Introduction
Epidemiological reports show that the consumption of
vegetables and fruits rich in polyphenols can reduce the incidence
of various diseases, such as cardiovascular disease (1), allergic inflammation (2), Alzheimer’s disease (3), and cancer (4). Butein
(3,4,2′,4′-tetrahydroxychalcone), a bioactive polyphenol isolated
from the stem bark of cashews and Rhus verniciflua Stokes,
has been used as a food additive and traditional herb medicine
(5). Butein exhibits antioxidant
and anti-inflammatory activities (6,7). It
also exerts antitumor activities against a wide range of cancers,
including colon cancer (8),
osteosarcoma (9), lymphoma
(5), melanoma (10), and breast cancer (11). Treatment with butein modulates the
Bcl-2/Bax ratio, increases pro-caspase 3 activity, and subsequently
triggers apoptosis in cancer cells (9,12). In
addition, co-treatment of butein with TNF-related apoptosis
inducing ligand (TRAIL) enhances death receptor 5 (DR5) expression
and elevated caspase 3 activity, which, in turn, cause
TRAIL-resistant leukemia and hepatoma cell lines to undergo
apoptosis (13,14). Very recently, butein was found to
inhibit the migration of breast and pancreatic cancer via abrogated
nuclear factor κB (NFκB) activity and downregulated chemokine
receptor CXCR4 expression and function (15).
Neuroblastoma, a highly prevalent solid tumor in
children, accounts for 15% of all pediatric cancer-related deaths
worldwide (16). Due to its easy
metastasis and drug resistance, the mortality of neuroblastoma
patients is very high. Therefore, determination of novel drugs that
induce apoptosis in neuroblastoma cells without causing toxicity to
normal cells is of utmost importance in the treatment of
neuroblastoma. In the current study, we show for the first time
that butein markedly triggers apoptosis in neuroblastoma cells
through the generation of reactive oxygen species (ROS), alteration
of Bcl-2 and Bax expression, and elevation of pro-caspase 3
activity. Pre-treatment with antioxidant agents reverse
butein-induced effects. In general, butein may be a potential
chemotherapeutic agent for the effective treatment of
neuroblastoma.
Materials and methods
Materials
Butein,
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT),
NAC, and propidium iodide were purchased from the Sigma Chemical
Company (St. Louis, MO, USA). Antibodies against pro-caspase 3 and
PARP were purchased from Cell Signaling Technology (Beverly, MA,
USA). Antibodies against Bcl-2 and Bax were obtained from Santa
Cruz Biotechnology (Santa Cruz, CA, USA). Anti- β-actin and
HRP-conjugated secondary antibodies were obtained from Sigma
Chemical Company.
Cell culture
Mouse neuroblastoma Neuro-2A cells were maintained
in modified Eagle’s medium (Gibco; Gaithersburg, MD, USA)
supplemented with 10% fetal bovine serum (FBS), 100 U/ml penicillin
and 100 g/ml streptomycin (Gibco) at 37°C in a humidified 5%
CO2 incubator.
MTT assay
Cells were seeded in 24-well plates at a density of
4×104/ml and treated with the indicated concentrations
of butein for 24 or 48 h. After removal of the supernatant, the
cells were incubated with a medium containing 5.0 g/l MTT and
incubated at 37°C for an additional 3 h. The formazan was dissolved
in 1 ml isopropanol and the absorbance was measured at 563 nm. The
experiments were performed in duplicate and repeated at least 3
times.
Cell cycle distribution and apoptosis
analysis
Cells exposed to different concentrations of butein
for 24 h were collected and fixed with ice-cold ethanol overnight
at −20°C. Subsequently, cells were stained with 50 μg/ml propidium
iodide (PI) in the dark for 15 min. The cell cycle distribution was
analyzed on a BD Biosciences FACScan system using the CellQuest™
Pro software. The proportion of hypodiploid cells (sub-G1 phase)
was also analyzed.
Reactive oxygen species (ROS)
analysis
Cells were pretreated using 2 mM NAC for 1 h,
followed by treatment with an indicated concentration of butein for
an additional 24 h. The cells were then loaded with a 5 μM
fluorescent probe 2′,7′-dichlorodihydrofluorescein diacetate
(H2DCFDA) at 37°C for 1 h. The fluorescent intensity was analyzed
using Molecular Devices Flexstation 3.
Western blot analysis
Cells were treated with the indicated concentration
of butein for 24 h. Cells were rinsed with ice-cold phosphate
buffered saline (PBS) and lysed in buffer containing 150 mM NaCl,
1% Triton X-100, 5 mM EDTA, 5 mM phenylmethylsulfonyl fluoride, 1%
aprotinin, 1 g/ml leupeptin, and 500 μM
Na3VO4. The protein concentration was
detected using Bradford protein assay kit (Bio-Rad Laboratories,
Hercules, CA, USA). Twenty micrograms of protein were separated by
10% polyacrylamide gel and transferred into a nitrocellulose
membrane. The membrane was blocked by PBS containing 0.5% non-fat
milk for 1 h at room temperature. After washing with PBS containing
0.1% Tween-20 (PBST), the membrane was probed with the indicated
antibodies at 4°C overnight, washed with PBST, and subsequently
incubated with horseradish peroxidase-conjugated goat anti-mouse
IgG antibody (Santa Cruz Biotechnology, Inc., Santa Cruz, CA;
1:5,000 dilution) at room temperature for 1 h. The bands were
detected by enhanced chemiluminescence kit (Perkin-Elmer Life
Science, Boston, MA, USA). β-actin expression was used as the
loading control.
Statistical analysis
Data reported are the mean ± SD deviation of 3
independent experiments and evaluated by Student t-tests.
Significant differences were established at P<0.05.
Results
Effects of butein on the viability and
apoptosis of neuroblastoma cells
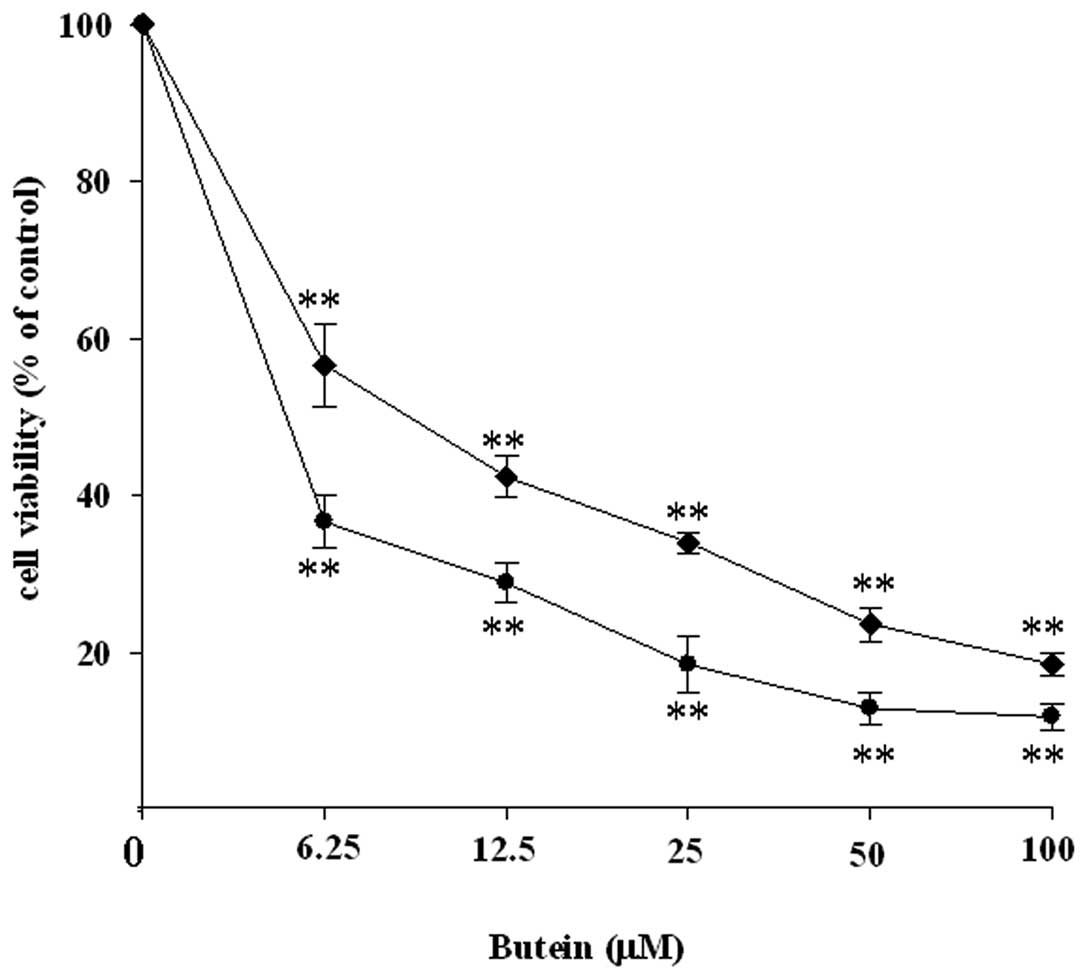
To determine the effects of butein on the cell
viability of neuroblastoma, Neuro-2A cells were treated with
various concentrations of butein for 24 and 48 h. Cell viability
was measured by the MTT assay. After treatment with 6.25, 12.5, 25,
50, and 100 μM butein for 24 h, cell viabilities were determined to
be around 56.65, 42.41, 34.08, 23.68 and 18.58%, respectively,
compared with the vehicle control. Cell viabilities were 36.70,
28.91, 18.54, 12.98 and 11.91% compared with the vehicle control
when the cells were exposed to 6.25, 12.5, 25, 50 and 100 μM butein
for 48 h. Collectively, treatment with butein resulted in a dose-
and time-dependent inhibition of cell viability in Neuro-2A cells
(Fig. 1).
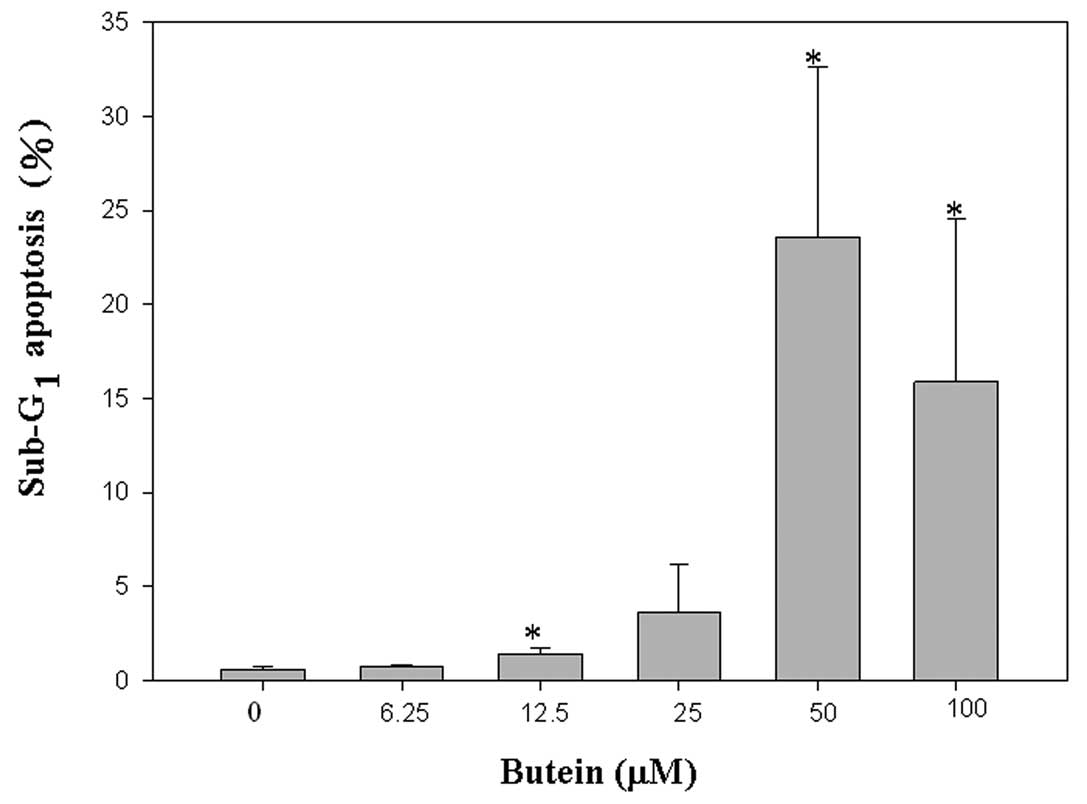
To further examine the inhibitory effects of butein
on cell viability, cell cycle progression was conducted by flow
cytometry analysis. Increased sub-G1 phase percentages were found
in proportion to the butein treatments (0.58, 0.71, 1.45, 3.63,
23.60 and 15.89% in the presence of 0, 6.25, 12.5, 25, 50, and 100
μM butein). Accordingly, treatment with butein diminished cell
viability mainly by triggering apoptosis (Fig. 2).
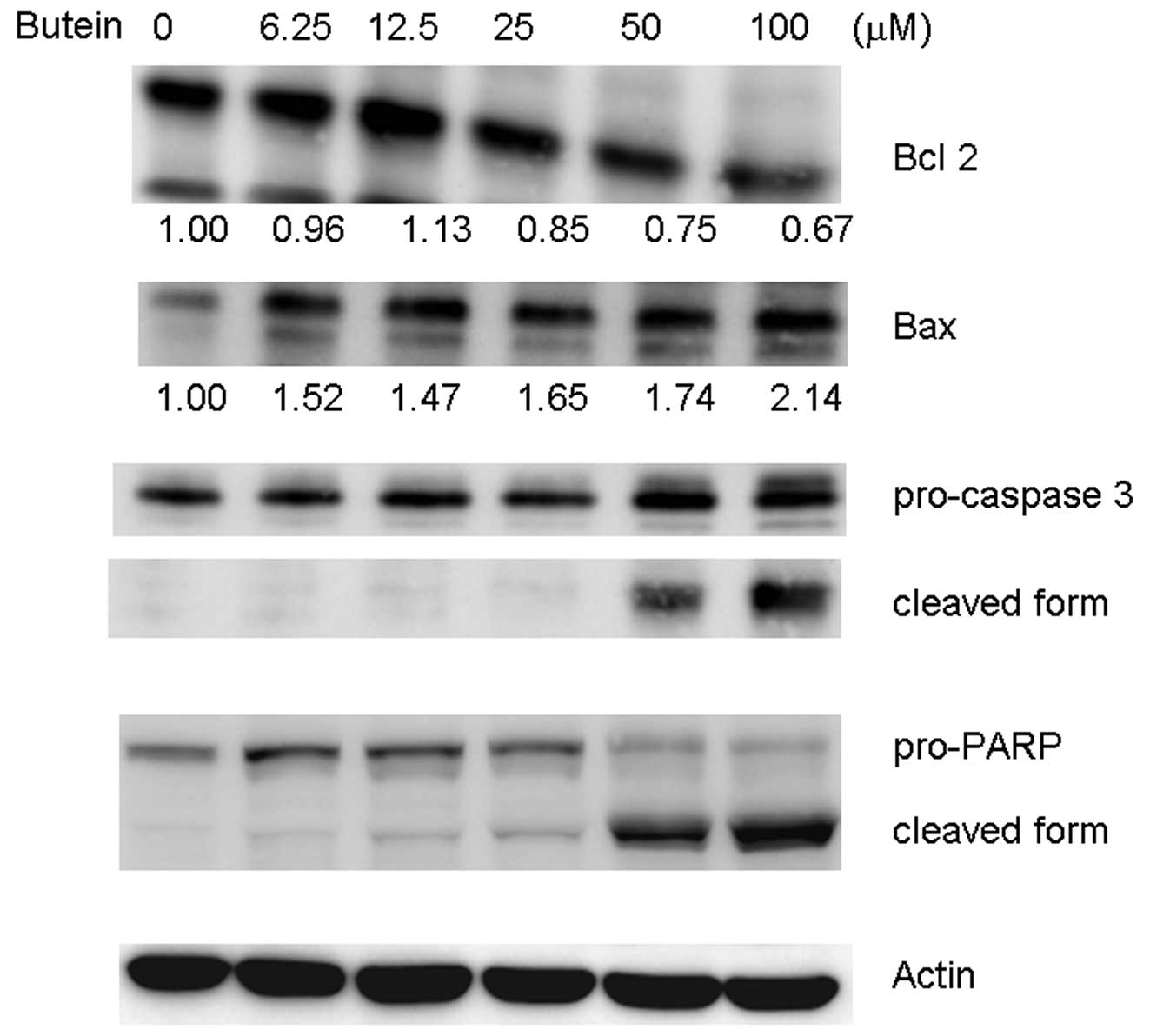
Effects of butein on apoptosis-related
protein expression
Bcl-2 family proteins are involved in apoptosis
(17,18). Decreased Bcl-2/Bax ratios may
trigger the release of cytochrome C from the mitochondria and
activation of pro-caspase 3 (18).
To elucidate the role of Bcl-2 family proteins in butein-induced
apoptosis, the expressions of Bcl-2, Bax, pro-caspase 3, and PARP
were measured in cell lysates derived from butein-treated cells. In
the presence of 25, 50, and 100 μM butein, Bcl-2 expression
decreased by 15, 25 and 30%, respectively, compared with the
vehicle control. Bax expression increased by 1.52, 1.47, 1.65, 1.74
and 2.14-fold compared with the control (Fig. 3). Significantly reduced pro-forms
and increased cleavage forms of caspase 3 and PARP were also
observed in the butein-treated groups (Fig. 3).
Effect of antioxidant agents on
butein-induced changes
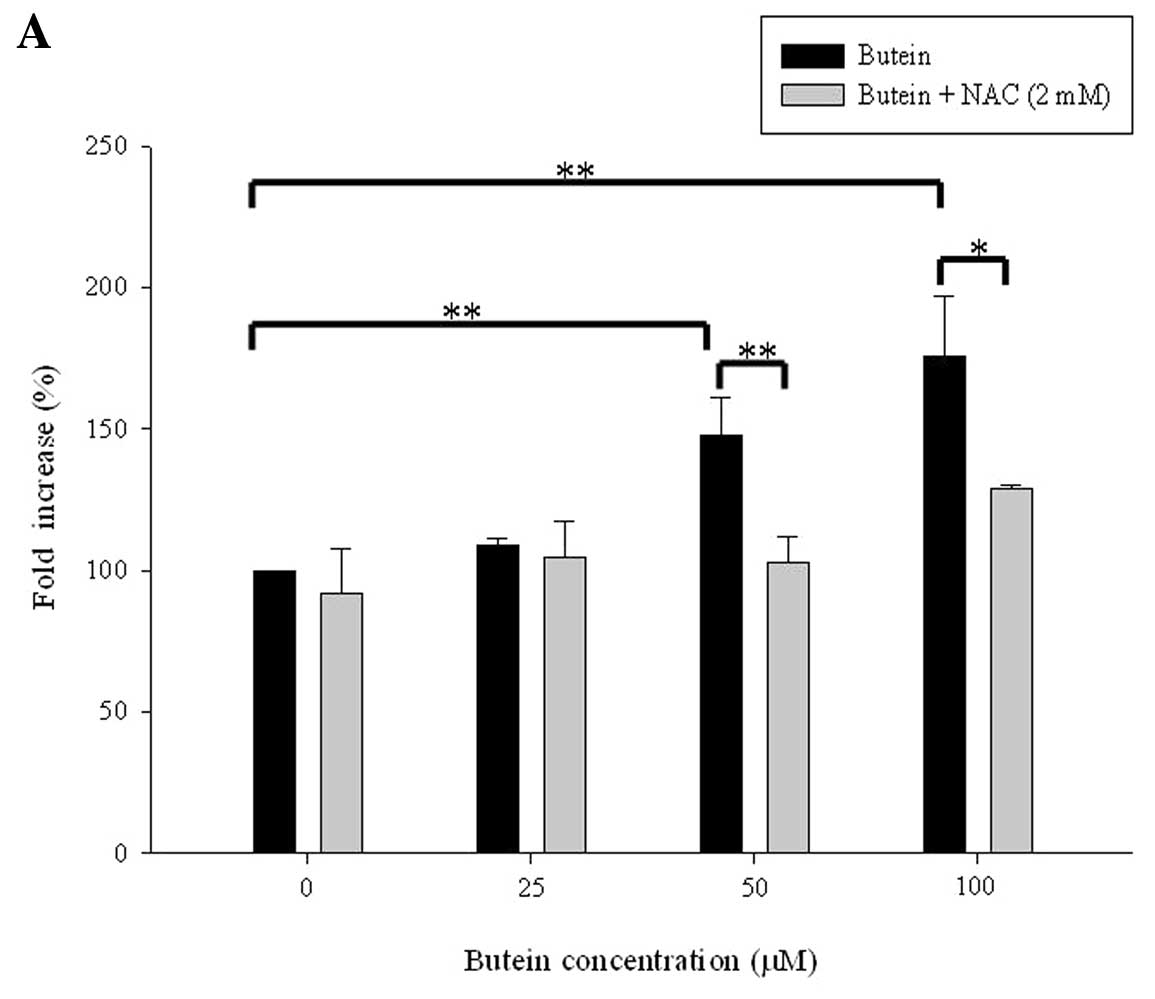
Recent studies demonstrated that generation of ROS
may function as a mediator in flavonoid-induced apoptosis in cancer
cells (19,20). To determine whether or not butein
induces oxidative stress, ROS generation by butein was determined
using the DCFDA staining method. As expected, treatment with 25, 50
and 100 μM butein significantly induced ROS production. ROS
generation was diminished by pretreatment with the antioxidant
agent NAC (Fig. 4A).
To determine whether or not elevated ROS levels
mediate the cytotoxic effects of butein, the effects of antioxidant
agents on cell viability was examined. Cells were pretreated with 2
mM NAC and then treated with 25, 50 and 100 μM butein for an
additional 24 h. As shown in Fig.
4B, butein-induced cell death was significantly prevented by
NAC. In the presence of NAC, cell viability recovered to about
61.56, 57.53 and 43.50% in the presence of 25, 50, and 100 μM
butein compared to the vehicle-treated group, respectively.
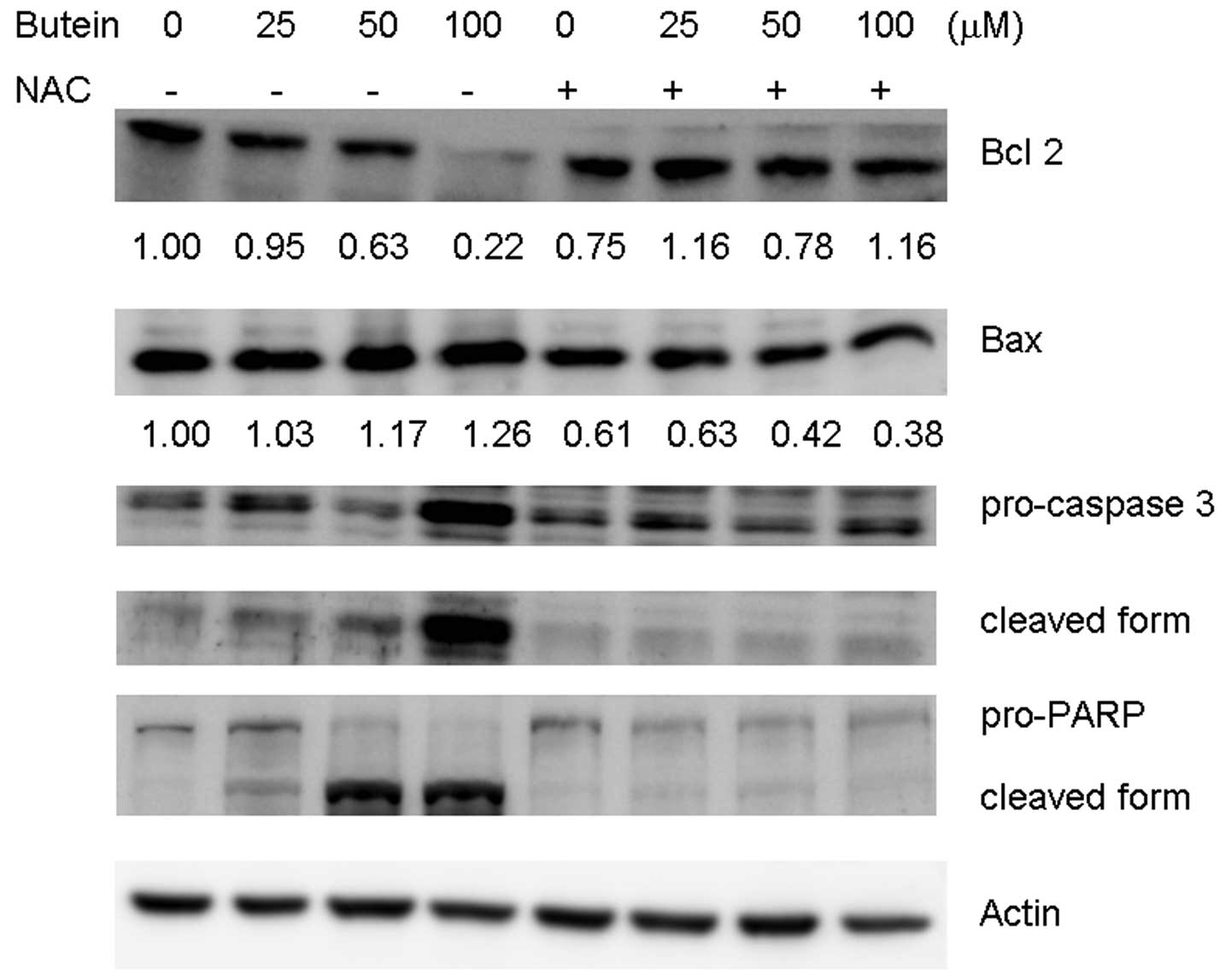
To determine whether or not NAC reverses
apoptosis-related protein expression, cell lysates derived from
combined treatments with NAC and butein were subjected to Western
blot analysis. NAC significantly recovered the expression of Bcl-2
and attenuated the expression of Bax expression (Fig. 5). Cleavage form of caspase 3 and
PARP was also prevented by NAC. Taken together, our results show
that ROS plays a critical role in butein-induced apoptosis.
Discussion
Extensive reports have revealed the inverse
relationship between the consumption of vegetables and fruits and
several chronic diseases, such as allergic inflammation,
hypertension, and cancer (21,22).
Flavonoids, one of the major bioactive compounds in vegetables and
fruits, exhibit anti-atherosclerosis (23), anti-inflammation (4), and antitumor effects (4). Butein, an ingredient isolated from
native plants, inhibits the proliferation of several human cancers,
such as melanoma (10), leukemia
(12), and osteosarcoma (9). In this presentation, the molecular
mechanisms of butein on the apoptosis of neuroblastoma cells were
delineated for the first time. Butein inhibited the cell viability
of neuroblastoma cells in a dose- and time-dependent manner.
Treatment with butein induced apoptosis, as evidenced by an
increased percentage of the sub-G1 population. In addition, butein
decreased the Bcl-2/Bax ratio, and enhanced the cleavage of caspase
3 and PARP. Pretreatment with antioxidant agents such as NAC
clearly attenuated butein-induced effects.
The balance of expression of pro-apoptotic and
anti-apoptotic members of the Bcl-2 family proteins plays a
critical role in apoptosis (24).
An increased pro-apoptotic/anti-apoptotic protein ratio altered the
membrane potential of the mitochondria, released apoptogenic
factors to the cytoplasma, activated the caspase cascade, and
eventually caused apoptosis (18,24).
Crude acetone extracts of R. verniciflua Stokes, which is
rich in fustin and fisetin, and butein-triggered osteosarcoma cells
undergo apoptosis via the activation of Bax and the down-regulation
of Bcl-2 expression (9). Similarly,
butein increased Bax expression and decreased Bcl-2 expression in
HL60 leukemia cells (12). In
accordance with previous reports, our findings also showed that
butein upregulated Bax expression and attenuated Bcl-2 expression
in neuroblastoma cells. Moreover, activation of caspase 3 and
cleavage of PARP, 2 hallmarks of apoptosis, were also observed in
butein-treated Neuro-2A cells. Collectively, our data suggest that
butein-induced apoptosis may mediate by increase ROS level and
change the pro-apoptotic/anti-apoptotic protein ratio.
Alternation of intracellular ROS production may
modulate several physiological functions (25). Elevated ROS can bind to lipids,
proteins, or DNA to produce oxidative stress and eventually cause
cell death (25). Flavonoid-induced
apoptosis in cancer cells via elevation of intracellular ROS levels
has been shown in several reports (19,20),
although previous reports have also shown that butein induces ROS
generation, modulates JNK, ATM, and Chk activity, and causes G2/M
arrest in hepatoma cells (26). The
consequence of butein-induced ROS production in cell death remains
unclear. In the current presentation, butein enhanced ROS levels up
to 1.5-fold and caused cell death. Blockage of ROS generation by an
antioxidant agent, NAC, significantly increased cell viability in
the presence of butein. Cotreatment with NAC recovered the
anti-apoptotic protein Bcl-2 and decreased pro-apoptotic Bax
levels, which, in turn, attenuated pro-caspase 3 activity. ROS
appears to play a pivotal role in butein-induced apoptosis in
neuroblastoma cells.
In summary, to the best of our knowledge, this is
the first report to show that butein inhibits cell viability in
neuroblastoma cells. Butein triggers apoptosis, as evidenced by an
increase in the sub-G1 population. Butein also elevated ROS levels,
altered Bcl-2/Bax ratios, induced pro-caspase 3 and PARP cleavage,
and consequently caused cells to undergo apoptosis. Pre-treatment
with NAC obviously abrogated the butein-induced effects.
Collectively, our findings suggest that butein may serve as a
chemotherapeutic agent for the future treatment of
neuroblastoma.
Acknowledgements
This study was supported by grants of National
Science Council of Taiwan (NSC-94-2311-B-040-002 and
NSC-93-2311-B-040-009). Molecular Devices Flexstation 3 was
performed in the Instrument Center of Chung Shan Medical
University, which is supported by National Science Council,
Ministry of Education and Chung Shan Medical University.
References
|
1
|
Michalska M, Gluba A, Mikhailidis DP,
Nowak P, Bielecka-Dabrowa A, Rysz J and Banach M: The role of
polyphenols in cardiovascular disease. Med Sci Monit.
16:RA110–RA119. 2010.PubMed/NCBI
|
|
2
|
Enomoto T, Nagasako-Akazome Y, Kanda T,
Ikeda M and Dake Y: Clinical effects of apple polyphenols on
persistent allergic rhinitis: a randomized double-blind
placebo-controlled parallel arm study. J Investig Allergol Clin
Immunol. 16:283–289. 2006.PubMed/NCBI
|
|
3
|
Singh M, Arseneault M, Sanderson T, Murthy
V and Ramassamy C: Challenges for research on polyphenols from
foods in Alzheimer’s disease: bioavailability, metabolism, and
cellular and molecular mechanisms. J Agric Food Chem. 56:4855–4873.
2008.PubMed/NCBI
|
|
4
|
Guo W, Kong E and Meydani M: Dietary
polyphenols, inflammation, and cancer. Nutr Cancer. 61:807–810.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lee JC, Lee KY, Kim J, Na CS, Jung NC,
Chung GH and Jang YS: Extract from Rhus verniciflua Stokes
is capable of inhibiting the growth of human lymphoma cells. Food
Chem Toxicol. 42:1383–1388. 2004.
|
|
6
|
Jung CH, Jun CY, Lee S, Park CH, Cho K and
Ko SG: Rhus verniciflua stokes extract: radical scavenging
activities and protective effects on
H2O2-induced cytotoxicity in macrophage RAW
264.7 cell lines. Biol Pharm Bull. 29:1603–1607. 2006. View Article : Google Scholar
|
|
7
|
Jung CH, Kim JH, Hong MH, Seog HM, Oh SH,
Lee PJ, Kim GJ, Kim HM, Um JY and Ko SG: Phenolic-rich fraction
from Rhus verniciflua Stokes (RVS) suppress inflammatory
response via NF-kappaB and JNK pathway in
lipopolysaccharide-induced RAW 264.7 macrophages. J Ethnopharmacol.
110:490–497. 2007.
|
|
8
|
Yit CC and Das NP: Cytotoxic effect of
butein on human colon adenocarcinoma cell proliferation. Cancer
Lett. 82:65–72. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jang HS, Kook SH, Son YO, Kim JG, Jeon YM,
Jang YS, Choi KC, Kim J, Han SK, Lee KY, Park BK, Cho NP and Lee
JC: Flavonoids purified from Rhus verniciflua Stokes
actively inhibit cell growth and induce apoptosis in human
osteosarcoma cells. Biochim Biophys Acta. 1726:309–316.
2005.PubMed/NCBI
|
|
10
|
Iwashita K, Kobori M, Yamaki K and
Tsushida T: Flavonoids inhibit cell growth and induce apoptosis in
B16 melanoma 4A5 cells. Biosci Biotechnol Biochem. 64:1813–1820.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kim JH, Jung CH, Jang BH, Go HY, Park JH,
Choi YK, Hong SI, Shin YC and Ko SG: Selective cytotoxic effects on
human cancer cell lines of phenolic-rich ethyl-acetate fraction
from Rhus verniciflua Stokes. Am J Chin Med. 37:609–620.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kim NY, Pae HO, Oh GS, Kang TH, Kim YC,
Rhew HY and Chung HT: Butein, a plant polyphenol, induces apoptosis
concomitant with increased caspase-3 activity, decreased Bcl-2
expression and increased Bax expression in HL-60 cells. Pharmacol
Toxicol. 88:261–266. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kim N: Butein sensitizes human leukemia
cells to apoptosis induced by tumor necrosis factor-related
apoptosis inducing ligand (TRAIL). Arch Pharm Res. 31:1179–1186.
2008. View Article : Google Scholar
|
|
14
|
Moon DO, Kim MO, Choi YH and Kim GY:
Butein sensitizes human hepatoma cells to TRAIL-induced apoptosis
via extracellular signal-regulated kinase/Sp1-dependent DR5
upregulation and NF-kappaB inactivation. Mol Cancer Ther.
9:1583–1595. 2010. View Article : Google Scholar
|
|
15
|
Chua AW, Hay HS, Rajendran P, Shanmugam
MK, Li F, Bist P, Koay ES, Lim LH, Kumar AP and Sethi G: Butein
downregulates chemokine receptor CXCR4 expression and function
through suppression of NF-kappaB activation in breast and
pancreatic tumor cells. Biochem Pharmacol. 80:1553–1562. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mueller S and Matthay KK: Neuroblastoma:
biology and staging. Curr Oncol Rep. 11:431–438. 2009. View Article : Google Scholar
|
|
17
|
Llambi F and Green DR: Apoptosis and
oncogenesis: give and take in the BCL-2 family. Curr Opin Genet
Dev. 21:12–20. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ola MS, Nawaz M and Ahsan H: Role of Bcl-2
family proteins and caspases in the regulation of apoptosis. Mol
Cell Biochem. 351:41–58. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chang IC, Huang YJ, Chiang TI, Yeh CW and
Hsu LS: Shikonin induces apoptosis through reactive oxygen
species/extracellular signal-regulated kinase pathway in
osteosarcoma cells. Biol Pharm Bull. 33:816–824. 2010. View Article : Google Scholar
|
|
20
|
Juan ME, Wenzel U, Daniel H and Planas JM:
Resveratrol induces apoptosis through ROS-dependent mitochondria
pathway in HT-29 human colorectal carcinoma cells. J Agric Food
Chem. 56:4813–4818. 2008. View Article : Google Scholar
|
|
21
|
Kelloff GJ, Boone CW, Crowell JA, Steele
VE, Lubet R and Sigman CC: Chemopreventive drug development:
perspectives and progress. Cancer Epidemiol Biomarkers Prev.
3:85–98. 1994.PubMed/NCBI
|
|
22
|
Khan N, Afaq F and Mukhtar H: Cancer
chemoprevention through dietary antioxidants: progress and promise.
Antioxid Redox Signal. 10:475–510. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Grassi D, Desideri G, Croce G, Tiberti S,
Aggio A and Ferri C: Flavonoids, vascular function and
cardiovascular protection. Curr Pharm Des. 15:1072–1084. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cory S and Adams JM: The Bcl2 family:
regulators of the cellular life-or-death switch. Nat Rev Cancer.
2:647–656. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Valko M, Leibfritz D, Moncol J, Cronin MT,
Mazur M and Telser J: Free radicals and antioxidants in normal
physiological functions and human disease. Int J Biochem Cell Biol.
39:44–84. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Moon DO, Kim MO, Choi YH, Hyun JW, Chang
WY and Kim GY: Butein induces G(2)/M phase arrest and apoptosis in
human hepatoma cancer cells through ROS generation. Cancer Lett.
288:204–213. 2010. View Article : Google Scholar : PubMed/NCBI
|