Introduction
Endometrial carcinoma is the most common malignancy
of the female genital tract, with an estimated 46,470 new cases and
8,120 deaths in the United States in 2011 (1). In other countries, such as China, its
incidence is on the rise (2). For
young women who wish to retain their fertility, only 50% show a
complete response to hormonal treatment (usually progestins), while
25% suffer a relapse after a temporary response (3). Patients with metastatic and refractory
cancer are at a significantly higher risk of morbidity and
mortality, with a median survival of 8–16 months (4). The poor prognosis is attributed to the
significant failure rate of traditional adjuvant therapy, including
radiotherapy, chemotherapy and hormonal therapy, following tumor
debulking (5). To improve the
outcome of patients with endometrial carcinoma, it is important to
investigate molecular pathways that are critical to the development
of the disease, and to identify novel targets for therapy.
The p21-activated kinases (Paks) are a family of
conserved non-receptor serine/threonine kinases that integrate
various signaling pathways that are vital to normal cell survival
and function (6). Based on their
sequence and functional similarities, six mammalian Paks are
subdivided into two groups: group I (pak1–3) and group II (pak4–6)
(7). Pak1 was originally identified
as a downstream effector of the Rho family small GTPases Cdc42 and
Rac1, which play a fundamental role in cytoskeleton reorganization
and motility (8,9). Subsequent studies have revealed that
Pak1 is also involved in regulating cell growth (10), apoptosis (11), adhesion (12) and angiogenesis (13), all of which are important for
tumorigenesis and metastasis.
Pak1 is widely expressed in a variety of normal
tissues, such as brain, muscle and spleen (14). Previous studies indicated that Pak1
expression is significantly increased in breast, colorectal and
ovarian cancer (15–17). Functional studies have also
implicated Pak1 in cell transformation and tumor cell invasion
(14). For instance, Pak1 has been
shown to promote non-small cell lung cancer (NSCLC) cell motility
and invasiveness by phosphorylation of CRK-II (c-Crk) (18). In addition, Pak1 was able to promote
mammary epithelial hyperplasia through phosphorylation and
transactivation of estrogen receptor-α (ERα) (10), which is also involved in the
pathogenesis of endometrial cancer (EC) (19). However, the expression pattern and
the function of Pak1 in EC remain unknown.
In this study, we sought to determine whether Pak1
is involved in the pathogenesis of EC. The expression of Pak1 in EC
cell lines and tissues was assessed and correlated with
clinicopathological parameters. We also investigated the effect of
Pak1 on EC cell proliferation, migration, invasion,
anchorage-independent growth and survival. Our results indicate
that Pak1 is a multifunctional regulator of the progression of
EC.
Materials and methods
Clinical samples
A total of 42 paraffin embedded EC tissues
(including 10 metastasis-positive lymph nodes) were retrieved from
32 patients with endometrial carcinoma at the Department of
Pathology at the International Peace Maternity and Child Health
Hospital, affiliated to Shanghai Jiao Tong University, China, from
December 2009 to November 2011. The diagnosis of each sample was
established by two pathologists according to the criteria of the
International Federation of Gynecology and Obstetrics (FIGO). These
patients ranged in age from 44 to 78 years (mean, 59 years). None
had hormone therapy, radiotherapy, or chemotherapy prior to
surgery.
Twenty-three normal endometrial samples were
obtained from patients who underwent a hysterectomy to treat other
diseases, such as myoma or adenomyosis. These patients ranged in
age from 27 to 64 years (mean, 46 years). Ten endometrial
hyperplasia tissues (5 without atypical cases and 5 with atypical
cases) were collected from patients who had undergone hysteroscope
examination due to irregular bleeding. The collection of clinical
samples was approved by the Ethics Committee of the Medical Faculty
of Shanghai Jiao Tong University, China. All patients provided
written informed consent.
Immunohistochemistry analysis
The immunohistochemical staining procedure was as
follows: formalin-fixed sections were deparaffinized with xylene
and hydrated in alcohol; for antigen retrieval, slides were
immersed in citrate buffer (pH 6.0) and boiled for 20 min. These
sections were then incubated in 0.3% H2O2 in
methanol for 30 min to inactivate endogenous peroxidase activity.
Non-specific reaction was blocked by incubating slides with 5%
bovine serum albumin for 30 min at room temperature. Sections were
incubated overnight at 4°C with a 1:100 dilution of rabbit
anti-Pak1 antibody (Abcam, Cambridge, MA, USA). Preimmune serum
from the same species was used as a negative control. Subsequently,
slides were incubated with biotinylated secondary antibody
(Mrbiotech, Emeryville, CA, USA) for 10 min.
The staining intensity was scored in the following
manner: 0, negative; 1, weak; 2, moderate; 3, strong. The staining
percentage was scored as: 0, 0–5%; 1, 5–25%; 2, 25–50%; 3, 50–75%;
and 4, 75–100%. We obtained a composite histoscore by multiplying
the values of the 2 parameters by a maximum of 12 (20).
Cell culture and isolation of human
endometrial epithelial cells
Four EC cell lines were purchased from ATCC
(American Type Culture Collection, Rockville, MD, USA). All cell
lines were cultured in DMEM/F12 (1:1) (Gibco, Auckland, New
Zealand) and supplemented with 10% fetal bovine serum (FBS)
(HyClone, Logan, UT, USA) in a 37°C, 5% CO2
incubator.
Primary human endometrial epithelial cells were
cultured as described by Zhang et al(21) and Osteen et al(22). Briefly, the normal endometrial
tissue was minced into small pieces and digested with 0.1%
trypsin-EDTA for 30 min at 37°C. Using serial filtration, tissue
debris was separated with 150-μm aperture sieves. Epithelial glands
were retained on 40-μm aperture sieves and backwashed with PBS. The
separated endometrial cells were cultured in DMEM/F12 (1:1)
supplemented with 10% FBS. All specimens were confirmed as normal
by histological analysis.
Plasmid and transfection
To stably express Pak1 in AN3CA and Ishikawa ells,
the cells were transfected with Pak1 expressing vector or the empty
control vector pEGFP-N1 (Clontech Laboratories, Palo Alto, CA, USA)
using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA), and then
selected with G418 (800 μg/ml; Sigma Chemical, St. Louis, MO, USA).
To stably silence Pak1, the cells were transfected with a set of
shRNA constructs (GeneCopoeia, Germantown, MD, USA) against human
Pak1 or the scrambled control vector psiHIV-U6 (GeneCopoeia), and
then selected with puromycin (0.5 μg/ml; Sigma Chemical).
Real-time qRT-PCR
Total RNA was extracted from cells using TRIzol
reagent (Invitrogen). cDNA was reverse-transcribed from total RNA
using Prime Script RT reagent kit (Takara Inc., Otsu, Japan).
Real-time PCR was performed using SYBR Premix Ex Taq (Takara Inc.)
and analyzed with an ABI Prism 7000 Sequence Detection System. The
oligonucleotide primers used included: Pak1 forward:
5′-AGTTTCAGAAGATGAG GATGATGA-3′, reverse: 5′-AATCACAGACCGTGTGTAT
ACAG-3′; and β-actin forward: 5′-CAGCCATGTACGTTG CTATCCAGG-3′,
reverse: 5′-AGGTCCAGACGCAGGATG GCATG-3′. Relative transcript
expression levels were calculated using the 2−ΔΔCT
method using β-actin as internal reference. All data were obtained
in triplicate in three independent experiments.
Western blot analysis
Cells were harvested with ProteoJET Mammalian Cell
Lysis Reagent (MBI Fermentas, Burlington, ON, Canada) with a
protease inhibitor cocktail (Roche Diagnostics, Basel,
Switzerland). A total of 60 μg protein was separated by SDS-PAGE,
and transferred to a polyvinylidene fluoride (PVDF) membrane.
Membranes were incubated with primary antibodies. Signal was
detected using BeyoECL Plus (Beyotime, Shanghai, China). Primary
antibodies included: rabbit anti-Pak1 (1:1000; Abcam), mouse
anti-β-actin (1:2000; ProteinTech Group, Chicago, IL, USA).
Proliferation assays
Cells (2×103 cells/well) were plated in
96-well plates. Cell number was measured every 24 h by MTT assay
following the manufacturer’s instructions (Beyotime). The OD value
of each well was detected at 570 nm. Medium was changed every other
day. Each experiment was repeated in triplicate.
In vitro migration and invasion
assays
Cell migration and invasion assays were performed
using culture medium-treated 6.5-mm transwell chambers with 8.0 μm
polycarbonate membranes (Corning LifeScience, Corning, NY, USA).
Matrigel (BD Biosciences, San Jose, CA, USA) coated transwell
membrane was used for invasion assay. Cells were seeded in
serum-free medium on the upper compartment of each transwell
chamber at a density between 2×105–1×106
cells/ml (200 μl/chamber). The lower chamber was filled with
culture medium containing 10% FBS. After 24 h of incubation,
non-migrated or non-invaded cells were removed from the upper
chamber. The invaded or migrated cells on the lower side of inserts
were fixed and stained with 5% crystal violet. Attached cells were
lysed with 10% ethylic acid, and absorbance (proportional to the
number of cells) was measured at 590 nm.
Soft agar colony assays
Cells were seeded in 0.3% top agar in growth medium
over a layer of 0.6% agar in a 6-well plate at a density of
1×104 cells/ml. After 3 weeks of incubation, colonies of
>50 cells were produced. Only colonies with >50 cells were
counted and photographed with an inverted microscope. All assays
were performed at least three times in triplicate.
Hoechst staining
Cells were seeded in 24-well plates and treated with
TNF-α (20 ng/ml; Peprotech, Rocky Hill, NJ, USA). At the indicated
time point, cells were washed in PBS and fixed in 4% (wt/vol)
paraformaldehyde for 16 h at 4°C. Cells were washed in PBS and
permeabilized with 0.2% Triton X-100 for 20 min at room
temperature. Permeabilized cells were then treated with Hoechst
33258 (10 mg/ml) for 30 min at room temperature. Photomicrographs
of cells were captured under fluorescence at a magnification of
×400.
Flow cytometry
Annexin V-TRITC apoptosis detection kit (KeyGen
Biotech, Shanghai, China) was used to identify the percentage of
apoptosis, following the manufacturer’s instructions. Briefly,
after stimulation, both floating and attached cells were collected
by low speed centrifugation. After washing in PBS, cells were
stained with Annexin V-TRITC and 7-AAD. Following incubation for 30
min at room temperature, the cells were analyzed by flow cytometry
(Becton Dickinson FACScan, Immunocytochemistry Systems, San Jose,
CA, USA). Tests were performed in triplicate.
Caspase activity assay
The enzymatic activity of caspase-3 was measured
according to the manufacturer’s instructions (Caspase-3 Activity
Assay kit, Beyotime). After 0–12 h of TNF-α exposure,
2×106 cells were collected and suspended in cell lysis
buffer. Cell lysates were mixed with reaction buffer at 37°C for 1
h. An increase in absorbance at 405 nm was used to quantify the
activation of caspase-3 activity. Caspase activity was expressed as
percentage of enzyme activity compared to control. Control groups
received 0.1% DMSO. All the experiments were carried out in
triplicate.
Statistical analysis
All tests were carried out with SPSS 16.0
(Microsoft, Redmond, WA, USA) or Prism (GraphPad, San Diego, CA,
USA). Each experiment was performed at least three times. Where
applicable, data are shown as the means ± SD. The two-tailed
Student’s t-test or Mann-Whitney U test were used for comparison of
data between two groups. Differences were considered statistically
significant at P<0.05.
Results
Pak1 is overexpressed in endometrial
cancer tissues and cell lines
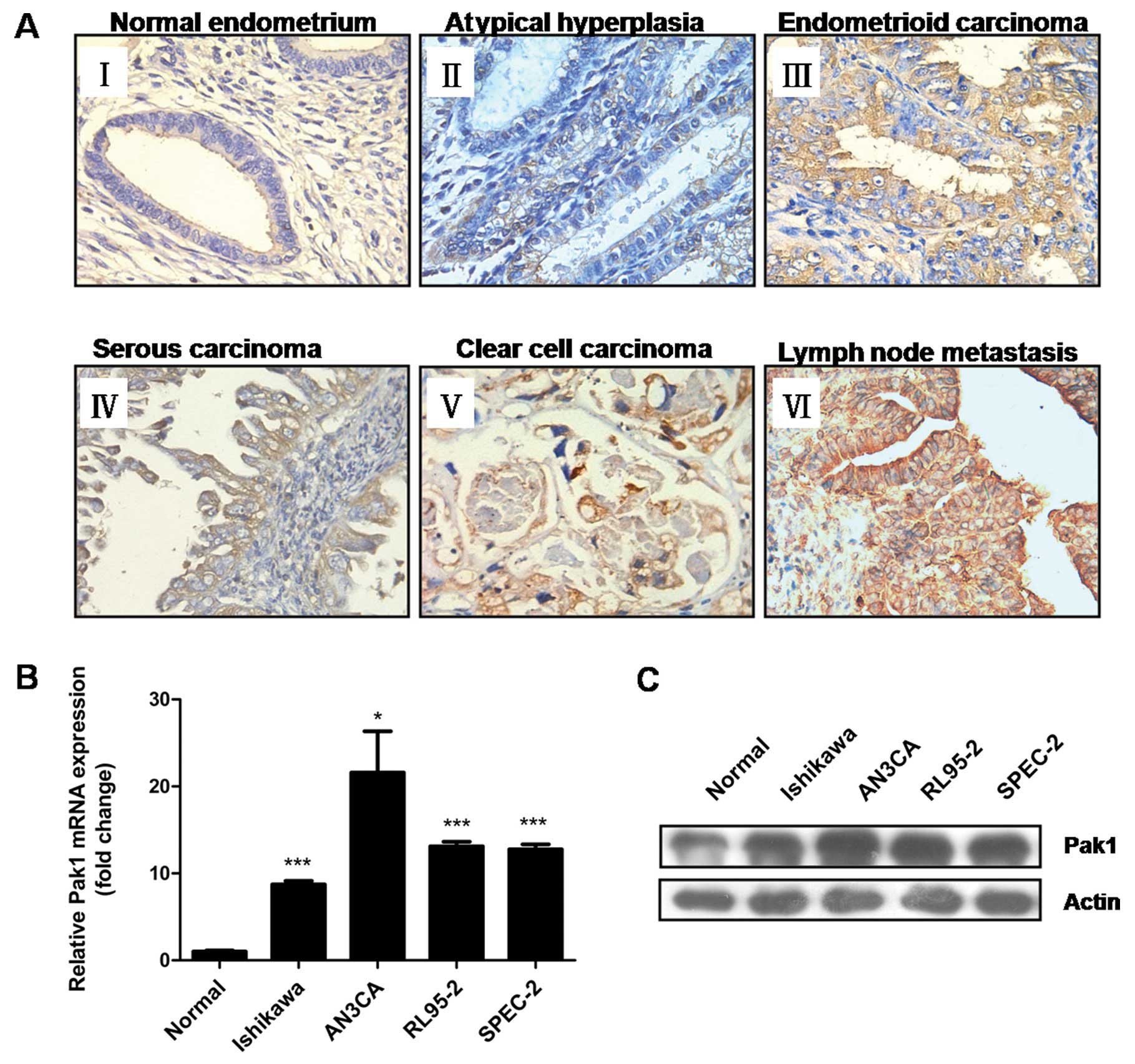
Immunochemistry staining showed that Pak1 protein
was predominantly localized to the cytoplasm of endometrial
epithelial cells. There was only weak or no staining in normal
endometrium, whereas moderate to strong Pak1 immunostaining was
found in endometrial hyperplasia and endometrial carcinoma tissues.
The most intense staining was found in lymph node metastases
(Fig. 1A).
To account for both stain intensity and the
uniformity of stain, a composite histoscore (percentage epithelium
stained × stain intensity) was calculated. Analyzed by the
Kruskal-Wallis rank test, cytoplasmic Pak1 expression (Pak1
composite histoscore) progressively increased in the epithelial
cells of endometrial hyperplasia, invasive carcinomas, and lymph
node metastases relative to the normal endometrium (P<0.001;
Table I).
 | Table ICorrelation of Pak1 expression with
clinicopathological parameters in endometrial carcinomas. |
Table I
Correlation of Pak1 expression with
clinicopathological parameters in endometrial carcinomas.
| | Pak1
histoscore |
|---|
| |
|
|---|
|
Characteristics | Case (n) | Mean ± SD | P-value |
|---|
| Diagnostic
categories |
| Normal | 23 | 4.70±2.23 | |
| Hyperplasia | 10 | 6.70±1.50 | |
| Carcinomas | 32 | 9.63±2.85 | |
| Lymph node
metastasisc | 10 | 12.00±0.00 | <0.001a |
| Age |
| <60 | 19 | 9.58±2.80 | |
| ≥60 | 13 | 9.69±3.04 |
0.966b |
| Histology |
| Endometrioid | 24 | 9.00±2.95 | |
| Non-endometrioid
(serous/clear) | 8 | 11.50±1.41 |
0.051b |
| FIGO stage |
| Early (I-II) | 24 | 8.83±2.88 | |
| Late (III-IV) | 8 | 12.00±0.00 | 0.013b |
| Histological
grade |
| Low (1–2) | 16 | 7.75±2.72 | |
| High (3) | 16 | 11.50±1.37 | <0.001b |
| Myometrial
invasion |
| ≤1/2 | 27 | 9.19±2.90 | |
| >1/2 | 5 | 12.00±0.00 | 0.043b |
| Involving
cervix |
| Negative | 27 | 9.19±2.90 | |
| Positive | 5 | 11.20±1.79 | 0.043b |
| Vascular space
invasion |
| Negative | 18 | 8.44±3.03 | |
| Positive | 14 | 11.14±1.70 | 0.014b |
We next explored the correlation of Pak1 expression
levels with clinicopathological parameters in endometrial
carcinomas. Significantly higher cytoplasmic Pak1 expression was
found in carcinomas of advanced stage (stages III and IV) and poor
histological differentiation (grade 3) (all P<0.05; Table I). Additionally, increased Pak1
expression was significantly associated with depth of myometrial
invasion, cervix involvement and vascular space invasion (all
P<0.05; Table I). However, there
was no significant difference between endometrioid and
non-endometrioid (serous and clear cell histological subtypes)
endometrial carcinomas (P=0.051; Table
I).
Moreover, using real-time qRT-PCR and
immunoblotting, we detected higher levels of Pak1 mRNA (Fig. 1B) and protein (Fig. 1C) in EC cell lines compared with
primary cultured normal endometrial epithelial cells. The elevated
Pak1 expression in endometrial carcinoma determined above suggests
that overexpression of Pak1 might contribute to the malignant
progression of human EC.
Pak1 promotes endometrial cancer cell
proliferation
Immuno-histochemical staining showed that Pak1
expression was associated with the progression of EC. To
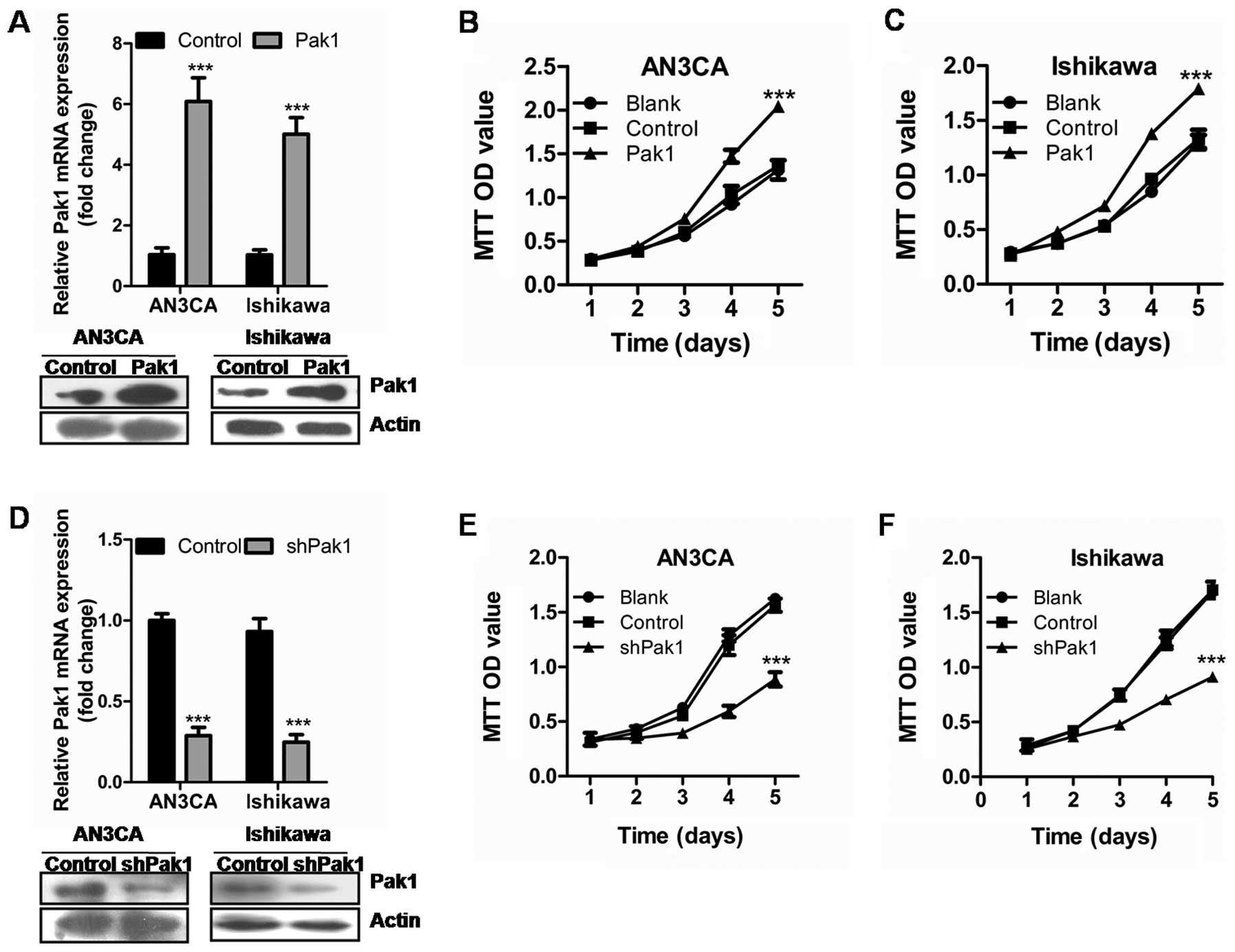
investigate the potential role of Pak1 in EC cell proliferation,
AN3CA and Ishikawa cells were stably transfected with Pak1
expressing vector or empty control vector (Fig. 2A). Proliferation assays revealed
that stable overexpression of Pak1 significantly induced AN3CA and
Ishikawa cell growth (Fig. 2B and
C) (P<0.001, P<0.001).
To further investigate the effect of Pak1 on EC cell
proliferation, we performed short hairpin RNAs (shRNA)-mediated
stable knockdown of Pak1 in AN3CA and Ishikawa cells. As shown in
Fig. 2D, the shRNAs led to an
almost complete loss of Pak1 in EC cell lines compared to the
scrambled control cells. Significantly reduced cell proliferation
was observed after Pak1 stable knockdown (Fig. 2E and F) (P<0.001,
P<0.001).
Pak1 promotes endometrial cancer cell
migration and invasion
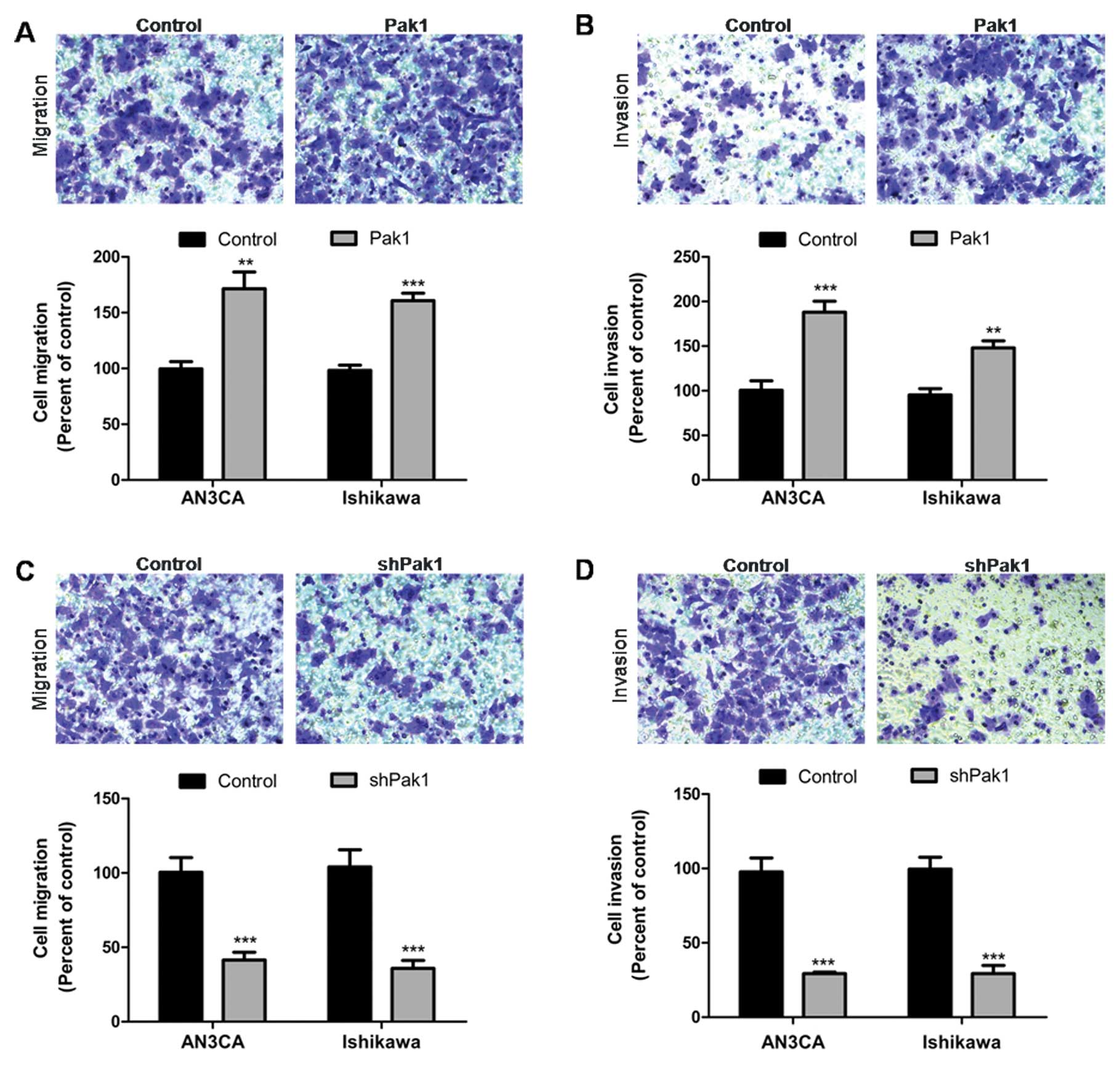
We next examined the effect of Pak1 on EC cell
migration and invasion. The transwell migration and invasion assays
were performed to study the migratory and invasive ability of EC
cells. As shown in Fig. 3A and B,
stable overexpression of Pak1 in AN3CA and Ishikawa cells
significantly increased cell migration and invasion compared with
cells transfected with control vector (P<0.001, P<0.001).
Conversely, shRNA-mediated knockdown of Pak1 in AN3CA and Ishikawa
cells resulted in significantly decreased cell migration and
invasion. Cells transfected with shRNA expression vector showed
>50% defective migration and invasion compared with cells
transfected with scrambled control vector (Fig. 3C and D) (P<0.001, P<0.001).
These results indicate that Pak1 is an important participant in EC
cell invasion and metastasis.
Pak1 promotes endometrial cancer cell
anchorage-independent growth
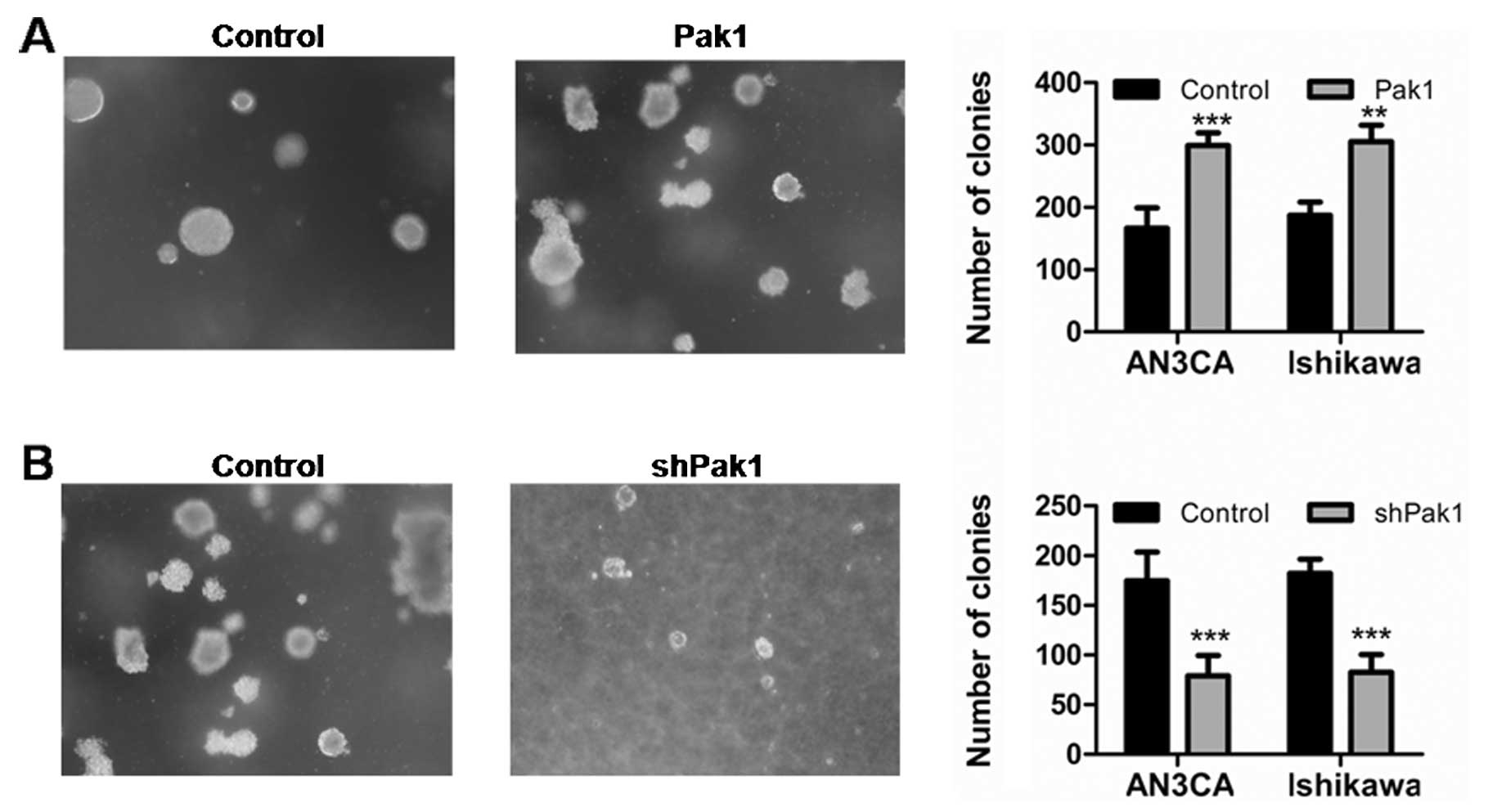
We next investigated whether Pak1 is required in
anchorage-independent growth, a hallmark of oncogenic
transformation (23). The soft agar
assays were performed to test the transformation of EC cells
(24). Pak1 expressing vector and
control vector transfected EC cells were plated in soft agar and
scored for colony formation after 3 weeks. As shown in Fig. 4A, there was a significant difference
in the number and size of colonies between vector-transfected
control cells and Pak1 overexpression AN3CA and Ishikawa cells
(P<0.001, P<0.01). By contrast, stable knockdown of Pak1
almost completely abolished colony formation of EC cells in soft
agar (Fig. 4B). These findings
indicate that Pak1 promoted EC cell anchorage-independent
growth.
Pak1 protects endometrial cancer cells
from apoptosis via inhibition of caspase-3 activation
Since overexpression of Pak1 results in increased
growth of EC cells, we sought to determine whether Pak1 also plays
a protective role against apoptosis in EC cells. To elucidate this
possibility, vector control cells and Pak1 overexpressing EC cells
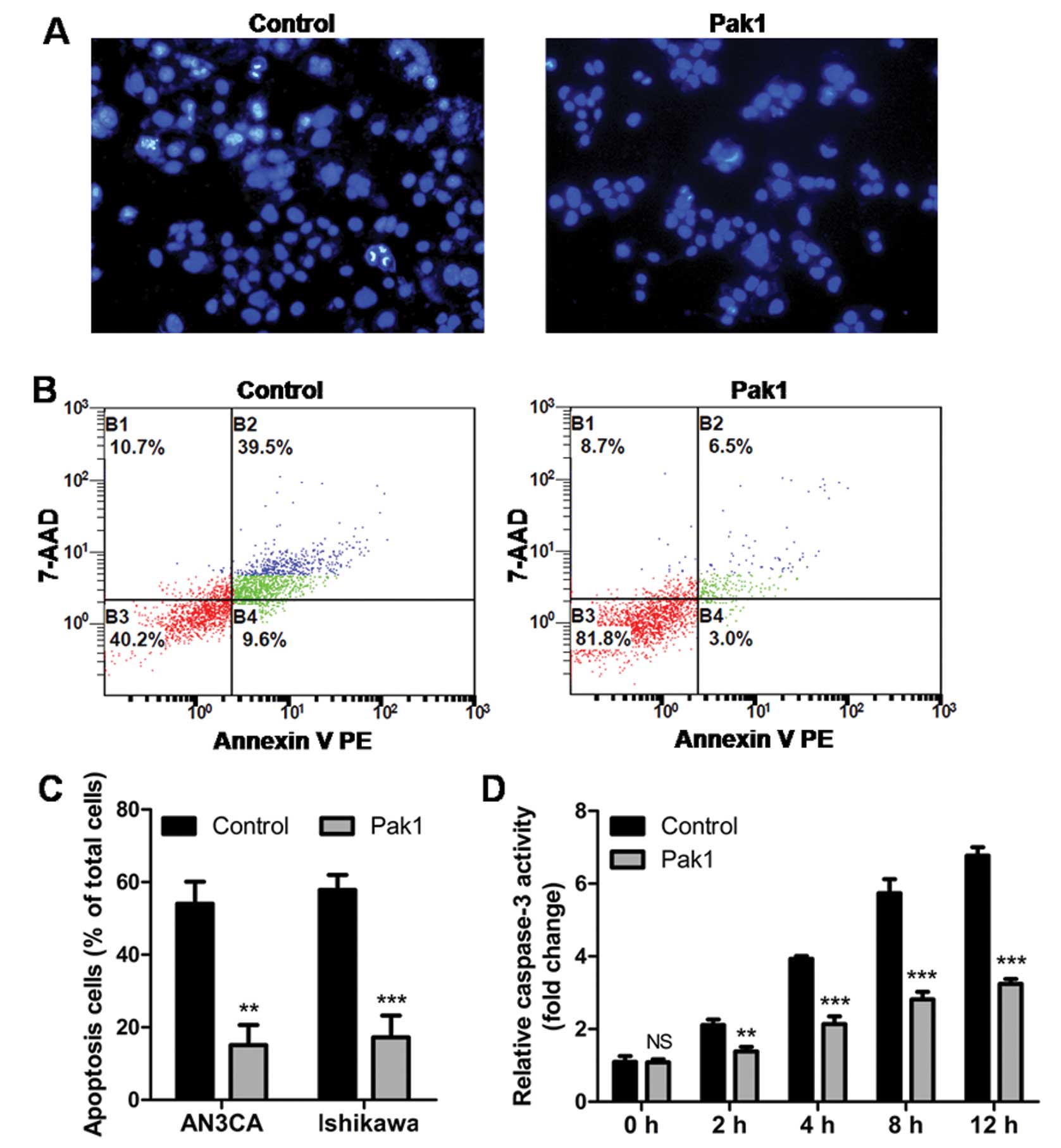
were treated with TNF-α. As shown in Fig. 5A, at 2 h, more control cells had
condensed nuclei compared to Pak1 overexpressing cells, as
determined by Hoechst staining. At 12 h, flow cytometry using
Annexin V and 7-AAD showed that a higher percentage of apoptotic
cells were observed in control cells compared with Pak1
overexpressing cells (Fig. 5B and
C). To further elucidate the mechanism by which Pak1 acts, we
tested the activation of caspase-3, a well known downstream target
of the TNF-α induced death pathway (25). We found that overexpression of Pak1
markedly abolished TNF-α induced caspase-3 activation (Fig. 5D). Our results indicate that, at
least in response to TNF-α, Pak1 plays an essential role in
protecting cells from apoptosis via inhibition of caspase-3
activation.
Discussion
Pak1 is upregulated and activated in several human
tumor types (6). However, the
expression of Pak1 in EC tissue and its relationship with
pathological parameters remains unknown. In the present study,
significantly increased Pak1 protein expression in clinical EC
samples and cell lines was detected relative to normal endometrial
tissues and primary cultured normal endometrial epithelial cells.
Furthermore, Pak1 protein expression is additionally increased in
lymph node metastases. These data indicate that Pak1 expression is
specifically increased in the most advanced lesions of EC. We found
a significantly positive correlation between Pak1 expression with
advanced disease stage and poor histological differentiation, which
further suggests that Pak1 expression is increased with EC
progression. Collectively, these data indicate that overexpression
of Pak1 might contribute to malignant progression of human EC.
Pak1 has been shown to be involved in the rapid
proliferation of cancer cells (17,26).
Pak1 promotes proliferation through the NF-κB-dependent pathway
that regulates cyclin D1 transcription in breast cancer (17). Knockdown of Pak1 in colorectal
cancer cells decreased proliferation and delayed the G1/S
cell-cycle transition (26). In
this study, we were able to confirm effect of Pak1 on proliferation
from two aspects: Pak1 ectopic expression significantly enhanced
proliferation whereas shRNA-mediated knockdown of Pak1 efficiently
impeded proliferation in EC cells. Pak1 is a common point of
convergence of growth factor signaling (27), and also directly interacts with
estrogen receptor (ER) in ER-positive breast cancer (28). It has been proposed that Pak1 may
constitute an important point of cross-talk between growth factors
and the ER (29). Cross-talk
between ER and growth factor signaling has emerged as a critical
factor in endocrine resistance in EC (30). If this is the case, the levels and
activation of Pak1 could potentially affect the action of
anti-estrogen therapies. In experimental breast cancer model
systems, overexpression of Pak1 correlates with the development of
ligand-independent stimulation of ERα signaling, which is
associated with sensitivity to tamoxifen (a selective
anti-estrogen, which has anti-estrogenic effects in the breast but
weak estrogenic effects in the endometrium) (28,31).
Therapies that target Pak1 may therefore represent a strategy to
increase the hormonal treatment response in EC.
We further investigated the effect of Pak1 on
enhancing cell migration and invasion of EC cell lines. Tumor cell
migration and invasion requires coordinated reorganization of the
actin cytoskeleton (32). Pak1 is
thought to regulate actin reorganization through several reported
substrates (6). For example, Pak1
phosphorylates LIM-kinase at threonine 508 within kinase’s
activation loop, and increases the phosphorylation and activation
of cofilin, thus regulating actin depolymerization (9). Inhibition of Pak1 abolishes
filamentous actin (F-actin) flow in the lamella, displaces myosin
IIA from the cell edge, and decreases focal adhesion turnover
(33). Pak1-knockdown cells have
significantly impaired migration and invasion (16,34,35).
In the present study, we also showed that downregulation of Pak1
expression reduced migration and invasion of EC cells.
Anchorage-independent growth is an important
hallmark of oncogenic transformation (23). The expression of kinase-active Pak1
mutants significantly stimulates anchorage-independent growth of
breast cancer cells in soft agar in a preferential
mitogen-activated protein kinase (MAPK)-sensitive manner (36). Pak1 also phosphorylates dynein light
chain 1 (DLC1) on serine 88, and promotes breast cancer cell
anchorage-independent growth and tumor formation (37). In addition, Pak1 simultaneously
activates MAPK and MET signaling, that permit human mammary
epithelial cells to form anchorage-independent colonies (38). Herein, we showed that knockdown of
Pak1 reduces, and overexpression enhances, the colony-forming
ability of EC cells. These results demonstrate the ability of Pak1
expression to stimulate the growth of EC cells in an
anchorage-independent manner, and indicate that Pak1 may play an
essential role in oncogenic transformation.
The efficacy of cancer treatments depends not only
on the cellular damage they cause but also on the cell’s ability to
respond to these damages by inducing the apoptotic response
(5). Pak1 has been shown to be
involved in the regulation of several important pro-apoptotic
pathways (6). For instance, Pak1
has been reported to directly phosphorylate and inactivate the
pro-apoptotic functions of B-cell lymphoma 2 (BCL2) antagonist of
cell death (BAD), which results in cell survival (39). In our study, Pak1 was found to
protect EC cells from apoptosis induced by TNF-α via inhibition of
caspase-3 activation. Alteration of this signal transduction
pathway leading to apoptosis has been reported in drug-resistant
cells (5). For instance, reduced
caspase-3 activity is associated with cisplatin resistance in human
ovarian cancer (40). In human
bladder carcinoma, activation of AKT increases paclitaxel
resistance by increasing Bad phosphorylation, leading to decreased
release of mitochondrial cytochrome c and caspase-3-mediated
apoptosis (41). In EC, caspase-3
expression is associated with patient survival (42). We found that expression of Pak1
leads to an inhibition of caspase-3 activation, which might be
associated with cisplatin and/or paclitaxel resistance in EC. A
recent study showed that dual inhibition of Pak1 and inhibitor of
apoptosis proteins (IAPs) efficiently increased effector caspase
activation and apoptosis of NSCLC cells (11). Thus, inhibition of Pak1 to enhance
chemosensitivity of EC may provide a promising therapeutic option
to putative targeted therapy. However, whether Pak1 is involved in
protecting cancer cells from apoptosis induced by chemotherapy
remains to be determined.
In summary, to the best of our knowledge, we showed
for the first time that overexpression of Pak1 plays important
roles at multiple stages of EC progression. Our results showed that
Pak1 protein is overexpressed in endometrial carcinoma,
particularly in the most advanced lesions. We also found that Pak1
affects EC cell proliferation, migration, invasion and
anchorage-independent growth in vitro. In addition, Pak1
regulates cell apoptosis through inhibition of caspase-3
activation. Our study demonstrated that Pak1 is a multifunctional
regulator of EC, and supports further characterization of Pak1 as a
therapeutic target.
Acknowledgements
This study was supported by Grants from the National
Natural Science Funds of China (nos. 81072139, 81172476 and
81272885). The authors thank Professor Hui-Juan Zhang and Dr Yuan
Liu for pathology revision.
Abbreviations:
|
BAD
|
Bcl-2-associated death promoter
|
|
BCL2
|
B-cell lymphoma 2
|
|
DLC1
|
dynein light chain 1
|
|
EC
|
endometrial cancer
|
|
ERα
|
estrogen receptor α
|
|
MAPK
|
mitogen-activated protein kinase
|
|
NSCLC
|
non-small cell lung cancer
|
|
Pak1
|
p21-activated kinase 1
|
|
qRT-PCR
|
quantitative reverse
transcription-polymerase chain reaction
|
|
TNF-α
|
tumor necrosis factor α
|
References
|
1
|
Siegel R, Ward E, Brawley O and Jemal A:
Cancer statistics, 2011: the impact of eliminating socioeconomic
and racial disparities on premature cancer deaths. CA Cancer J
Clin. 61:212–236. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang D, Zheng W, Wang SM, et al:
Estimation of cancer incidence and mortality attributable to
overweight, obesity, and physical inactivity in China. Nutr Cancer.
64:48–56. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chiva L, Lapuente F, Gonzalez-Cortijo L,
et al: Sparing fertility in young patients with endometrial cancer.
Gynecol Oncol. 111:S101–S104. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dedes KJ, Wetterskog D, Ashworth A, Kaye
SB and Reis-Filho JS: Emerging therapeutic targets in endometrial
cancer. Nat Rev Clin Oncol. 8:261–271. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chaudhry P and Asselin E: Resistance to
chemotherapy and hormone therapy in endometrial cancer. Endocr
Relat Cancer. 16:363–380. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kumar R, Gururaj AE and Barnes CJ:
p21-activated kinases in cancer. Nat Rev Cancer. 6:459–471. 2006.
View Article : Google Scholar
|
|
7
|
Eswaran J, Soundararajan M, Kumar R and
Knapp S: UnPAKing the class differences among p21-activated
kinases. Trends Biochem Sci. 33:394–403. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Manser E, Leung T, Salihuddin H, Zhao ZS
and Lim L: A brain serine/threonine protein kinase activated by
Cdc42 and Rac1. Nature. 367:40–46. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Edwards DC, Sanders LC, Bokoch GM and Gill
GN: Activation of LIM-kinase by Pak1 couples Rac/Cdc42 GTPase
signalling to actin cytoskeletal dynamics. Nat Cell Biol.
1:253–259. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang RA, Mazumdar A, Vadlamudi RK and
Kumar R: P21-activated kinase-1 phosphorylates and transactivates
estrogen receptor-alpha and promotes hyperplasia in mammary
epithelium. EMBO J. 21:5437–5447. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ong CC, Jubb AM, Haverty PM, et al:
Targeting p21-activated kinase 1 (PAK1) to induce apoptosis of
tumor cells. Proc Natl Acad Sci USA. 108:7177–7182. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhou H and Kramer RH: Integrin engagement
differentially modulates epithelial cell motility by RhoA/ROCK and
PAK1. J Biol Chem. 280:10624–10635. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bagheri-Yarmand R, Vadlamudi RK, Wang RA,
Mendelsohn J and Kumar R: Vascular endothelial growth factor
up-regulation via p21-activated kinase-1 signaling regulates
heregulin-beta1-mediated angiogenesis. J Biol Chem.
275:39451–39457. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Vadlamudi RK and Kumar R: P21-activated
kinases in human cancer. Cancer Metastasis Rev. 22:385–393. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Carter JH: Pak-1 expression increases with
progression of colorectal carcinomas to metastasis. Clin Cancer
Res. 10:3448–3456. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Siu MK, Wong ES, Chan HY, et al:
Differential expression and phosphorylation of Pak1 and Pak2 in
ovarian cancer: effects on prognosis and cell invasion. Int J
Cancer. 127:21–31. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Balasenthil S: p21-activated kinase-1
signaling mediates cyclin D1 expression in mammary epithelial and
cancer cells. J Biol Chem. 279:1422–1428. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rettig M, Trinidad K, Pezeshkpour G, et
al: AK1 kinase promotes cell motility and invasiveness through
CRK-II serine phosphorylation in non-small cell lung cancer cells.
PLoS One. 7:e420122012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Di Cristofano A and Ellenson LH:
Endometrial carcinoma. Annu Rev Pathol. 2:57–85. 2007.
|
|
20
|
Siu MK, Chan HY, Kong DS, et al:
p21-activated kinase 4 regulates ovarian cancer cell proliferation,
migration, and invasion and contributes to poor prognosis in
patients. Proc Natl Acad Sci USA. 107:18622–18627. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang L, Rees MC and Bicknell R: The
isolation and long-term culture of normal human endometrial
epithelium and stroma. Expression of mRNAs for angiogenic
polypeptides basally and on oestrogen and progesterone challenges.
J Cell Sci. 108:323–331. 1995.
|
|
22
|
Osteen KG, Hill GA, Hargrove JT and
Gorstein F: Development of a method to isolate and culture highly
purified populations of stromal and epithelial cells from human
endometrial biopsy specimens. Fertil Steril. 52:965–972.
1989.PubMed/NCBI
|
|
23
|
Stoker M, O’Neill C, Berryman S and Waxman
V: Anchorage and growth regulation in normal and virus-transformed
cells. Int J Cancer. 3:683–693. 1968. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Menendez JA, Vellon L, Colomer R and Lupu
R: Oleic acid, the main monounsaturated fatty acid of olive oil,
suppresses Her-2/neu (erbB-2) expression and synergistically
enhances the growth inhibitory effects of trastuzumab (Herceptin)
in breast cancer cells with Her-2/neu oncogene amplification. Ann
Oncol. 16:359–371. 2005. View Article : Google Scholar
|
|
25
|
Janicke RU, Sprengart ML, Wati MR and
Porter AG: Caspase-3 is required for DNA fragmentation and
morphological changes associated with apoptosis. J Biol Chem.
273:9357–9360. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Qing H, Gong W, Che Y, et al:
PAK1-dependent MAPK pathway activation is required for colorectal
cancer cell proliferation. Tumour Biol. 33:985–994. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gururaj AE, Rayala SK and Kumar R:
p21-activated kinase signaling in breast cancer. Breast Cancer Res.
7:5–12. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Rayala SK: P21-activated kinase 1
regulation of estrogen receptor-alpha activation involves serine
305 activation linked with serine 118 phosphorylation. Cancer Res.
66:1694–1701. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kumar R: Signaling intricacies take center
stage in cancer cells. Cancer Res. 65:2511–2515. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gururaj AE: Novel mechanisms of resistance
to endocrine therapy: genomic and nongenomic considerations. Clin
Cancer Res. 12:S1001–S1007. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kok M, Zwart W, Holm C, et al: PKA-induced
phosphorylation of ERα at serine 305 and high PAK1 levels is
associated with sensitivity to tamoxifen in ER-positive breast
cancer. Breast Cancer Res Treat. 125:1–12. 2010.PubMed/NCBI
|
|
32
|
Gimona M: Mechanics and dynamics of the
cytoskeleton: a special issue stemming from the 2008 ECF
Meeting/FEBS Workshop (Mechanics and Dynamics of the Cytoskeleton)
in Potsdam, Germany. Cell Motil Cytoskeleton. 66:ii–iii. 2009.
View Article : Google Scholar
|
|
33
|
Delorme-Walker VD, Peterson JR, Chernoff
J, et al: Pak1 regulates focal adhesion strength, myosin IIA
distribution, and actin dynamics to optimize cell migration. J Cell
Biol. 193:1289–1303. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
McCarty SK, Saji M, Zhang X, et al: Group
I p21-activated kinases regulate thyroid cancer cell migration and
are overexpressed and activated in thyroid cancer invasion. Endocr
Relat Cancer. 17:989–999. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li LH, Luo Q, Zheng MH, et al:
P21-activated protein kinase 1 is overexpressed in gastric cancer
and induces cancer metastasis. Oncol Rep. 27:1435–1442.
2012.PubMed/NCBI
|
|
36
|
Vadlamudi RK, Adam L, Wang RA, et al:
Regulatable expression of p21-activated kinase-1 promotes
anchorage-independent growth and abnormal organization of mitotic
spindles in human epithelial breast cancer cells. J Biol Chem.
275:36238–36244. 2000. View Article : Google Scholar
|
|
37
|
Vadlamudi RK, Bagheri-Yarmand R, Yang Z,
et al: Dynein light chain 1, a p21-activated kinase 1-interacting
substrate, promotes cancerous phenotypes. Cancer Cell. 5:575–585.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Shrestha Y, Schafer EJ, Boehm JS, et al:
PAK1 is a breast cancer oncogene that coordinately activates MAPK
and MET signaling. Oncogene. 31:3397–3408. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Schurmann A, Mooney AF, Sanders LC, et al:
p21-activated kinase 1 phosphorylates the death agonist bad and
protects cells from apoptosis. Mol Cell Biol. 20:453–461. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yang X, Zheng F, Xing H, et al: Resistance
to chemotherapy-induced apoptosis via decreased caspase-3 activity
and overexpression of antiapoptotic proteins in ovarian cancer. J
Cancer Res Clin Oncol. 130:423–428. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Szanto A, Bognar Z, Szigeti A, Szabo A,
Farkas L and Gallyas F Jr: Critical role of bad phosphorylation by
Akt in cytostatic resistance of human bladder cancer cells.
Anticancer Res. 29:159–164. 2009.PubMed/NCBI
|
|
42
|
Peiro G, Diebold J, Baretton GB, Kimmig R
and Lohrs U: Cellular apoptosis susceptibility gene expression in
endometrial carcinoma: correlation with Bcl-2, Bax, and caspase-3
expression and outcome. Int J Gynecol Pathol. 20:359–367. 2001.
View Article : Google Scholar : PubMed/NCBI
|