Introduction
Despite the progress of multimodal therapies for
non-small cell lung cancer (NSCLC) including surgery, radiotherapy,
and chemotherapy, the prognosis of patients remains poor.
Therefore, accurate molecular analyses are essential for the
establishment of an optimal treatment modality for each NSCLC
patient.
CpG islands stretch over 500–2000 bp. These
regulatory elements are located within promoter regions or the
first exons of genes (1). In a
normal genome, most cytosines of CpG sites in CpG islands are
unmethylated, while methylated in X chromosome inactivation. Gene
silencing associated with aberrant methylation of tumor-suppressor
genes (TSGs) is a common mechanism of tumorigenesis in NSCLC
(2). Global genomic DNA
hypomethylation also plays an important role in genomic instability
during tumorigenesis (3). Thus, the
mechanisms of both DNA hypermethylation of TSGs and global genomic
hypomethylation should be elucidated in order to understand and
manage NSCLC.
The methods used to measure methylation in cancer
cells or tissues varies among methylation-specific PCR (MSP)
(4), real-time MSP (5), direct sequencing followed by
subcloning, or combined bisulfite restriction analysis (COBRA)
(6). Although all are technically
sound, have widespread appeal and are frequently reported in the
literature (7–10), each has limitations. Since the
primers used for MSP and real-time MSP are designed to detect
methylation at all of the CpG sites in the binding sequence,
amplicons are generated only when the CpG island being examined is
completely methylated. Subcloning and COBRA procedures are labor
intensive and are thus not suitable for analysis of numerous
samples; the non-quantitative nature of these methods is also a
drawback.
In such a situation, the development of a simple,
easy and accurate methylation assay seems necessary. Pyrosequencing
is a nonelectrophoretic nucleotide extension sequencing technology
with various applications including quantitative methylation
analysis (11). Pyrosequencing has
previously been shown to be more precise than COBRA and MethyLight
(12). We previously demonstrated
that one bisulfite-treated DNA specimen can provide precise
measurement of a gene’s methylation value using pyrosequencing
(13). Since a single run of PCR
pyrosequencing can provide a reasonably precise measurement of gene
methylation in a given specimen, and the variation in gene
methylation values between different tissue sections from one tumor
is relatively small, the gene methylation level of a representative
tissue sample is most likely similar to the level of the entire
tumor.
In the present study, the methylation levels of long
interspersed nucleotide element 1 (LINE-1), Slit homolog 2 (SLIT2),
T-cell differentiation/myelin and lymphocyte protein gene (MAL),
and insulin-like growth factor binding protein 7 (IGFBP7, also
named insulin-like growth factor binding protein-related protein)
were assessed using sodium bisulfite conversion and a PCR
pyrosequencing assay. Since these genes are known to be TSGs in
various tumors, their role in tumorigenesis of NSCLC needs to be
clarified. We correlated our results with clinicopathological
features of the NSCLCs to learn more about how methylation profiles
differ in NSCLC subtypes.
Materials and methods
Patients and cell lines
Specimens were obtained from 56 serial patients who
underwent thoracic surgery at Kumamoto University Hospital from
2010 to 2012. None of these patients underwent preoperative
chemotherapy, radiotherapy, or chemoradiotherapy. Informed consent
for the research was obtained from each patient. The study design
was approved by the ethics review board of our university.
Eight NSCLC cell lines (HCC15, H63, H157, H460,
HUT15, HUT29, HUT70 and PC10) and one SCLC cell line (SBC5) were
used in this study. We established several of the cell lines used
(14), and others were purchased,
or generously donated by Dr Adi F. Gazdar of the University of
Texas Southwestern Medical Center.
5-Aza-2′-deoxycytidine (5-aza-CdR)
treatment
The 9 tumor cell lines were incubated in culture
medium with 1 μM of the demethylating agent 5-aza-CdR
(Sigma-Aldrich, USA) for 6 days, with replacement of the medium on
days 1, 3 and 5. Cells were harvested and RNA was extracted at day
6 as previously described (15).
Reverse transcriptase-PCR (RT-PCR)
A reverse transcriptase-PCR (RT-PCR) assay was used
to examine mRNA expression. Total RNA was extracted from samples
with TRIzol (Life Technologies, USA) following the manufacturer’s
instructions. The RT reaction was performed on 4 μg of total RNA
using Deoxyribonuclease I and the SuperScript II First-Strand
Synthesis System with the oligo(dT) Primer System (Life
Technologies), and aliquots of the reaction mixture were
subsequently used for PCR amplification. Real-time RT-PCR was
performed using SYBR Premix Ex Taq (Perfect Real-Time) (Takara,
Japan) and Thermal Cycler Dice® Real-Time System TP850
software. The results were analyzed using the comparative Ct method
(ΔΔCt) to compare the relative expression of each target gene
before and after 5-aza-CdR treatment according to the user manual,
and the ratio (5-aza-CdR/mRNA) was obtained. GAPDH was coamplified
with target genes and served as an internal standard. Primer
sequences were identical to those of the endogenous human target
genes as confirmed by a BLAST search. For all RNA examined, the
Gene Accession numbers are listed in parentheses. The real-time
RT-PCR primer sequences used were (sequences following F, forward
primer sequences; R, reverse primer sequences): SLIT2 (NM_004787.1)
F, GGTGTCCTCTGTGATGAAGAG and R, GTGTTTAGGACACACACCTCG; MAL
(NM_002371.2) F, GCAAGACGGCTTCACCTACAG and R, GCAGAGTG
GCTATGTAGGAGAACA; IGFBP7 (NM_001553.1) F, CAC TGGTGCCCAGGTGTACT and
R, TTGGATGCATGGCAC TCACA; GAPDH (NM_002046.3) F, TGAACGGGAAG
CTCACTGG and R, TCCACCACCCTGTTGCTGTA. We confirmed that genomic DNA
was not amplified with these primers, because all sequences cross
an intron.
DNA extraction and pyrosequencing
assay
Genomic DNA was obtained from primary tumors and
nonmalignant tissues by digestion with proteinase K, and
phenol/chloroform (1:1) extraction using PureLink™ Genomic DNA kits
(Invitrogen, USA).
DNA was treated with sodium bisulfite using EpiTect
Bisulfite kits according to the manufacturer’s instructions
(Qiagen, USA). Subsequent PCR and pyrosequencing for each gene was
performed using the PyroMark kit (Qiagen) as described previously
(16). The PCR conditions were 45
cycles of 95°C for 20 sec, 50°C for 20 sec and 72°C for 20 sec,
followed by 72°C for 5 min. The biotinylated PCR products were
purified and denatured prior to pyrosequencing with the
Pyrosequencing Vacuum Prep Tool (Qiagen) in the PyroMark Q96 MD
System (Qiagen). The nucleotide dispensation orders were: LINE-1
(X58075), ACTCAGTGTGTCAGTCAGTTAGTCTG; SLIT2,
GTCGTCGTTGATTAGAGTGATCTGTCG; MAL, ATGTC GTCATGATAGTCGAGTTCGTCG;
IGFBP7, GTCGTCGA TGTAGTTGTCG. The amount of C relative to the sum
of the amounts of C and T at each CpG site was calculated as a
percentage (i.e., 0–100). The average of the relative amounts of C
in the CpG sites was used as the overall methylation level of each
gene in a given tumor.
Detection of epidermal growth factor receptor (EGFR)
gene mutations was performed using the Cycleave method as reported
previously (17).
Statistical analysis
The Fisher’s exact test and Mann-Whitney U test were
applied to assess the association between categorical variables.
Statistical significance was defined as a P-value <0.05. All
P-values were two-sided. To determine the appropriate methylation
cut-off value, each methylation level was subdivided into two
cohorts using receiver operating characteristic (ROC) curve
analysis (18). All statistical
analyses were performed using SPSS 16.0 for Windows (SPSS, Inc.,
USA).
Results
Aberrant methylation of LINE-1, SLIT2,
MAL and IGFBP7 in primary tumors
We examined the methylation level of LINE-1, SLIT2,
MAL and IGFBP7 in 56 NSCLC tissues and matched non-malignant lung
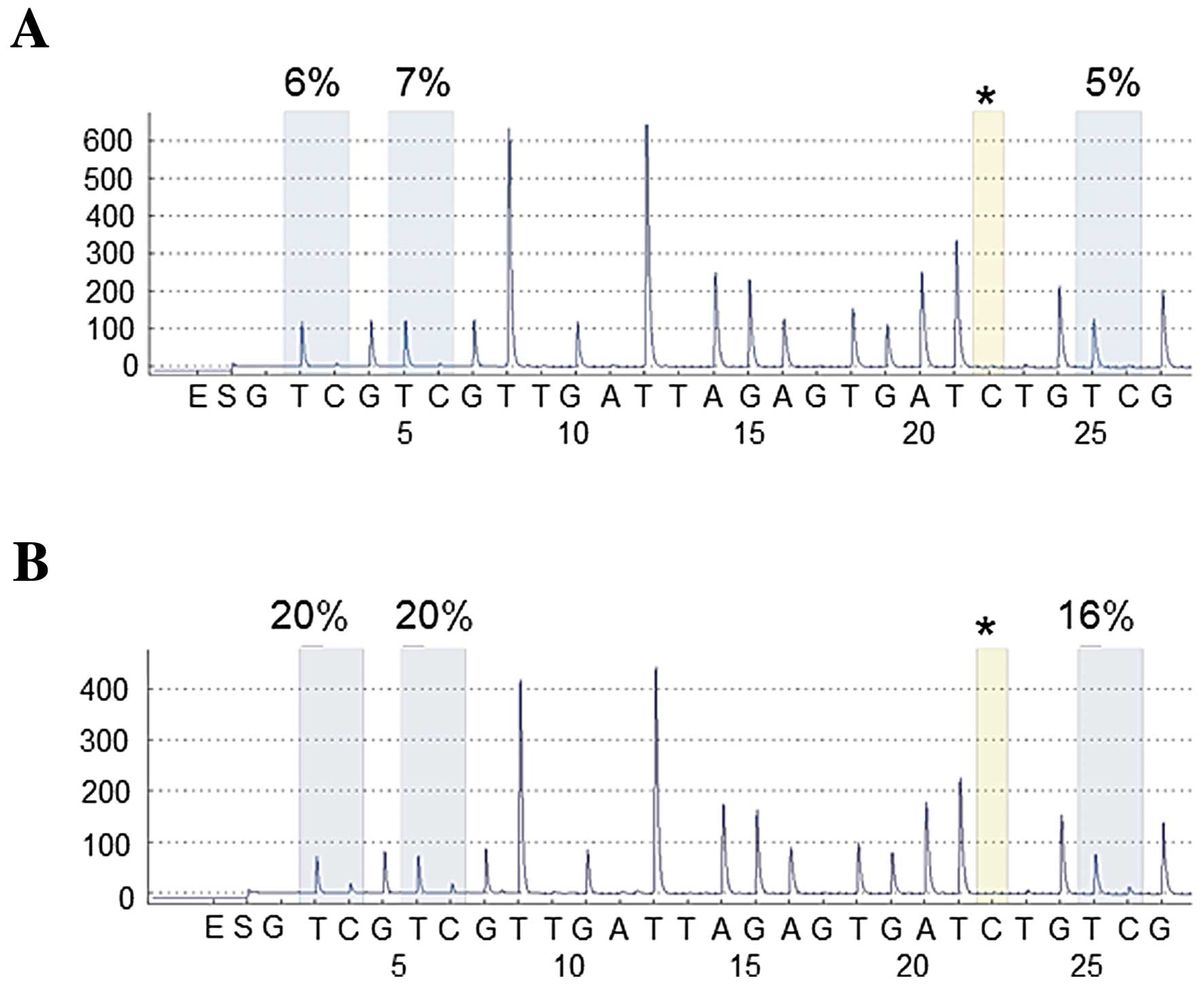
tissues, with representative cases shown in Fig. 1. Tumor tissues showed significantly
lower levels of LINE-1 methylation when compared with matched
non-malignant lung tissues (P<0.0001). In contrast, methylation
levels of SLIT2, MAL and IGFBP7 in tumor samples were higher when
compared with levels in matched non-malignant lung tissues
(P<0.0001, P=0.0010 and P<0.0001, respectively) (Table I). These data indicate that aberrant
methylation was a tumor-specific event in NSCLCs.
 | Table IAberrant methylation levels of genes
tested and cut-off values obtained from ROC curves. |
Table I
Aberrant methylation levels of genes
tested and cut-off values obtained from ROC curves.
| Genes | Cut-off value | Tumor tissue | Matched non-malignant
lung tissue | P-valueb | P-valuec |
|---|
|
|
|---|
| Median | Mean ± SD | Range | Methylation
(%)a | Median | Mean ± SD | Range | Methylation
(%)a |
|---|
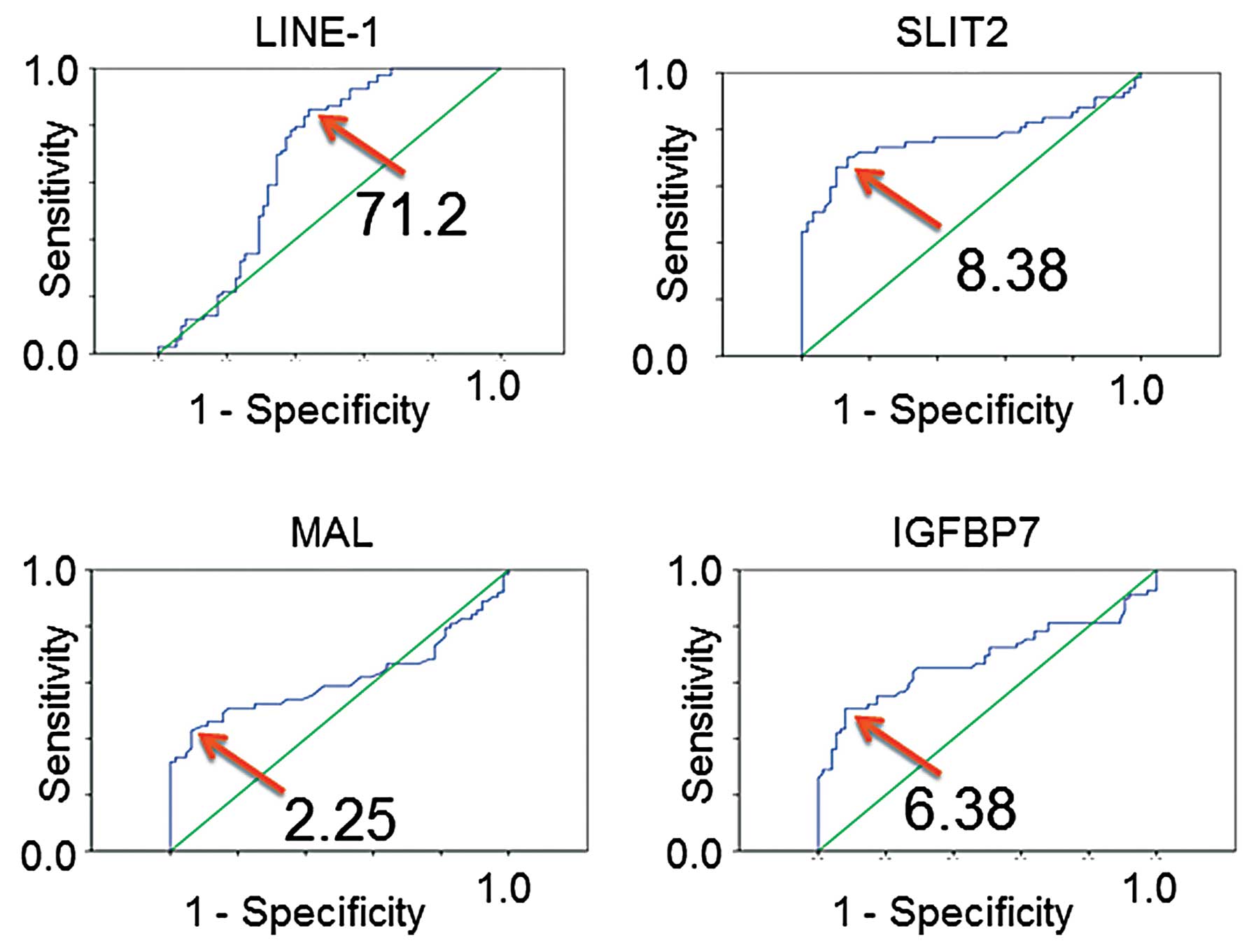
| LINE-1 | 71.2 | 71.0 | 67.9±10.0 | 35.0–80.4 | 31 (55) | 73.4 | 73.8±2.9 | 68.0–86.5 | 9 (16) | <0.0001 | <0.0001 |
| SLIT2 | 8.38 | 11.0 | 12.3±7.6 | 3.4–42.6 | 36 (64) | 6.1 | 6.6±2.0 | 3.4–11.8 | 8 (14) | <0.0001 | <0.0001 |
| MAL | 2.25 | 2.0 | 2.5±1.6 | 1.0–8.2 | 26 (46) | 1.8 | 1.8±0.3 | 1.0–2.6 | 4 (7) | 0.0010 | <0.0001 |
| IGFBP7 | 6.38 | 6.6 | 7.9±4.2 | 2.1–19.1 | 30 (54) | 4.9 | 5.1±1.0 | 3.4–9.3 | 3 (5) | <0.0001 | <0.0001 |
Studies that define cut-off values of aberrant
methylation are important as they are useful for further discussion
or cancer screening. Therefore, ROC curve analyses were conducted
to obtain cut-off values for methylation within each genomic
region. As shown in Fig. 2, cut-off
values were determined, and the frequencies of aberrant methylation
in both tumor and matched non-malignant lung tissue were obtained
(Table I). Comparisons of tumor
tissue with matched non-malignant lung tissue indicated that
aberrant methylation of LINE-1, SLIT2, MAL and IGFBP7 were all
tumor-specific events (all P<0.0001), despite the fact that
tumor tissues consist of mixtures of tumor cells and non-malignant
cells.
We next compared the clinicopathologic features with
values of the methylation level and frequencies of aberrant
methylation of LINE-1, SLIT2, MAL and IGFBP7 in NSCLCs (Table II). The level of LINE-1 methylation
was significantly lower in squamous cell carcinoma when compared
with the level in adenocarcinoma (P=0.0001). The methylation level
of SLIT2 was significantly higher in cases of chronic obstructive
pulmonary disease (COPD) when compared with the level in non-COPD
cases (P=0.05). Frequencies of MAL hypermethylation were
significantly higher in men than in women (P=0.035). In addition,
the methylation level and methylation frequency of MAL were both
significantly higher in smokers than in non-smokers (P=0.037;
P=0.032) and in patients without EGFR mutations (P=0.001; P=0.002)
compared to patients with EGFR mutations.
| Table IIAberrant methylation levels and
frequencies of LINE-1, SLIT2, MAL and IGFBP7 in NSCLC. |
Table II
Aberrant methylation levels and
frequencies of LINE-1, SLIT2, MAL and IGFBP7 in NSCLC.
| Patient
characteristics | 56 cases | LINE-1 | P-valuea
P-valueb | SLIT2 | P-valuea
P-valueb | MAL | P-valuea
P-valueb | IGFBP7 | P-valuea
P-valueb |
|---|
|
|
|
|
|---|
| Mean | n | % | Mean | n | % | Mean | n | % | Mean | n | % |
|---|
| Agec (years) |
| <72 | 27 | 70.4 | 15 | 56 | NS | 11.1 | 17 | 63 | NS | 2.51 | 13 | 48 | NS | 7.93 | 14 | 52 | NS |
| ≥72 | 29 | 65.6 | 16 | 55 | NS | 13.4 | 19 | 66 | NS | 2.57 | 13 | 45 | NS | 7.83 | 16 | 55 | NS |
| Gender |
| Male | 30 | 67.8 | 16 | 53 | NS | 11.8 | 20 | 67 | NS | 2.75 | 18 | 60 | NS | 8.75 | 19 | 63 | NS |
| Female | 26 | 68.1 | 14 | 54 | NS | 12.9 | 16 | 62 | NS | 2.31 | 8 | 31 | 0.035 | 6.88 | 11 | 42 | NS |
| Smoking
history |
| Smoker | 32 | 67.1 | 18 | 56 | NS | 12.5 | 20 | 63 | NS | 2.93 | 19 | 59 | 0.037 | 8.72 | 20 | 63 | NS |
| Never smoker | 24 | 69.1 | 13 | 54 | NS | 12.0 | 16 | 67 | NS | 2.03 | 7 | 29 | 0.032 | 6.76 | 10 | 42 | NS |
| COPD |
| (+) | 21 | 66.3 | 11 | 52 | NS | 14.8 | 15 | 71 | 0.05 | 2.89 | 10 | 48 | NS | 9.03 | 14 | 67 | NS |
| (−) | 35 | 68.9 | 20 | 57 | NS | 10.7 | 21 | 60 | NS | 2.34 | 16 | 46 | NS | 7.19 | 16 | 46 | NS |
| Histology |
| ADC | 47 | 70.1 | 25 | 53 | 0.0001 | 12.2 | 32 | 68 | NS | 2.44 | 21 | 45 | NS | 8.09 | 26 | 55 | NS |
| SCC | 9 | 56.7 | 6 | 67 | NS | 12.5 | 4 | 44 | NS | 3.08 | 5 | 56 | NS | 6.79 | 4 | 44 | NS |
| p-stage |
| IA | 31 | 69.5 | 15 | 48 | NS | 11.1 | 18 | 58 | NS | 2.44 | 12 | 39 | NS | 7.90 | 17 | 55 | NS |
| IB-III | 25 | 66.0 | 16 | 64 | NS | 13.7 | 18 | 72 | NS | 3.32 | 14 | 56 | NS | 7.86 | 13 | 52 | NS |
| EGFR
mutationd |
| (+) | 17 | 68.0 | 12 | 71 | NS | 11.5 | 13 | 76 | NS | 1.67 | 3 | 18 | 0.001 | 7.34 | 8 | 47 | NS |
| (−) | 18 | 69.7 | 9 | 50 | NS | 16.1 | 15 | 83 | NS | 3.61 | 13 | 72 | 0.002 | 9.41 | 13 | 72 | NS |
Aberrant methylation and mRNA expression
of SLIT2, MAL and IGFBP7 in the cell lines
We next examined the methylation level of LINE-1,
SLIT2, MAL and IGFBP7 in cell lines and determined whether the
observed levels were aberrant. According to the cut-off values
obtained from primary tumor sample analyses (Table I), LINE-1 hypomethylation was
present in 9 out of 9 cell lines. Hypermethylation of SLIT2, MAL
and IGFBP7 was present in 7 out of 9, 9 out of 9, and 7 out of 9
cell lines, respectively (Table
III). Next, expression of SLIT2, MAL and IGFBP7 was examined by
real-time RT-PCR before and after demethylation with 5-aza-CdR
treatment. By calculating the ratios of gene expression for the two
treatments, we determined that SLIT2, MAL and IGFBP7 expression was
higher after 5-aza-CdR treatment in 4 out of 9, 8 out of 9 and 7
out of 9 cell lines, respectively (Table III). The overall concordances
between gene expression ratios and methylation levels of SLIT2, MAL
and IGFBP7 were 67, 89 and 78% in lung cancer cell lines. The
observed discordance could not be dissolved even when we changed
the cut-off value of methylation (data not shown). Finally, we
compared the methylation level of LINE-1 with methylation levels of
other genes. There was no significant correlation between LINE-1
methylation and methylation of other genes (data not shown).
| Table IIIAberrant methylation and expression
change ratio of lung cancer cell lines. |
Table III
Aberrant methylation and expression
change ratio of lung cancer cell lines.
| LINE-1 | SLIT2 | MAL | IGFBP7 |
|---|
|
|
|
|
|
|---|
| Cell line | Methylation | Methylation | 5-Aza-CdR/mRNA | Methylation | 5-Aza-CdR/mRNA | Methylation | 5-Aza-CdR/mRNA |
|---|
| SBC5 | 48.5↓ | 72.5↑ | 2.46 | 93.5↑ | 37.5 | 98.0↑ | 69.00 |
| HCC15 | 33.4↓ | 44.6↑ | 1.73 | 6.4↑ | 23.5 | 7.6↑ | 7.84 |
| H63 | 33.6↓ | 3.50 | 0.40 | 3.4↑ | 3.37 | 35.7↑ | 0.47 |
| H157 | 40.4↓ | 22.8↑ | 1.91 | 26.4↑ | 30.0 | 64.7↑ | 1.26 |
| H460 | 46.7↓ | 10.1↑ | 0.09 | 11.2↑ | 25.0 | 52.7↑ | 2.57 |
| HUT15 | 29.1↓ | 28.4↑ | 0.33 | 4.5↑ | 2.40 | 4.7 | 0.54 |
| HUT29 | 36.1↓ | 2.35 | 1.00 | 2.6↑ | 1.20 | 3.3 | 1.72 |
| HUT70 | 69.6↓ | 83.0↑ | 4.14 | 3.8↑ | 25.9 | 95.6↑ | 5.82 |
| PC10 | 14.4↓ | 75.7↑ | 0.63 | 6.6↑ | 1.00 | 6.8↑ | 2.23 |
Discussion
LINE-1 methylation is a good indicator of cellular
5-methylcytosine levels, and thus is correlated with the global DNA
methylation level of cells or tissues. Previously, it was reported
that hypomethylation of LINE-1 measured by real-time MSP was a
prognostic marker in stage IA NSCLC (19). In our study, we first measured
LINE-1 methylation in NSCLC using pyrosequencing, and found that
LINE-1 was hypomethylated at higher frequencies in tumor tissues
when compared with the frequencies in non-malignant lung tissue of
NSCLCs. Although the number examined was relatively small and the
prognosis was not examined, we detected significantly lower
methylation levels in squamous cell carcinomas as compared to
adenocarcinomas. The role of LINE-1 hypomethylation in
carcinogenesis may be different between squamous cell carcinoma and
adenocarcinoma.
The re-expression of SLIT2, MAL and IGFBP7 that was
observed in lung cancer cell lines after treatment with a
demethylating agent exhibited good concordance with the methylation
status, suggesting that expression of those genes was downregulated
mainly by hypermethylation. However, differences in the methylation
level of each gene did not correlate linearly with the changes in
expression measured before and after demethylation. Moreover, the
discordance between methylation and expression remained even after
modifying the cut-off value of methylation levels. The expression
of each gene may be regulated partially by other mechanisms such as
histone acetylation, loss of heterozygosity or miRNA, or may be due
to toxicity of the demethylating agent.
SLIT2 suppresses tumor growth by coordinating
regulation of the β-catenin and PI3K/AKT pathways. The
hypermethylation of SLIT2 in NSCLC was examined by MSP (20). We also conducted similar
experiments, but were not able to obtain clear results (data not
shown), indicating that MSP can be influenced by conditions of the
experiments, laboratory or reagents. In a previous study, SLIT2
methylation was examined by genome-wide DNA methylation analysis,
using a microarray, in squamous cell carcinoma of the lung
(21). Since SLIT2 is a well-known
TSG and may play an important role in NSCLC tumorigenesis, we used
pyrosequencing to obtain accurate and quantitative measurements of
SLIT2 methylation in NSCLCs. Our results indicated that SLIT2 was
hypermethylated in both tumor tissues and tumor cell lines. In
addition, SLIT2 methylation levels were significantly higher in
COPD cases than in non-COPD cases. We previously examined the
molecular influence of COPD on the pathogenesis of NSCLC, and found
that the genetic profile of COPD-related NSCLC was quite different
from that of non-COPD-related NSCLC (22). Aberrant methylation of SLIT2 may
also play a different role between COPD and non-COPD NSCLCs.
MAL is a T-cell differentiation protein. The MAL
gene, which was initially isolated and cloned in 1987, maps to the
long arm of chromosome 2, encodes a 16.7-kDa membrane protein, and
contains a CpG island (23). MAL
has been shown to possess tumor-suppressor capabilities by
suppressing motility, invasion and tumorigenicity and by enhancing
apoptosis in esophageal cancer (24). Previous studies have indicated that
MAL is frequently methylated and plays a critical role in
tumorigenesis of various types of tumors (25–27).
However, to date there have been no studies examining MAL
methylation in NSCLC. We examined methylation using pyrosequencing.
Although the aberrant methylation level of the MAL gene was not
high in comparison to the other genes examined here, methylation
was significantly higher in tumor tissue than in non-malignant lung
tissue. Moreover, the frequency of MAL methylation was
significantly higher in males than in females, and both methylation
level and methylation frequency were significantly higher in
smokers and patients without EGFR mutations than in non-smokers and
patients with EGFR mutations. Thus, the hypermethylation of MAL may
play an important role in tumorigenesis of NSCLC, particularly in
male smokers.
IGFBP7 is expressed in normal epithelial cells and
acts as a tumor suppressor by inducing apoptosis in various types
of tumors (28–30). The IGFBP7 gene is aberrantly
methylated in various tumors (31,32),
and methylation was previously detected in 46 out of 90 lung
cancers examined by MSP (33). In
our study, we evaluated the methylation level of the IGFBP7 gene in
NSCLC using pyrosequencing, and detected hypermethylation in 30 out
of 56 NSCLCs. The frequency of IGFBP7 hypermethylation tha was
measured was quite similar to that reported by Chen et al,
although the methodology and population were different (33).
In summary, through accurate measurement of
methylation levels using a pyrosequencing assay, hypomethylation of
LINE-1 and hypermethylation of SLIT2, MAL, and IGFBP7 were
frequently detected in NSCLCs. Furthermore, these aberrant
methylation profiles were accompanied by distinct clinical
features. Further basic and clinical studies are required to
investigate the role of aberrant methylation in NSCLC. Ultimately,
further research in this area can contribute to the establishment
of personalized treatment for NSCLC.
Acknowledgements
This study was supported by a Grant-in-Aid for
Scientific Research from the Ministry of Education, Science,
Sports, Culture and Technology of Japan (23592069), and a grant
from the Smoking Research Foundation (2010–2012).
Abbreviations:
|
LINE-1
|
long interspersed nucleotide element
1
|
|
SLIT2
|
slit homolog 2
|
|
MAL
|
myelin and lymphocyte protein gene
|
|
IGFBP7
|
insulin-like growth factor binding
protein 7
|
|
NSCLC
|
non-small cell lung cancer
|
References
|
1
|
Takai D and Jones PA: Comprehensive
analysis of CpG islands in human chromosomes 21 and 22. Proc Natl
Acad Sci USA. 99:3740–3745. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Suzuki M and Yoshino I: Aberrant
methylation in non-small cell lung cancer. Surg Today. 40:602–607.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gaudet F, Hodgson JG, Eden A, et al:
Induction of tumors in mice by genomic hypomethylation. Science.
300:489–492. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Herman JG, Graff JR, Myohanen S, Nelkin BD
and Baylin SB: Methylation-specific PCR: a novel PCR assay for
methylation status of CpG islands. Proc Natl Acad Sci USA.
93:9821–9826. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lo YM, Wong IH, Zhang J, Tein MS, Ng MH
and Hjelm NM: Quantitative analysis of aberrant p16 methylation
using real-time quantitative methylation-specific polymerase chain
reaction. Cancer Res. 59:3899–3903. 1999.PubMed/NCBI
|
|
6
|
Xiong Z and Laird PW: COBRA: a sensitive
and quantitative DNA methylation assay. Nucleic Acids Res.
25:2532–2534. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Suzuki M, Toyooka S, Shivapurkar N, et al:
Aberrant methylation profile of human malignant mesotheliomas and
its relationship to SV40 infection. Oncogene. 24:1302–1308. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Takahashi T, Shivapurkar N, Riquelme E, et
al: Aberrant promoter hypermethylation of multiple genes in
gallbladder carcinoma and chronic cholecystitis. Clin Cancer Res.
10:6126–6133. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tamura H, Suzuki M, Moriya Y, et al:
Aberrant methylation of N-methyl-D-aspartate receptor type 2B
(NMDAR2B) in non-small cell carcinoma. BMC Cancer. 11:2202011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tian L, Suzuki M, Nakajima T, et al:
Clinical significance of aberrant methylation of prostaglandin E
receptor 2 (PTGER2) in nonsmall cell lung cancer: association with
prognosis, PTGER2 expression, and epidermal growth factor receptor
mutation. Cancer. 113:1396–1403. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ronaghi M: Pyrosequencing sheds light on
DNA sequencing. Genome Res. 11:3–11. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sepulveda AR, Jones D, Ogino S, et al: CpG
methylation analysis - current status of clinical assays and
potential applications in molecular diagnostics: a report of the
Association for Molecular Pathology. J Mol Diagn. 11:266–278. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Iwagami S, Baba Y, Watanabe M, et al:
Pyrosequencing assay to measure LINE-1 methylation level in
esophageal squamous cell carcinoma. Ann Surg Oncol. 19:2726–2732.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yamaji H, Iizasa T, Koh E, et al:
Correlation between interleukin 6 production and tumor
proliferation in non-small cell lung cancer. Cancer Immunol
Immunother. 53:786–792. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Suzuki M, Sunaga N, Shames DS, Toyooka S,
Gazdar AF and Minna JD: RNA interference-mediated knockdown of DNA
methyltransferase 1 leads to promoter demethylation and gene
re-expression in human lung and breast cancer cells. Cancer Res.
64:3137–3143. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Baba Y, Huttenhower C, Nosho K, et al:
Epigenomic diversity of colorectal cancer indicated by LINE-1
methylation in a database of 869 tumors. Mol Cancer. 9:1252010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yoshida K, Yatabe Y, Park JY, et al:
Prospective validation for prediction of gefitinib sensitivity by
epidermal growth factor receptor gene mutation in patients with
non-small cell lung cancer. J Thorac Oncol. 2:22–28. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Moses LE, Shapiro D and Littenberg B:
Combining independent studies of a diagnostic test into a summary
ROC curve: data-analytic approaches and some additional
considerations. Stat Med. 12:1293–1316. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Saito K, Kawakami K, Matsumoto I, Oda M,
Watanabe G and Minamoto T: Long interspersed nuclear element 1
hypomethylation is a marker of poor prognosis in stage IA non-small
cell lung cancer. Clin Cancer Res. 16:2418–2426. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dallol A, Da Silva NF, Viacava P, et al:
SLIT2, a human homologue of the Drosophila Slit2 gene, has
tumor suppressor activity and is frequently inactivated in lung and
breast cancers. Cancer Res. 62:5874–5880. 2002.
|
|
21
|
Kwon YJ, Lee SJ, Koh JS, et al:
Genome-wide analysis of DNA methylation and the gene expression
change in lung cancer. J Thorac Oncol. 7:20–33. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Suzuki M, Wada H, Yoshino M, et al:
Molecular characterization of chronic obstructive pulmonary
disease-related non-small cell lung cancer through aberrant
methylation and alterations of EGFR signaling. Ann Surg Oncol.
17:878–888. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Puertollano R, Martin-Belmonte F, Millan
J, et al: The MAL proteolipid is necessary for normal apical
transport and accurate sorting of the influenza virus hemagglutinin
in Madin-Darby canine kidney cells. J Cell Biol. 145:141–151. 1999.
View Article : Google Scholar
|
|
24
|
Mimori K, Shiraishi T, Mashino K, et al:
MAL gene expression in esophageal cancer suppresses motility,
invasion and tumorigenicity and enhances apoptosis through the Fas
pathway. Oncogene. 22:3463–3471. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lind GE, Ahlquist T, Kolberg M, et al:
Hypermethylated MAL gene - a silent marker of early colon
tumorigenesis. J Transl Med. 6:132008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Buffart TE, Overmeer RM, Steenbergen RD,
et al: MAL promoter hypermethylation as a novel prognostic marker
in gastric cancer. Br J Cancer. 99:1802–1807. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Overmeer RM, Henken FE, Bierkens M, et al:
Repression of MAL tumour suppressor activity by promoter
methylation during cervical carcinogenesis. J Pathol. 219:327–336.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ruan W, Xu E, Xu F, et al: IGFBP7 plays a
potential tumor suppressor role in colorectal carcinogenesis.
Cancer Biol Ther. 6:354–359. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Vizioli MG, Sensi M, Miranda C, et al:
IGFBP7: an oncosuppressor gene in thyroid carcinogenesis. Oncogene.
29:3835–3844. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mutaguchi K, Yasumoto H, Mita K, et al:
Restoration of insulin-like growth factor binding protein-related
protein 1 has a tumor-suppressive activity through induction of
apoptosis in human prostate cancer. Cancer Res. 63:7717–7723.
2003.
|
|
31
|
Smith P, Nicholson LJ, Syed N, et al:
Epigenetic inactivation implies independent functions for
insulin-like growth factor binding protein (IGFBP)-related protein
1 and the related IGFBPL1 in inhibiting breast cancer phenotypes.
Clin Cancer Res. 13:4061–4068. 2007. View Article : Google Scholar
|
|
32
|
Ye F, Chen Y, Knosel T, et al: Decreased
expression of insulin-like growth factor binding protein 7 in human
colorectal carcinoma is related to DNA methylation. J Cancer Res
Clin Oncol. 133:305–314. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chen Y, Cui T, Knosel T, Yang L, Zoller K
and Petersen I: IGFBP7 is a p53 target gene inactivated in human
lung cancer by DNA hypermethylation. Lung Cancer. 73:38–44. 2011.
View Article : Google Scholar : PubMed/NCBI
|