Introduction
Glioblastoma multiforme (GBM) is the most common and
fatal intracranial tumor (1). Till
now, multimodality treatments with surgery, radiation and
chemotherapy have led to some improvement in prognosis for patients
with GBM (2,3). However, the 5-year survival rate for
patients with GBM remains less than 5% and the median survival time
is approximately one year (4).
Multiple lines of evidence indicate that oxidative
stress plays an important role in the occurrence and progress of
malignant tumors (5,6). Tumor cells themselves can induce
oxidative stress. NF-E2-related factor 2 (Nrf2) is considered as a
critical regulator of intracellular antioxidants and phase II
detoxification enzymes by transcriptional upregulation of many
antioxidant response element (ARE)-containing genes (7–10).
Nrf2/Kelch-like ECH-associated protein 1 (Keap1) signaling pathway
is the main pathway for cell adaptation to oxidative stress.
Furthermore, current studies agree that Nrf2 plays a dual role in
tumor cells. Apart from its positive role in normal cells, Nrf2
also has its dark side. It is responsible for drug resistance of
certain cancer cells (11).
In addition to the activation of Nrf2 response,
oxidative stress may cause the induction of macroautophagy
(referred to hereafter as autophagy). Autophagy is a
self-cannibalization process consisting of the degradation and
recycling of organelles and portions in cytosol (12). This process plays a major role in
maintaining cellular homeostasis and some reports suggest that the
main role of autophagy in cancer cells is cell protection (13–16).
However, autophagic cell death could also be initiated in response
to sustained intracellular damage caused by hypoxia,
chemotherapeutic agents, virus infection, or toxins, especially in
gliomas (17). Recent studies found
that GBM is resistant to chemotherapy protocols that induce
apoptosis but seems to be less resistant to therapies that induce
autophagy (18–20). As an effective cytotoxic drug,
temozolomide (TMZ) induces significant autophagic cell death in
many GBM cells, such as U251 human glioma cell line (18,21,22).
Thus, autophagy has potential utility as a target for GBM therapy
(23,24). Recently, several experts proposed
the hypothesis that there may be cross-talk between Nrf2/Keap1 and
autophagy pathways (25). Thus, the
aim of the current study was to determine whether Nrf2 can
influence autophagy in U251 human glioma cell line after TMZ.
Materials and methods
Reagents
TMZ was kindly supplied by Schering-Plough Co.
(Kenilworth, NJ, USA) and was dissolved in dimethyl sulfoxide
(DMSO, Sigma Chemical Co., St. Louis, MO, USA) in 100 mM stock
solution. TMZ was used at a concentration of 100 μM. The final
concentration of DMSO which did not exceed 0.1% in the culture
medium did not influence cell viability and the expression of the
proteins studied as previously described (26). Acridine orange was purchased from
Sigma Chemical Co.
Cell culture and transient
transfection
Human U251 glioma cells were obtained from American
Tissue Culture Colection (ATCC) and cultured in Dulbecco’s modified
Eagle medium (DMEM, HyClone, IL, USA) with 10% fetal bovine serum
(HyClone) at 37°C and 5% CO2 incubator. The vector
pGPH1/GPF/Neo used for cloning Nrf2 short hairpin RNA (shRNA) was
purchased from GenePharma (Shanghai, China). The target sequence
was GCAGTTCAATGAAGCTCAACT. The new plasmid was named as Si-Nrf2.
Random sequence was used as negative control, which was named as
Si-control. Cells were seeded in 6-well plates at
1×106/well and allowed to attach for 24 h before
transfection. Then Si-Nrf2 and Si-control were transfected by
Lipofectamine 2000 (Invitrogen, CA, USA) according to the
manufacturer’s protocol. Cells treated with Lipofectamine 2000
alone were set up as blank control, which was named as group Lipo.
After incubation at 37°C and 5% CO2 for 48 h, cells were
collected.
Ultrastructural analysis of autophagy by
transmission electron microscopy (TEM)
For the TEM analysis, transfected cells after 48 h
were trypsinized, washed twice with PBS and fixed with ice-cold
glutaraldehyde (2.5% in 0.1 M cacodylate buffer, pH 7.4) for 30
min. After washing in PBS the cells were postfixed in 1% osmium
tetroxide (OsO4) and embedded in Epon; 0.1 mm thin
sections were stained with uranyl acetate/lead citrate (Fluka, St.
Louis, MO, USA) and viewed in a JEOL JEM-1011 TEM (Tokyo,
Japan).
Western blot analysis
To obtain total protein lysates, transfected cells
after 48 h and further treated with or without TMZ (100 μM) for 3
days were homogenized in RIPA buffer (1% NP40, 0.5% sodium
deoxycholate, 0.1% sodium dodecyl sulfate, 1 mM ethylene diamine
tetraacetic acid, 1 mM ethylene glycol tetraacetic acid, 1 mM
Na3VO4, 20 mM NaF, 0.5 mM DL-dithiothreitol,
1 mM phenylmethane-sulfonyl fluoride, and protease inhibitor
cocktail in PBS, pH 7.4) and centrifuged at 14,000 × g for 15 min
at 4°C. Protein concentrations were estimated by Coomassie Plus
Protein Assay Reagent (Pierce, IL, USA). Equal amounts of protein
from each sample was separated by SDS-PAGE on 8–12% gels and
transferred to polyvinylidene difluoride (PVDF) membrane
(Millipore, Bedford, MA, USA). Following incubation with primary
antibodies against Nrf2 (Abcam, Cambridge, UK; 1:500),
Microtubule-associated protein light chain 3B (LC3B, 1:3000),
β-actin (Santa Cruz Biotechnology, Inc., CA, USA; 1:500), GAPDH
(Santa Cruz Biotechnology, Inc.; 1:1000) and peroxidase-conjugated
goat anti-rabbit IgG (KeyGen Biotech, Nanjing, China; 1:5000) as
the secondary antibody, specific protein bands were visualized
using ECL detection system (Amersham Biosciences, Bucks, UK) and
exposed radiographic film (Fuji Hyperfilm, Tokyo, Japan). The
developed film was digitized using an Epson Perfection 2480 scanner
(Seiko Co., Nagano, Japan). The levels of Nrf2 and LC3B were
quantified by densitometry using ImageJ program and expressed
relative to β-actin and GAPDH signals, respectively.
Acridine orange (AO) immunofluorescent
staining and flow cytometric analysis
Quantification of autophagy by AO staining using
flow cytometry was performed as described previously (5). Transfected cells after 48 h treated
with or without TMZ (100 μM) for 3 days were stained with 1 μg/ml
AO for 15 min at 37°C, trypsinized, washed, and collected in phenol
red-free growth medium. Green (510–530 nm) and red (650 nm)
fluorescence emission from cells illuminated with blue (488 nm)
excitation light was measured with flow cytometer. Depending on
their acidity, autophagic lysosomes appeared as the orange/red
fluorescent cytoplasmic vesicles, while nuclei were stained green.
AO-stained cells were analyzed on a FACSCalibur flow cytometer
(Becton, Dickinson and Company, NJ, USA) using FlowJo7.6.1 software
(Tree Star, Inc., San Carlos, CA, USA). Autophagy was quantified as
a ratio between geomean fluorescence intensity of red vs. green
fluorescence (FL3/FL1).
Cell viability assay by cell counting
kit-8 (CCK-8) assay
Cell viability was assessed by CCK-8 assay (Dojindo,
Kumamoto, Japan). Cells (1×103/well) were seeded to
96-well culture plates and cultivated for 24 h to adhere, then
transfected according to the manufacturer’s protocol. After 48 h,
total medium was changed to 100 μl complete culture medium with or
without 100 μM TMZ and cells were incubated for a further 72 h.
DMSO was used as solvent control. CCK-8 (10 μl) was added into
every well and incubated for 2 h. Then the OD value was read at 450
nm using a Bio-Rad ELISA microplate reader (Bio-Rad Laboratories,
CA, USA). The viability rate of tumor cells equals (the OD values
of treated groups/the OD values of control group) ×100%.
Statistical analysis
All experiments were done at least three times. Data
were presented as mean ± SD. The statistical significance of the
differences between treatments was assessed using one-way ANOVA
followed by Student-Neuman-Keuls test for multiple comparisons. The
value of p<0.05 was considered significant. All analyses were
performed by SPSS 19.0.
Results
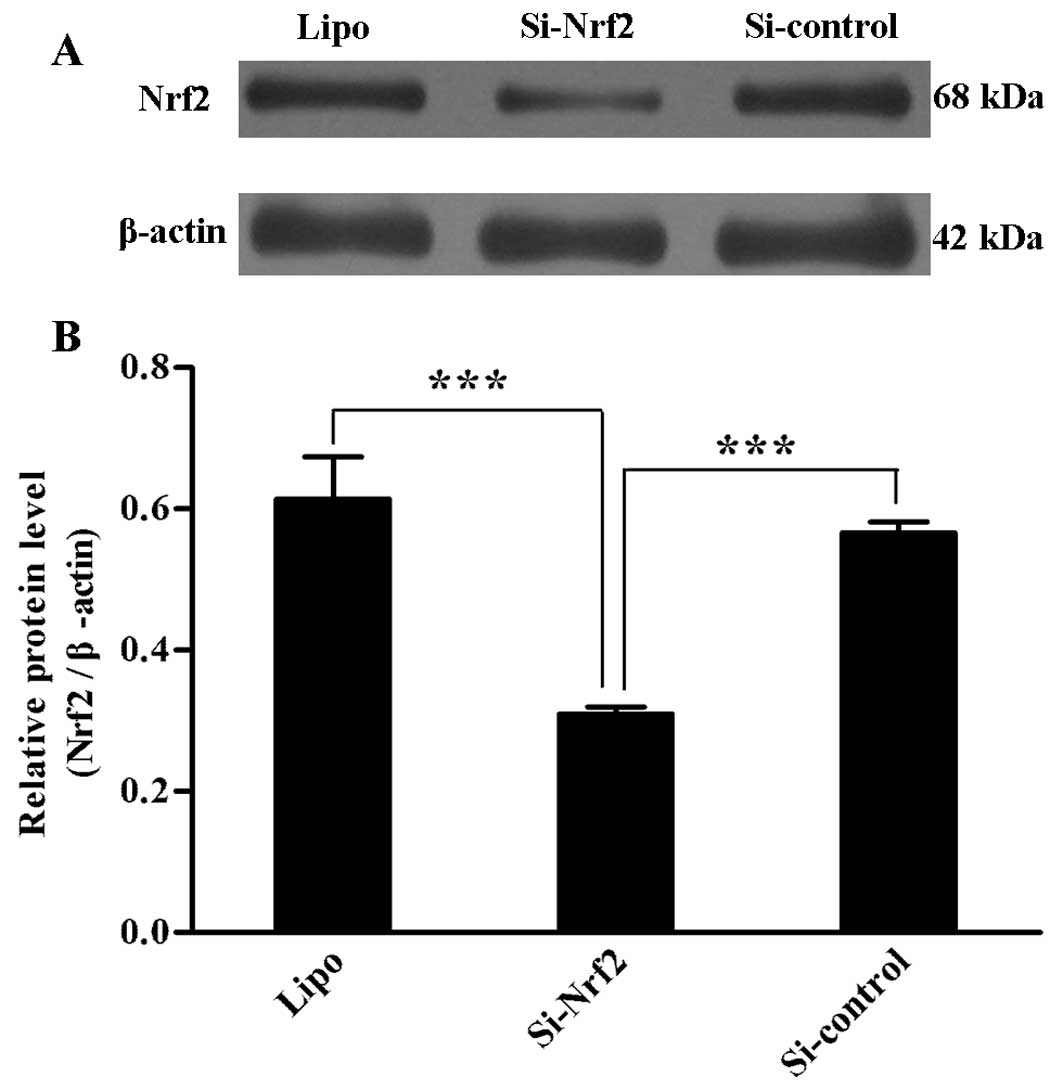
Transient transfection effect on Nrf2
protein level
To validate the transient transfection effect of two
plasmids mentioned above, we tested the protein level of three cell
groups 48 h after transfection, respectively. Si-Nrf2 transfection
reduced the Nrf2 protein level compared with transfection of
Si-control (p=0.01) and group Lipo (p=0.003). Group Si-control and
Lipo showed no difference in Nrf2 protein level (Fig. 1A and B). The efficiency of each
transfection was verified by this method.
Effect of Nrf2 downregulation on the
basal level of autophagy in U251 cells
To examine the role of Nrf2 downregulation on the
basal level of autophagy in U251 cells, autophagy-related
qualitative and quantitative detection were performed
accordingly.
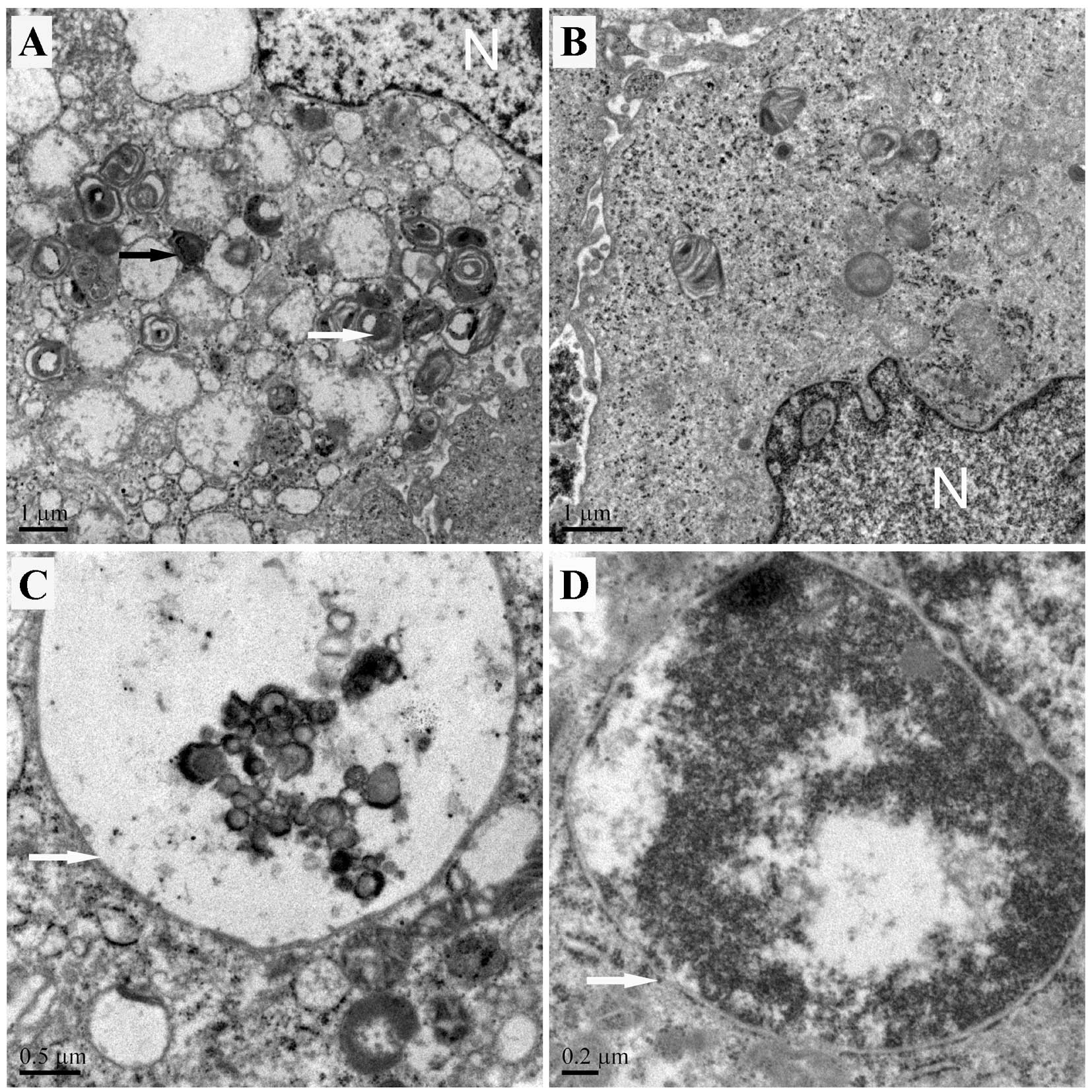
We observed autophagic structures by TEM. The result
showed that autophagic vacuoles increased in group Si-Nrf2 compared
to group Si-control. Most of the autophagosomes in group Si-Nrf2
contained lamellar structures or residual digested components,
whereas tumor cells of group Si-control exhibited few such features
(Fig. 2A and B). Also shown were
representative images of an autophagolysosome containing undegraded
cellular contents (Fig. 2C) and the
fusion of a late stage autophagic vacuoles with degraded cellular
contents (Fig. 2D).
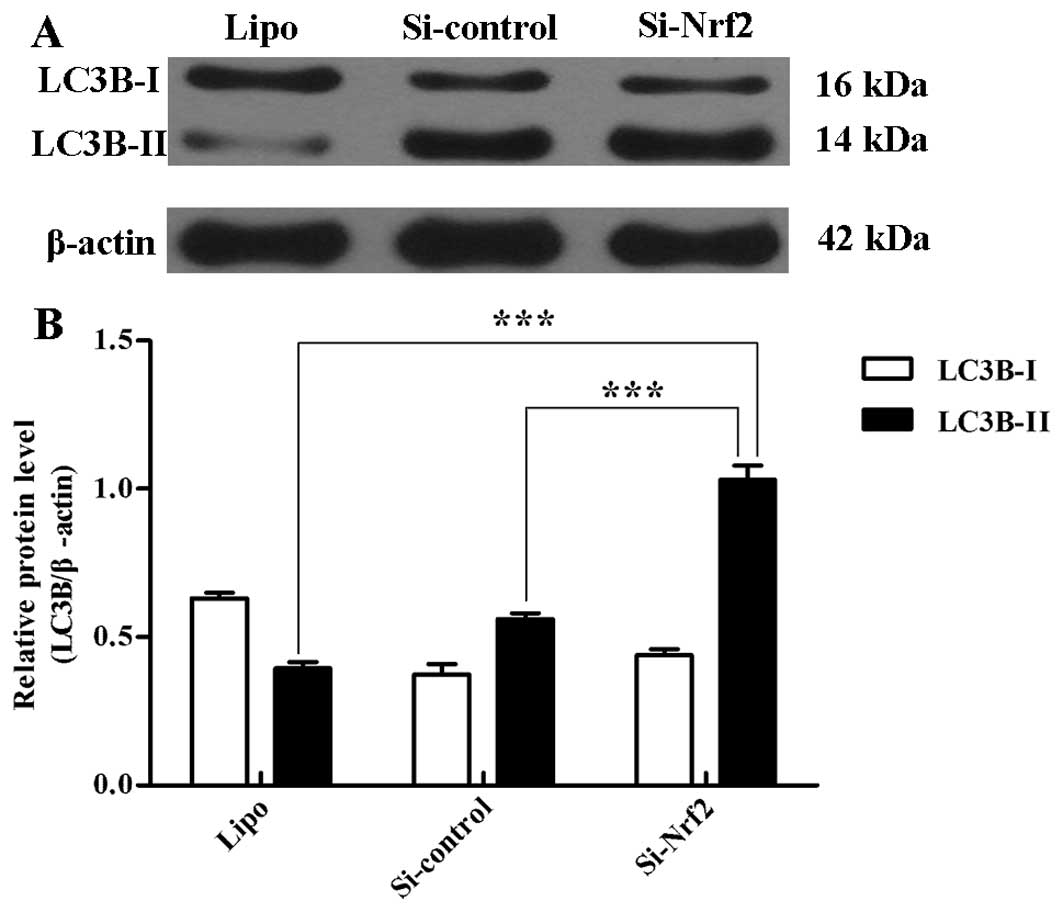
To quantify the incidence of the basal level of
autophagy, we tested the expression of two LC3B forms (LC3B-I and
LC3B-II) using western blot analysis. Si-Nrf2 transfection
significantly raised the LC3B-II protein level compared with
transfection of Si-control (p<0.001) and Lipo (p<0.001)
(Fig. 3A and B).
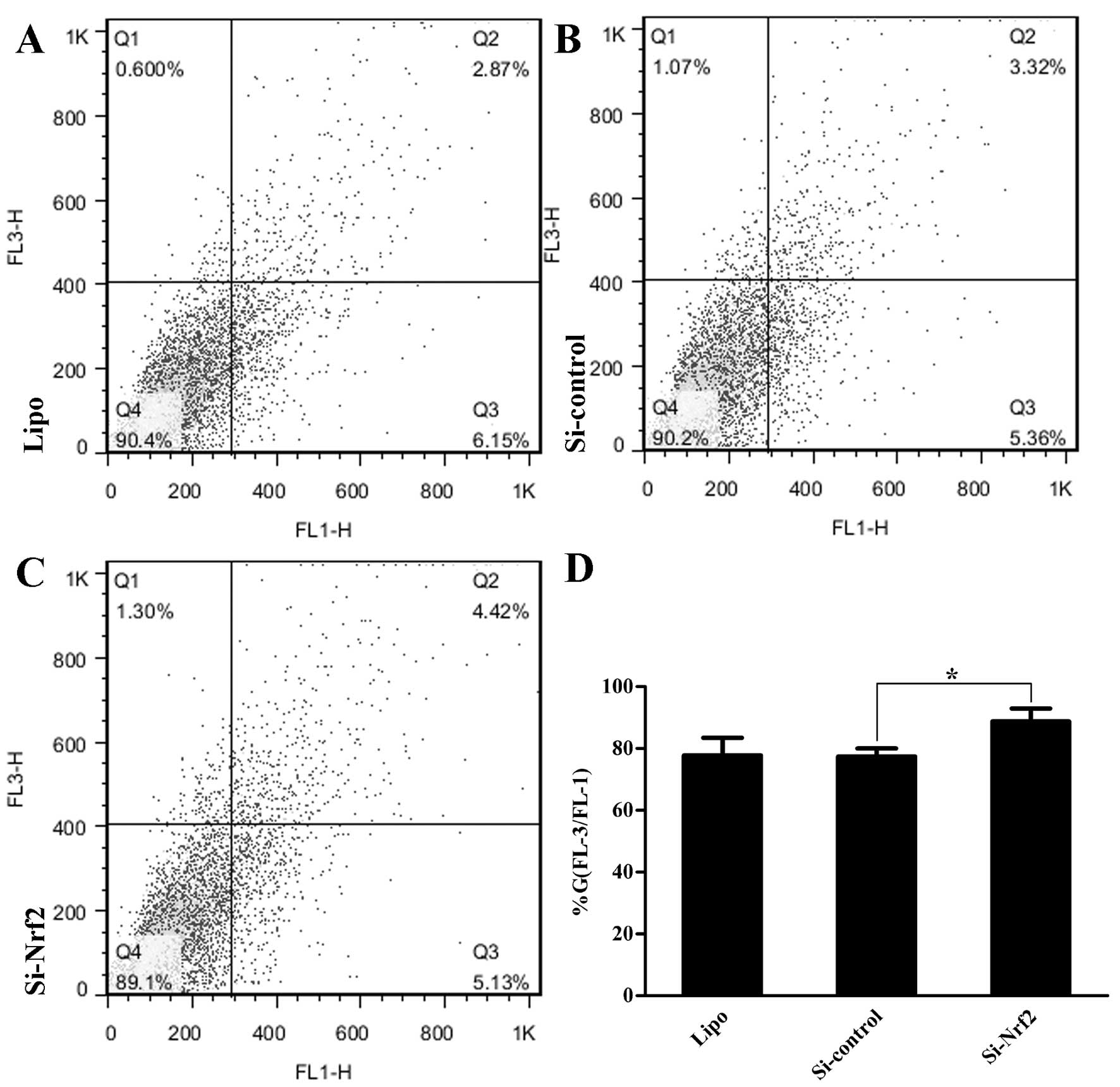
In addition, we quantified the presence of acidic
vesicular organelles (AVOs) by flow cytometry with AO staining
(4). Group Si-Nrf2 had more AVOs
(p=0.017) (Fig. 4A–D) in comparison
with group Si-control. Collectively, the findings demonstrated that
group Si-Nrf2 had a higher level of basal autophagy than control
groups.
Effect of Nrf2 downregulation on
autophagy induced by TMZ in U251 cells
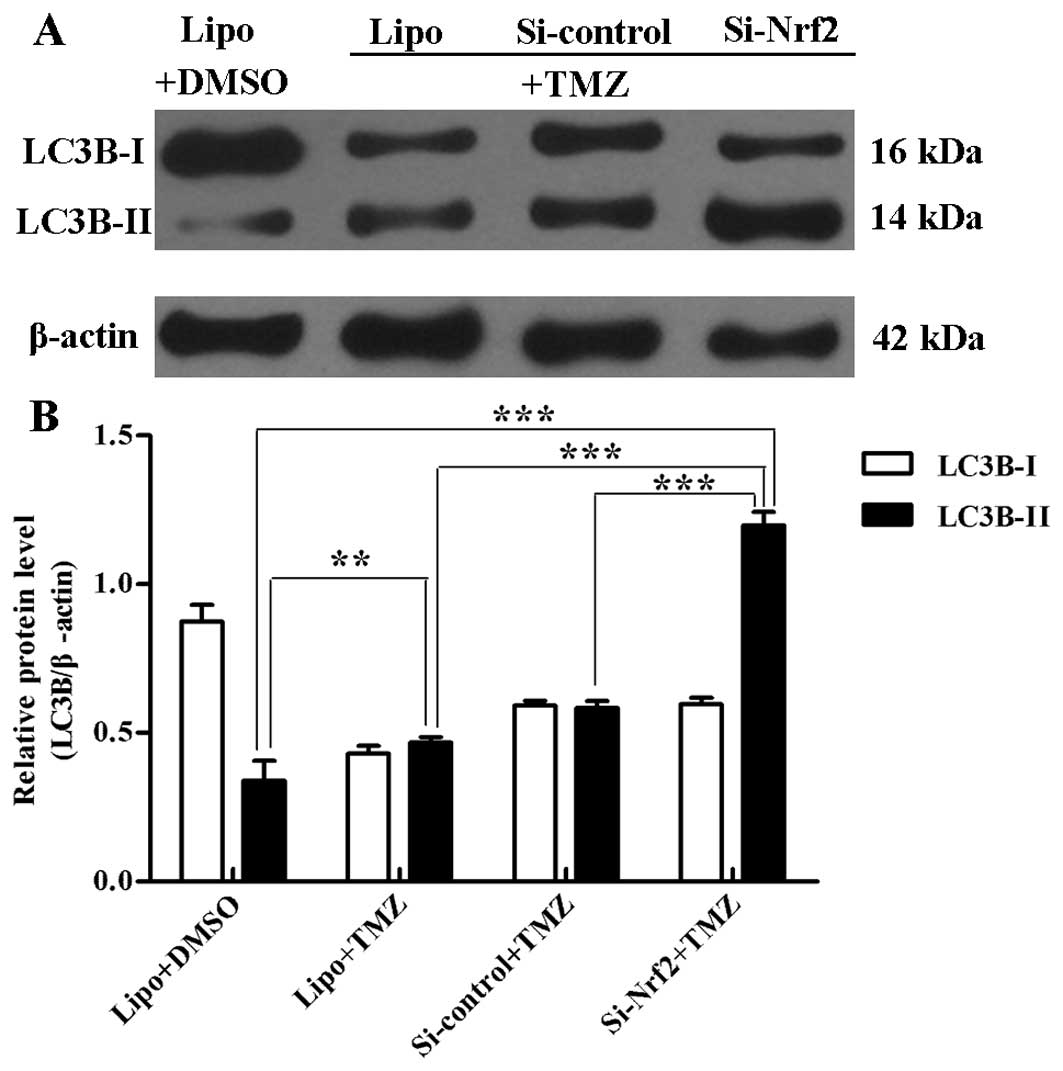
After treatment with TMZ (100 μM) for 3 days,
transfected U251 cells were treated similar to the above methods.
Western blot results showed that group Si-Nrf2 treated with TMZ
raised the protein level of LC3B-II compared with other groups
(p<0.001). In group Lipo, cells treated with TMZ had more
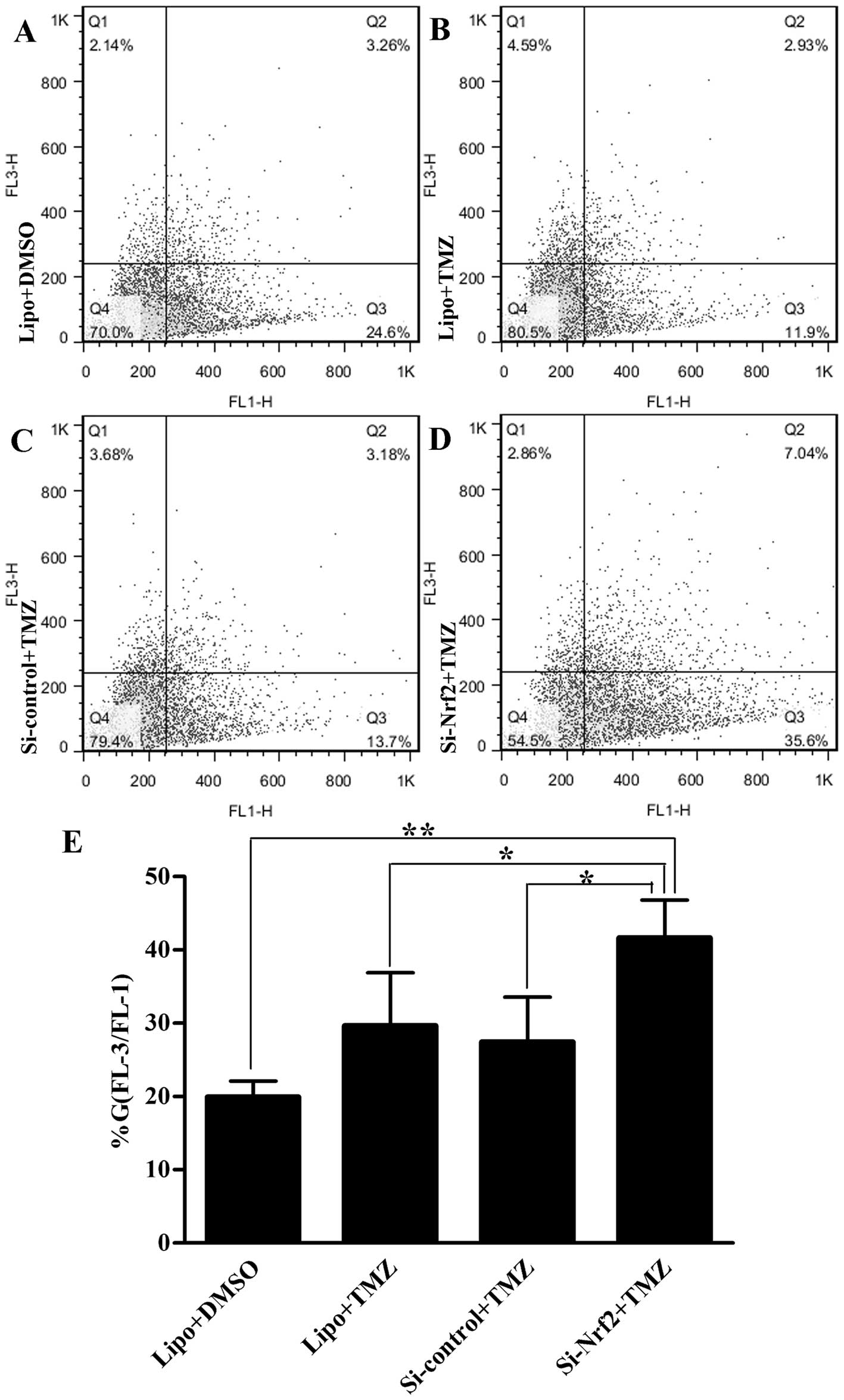
LC3B-II expressions than with DMSO treatment (p=0.007) (Fig. 5A and B). Likewise, there were more
AVOs in group Si-Nrf2 than other groups (p<0.05) detected by
flow cytometry (Fig. 6A–E).
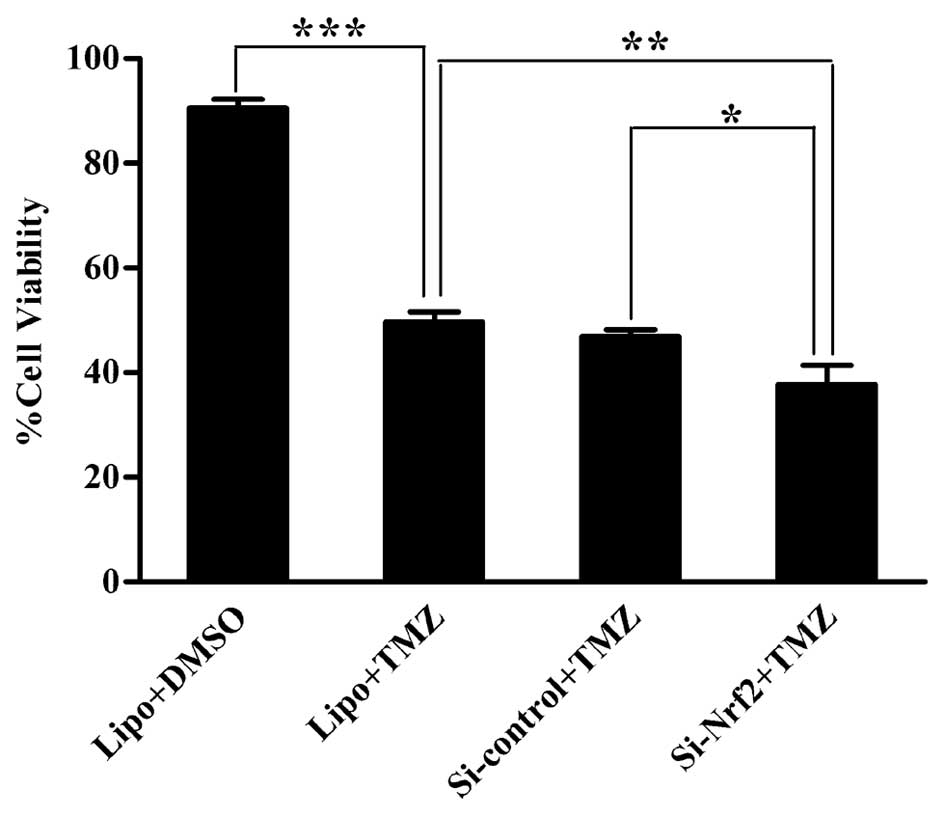
Effect of Nrf2 downregulation on cell
viability in U251 cells after TMZ
To characterize the effect of Nrf2 downregulation on
viability of glioma cells after TMZ, we performed CCK-8 assay.
Group Lipo treated with TMZ had much less cells than the one
treated with DMSO (p<0.001). After TMZ, cells in group Si-Nrf2
showed a decrease compared to those in group Si-control (p=0.015)
and Lipo (p=0.007) in cell viability. Group Si-control and Lipo
showed no significant difference (Fig.
7).
Discussion
In this study, the results showed that knockdown of
Nrf2 significantly enhanced the basal level of autophagy in U251
cells characterized by increased autophagic structures by TEM,
elevated expression of LC3B-II at protein level and increased AVOs
by flow cytometry. In addition, higher level of autophagy and lower
level of cell viability were found in group Si-Nrf2 than in the
other groups after TMZ. To our knowledge, the findings showed for
the first time the Nrf2 downregulation enhanced autophagy induced
by TMZ in U251 cells.
We evaluated the protein level of Nrf2 after
transfection because it reflected the transient transfection
efficiency and correlated with the effectiveness of follow-up
results. It has been revealed that oxidative stress can be induced
by tumor cells. Nrf2 is mutational and overexpressed in many tumor
tissues and cell lines (27,28).
Nrf2 transcription complex specifically recognizes ARE, which is
located in the promoter region of all Nrf2-targeted genes and
subsequently initiates the expression of a group of genes. These
Nrf2-targeted gene products exert protection for cells from
oxidative damage (5). As previously
reported, Nrf2 was highly expressed in U251 cells and the
transfection efficiency was most predominant at 48 h after
transfection (5,29). Hence, we chose the U251 cells and
accessed the transient transfection 48 h after transfection.
Consistent with previous results, Si-Nrf2 transfection
significantly reduced the Nrf2 protein level compared with
transfection of Si-control.
The basal level of autophagy in GBM cells is higher
than that in normal cells. Oxidative stress may be the fundamental
cause. The basal level of autophagy is vital for the biological
activity of tumors. In order to evaluate whether Nrf2 can regulate
basal autophagy, TEM, quantification of LC3B-II protein levels and
AVOs were performed. The use of TEM is a valid and important
morphological method for various autophagic structures (30,31).
In this study, a larger number of autophagosomes and
autophagolysosomes that correlated with the induction of autophagy
were found in group Si-Nrf2 than group Si-control. It is known that
LC3B-II is closely associated with the membrane of autophagosomes
and AVOs are characteristic of autophagy (32–34).
Elevated expression of LC3B-II and number of AVOs reflected that
knockdown of Nrf2 caused an increased basal level of autophagy.
This indicated that Nrf2 was responsible for regulating basal
levels of autophagy in U251 cells. Our finding was in concordance
with previous reports in other tumors (35). Nevertheless, from our point of view,
further investigation still needs to be done because
Nrf2-responsive genes that stimulate autophagy are not well
known.
Earlier studies indicated that TMZ may exert its
cytotoxicity mainly by apoptosis pathway, while recent studies
investigating the cytotoxic effects of TMZ in malignant gliomas had
focused more on autophagy (32).
Glioma studies in vitro also showed that autophagy, not
apoptosis, was induced by TMZ (19). Autophagy induced by TMZ is important
for GBM chemotherapy. Therefore, the present study paid attention
to the effect of Nrf2 downregulation on autophagy induced by TMZ.
Substantial evidence showed that Nrf2 and apoptosis pathway may
cross-talk mediated by the Keap1-binding proteins such as
phosphoglycerate mutase 5 (PGAM5), prothymosin α (ProTα), fetal
Alz-50 clone 1 (FAC1), and p62 (SQSTM1) (25). Hence, in order to exclude the role
of Nrf2 on apoptosis induced by TMZ, we chose a certain dose and
time of TMZ based on previous studies. At this clinically
achievable dose (100 μM), TMZ induced autophagy, not apoptosis in
U251 cells (19,21,36).
With TMZ, we observed a significant decrease in cell viability and
increase in the level of LC3B-II compared to DMSO. Nevertheless,
the difference detected by flow cytometry lacked statistical
significance. This may be attributed to the error of operation and
poor state of the cells caused by toxicity of TMZ. We speculate
that lower cell viability may be connected with higher level of
autophagy induced by TMZ, which may be caused by autophagic cell
death induced by sustained TMZ treatment. Our data suggested that
Nrf2 limited the extent of autophagy induced by TMZ, which was in
agreement with the role on basal autophagy, also, Nrf2 protected
tumor cells from the death induced by TMZ.
The findings above illustrate the regulation of Nrf2
on autophagy induced by TMZ and its protective actions in tumor
cells. Consistent with our results in U251 cells, the drug
resistance of Nrf2 has also been demonstrated in a variety of
experimental models (37).
Transfection of Nrf2 siRNA of cisplatin-resistant human ovarian
cancer SK-OV cells showed exacerbated cytotoxicity to cisplatin.
Similar result was discovered in other cancer types and
chemotherapeutic agents (10). In
this study, knockdown of Nrf2 enhanced both the level of basal
autophagy and autophagy induced by TMZ. Nevertheless, how Nrf2
regulates autophagy remains unknown. Others have also reported that
Nrf2 negatively regulated autophagy induced by anticancer redox
agent MitoQ and the Nrf2-regulated enzyme NQO1 was partly
responsible for adjusting the level of autophagy (35).
Recent experimental evidence showed that p62, a
multidomain adapter protein, has a critical role in an oxidative
stress response pathway by its direct interaction with the
ubiquitin ligase adaptor Keap1, which resulted in constitutive
activation of Nrf2 (38,39). Another study presented evidence that
p62 was an Nrf2 target gene, which created a positive feedback loop
in the Nrf2-mediated transcriptional response (40). During autophagy, p62 acted not only
as an adaptor protein in selective autophagy of ubiquitinated
proteins, but also a substrate for degradation (41). Based on the above, we speculate that
molecular foundation for possible cross-talk between Nrf2 and
autophagy may be p62 or other Nrf2-targeted gene products, such as
NQO1. This question and whether the protective function of Nrf2 in
GBM cells after TMZ corresponds with autophagy regulated by Nrf2
are the issues of importance we would like to address in our future
work.
In conclusion, we demonstrated for the first time
that knockdown of Nrf2 enhanced autophagy induced by TMZ in U251
cells. We speculate that the combination of TMZ and the knockdown
of Nrf2 may point to a novel therapeutic opportunity for GBM to
enhance the antitumor effects of TMZ, which requires further
studies.
Acknowledgements
This work was supported by grants of the Jiangsu
Provincial Key Subject (X4200722) and the Jinling Hospital of
Nanjing (Grant no. 2012035). We are thankful to Professor Shao-jun
Jiang for his TEM assessment.
References
|
1
|
Germano IM, Emdad L, Qadeer ZA, Binello E
and Uzzaman M: Embryonic stem cell (ESC)-mediated transgene
delivery induces growth suppression, apoptosis and
radiosensitization, and overcomes temozolomide resistance in
malignant gliomas. Cancer Gene Ther. 17:664–674. 2010. View Article : Google Scholar
|
|
2
|
Ulasov IV, Sonabend AM, Nandi S, Khramtsov
A, Han Y and Lesniak MS: Combination of adenoviral virotherapy and
temozolomide chemotherapy eradicates malignant glioma through
autophagic and apoptotic cell death in vivo. Br J Cancer.
100:1154–1164. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Stupp R, Mason WP, van den Bent MJ, et al:
Radiotherapy plus concomitant and adjuvant temozolomide for
glioblastoma. New Engl J Med. 352:987–996. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jiang H, White EJ, Conrad C, Gomez-Manzano
C and Fueyo J: Autophagy pathways in glioblastoma. Method Enzymol.
453:273–286. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pan H, Wang H, Zhu L, Mao L, Qiao L and Su
X: The Role of Nrf2 in migration and invasion of human glioma cell
U251. World Neurosurg. Nov 7–2011.(Epub ahead of print).
|
|
6
|
Zhang DD: The Nrf2-Keap1-ARE signaling
pathway: The regulation and dual function of Nrf2 in cancer.
Antioxid Redox Signal. 13:1623–1626. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lau A, Villeneuve NF, Sun Z, Wong PK and
Zhang DD: Dual roles of Nrf2 in cancer. Pharmacol Res. 58:262–270.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Akhdar H, Loyer P, Rauch C, Corlu A,
Guillouzo A and Morel F: Involvement of Nrf2 activation in
resistance to 5-fluorouracil in human colon cancer HT-29 cells. Eur
J Cancer. 45:2219–2227. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang J, Stevens MF and Bradshaw TD:
Temozolomide: mechanisms of action, repair and resistance. Curr Mol
Pharmacol. 5:102–114. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang XJ, Sun Z, Villeneuve NF, et al: Nrf2
enhances resistance of cancer cells to chemotherapeutic drugs, the
dark side of Nrf2. Carcinogenesis. 29:1235–1243. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Levine B, Mizushima N and Virgin HW:
Autophagy in immunity and inflammation. Nature. 469:323–335. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Klionsky DJ, Abeliovich H, Agostinis P, et
al: Guidelines for the use and interpretation of assays for
monitoring autophagy in higher eukaryotes. Autophagy. 4:151–175.
2008. View Article : Google Scholar
|
|
13
|
Mathew R and White E: Autophagy in
tumorigenesis and energy metabolism: friend by day, foe by night.
Curr Opin Genet Dev. 21:113–119. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jeon SH, Kim SH, Kim Y, et al: The
tricyclic antidepressant imipramine induces autophagic cell death
in U-87MG glioma cells. Biochem Biophys Res Commun. 413:311–317.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Park EJ, Choi KS and Kwon TK:
β-Lapachone-induced reactive oxygen species (ROS) generation
mediates autophagic cell death in glioma U87 MG cells. Chem Biol
Interact. 189:37–44. 2011.
|
|
16
|
Jiang H, Gomez-Manzano C, Aoki H, et al:
Examination of the therapeutic potential of Delta-24-RGD in brain
tumor stem cells: role of autophagic cell death. J Natl Cancer
Inst. 99:1410–1414. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kaza N, Kohli L and Roth KA: Autophagy in
brain tumors: a new target for therapeutic intervention. Brain
Pathol. 22:89–98. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jakubowicz-Gil J, Langner E and Rzeski W:
Kinetic studies of the effects of Temodal and quercetin on
astrocytoma cells. Pharmacol Rep. 63:403–416. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kanzawa T, Germano IM, Komata T, Ito H,
Kondo Y and Kondo S: Role of autophagy in temozolomide-induced
cytotoxicity for malignant glioma cells. Cell Death Differ.
11:448–457. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Natsumeda M, Aoki H, Miyahara H, et al:
Induction of autophagy in temozolomide treated malignant gliomas.
Neuropathology. 31:486–493. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Katayama M, Kawaguchi T, Berger MS and
Pieper RO: DNA damaging agent-induced autophagy produces a
cytoprotective adenosine triphosphate surge in malignant glioma
cells. Cell Death Differ. 14:548–558. 2007. View Article : Google Scholar
|
|
22
|
Kato T, Natsume A, Toda H, et al:
Efficient delivery of liposome-mediated MGMT-siRNA reinforces the
cytotoxity of temozolomide in GBM-initiating cells. Gene Ther.
17:1363–1371. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Voss V, Senft C, Lang V, et al: The
pan-Bcl-2 inhibitor (−)-gossypol triggers autophagic cell death in
malignant glioma. Mol Cancer Res. 8:1002–1016. 2010.
|
|
24
|
Lefranc F: Glioblastomas are resistant to
apoptosis but less resistant to the autophagic process. Bull Mem
Acad R Med Belg. 162:331–338. 2007.(In French).
|
|
25
|
Stepkowski TM and Kruszewski MK: Molecular
cross-talk between the NRF2/KEAP1 signaling pathway, autophagy, and
apoptosis. Free Radic Biol Med. 50:1186–1195. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gao S, Yang XJ, Zhang WG, Ji YW and Pan Q:
Mechanism of thalidomide to enhance cytotoxicity of temozolomide in
U251-MG glioma cells in vitro. Chin Med J. 122:1260–1266.
2009.PubMed/NCBI
|
|
27
|
Klingelhoeffer C, Kammerer U, Koospal M,
et al: Natural resistance to ascorbic acid induced oxidative stress
is mainly mediated by catalase activity in human cancer cells and
catalase-silencing sensitizes to oxidative stress. BMC Complement
Altern Med. 12:612012. View Article : Google Scholar
|
|
28
|
Lau AT, Wang Y and Chiu JF: Reactive
oxygen species: current knowledge and applications in cancer
research and therapeutic. J Cell Biochem. 104:657–667. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bjorkoy G, Lamark T, Brech A, et al:
p62/SQSTM1 forms protein aggregates degraded by autophagy and has a
protective effect on huntingtin-induced cell death. J Cell Biol.
171:603–614. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mizushima N and Yoshimori T: How to
interpret LC3 immunoblotting. Autophagy. 3:542–545. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Aoki H, Kondo Y, Aldape K, et al:
Monitoring autophagy in glioblastoma with antibody against isoform
B of human microtubule-associated protein 1 light chain 3.
Autophagy. 4:467–475. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lin CJ, Lee CC, Shih YL, et al:
Resveratrol enhances the therapeutic effect of temozolomide against
malignant glioma in vitro and in vivo by inhibiting autophagy. Free
Radic Biol Med. 52:377–391. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chen H-Y and White E: Role of autophagy in
cancer prevention. Cancer Prev Res. 4:973–983. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Moreau K, Luo S and Rubinsztein DC:
Cytoprotective roles for autophagy. Curr Opin Cell Biol.
22:206–211. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Rao VA, Klein SR, Bonar SJ, et al: The
antioxidant transcription factor Nrf2 negatively regulates
autophagy and growth arrest induced by the anticancer redox agent
mitoquinone. J Biol Chem. 285:34447–34459. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Pan Q, Yang XJ, Wang HM, et al:
Chemoresistance to temozolomide in human glioma cell line U251 is
associated with increased activity of O6-methylguanine-DNA
methyltransferase and can be overcome by metronomic temozolomide
regimen. Cell Biochem Biophys. 62:185–191. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Cho JM, Manandhar S, Lee HR, Park HM and
Kwak MK: Role of the Nrf2-antioxidant system in cytotoxicity
mediated by anticancer cisplatin: implication to cancer cell
resistance. Cancer Lett. 260:96–108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Nezis IP and Stenmark H: p62 at the
interface of autophagy, oxidative stress signaling, and cancer.
Antioxid Redox Signal. 17:786–793. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Inami Y, Waguri S, Sakamoto A, et al:
Persistent activation of Nrf2 through p62 in hepatocellular
carcinoma cells. J Cell Biol. 193:275–284. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Jain A, Lamark T, Sjottem E, et al:
p62/SQSTM1 is a target gene for transcription factor NRF2 and
creates a positive feedback loop by inducing antioxidant response
element-driven gene transcription. J Biol Chem. 285:22576–22591.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Komatsu M, Kurokawa H, Waguri S, et al:
The selective autophagy substrate p62 activates the stress
responsive transcription factor Nrf2 through inactivation of Keap1.
Nat Cell Biol. 12:213–223. 2010.PubMed/NCBI
|