structure and function.
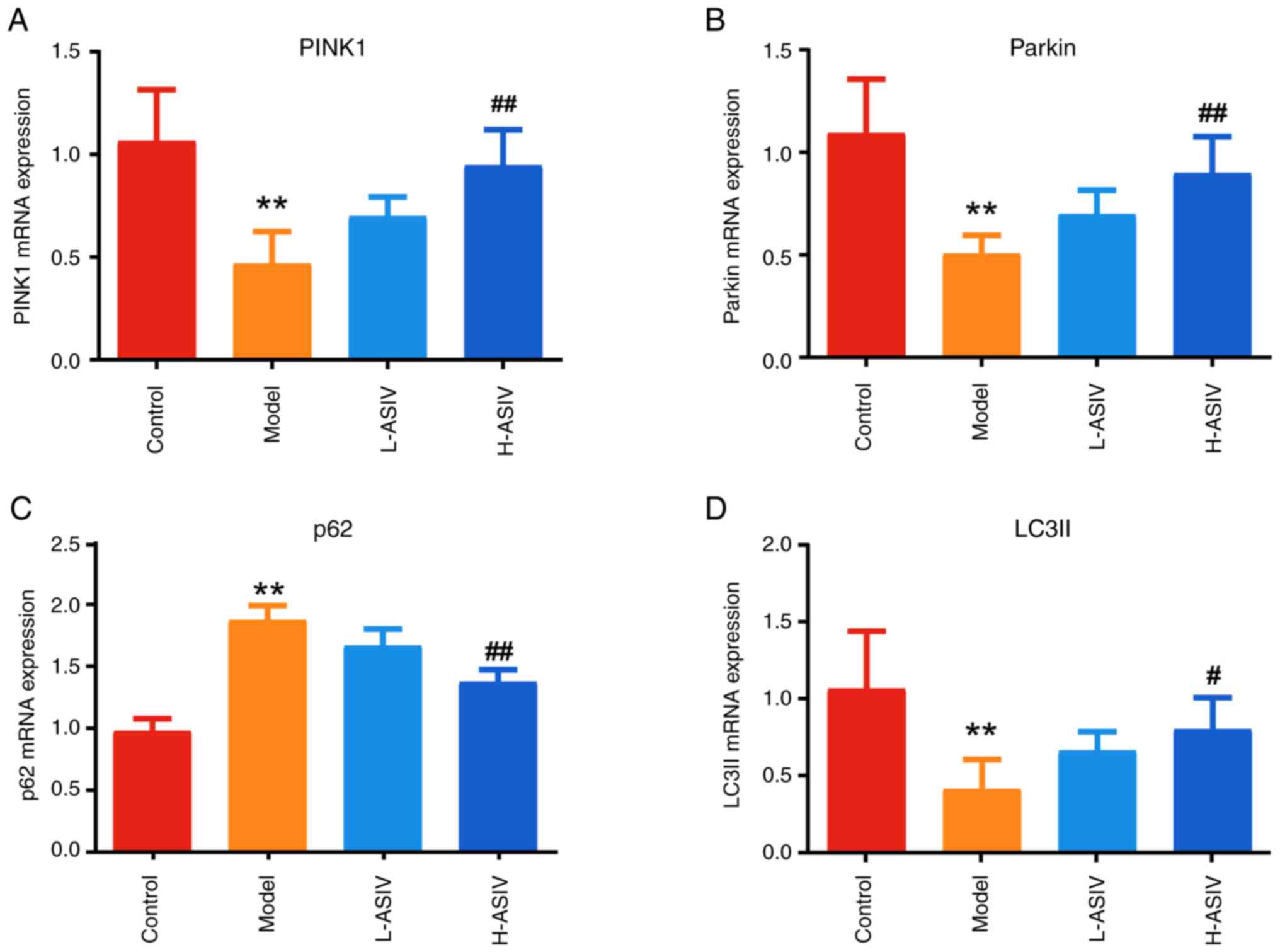
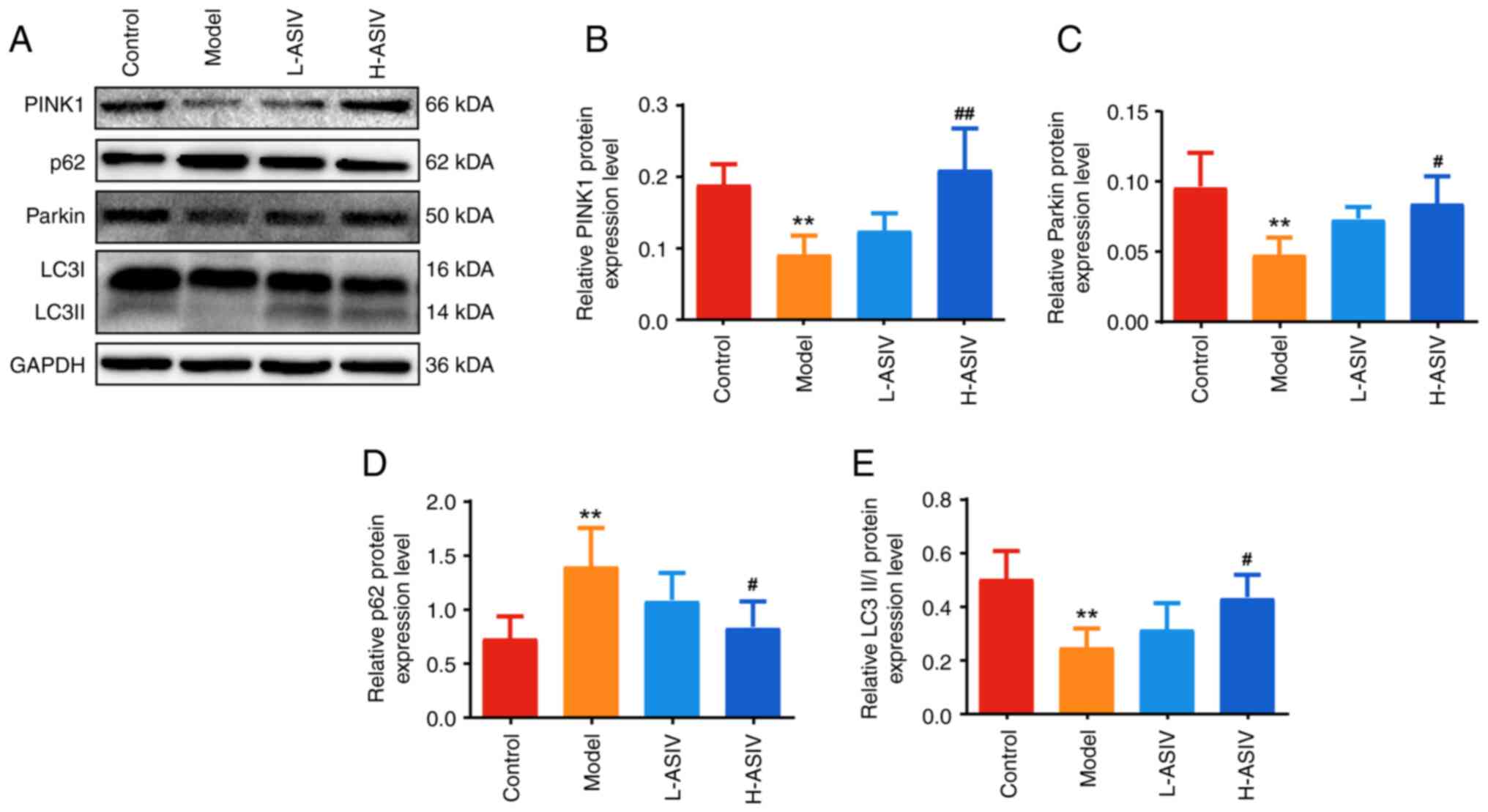
AS-IV promotes PINK1/Parkin-mediated
mitophagy in the skeletal muscle of EAMG rats
To further understand the mechanism of action of
AS-IV, its effects on the mRNA and protein expression levels of
members of the mitophagy-related signaling pathway were determined.
As shown in Fig. 5A-D, compared
with those in the control group, the mRNA expression levels of
PINK1, Parkin and LC3II in the model group were decreased, whereas
the mRNA expression levels of p62 were increased (all P<0.01).
Compared with in the model group, the difference in the mRNA
expression levels of PINK1, Parkin, p62 and LC3II in the L-ASIV
group was not significant, whereas the mRNA expression levels of
PINK1, Parkin and LC3II were significantly increased in the H-ASIV
group (P<0.05 or P<0.01). Additionally, the mRNA expression
levels of p62 were significantly decreased in the H-ASIV group
(P<0.01).
The protein expression levels of the aforementioned
molecules were further assessed using western blotting following
AS-IV treatment. As shown in Fig.
6A-E, the protein expression levels of PINK1, Parkin, and
LC3II/I in the model group were significantly lower than those in
the control group, whereas the protein expression levels of p62
were significantly increased (all P<0.01). After treatment with
AS-IV, compared with those in the model group, the protein
expression levels of PINK1, Parkin, and LC3II/I in the H-ASIV group
were significantly higher, and the expression levels of p62 were
significantly decreased (P<0.05 or P<0.01). By contrast,
there was no significant difference in the protein expression
levels in the L-ASIV group. Taken together, these results suggested
that AS-IV can promote PINK1/Parkin-mediated mitophagy in the
skeletal muscle of EAMG rats.
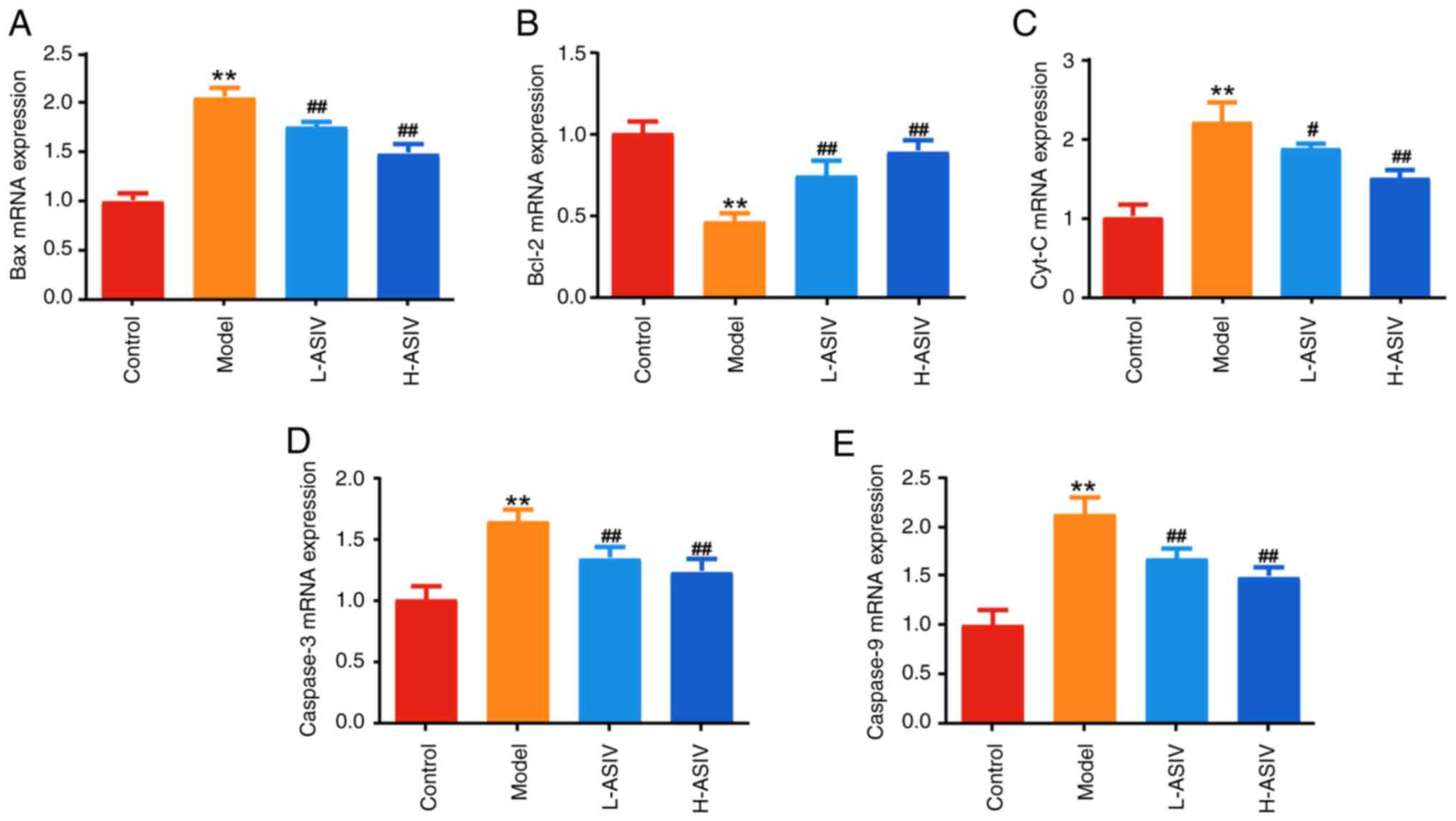
AS-IV ameliorate expression of
mitochondrial apoptosis- associated proteins/genes in the skeletal
muscle of EAMG rats
Subsequently, we conducted further investigations to
observe the impact of AS-IV on the mRNA and protein expression
levels of mitochondrial apoptosis-related signaling pathway
components. As shown in Fig. 7A-E,
the mRNA expression levels of Bax, Cyt-C, caspase 3 and caspase 9
were significantly higher (P<0.01), and the mRNA expression
levels of Bcl-2 were significantly lower (P<0.01), in the
skeletal muscle tissues of the model group compared with in the
control group. After 4 weeks of AS-IV treatment, the expression
levels of Cyt-C, Bax, caspase 3 and caspase 9 were significantly
lower in the L-ASIV and H-ASIV groups than those in the model group
(all P<0.01; except for Cyt-C in the L-ASIV group, P<0.05).
By contrast, Bcl-2 expression in the H-ASIV and L-ASIV groups was
significantly higher than that in the model group (P<0.01).
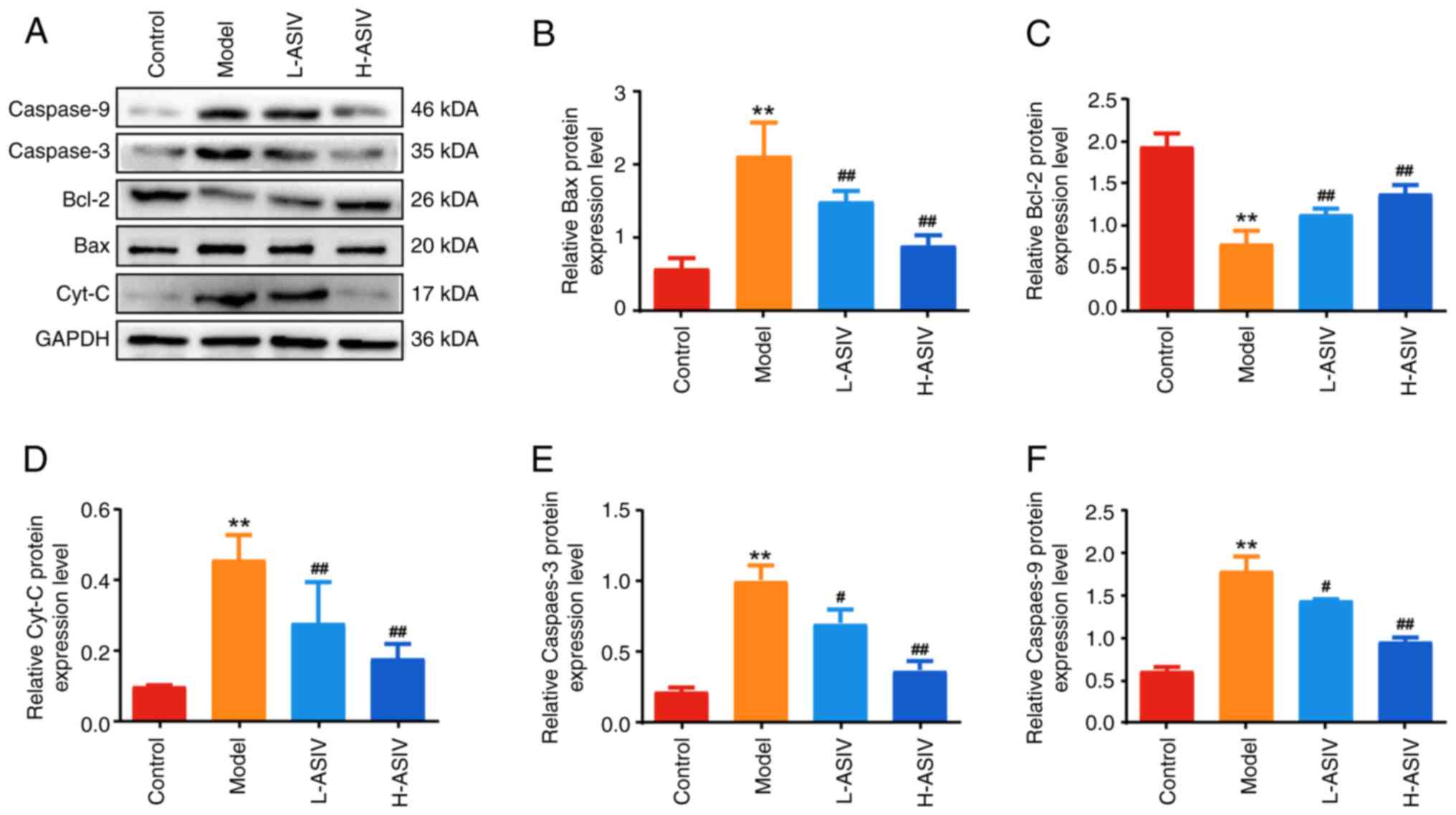
Western blotting was used to detect the protein
expression levels of the aforementioned genes. As shown in Fig. 8A-F, the protein expression levels
of Cyt-C, Bax, caspase 3 and caspase 9 were significantly higher
(all P<0.01), and the expression levels of Bcl-2 were
significantly lower (P<0.01) in the skeletal muscle of the model
group compared with those in the control group. After 4 weeks of
AS-IV treatment, the expression levels of Cyt-C, Bax, caspase 3 and
caspase 9 in the H-ASIV and L-ASIV groups were significantly lower
than those in the model group (all P<0.01; except for caspase 3
and 9 in the L-ASIV group, P<0.05). In addition, the protein
expression levels of Bcl-2 in the H-ASIV and L-ASIV groups were
significantly higher than those in the model group (P<0.01). In
summary, these results indicated that AS-IV can ameliorate the
expression of mitochondrial apoptosis-associated proteins/genes in
the skeletal muscle of EAMG rats.
Discussion
AS-IV has been shown to exert pharmacological
effects, such as antioxidant, anti-inflammatory, anti-apoptotic and
immune-enhancing activities. Previous studies have also confirmed
that AS-IV alleviates neuron and Schwann cell damage and improves
neural functional deficits (51–53),
which significantly improves neurodegenerative diseases, such as
Alzheimer's disease (AD) and PD (54). Moreover, AS-IV has demonstrated
favorable therapeutic effects on neuromuscular disorders, such as
amyotrophic lateral sclerosis (ALS), which has been attributed to
its ability to ameliorate inflammatory responses and
neurotransmission (29,34,55,56).
Since MG is a representative neuromuscular disease (57), it is reasonable to speculate that
AS-IV may offer therapeutic effects in its management. In the
present study, an EAMG rat model was established, and a reduction
in body weight and muscle strength was observed, as well as
impaired neurotransmission and a significant decrease in the action
potential. Furthermore, the structure of the gastrocnemius muscle
was disturbed, and muscle fibers appeared broken based on
microscopic observation with H&E staining. In the rats treated
with AS-IV, the aforementioned observations were reversed. AS-IV
could improve symptoms of muscle weakness and pathological changes
in the gastrocnemius muscle and could improve the attenuation of
the action potential of the gastrocnemius muscle and restore
neurotransmission, thus indicating that AS-IV may exert a
protective effect on EAMG rats.
The primary autoimmune antibody associated with MG
onset is AChR-Ab, which targets the postsynaptic membrane of AChR,
leading to disrupted ACh transmission and impaired NMJ function,
resulting in symptoms of muscle weakness (58). The current treatment approach for
MG primarily involves the use of acetylcholinesterase inhibitors,
corticosteroids and immunosuppressants, which can alleviate
symptoms in the majority of patients with MG (59). However, ~15% of patients with MG
have been reported to show limited or no response to these
therapies (60), while
simultaneously enduring the burden of numerous adverse reactions
(61,62). AS-IV, a natural medicinal compound
derived from A. membranaceus, is a traditional Chinese
medicine that presents advantages, such as cost-effectiveness and
low toxicity (63,64). In the present study, the levels of
AChR-Ab were increased in EAMG rats in response to modeling, but
were reversed after AS-IV treatment, suggesting the potential for
AS-IV to regulate AChR-Ab levels in EAMG rats. Nevertheless, ~20%
of patients with MG do not exhibit AChR-Ab in their serum (65), and the severity of symptoms is not
correlated with antibody levels (66–68),
which presents challenges in diagnosis, treatment selection and
evaluation of prognosis. The present findings indicated that AS-IV
not only reduced serum AChR-Ab levels in EAMG rats, but also
significantly improved the morphology and function of skeletal
muscle mitochondria. It may be hypothesized that mitochondrial
damage serves a role in MG, and AS-IV could exert its therapeutic
effects by improving mitochondrial function.
The NMJ is a highly active site that requires a
significant amount of energy to sustain the transmission of nerve
impulses and muscle contraction (17). Mitochondria have a vital role in
providing energy for these processes through the production of ATP
(69). Furthermore, mitochondria
serve a role in antioxidant defense and in the regulation of
calcium ions (18). Certain
proteins within mitochondria can also promote the aggregation of
AChRs at the postsynaptic membrane (70). In summary, the proper functioning
of mitochondria ensures the coordinated progression of NMJ activity
and skeletal muscle physiology.
The pathological process of MG may be significantly
influenced by mitochondrial dysfunction. Previous studies have
shown mitochondrial accumulation and atypical abnormal
ultrastructure in muscle biopsies obtained from patients with MG
(15). Additionally, there is a
deficiency of mitochondrial respiratory chain complexes in patients
with MG (71). Mitochondrial
dysfunction and depleted energy levels have also been observed in
the muscle tissue of patients with MG (16). The impairment of mitochondria can
diminish muscle energy production and trigger oxidative stress,
further exacerbating symptoms of muscle weakness (72). Consequently, reversing
mitochondrial damage, and restoring the normal structure and
function of mitochondria, are crucial for effective MG treatment.
In the present study, after modeling, compared with in the control
group, the model group rats demonstrated a marked decrease in the
number of mitochondria in the gastrocnemius muscle, along with
incomplete mitochondrial structure, swelling and vacuolization, and
there was a lack of cristae based on TEM. Subsequent administration
of AS-IV at low or high doses led to an increase in mitochondrial
quantity and pronounced improvement in mitochondrial structure,
particularly in the high-dose group, where well-defined cristae and
relatively intact mitochondrial structure were evident. These
findings suggested that AS-IV may exert a protective effect against
mitochondrial damage in EAMG.
The activity of ATPase can reflect the function of
mitochondria. ATPase hydrolyzes ATP into ADP and phosphate, and
releases energy for almost all essential cellular processes
(73). In our previous study,
myocyte damage alongside decreased
Ca2+/Mg2+-ATP and
Na+/K+-ATP enzyme activity were observed in
the skeletal muscle of EAMG rats, indicative of abnormal energy
metabolism in the skeletal muscle of EAMG rats, resulting in
impaired muscle diastolic movement (19). COX, also known as mitochondrial
respiratory chain complex IV, is one of the oxidases essential for
mitochondrial respiratory function (74). Previous studies have shown that
defects in the mitochondrial respiratory chain are important
factors that interfere with cellular energy metabolism and are an
important cause of skeletal muscle injury (75–77).
In the present study, by staining frozen sections of the rat
gastrocnemius muscle, the results showed that the model group had a
reduced staining and markedly lower activity in the positively
stained ATPase area, low COX activity, and a lower number of cells
positive for both ATPase and COX activity, all of which were
restored after treatment with AS-IV. These results further
confirmed that AS-IV could reduce skeletal muscle injury in EAMG
rats by improving mitochondrial energy metabolism.
Mitophagy is a process that selectively removes
damaged or abnormal mitochondria and serves a crucial role in
maintaining intracellular homeostasis (78). PINK1/Parkin is the classical
pathway mediating mitophagy. PINK1 acts as a mitochondrial Ser/Thr
kinase that recruits Parkin to depolarized mitochondria and
interacts with the outer mitochondrial membrane complex to regulate
Parkin translocation and activation. Parkin acts as an E3 ubiquitin
ligase and is typically located in the cytoplasm. In response to
mitochondrial stress, it is rapidly recruited to damaged
mitochondria, thereby phosphorylating the ubiquitin ligase p62 and
promoting its binding to LC3 in the mitochondria to initiate
mitophagy (79).
Apoptosis serves an important role in the regulation
of homeostasis in the human body, and mitochondrial apoptosis is
one of the major routes of apoptosis, involving a variety of key
signaling molecules (80).
Apoptosis-promoting Bax protein and apoptosis-inhibiting Bcl-2
protein, primarily localized on the mitochondria and endoplasmic
reticulum, jointly regulate mitochondrial membrane permeability and
apoptosis signal transmission. When cells are stimulated by
apoptotic signals, Bax proteins excessively accumulate in the outer
mitochondrial membrane, enhancing mitochondrial membrane
permeability. This leads to the release of soluble proteins such as
Cyt-C from the intermembrane space to the cytoplasm. Subsequently,
Cyt-C can recruit and activate proenzymes of caspase 9, initiating
the amplification of the caspase cascade effect, and further
activating the cleavage of caspase 3 precursors, ultimately leading
to the initiation of apoptosis (81,82).
A growing number of studies have confirmed the close
association between mitophagy and the development of progressive
myasthenic diseases, such as neurogenic distal myopathy and ALS
(83,84). Similarly, mitochondrial apoptosis
has a key role in several neurodegenerative and autoimmune
diseases, such as AD, PD and rheumatoid arthritis (RA) (83–86).
AS-IV has been reported to prevent dopaminergic neurodegeneration
in PD by promoting mitophagy to inhibit astrocyte senescence
(87). AS-IV also attenuates
Schwann cell injury in diabetic peripheral neuropathy by promoting
autophagy (88). In addition,
AS-IV attenuates neuroinflammation and delays smooth muscle cell
senescence in spinal cord injury by promoting autophagy (89,90).
AS-IV has also been shown to exert its anti-apoptotic effects in
several diseases via the regulation of key mitochondrial apoptotic
signaling molecules, such as Bax/Bcl-2 and the caspase family of
proteins (91–94). However, the involvement of
mitophagy and apoptosis in the development of MG and the potential
of AS-IV to mitigate skeletal muscle injury in MG through the
regulation of mitophagy and apoptosis remains unclear.
In the present study, the results showed there was a
decrease in the mRNA and protein expression levels of PINK1 and
Parkin, as well as an increase in the expression of p62 in the
skeletal muscle tissues of the EAMG rat model. The expression of
LC3, as a marker of autophagy, holds significant importance.
Additionally, the ratio of LC3II/LC3I is often used to measure the
degree of autophagy (95). A
significant reduction in LC3II/I was observed in the skeletal
muscle of EAMG rats, along with a decrease in the number of
autophagosomes observed by TEM in the model group, indicating a
marked attenuation of mitophagy in the skeletal muscle of EAMG
rats. Furthermore, the mRNA and protein expression levels of
pro-apoptosis-related signaling molecules, such as Cyt-C, Bax,
caspase 3 and caspase 9, were increased, whereas the expression
levels of the apoptosis-suppressing protein Bcl-2 were decreased,
leading to an elevated level of mitochondrial apoptosis. All of the
aforementioned changes were reversed by AS-IV treatment. Therefore,
these findings indicated that mitophagy was reduced and
mitochondrial apoptosis was elevated in the skeletal muscle of EAMG
rats, resulting in skeletal muscle cell damage. Conversely, AS-IV
improved the morphology, structure and function of mitochondria by
promoting mitophagy and inhibiting mitochondrial apoptosis. This
effectively reduced the pathological damage to skeletal muscle and
improved the symptoms in EAMG rats.
However, the present study has some limitations. In
the present study, AS-IV could decrease the levels of AChR-Ab and
alleviate aberrant mitophagy and apoptosis. However, the potential
link between mitophagy, apoptosis and AChR-Ab remains unclear;
investigating this association presents an intriguing question, and
further exploration of this topic will be pursued in future
research.
In conclusion, to the best of our knowledge, the
present study was the first to demonstrate that AS-IV protected
against EAMG in a rat model by modulating mitophagy and apoptosis.
This resulted in an improvement in mitochondrial structure and
function, as well as a reduction in gastrocnemius muscle damage.
These findings provide novel insights into the potential mechanism
of MG and highlight potential novel treatment strategies.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Natural Science
Foundation of Guangdong Province (grant no. 2023A1515011127), the
Project in Key Fields of Universities in Guangdong Province (grant
no 2021ZDZX2032) and the National Natural Science Foundation of
China (grant no. 82374391).
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
JZ, JH, JL, QL and LK performed experiments. JZ, JH
and JL wrote the manuscript. QL, LK, QJ, YL, HZha, HZho and PY
contributed to analysis and interpretation of data. HZho and PY
provided critical comments on the revision of the manuscript. YS
and TC contributed to the conception, design and supervision of the
study. YS provided funding. JZ, QL and LK confirm the authenticity
of all the raw data. All authors have read and approved the final
version of the manuscript.
Ethics approval and consent to
participate
All experimental procedures were approved by the
Animal Ethics Committee of Guangzhou University of Traditional
Chinese Medicine (Guangzhou, China; approval no. A202005017).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
AS-IV
|
astragaloside IV
|
|
L-ASIV
|
low-dose AS-IV
|
|
H-ASIV
|
high-dose AS-IV
|
|
MG
|
myasthenia gravis
|
|
EAMG
|
experimental autoimmune MG
|
|
AChR
|
acetylcholine receptor
|
|
Cyt-C
|
cytochrome c
|
|
COX
|
Cyt-C oxidase
|
|
PINK1
|
phosphatase and tensin homolog-induced
putative kinase 1
|
|
CFA
|
complete Freund's adjuvant
|
|
IFA
|
incomplete Freund's adjuvant
|
References
|
1
|
Payet CA, You A, Fayet OM, Dragin N,
Berrih-Aknin S and Le Panse R: Myasthenia gravis: An acquired
interferonopathy? Cells. 11:12182022. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gilhus NE, Tzartos S, Evoli A, Palace J,
Burns TM and Verschuuren J: Myasthenia gravis. Nat Rev Dis Primers.
5:302019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Huijbers MG, Marx A, Plomp JJ, Le Panse R
and Phillips WD: Advances in the understanding of disease
mechanisms of autoimmune neuromuscular junction disorders. Lancet
Neurol. 21:163–175. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cortés-Vicente E, Álvarez-Velasco R,
Segovia S, Paradas C, Casasnovas C, Guerrero-Sola A, Pardo J,
Ramos-Fransi A, Sevilla T, López de Munain A, et al: Clinical and
therapeutic features of myasthenia gravis in adults based on age at
onset. Neurology. 94:e1171–e1180. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gilhus NE and Verschuuren JJ: Myasthenia
gravis: Subgroup classification and therapeutic strategies. Lancet
Neurol. 14:1023–1036. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Punga AR, Maddison P, Heckmann JM, Guptill
JT and Evoli A: Epidemiology, diagnostics, and biomarkers of
autoimmune neuromuscular junction disorders. Lancet Neurol.
21:176–188. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
García Estévez DA and Pardo Fernández J:
Myasthenia gravis. Update on diagnosis and therapy. Med Clin
(Barc). 161:119–127. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mahic M, Bozorg A, DeCourcy J, Golden K,
Gibson G, Taylor C and Scowcroft A: Physician- and patient-reported
perspectives on myasthenia gravis in Europe: A real-world survey.
Orphanet J Rare Dis. 18:1692023. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Petersson M, Feresiadou A, Jons D, Ilinca
A, Lundin F, Johansson R, Budzianowska A, Roos AK, Kågström V,
Gunnarsson M, et al: Patient-Reported symptom severity in a
nationwide myasthenia gravis cohort: Cross-sectional analysis of
the swedish GEMG study. Neurology. 97:e1382–1391. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Verschuuren JJ, Palace J, Murai H,
Tannemaat MR, Kaminski HJ and Bril V: Advances and ongoing research
in the treatment of autoimmune neuromuscular junction disorders.
Lancet Neurol. 21:189–202. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Walker BR and Moraes CT:
Nuclear-mitochondrial interactions. Biomolecules. 12:4272022.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
de Beauchamp L, Himonas E and Helgason GV:
Mitochondrial metabolism as a potential therapeutic target in
myeloid leukaemia. Leukemia. 36:1–12. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xiang L, Shao Y and Chen Y: Mitochondrial
dysfunction and mitochondrion-targeted therapeutics in liver
diseases. J Drug Target. 29:1080–1093. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yang X, Xue P, Yuan M, Xu X, Wang C, Li W,
Machens HG and Chen Z: SESN2 protects against denervated muscle
atrophy through unfolded protein response and mitophagy. Cell Death
Dis. 12:8052021. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Martignago S, Fanin M, Albertini E,
Pegoraro E and Angelini C: Muscle histopathology in myasthenia
gravis with antibodies against MuSK and AChR. Neuropathol Appl
Neurobiol. 35:103–110. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shichijo K, Mitsui T, Kunishige M, Kuroda
Y, Masuda K and Matsumoto T: Involvement of mitochondria in
myasthenia gravis complicated with dermatomyositis and rheumatoid
arthritis: A case report. Acta Neuropathol. 109:539–542. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sousa-Soares C, Noronha-Matos JB and
Correia-de-Sá P: Purinergic tuning of the tripartite neuromuscular
synapse. Mol Neurobiol. 60:4084–4104. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ferrari R, Rodrigues-Simioni L and da Cruz
Höfling MA: Guanidine affects differentially the twitch response of
diaphragm, extensor digitorum longus and soleus nerve-muscle
preparations of mice. Molecules. 17:7503–7522. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jiao W, Hu F, Li J, Song J, Liang J, Li L,
Song Y, Chen Z, Li Q and Ke L: Qiangji Jianli Decoction promotes
mitochondrial biogenesis in skeletal muscle of myasthenia gravis
rats via AMPK/PGC-1α signaling pathway. Biomed Pharmacother.
129:1104822020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ke L, Li Q, Song J, Jiao W, Ji A, Chen T,
Pan H and Song Y: The mitochondrial biogenesis signaling pathway is
a potential therapeutic target for myasthenia gravis via energy
metabolism (Review). Exp Ther Med. 22:7022021. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li L, Cai D, Zhong H, Liu F, Jiang Q,
Liang J, Li P, Song Y, Ji A, Jiao W, et al: Mitochondrial dynamics
and biogenesis indicators may serve as potential biomarkers for
diagnosis of myasthenia gravis. Exp Ther Med. 23:3072022.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Song J, Lei X, Jiao W, Song Y, Chen W, Li
J and Chen Z: Effect of Qiangji Jianli decoction on mitochondrial
respiratory chain activity and expression of mitochondrial fusion
and fission proteins in myasthenia gravis rats. Sci Rep.
8:86232018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zheng Z, Guo C, Li M, Yang L, Liu P, Zhang
X, Liu Y, Guo X, Cao S, Dong Y, et al: Hypothalamus-habenula
potentiation encodes chronic stress experience and drives
depression onset. Neuron. 110:1400–1415.e6. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Su L, Zhang J, Gomez H, Kellum JA and Peng
Z: Mitochondria ROS and mitophagy in acute kidney injury.
Autophagy. 19:401–414. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gupta R, Ambasta RK and Pravir K:
Autophagy and apoptosis cascade: Which is more prominent in
neuronal death? Cell Mol Life Sci. 78:8001–8047. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lai Y, Xu X, Zhu Z and Hua Z: Highly
efficient siRNA transfection in macrophages using apoptotic
body-mimic Ca-PS lipopolyplex. Int J Nanomedicine. 13:6603–6623.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang J, Ma G, Guo Z, Yu Q, Han L, Han M
and Zhu Y: Study on the apoptosis mediated by
apoptosis-inducing-factor and influencing factors of bovine muscle
during postmortem aging. Food Chem. 266:359–367. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Piras A, Schiaffino L, Boido M, Valsecchi
V, Guglielmotto M, De Amicis E, Puyal J, Garcera A, Tamagno E,
Soler RM and Vercelli A: Inhibition of autophagy delays motoneuron
degeneration and extends lifespan in a mouse model of spinal
muscular atrophy. Cell Death Dis. 8:32232017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kuno A, Hosoda R, Sebori R, Hayashi T,
Sakuragi H, Tanabe M and Horio Y: Resveratrol ameliorates mitophagy
disturbance and improves cardiac pathophysiology of
Dystrophin-deficient mdx Mice. Sci Rep. 8:155552018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
De Palma C, Morisi F, Cheli S, Pambianco
S, Cappello V, Vezzoli M, Rovere-Querini P, Moggio M, Ripolone M,
Francolini M, et al: Autophagy as a new therapeutic target in
Duchenne muscular dystrophy. Cell Death Dis. 3:e4182012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bloemberg D and Quadrilatero J: Autophagy,
apoptosis, and mitochondria: Molecular integration and
physiological relevance in skeletal muscle. Am J Physiol Cell
Physiol. 317:C111–C130. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jing H, Xie R, Bai Y, Duan Y, Sun C, Wang
Y, Cao R, Ling Z and Qu X: The mechanism actions of astragaloside
IV prevents the progression of hypertensive heart disease based on
network pharmacology and experimental pharmacology. Front
Pharmacol. 12:7556532021. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Song J, Li Q, Ke L, Liang J, Jiao W, Pan
H, Li Y, Du Q, Song Y, Ji A, et al: Qiangji jianli decoction
alleviates hydrogen peroxide-induced mitochondrial dysfunction via
regulating mitochondrial dynamics and biogenesis in L6 myoblasts.
Oxid Med Cell Longev. 2021:66606162021. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Costa IM, Lima FOV, Fernandes LCB, Norrara
B, Neta FI, Alves RD, Cavalcanti JRLP, Lucena EES, Cavalcante JS,
Rego ACM, et al: Astragaloside IV supplementation promotes A
neuroprotective effect in experimental models of neurological
disorders: A systematic review. Curr Neuropharmacol. 17:648–665.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhu T, Wang L, Wang LP and Wan Q:
Therapeutic targets of neuroprotection and neurorestoration in
ischemic stroke: Applications for natural compounds from medicinal
herbs. Biomed Pharmacother. 148:1127192022. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhong Y, Liu W, Xiong Y, Li Y, Wan Q, Zhou
W, Zhao H, Xiao Q and Liu D: Astragaloside IV alleviates ulcerative
colitis by regulating the balance of Th17/Treg cells.
Phytomedicine. 104:1542872022. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bolduc JA, Collins JA and Loeser RF:
Reactive oxygen species, aging and articular cartilage homeostasis.
Free Radic Biol Med. 132:73–82. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jiang B, Yang YJ, Dang WZ, Li H, Feng GZ,
Yu XC, Shen XY and Hu XG: Astragaloside IV reverses
simvastatin-induced skeletal muscle injury by activating the
AMPK-PGC-1α signalling pathway. Phytother Res. 34:1175–1184. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
National Research Council (US) Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals, . Guide for the care and use of laboratory animals. 8th
edition. Washington (DC): National Academies Press (US); 2011
|
|
40
|
Baggi F, Annoni A, Ubiali F, Longhi R,
Milani M, Mantegazza R, Cornelio F and Antozzi C: Immunization with
rat-, but not Torpedo-derived 97–116 peptide of the AChR
alpha-subunit induces experimental myasthenia gravis in Lewis rat.
Ann N Y Acad Sci. 998:391–394. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
He X, Zhou S, Ji Y, Zhang Y, Lv J, Quan S,
Zhang J, Zhao X, Cui W, Li W, et al: Sorting nexin 17 increases
low-density lipoprotein receptor-related protein 4 membrane
expression: A novel mechanism of acetylcholine receptor aggregation
in myasthenia gravis. Front Immunol. 13:9160982022. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang CC, Li H, Zhang M, Li XL, Yue LT,
Zhang P, Zhao Y, Wang S, Duan RN, Li YB and Duan RS: Caspase-1
inhibitor ameliorates experimental autoimmune myasthenia gravis by
innate dendric cell IL-1-IL-17 pathway. J Neuroinflammation.
12:1182015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Claussen GC, Fesenmeier JT, Hah JS, Brooks
J and Oh SJ: The accessory nerve repetitive nerve stimulation test:
A valuable second-line test in myasthenia gravis. Eur J Neurol.
2:492–497. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Nie Q, Zhu L, Zhang L, Leng B and Wang H:
Astragaloside IV protects against hyperglycemia-induced vascular
endothelial dysfunction by inhibiting oxidative stress and
Calpain-1 activation. Life Sci. 232:1166622019. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Liu YL, Zhang QZ, Wang YR, Fu LN, Han JS,
Zhang J and Wang BM: Astragaloside IV improves High-Fat
Diet-Induced hepatic steatosis in nonalcoholic fatty liver disease
rats by regulating inflammatory factors level via TLR4/NF-κB
signaling pathway. Front Pharmacol. 11:6050642020. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yang J, Wang HX, Zhang YJ, Yang YH, Lu ML,
Zhang J, Li ST, Zhang SP and Li G: Astragaloside IV attenuates
inflammatory cytokines by inhibiting TLR4/NF-кB signaling pathway
in isoproterenol-induced myocardial hypertrophy. J Ethnopharmacol.
150:1062–1070. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Laferriere CA and Pang DS: Review of
intraperitoneal injection of sodium pentobarbital as a method of
euthanasia in laboratory rodents. J Am Assoc Lab Anim Sci.
59:254–263. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
van der Bliek AM, Sedensky MM and Morgan
PG: Cell biology of the mitochondrion. Genetics. 207:843–871. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Mansilla N, Racca S, Gras DE, Gonzalez DH
and Welchen E: The complexity of mitochondrial Complex IV: An
update of cytochrome c oxidase biogenesis in plants. Int J Mol Sci.
19:6622018. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Ni GX, Liang C, Wang J, Duan CQ, Wang P
and Wang YL: Astragaloside IV improves neurobehavior and promotes
hippocampal neurogenesis in MCAO rats though BDNF-TrkB signaling
pathway. Biomed Pharmacother. 130:1103532020. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Shi YH, Zhang XL, Ying PJ, Wu ZQ, Lin LL,
Chen W, Zheng GQ and Zhu WZ: Neuroprotective effect of
Astragaloside IV on cerebral Ischemia/Reperfusion injury rats
through Sirt1/Mapt pathway. Front Pharmacol. 12:6398982021.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Yin Y, Qu H, Yang Q, Fang Z and Gao R:
Astragaloside IV alleviates Schwann cell injury in diabetic
peripheral neuropathy by regulating microRNA-155-mediated
autophagy. Phytomedicine. 92:1537492021. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kuo YC, Chen IY and Rajesh R:
Astragaloside IV- and nesfatin-1-encapsulated phosphatidylserine
liposomes conjugated with wheat germ agglutinin and leptin to
activate anti-apoptotic pathway and block phosphorylated tau
protein expression for Parkinson's disease treatment. Mater Sci Eng
C Mater Biol Appl. 129:1123612021. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Tian Y, Jin S, Promes V, Liu X and Zhang
Y: Astragaloside IV and echinacoside benefit neuronal properties
via direct effects and through upregulation of SOD1 astrocyte
function in vitro. Naunyn Schmiedebergs Arch Pharmacol.
394:1019–1029. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Smith EF, Shaw PJ and De Vos KJ: The role
of mitochondria in amyotrophic lateral sclerosis. Neurosci Lett.
710:1329332019. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Attia M, Maurer M, Robinet M, Le Grand F,
Fadel E, Le Panse R, Butler-Browne G and Berrih-Aknin S: Muscle
satellite cells are functionally impaired in myasthenia gravis:
Consequences on muscle regeneration. Acta Neuropathol. 134:869–888.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Truffault F, de Montpreville V, Eymard B,
Sharshar T, Le Panse R and Berrih-Aknin S: Thymic germinal centers
and corticosteroids in myasthenia gravis: An immunopathological
study in 1035 cases and a critical review. Clin Rev Allergy
Immunol. 52:108–124. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Menon D and Bril V: Pharmacotherapy of
generalized myasthenia gravis with special emphasis on newer
biologicals. Drugs. 82:865–887. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Mantegazza R and Antozzi C: When
myasthenia gravis is deemed refractory: Clinical signposts and
treatment strategies. Ther Adv Neurol Disord.
11:17562856177491342018. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Lorenzoni PJ, Kay CSK, Zanlorenzi MF,
Ducci RD, Werneck LC and Scola RH: Myasthenia gravis and
azathioprine treatment: Adverse events related to thiopurine
S-methyl-transferase (TPMT) polymorphisms. J Neurol Sci.
412:1167342020. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Tao X, Wang W, Jing F, Wang Z, Chen Y, Wei
D and Huang X: Long-term efficacy and side effects of low-dose
tacrolimus for the treatment of Myasthenia Gravis. Neurol Sci.
38:325–330. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Stępnik K, Kukula-Koch W, Plazinski W,
Gawel K, Gaweł-Bęben K, Khurelbat D and Boguszewska-Czubara A:
Significance of astragaloside IV from the roots of astragalus
mongholicus as an acetylcholinesterase inhibitor-from the
computational and biomimetic analyses to the in vitro and in vivo
studies of safety. Int J Mol Sci. 24:91522023. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Yuan F, Yang Y, Liu L, Zhou P, Zhu Y, Chai
Y, Chen K, Tang W, Huang Q and Zhang C: Research progress on the
mechanism of astragaloside IV in the treatment of asthma. Heliyon.
9:e221492023. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Rivner MH, Pasnoor M, Dimachkie MM, Barohn
RJ and Mei L: Muscle-Specific tyrosine kinase and myasthenia gravis
owing to other antibodies. Neurol Clin. 36:293–310. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Howard FM Jr, Lennon VA, Finley J,
Matsumoto J and Elveback LR: Clinical correlations of antibodies
that bind, block, or modulate human acetylcholine receptors in
myasthenia gravis. Ann N Y Acad Sci. 505:526–538. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Lindstrom JM, Seybold ME, Lennon VA,
Whittingham S and Duane DD: Antibody to acetylcholine receptor in
myasthenia gravis. Prevalence, clinical correlates, and diagnostic
value. Neurology. 26:1054–1059. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Gilhus NE, Skeie GO, Romi F, Lazaridis K,
Zisimopoulou P and Tzartos S: Myasthenia gravis-autoantibody
characteristics and their implications for therapy. Nature Rev
Neurology. 12:259–268. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Anagnostou ME and Hepple RT: Mitochondrial
mechanisms of neuromuscular junction degeneration with aging.
Cells. 9:1972020. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Xiao Y, Zhang J, Shu X, Bai L, Xu W, Wang
A, Chen A, Tu WY, Wang J, Zhang K, et al: Loss of mitochondrial
protein CHCHD10 in skeletal muscle causes neuromuscular junction
impairment. Hum Mol Genet. 29:1784–1796. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Finsterer J, Oberman I and Reitner A:
Respiratory chain complex-I defect mimicking myasthenia. Metab
Brain Dis. 17:41–46. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Jang YC, Lustgarten MS, Liu Y, Muller FL,
Bhattacharya A, Liang H, Salmon AB, Brooks SV, Larkin L, Hayworth
CR, et al: Increased superoxide in vivo accelerates age-associated
muscle atrophy through mitochondrial dysfunction and neuromuscular
junction degeneration. FASEB. 24:1376–1390. 2010. View Article : Google Scholar
|
|
73
|
Holper L, Ben-Shachar D and Mann JJ:
Multivariate meta-analyses of mitochondrial complex I and IV in
major depressive disorder, bipolar disorder, schizophrenia,
Alzheimer disease, and Parkinson disease. Neuropsychopharmacology.
44:837–849. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Ferreira N, Andoniou CE, Perks KL, Ermer
JA, Rudler DL, Rossetti G, Periyakaruppiah A, Wong JKY, Rackham O,
Noakes PG, et al: Murine cytomegalovirus infection exacerbates
complex IV deficiency in a model of mitochondrial disease. PLoS
Genet. 16:e10086042020. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Hatakeyama H and Goto YI: Respiratory
chain complex disorganization impairs mitochondrial and cellular
integrity: Phenotypic variation in cytochrome c oxidase deficiency.
Am J Pathol. 187:110–121. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Wang XL, Feng ST, Wang ZZ, Chen NH and
Zhang Y: Role of mitophagy in mitochondrial quality control:
Mechanisms and potential implications for neurodegenerative
diseases. Pharmacol Res. 165:1054332021. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Chu CT: Mechanisms of selective autophagy
and mitophagy: Implications for neurodegenerative diseases.
Neurobiol Dis. 122:23–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Evans CS and Holzbaur ELF: Autophagy and
mitophagy in ALS. Neurobiol Dis. 122:35–40. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Carrascoso I, Sánchez-Jiménez C, Silion E,
Alcalde J and Izquierdo JM: A heterologous cell model for studying
the role of T-Cell intracellular antigen 1 in welander distal
myopathy. Mol Cell Biol. 39:e00299–18. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Wang TS, Coppens I, Saorin A, Brady NR and
Hamacher-Brady A: Endolysosomal targeting of mitochondria is
integral to BAX-Mediated mitochondrial permeabilization during
apoptosis signaling. Developmental cell. 53:627–645.e7. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Yin W, Li R, Feng X and James Kang Y: The
Involvement of cytochrome c Oxidase in mitochondrial fusion in
primary cultures of neonatal rat cardiomyocytes. Cardiovasc
Toxicol. 18:365–373. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Wang XR, Wang C and Wang XW, Qian LX, Chi
Y, Liu SS, Liu YQ and Wang XW: The functions of caspase in whitefly
Bemisia tabaci apoptosis in response to ultraviolet irradiation.
Insect Mol Biol. 27:739–751. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Huang G, Li H and Zhang H: Abnormal
expression of mitochondrial ribosomal proteins and their encoding
genes with cell apoptosis and diseases. Int J Mol Sci. 21:88792020.
View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Hazafa A, Batool A, Ahmad S, Amjad M,
Chaudhry SN, Asad J, Ghuman HF, Khan HM, Naeem M and Ghani U:
Humanin: A mitochondrial-derived peptide in the treatment of
apoptosis-related diseases. Life Sci. 264:1186792021. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Bock FJ and Tait SWG: Mitochondria as
multifaceted regulators of cell death. Nat Rev Mol Cell Biol.
21:85–100. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Zheng M, Kuang N, Zeng X, Wang J, Zou Y
and Fu Y: Daphnetin induces apoptosis in fibroblast-like
synoviocytes from collagen-induced arthritic rats mainly via the
mitochondrial pathway. Cytokine. 133:1551462020. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Xia ML, Xie XH, Ding JH, Du RH and Hu G:
Astragaloside IV inhibits astrocyte senescence: Implication in
Parkinson's disease. J Neuroinflammation. 17:1052020. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Yin Y, Qu H, Yang Q, Fang Z and Gao R:
Corrigendum to ‘Astragaloside IV alleviates Schwann cell injury in
diabetic peripheral neuropathy by regulating microRNA-155-mediated
autophagy’. Phytomedicine. 97:1539162022. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Lin J, Pan X, Huang C, Gu M, Chen X, Zheng
X, Shao Z, Hu S, Wang B, Lin H, et al: Dual regulation of microglia
and neurons by Astragaloside IV-mediated mTORC1 suppression
promotes functional recovery after acute spinal cord injury. J Cell
Mol Med. 24:671–685. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Li H, Xu J, Zhang Y, Hong L, He Z, Zeng Z
and Zhang L: Astragaloside IV alleviates senescence of vascular
smooth muscle cells through activating Parkin-mediated mitophagy.
Hum Cell. 35:1684–1696. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Feng M, Lv J, Zhang C, Chen D, Guo H, Tu
Y, Su L and Wang Z: Astragaloside IV protects sepsis-induced acute
kidney injury by attenuating mitochondrial dysfunction and
apoptosis in renal tubular epithelial cells. Curr Pharm Des.
28:2825–2834. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Li H, Yao C, Shi K, Zhao Y, Du J, Hu D and
Liu Z: Astragaloside IV attenuates hypoxia/reoxygenation
injury-induced apoptosis of type II alveolar epithelial cells
through miR-21-5p. Bioengineered. 12:7747–7754. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Wang F, Zhao Y, Chen S, Chen L, Sun L, Cao
M, Li C and Zhou X: Astragaloside IV alleviates ammonia-induced
apoptosis and oxidative stress in bovine mammary epithelial cells.
Int J Mol Sci. 20:6002019. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Qin S, Yin J, Huang S, Lin J, Fang Z, Zhou
Y and Huang K: Astragaloside IV protects ethanol-induced gastric
mucosal injury by preventing mitochondrial oxidative stress and the
activation of mitochondrial pathway apoptosis in rats. Front
Pharmacol. 10:8942019. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Yoshii SR and Mizushima N: Monitoring and
measuring autophagy. Int J Mol Sci. 18:18652017. View Article : Google Scholar : PubMed/NCBI
|