Introduction
Poly(ADP-ribosyl)ation is a
NAD+-dependent post-transcriptional modification of
chromosomal proteins mediated by poly(ADP-ribose) polymerases
(PARPs) and poly(ADP-ribose) glycohydrolase (PARG) (1). Reversible poly(ADP-ribosyl)ation of
chromosomal proteins has been suggested to play important roles in
various biological processes, including DNA replication (2,3),
repair (4–6), spindle assembly (7), transcription (8), telomere and chromosomal maintenance
(9) and epigenetic gene regulation
(10). Previous studies have
suggested that poly(ADP-ribose) metabolism is associated with
differentiation and proliferation (11,12),
cell death (13) and apoptosis
(14,15). Thus, poly(ADP-ribosyl)ation is
significantly involved in various biological activities, suggesting
that it requires precise controlling systems to adjust the amount
and length of poly(ADP-ribose) in eukaryotic cells.
Previous studies indicated that several
transcription factors, including Sp1 (16), YY1 (17), and NF-κB (18), are poly(ADP-ribosyl)ated and reduce
the transcription activity (16).
If there are binding sites for these transcription factors in the
promoter region, transcription might be affected by the
poly(ADP-ribosyl)ation. To date, the promoter regions of the human
PARP1(19) and
PARG(20,21) genes have been isolated and
characterized. In this study, we investigated the effect of short
interfering RNAs (siRNAs) for the human PARG cDNA on
poly(ADP-ribosyl)ation and PARP1 gene expression in HeLa S3
cells. The results showed that expression and promoter activity of
the PARP1 gene were reduced by the knockdown of the
PARG gene and that the amounts of poly(ADP-ribose) in the
cells did not increase compared to the control cells. Moreover,
PARG knockdown cells showed stronger cell death sensitivity
to staurosporine (STS) than the control cells, suggesting that
retarded turnover of poly(ADP-ribose)-NAD+ metabolism
might induce intracellular apoptosis signals.
It is well known that PARP1 activity is
downregulated by its augmented auto-poly(ADP-ribosyl)ation
(22,23), and artificially accumulated
poly(ADP-ribose) induces apoptosis (13). Collectively, our results indicate
that reduced poly(ADP-ribose) degradation subsequently suppresses
transcription of the PARP1 gene to escape excessive
poly(ADP-ribose) accumulation, thereby achieving a balance in
poly(ADP-ribose) levels for cell survival. Therefore,
poly(ADP-ribose) may act as a dual regulator for PARP1 activity not
only at the post-translational level but also at the
transcriptional level. Hence, we propose a molecular mechanism that
prevents cells from accumulating excess amounts of poly(ADP-ribose)
by regulating transcription of the PARP1 gene.
Materials and methods
Cell culture
Human cervical carcinoma (HeLa S3) cells (24) were grown in Dulbecco’s modified
Eagle’s medium (DMEM; Nacarai, Tokyo, Japan), supplemented with 10%
fetal bovine serum (FBS) (Sanko Pure Chemicals, Tokyo, Japan) and
penicillin-streptomycin at 37°C in a humidified atmosphere with 5%
CO2.
Transfection of siRNA
The ON-TARGETplus SMARTpool siRNAs used for
knockdown of the human PARG gene were purchased from Thermo
Fisher Scientific Inc. (Lafayette, CO, USA). They were introduced
into HeLa S3 cells with DharmaFECT Transfection reagent following
the manufacturer’s protocol (Thermo Fisher Scientific). In brief, 2
μM siRNA (50 μl) were added to serum-free DMEM (50 μl) in one tube,
and DharmaFECT1 (1.5 μl) was added to 98.5 μl of serum-free medium
in the other tube. They were gently mixed and incubated for 5 min
at room temperature, and were then combined, mixed and further
incubated for 20 min at room temperature. Subsequently, complete
medium (800 μl) was added and cells were cultivated with the medium
in a 35-mm culture dish.
Cell viability MTS assay
An MTS assay was performed according to the
manufacturer’s instructions. In brief, mock- or siRNA-transfected
cells were cultured in microtiter plate wells. MTS solution (20 μl)
(Promega, Madison, WI, USA) was added to each well (containing 100
μl of cell culture) and incubated for 3 h in a 37°C, 5%
CO2-humidified incubator. Then, the absorbance at 492 nm
was measured by a microtiter plate reader (Thermo Electron Corp.,
Vantaa, Finland) and normalized by the absorbance at 630 nm.
Reverse transcriptase and quantitative
real-time polymerase chain reaction (RT-qPCR)
RT-qPCR was carried out as previously described
(24). First-strand cDNAs were
synthesized with ReverTra Ace (Toyobo Corp., Tokyo, Japan), random
primers (Takara, Kyoto, Japan) and total RNAs were extracted from
HeLa S3 cells. A primer pair to amplify the human GAPDH
cDNAs was previously reported (24), and those for amplifying the
PARP1 and PARG cDNAs were: hPARP1S514,
5′-GCAGAGTATGCCAAGTCCAACAG-3′ and hPARP1AS813,
5′-ATCCACCTCATCGCCTTTTC-3′; and hPARG-S,
5′-ATGTGTAAGTGGCAAAATGAAGGG-3′ and hPARG-A952,
5′-CTTCTCTGGCCTGTTCATCTTC-3′, respectively. Real-time PCR analysis
was carried out using the Mx3000P Real-Time QPCR system
(Stratagene, La Jolla, CA, USA) as previously described (24). For PCR amplification, cDNAs were
amplified using SYBR-Green real-time PCR Master Mix (Toyobo) and
0.3 μM of each primer pair. Amplification of the PARP1 cDNA
was carried out, starting with an initial step for 1 min at 94°C,
followed by 42 cycles (94°C 30 sec, 55°C 30 sec and 72°C 1 min).
The conditions for amplification of the PARG and
GAPDH cDNAs were 1 min at 94°C, followed by 42 cycles (94°C
15 sec, 55°C 10 sec, and 72°C 15 sec).
Western blot analysis
Western blotting was carried out as previously
described (24,25) with antibodies against PARP1 (Santa
Cruz Biotechnology, Santa Cruz, CA, USA) and PAR (Calbiochem,
Darmstadt, Germany) followed by the addition of horseradish
peroxidase (HRP)-conjugated secondary antibody (Calbiochem). Signal
intensities were quantified with a LAS4000 system and Multi Gauge
Software (Fuji Film, Tokyo, Japan).
Construction of luciferase (Luc) reporter
plasmids
Luc reporter plasmids carrying 75-bp of the human
PARG promoter regions were designated pKBST-Δ6 (21). The 5′-flanking regions of the human
PARP1 gene were obtained by PCR with PrimeStar Taq
polymerase (Takara) and the template genomic DNA from HeLa S3 cells
as previously described (26). The
sense and antisense primers used for PCR were: hPARP1-2660,
5′-TCGGTACCGGGTCCTCCAAAGAGCTAC-3′; and AhPARP1-2895,
5′-ATCTCGAGCCGCCACCGAACACGC CGC-3′, respectively. The amplified DNA
fragments were digested with KpnI and XhoI and
ligated into the MCS of the pGL4-basic (pGL4.10[luc2]), vector
(Promega) to make pGL4-PARP1. Deletion derivatives, pGL4-PARP1Δ1
and pGL4-PARP1Δ2, were generated by PCR with pGL4-PARP1 as the
template and primer sets: hPARP1-2851, 5′-TCGGTA
CCGCCAGGCATCAGCAATCTA-3′ and AhPARP1-2895; and hPARP1-2660 and
AhPARP1-2851, 5′-ATCTCGAGTA GATTGCTGATGCCTGGC-3′, respectively. The
nucleotide sequences of the PCR products were determined by a DNA
Sequencing system (Applied Biosystems, Foster City, CA, USA) with
Rv (5′-TAGCAAAATAGGCTGTCCCC-3′) and GL
(5′-CTTTATGTTTTTGGCGTCTTCC-3′) primers.
Transient transfection and Luc
assays
Plasmid DNAs were transfected into HeLa S3 cells by
the DEAE-dextran method (24–26).
After a further 24 h of incubation, cells were collected and lysed
with 100 μl of 1X cell culture lysis reagent, mixed and centrifuged
at 12,000 × g for 5 sec. The supernatant was stored at −80°C. The
Dual Luciferase assay was performed with the Dual Luciferase assay
system (Promega), as previously described (21).
Results
Decrease in the amounts of PARG gene
transcripts after introducing its siRNAs
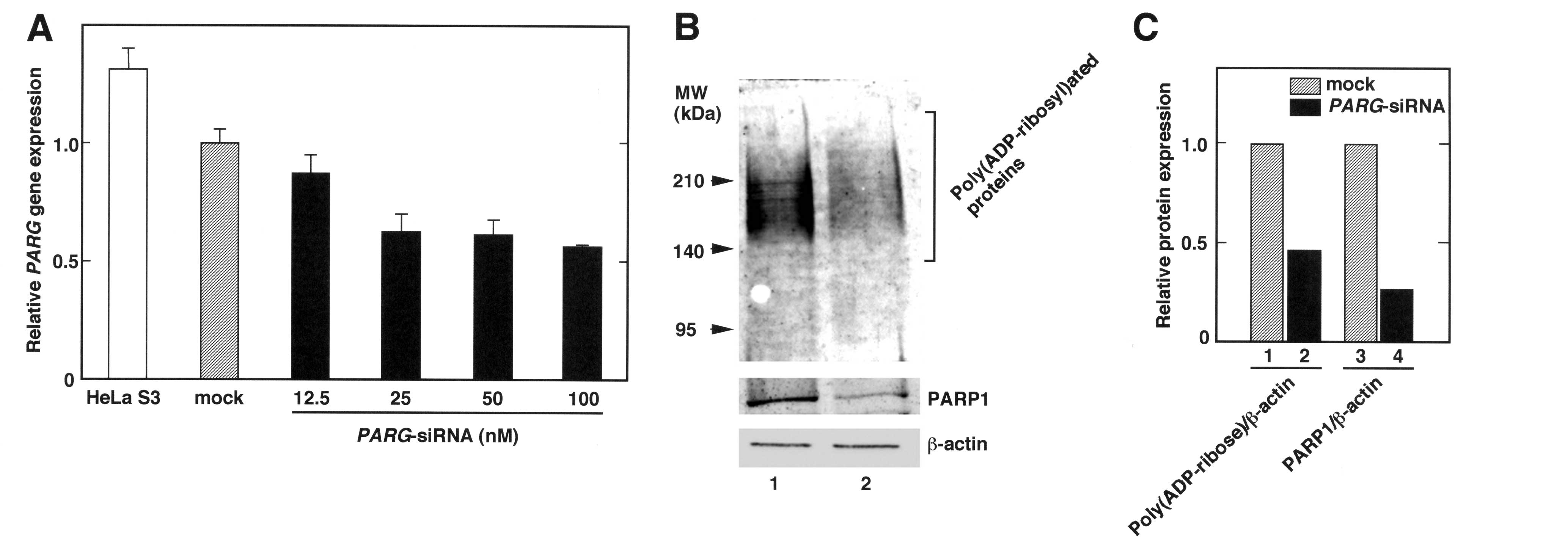
In order to suppress PARG gene expression,
ON-TARGETplus SMARTpool siRNAs (PARG-siRNA) were transfected
into HeLa S3 cells with DharmaFECT 1 reagent. The relative amounts
of PARG transcripts were reduced by PARG-siRNA treatment
(Fig. 1A). In 100 nM of
PARG-siRNA-treated cells, the PARG gene expression level
decreased to approximately half of that in the mock-transfected
cells. Since this treatment did not affect viability of cells
compared with the mock-transfected cells (data not shown), further
experiments were performed with 100 nM of PARG-siRNA.
Treatment of PARG-siRNA reduces the
amounts of poly(ADP-ribose) and PARP1
As the PARG gene encodes the main
poly(ADP-ribose) degrading enzyme, the level of
poly(ADP-ribosyl)ated proteins was assumed to increase following
the introduction of PARG-siRNA into HeLa S3 cells. Western
blotting revealed that the amounts of poly(ADP-ribose) decreased in
the PARG-siRNA-transfected HeLa S3 cells (Fig. 1B). In addition, the decrease of
poly(ADP-ribose) in the whole-cell extracts was accompanied by a
decrease in PARP1 protein levels (Fig.
1C), suggesting that the PARG-siRNA diminished
poly(ADP-ribose) and the PARP1 protein.
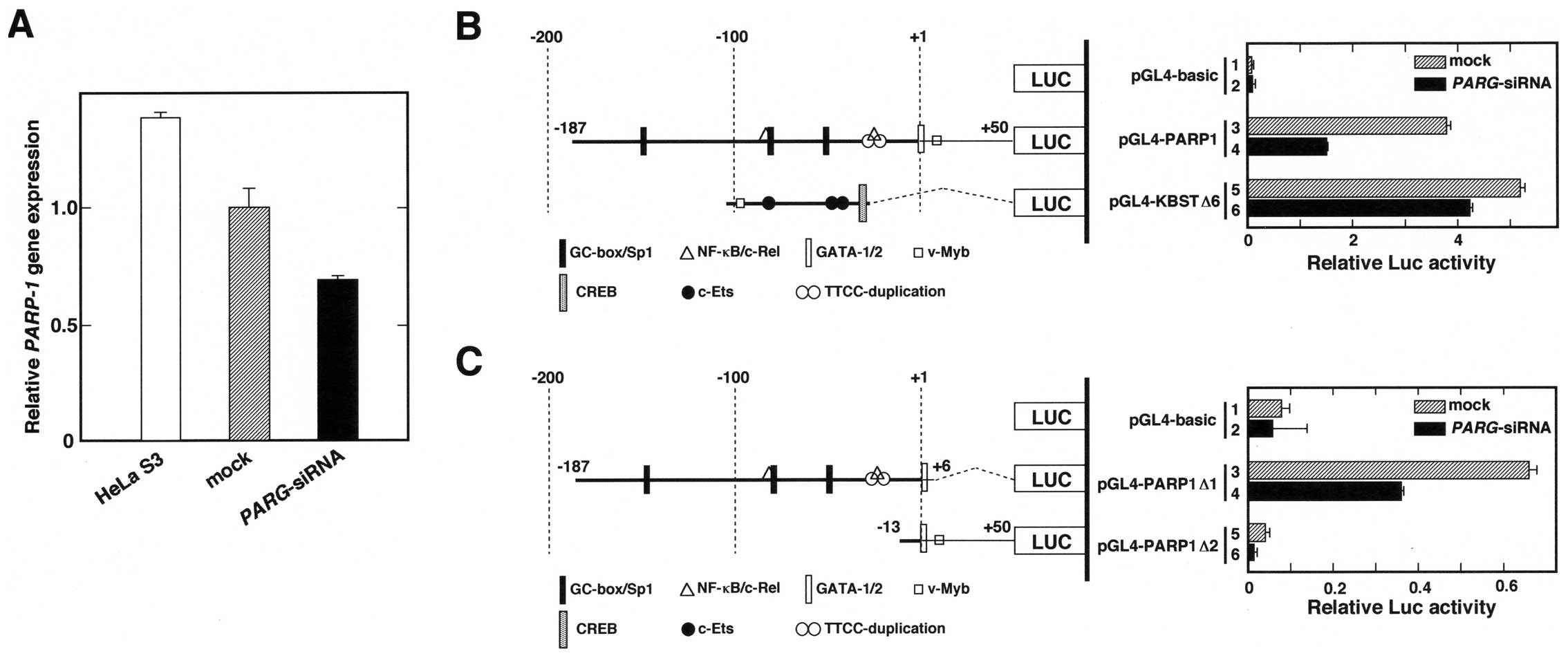
PARP1 gene expression and its promoter
activity are downregulated by transfection with PARG-siRNA
To examine whether the decrease in PARP1 protein
level is caused by changes in gene expression and promoter
activity, RT-qPCR analyses and transient transfection experiments
were performed. As shown in Fig.
2A, the relative PARP1 gene expression of
PARG-siRNA-transfected cells was 60% that of the control
(mock) cells. In addition, the transient transfection and Luc
reporter assay indicated that the PARP1 promoter activity
decreased in the PARG-siRNA-transfected cells compared to
the control cells (Fig. 2B). We
also tested whether the 75-bp core promoter of the PARG gene
(21) responds to PARG-siRNA
treatment (Fig. 2B, bars 5 and 6),
but only a 10% decrease was observed. To limit the
PARG-siRNA responsive element(s), we constructed two
deletion plasmids that contained −187 to +6 and −13 to +50 bp of
the human PARP1 promoter (Fig. 2C).
The results showed that the 193-bp fragment, which contains the
GC-box/Sp1, NF-κB/c-Rel, GATA-1 and GATA-2 binding sequences,
responded to the PARG-siRNA. The promoter activity obtained
from pGL4-PARP1Δ2-transfected cells was lower than that of the
pGL4-basic vector-transfected cells, indicating that the 63-bp
sequence from −13 to +50 is not essential for transcription of the
PARP1 gene. These results suggest that the transfection of
PARG-siRNA leads to suppression of PARP1 gene
expression through the 193-bp sequence of the PARP1 promoter
region.
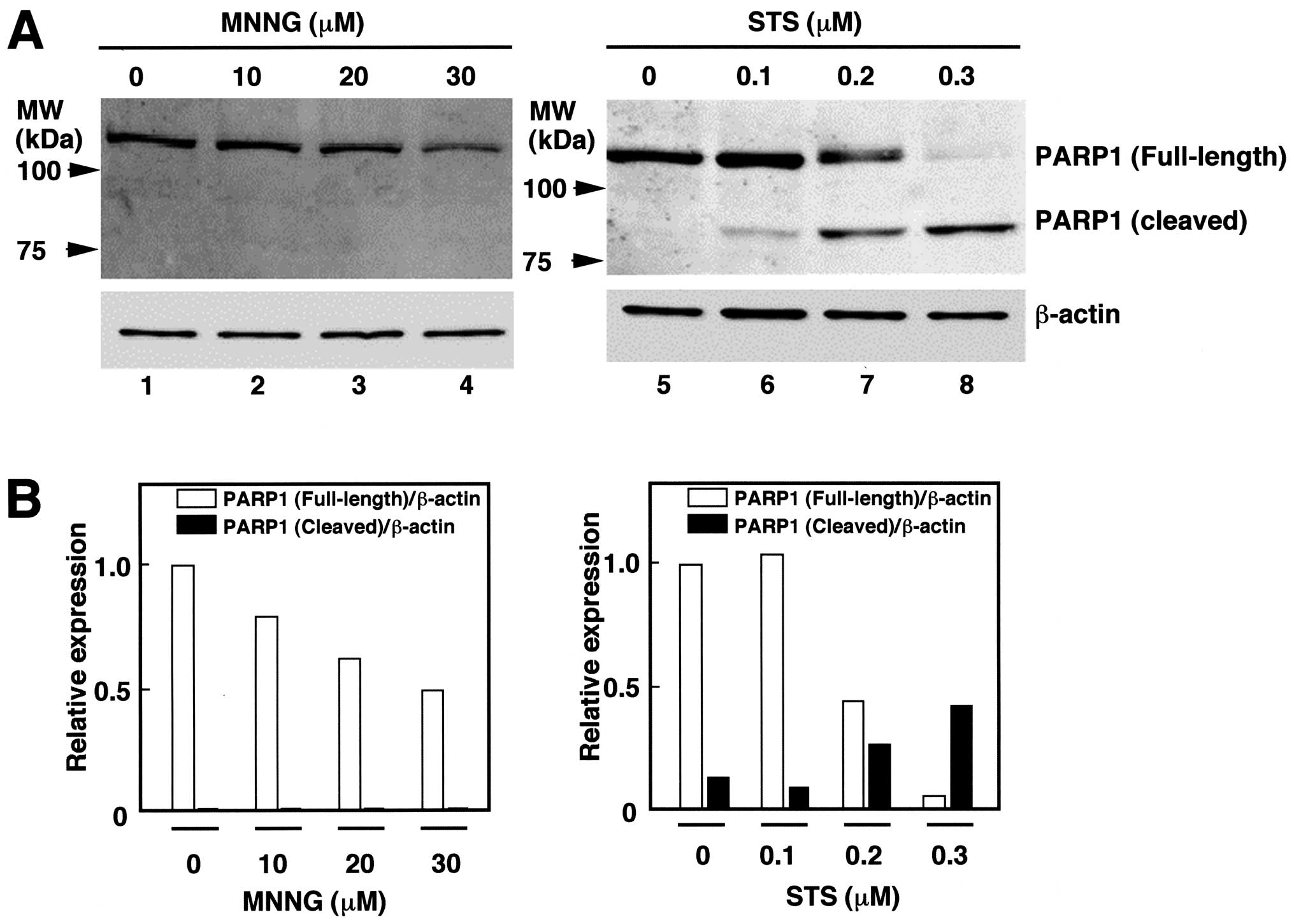
Viability of PARG siRNA-introduced HeLa
S3 cells following N-methyl-N′ nitro-N-nitrosoguanidine (MNNG) and
STS treatments
It has been demonstrated that poly(ADP-ribose)
metabolism is associated with cell death or apoptosis (13–15).
To examine the effects of MNNG and STS on PARP1 in HeLa S3 cells,
western blot analysis was performed (Fig. 3). The treatment with STS (0.1–0.3
μM) induced cleavage of PARP1, suggesting that caspases were
activated and cells underwent apoptosis. Although the relative
amount of PARP1 was diminished to 50%, cleaved forms of PARP1 were
not detected by the treatment with 30 μM of MNNG (Fig. 3).
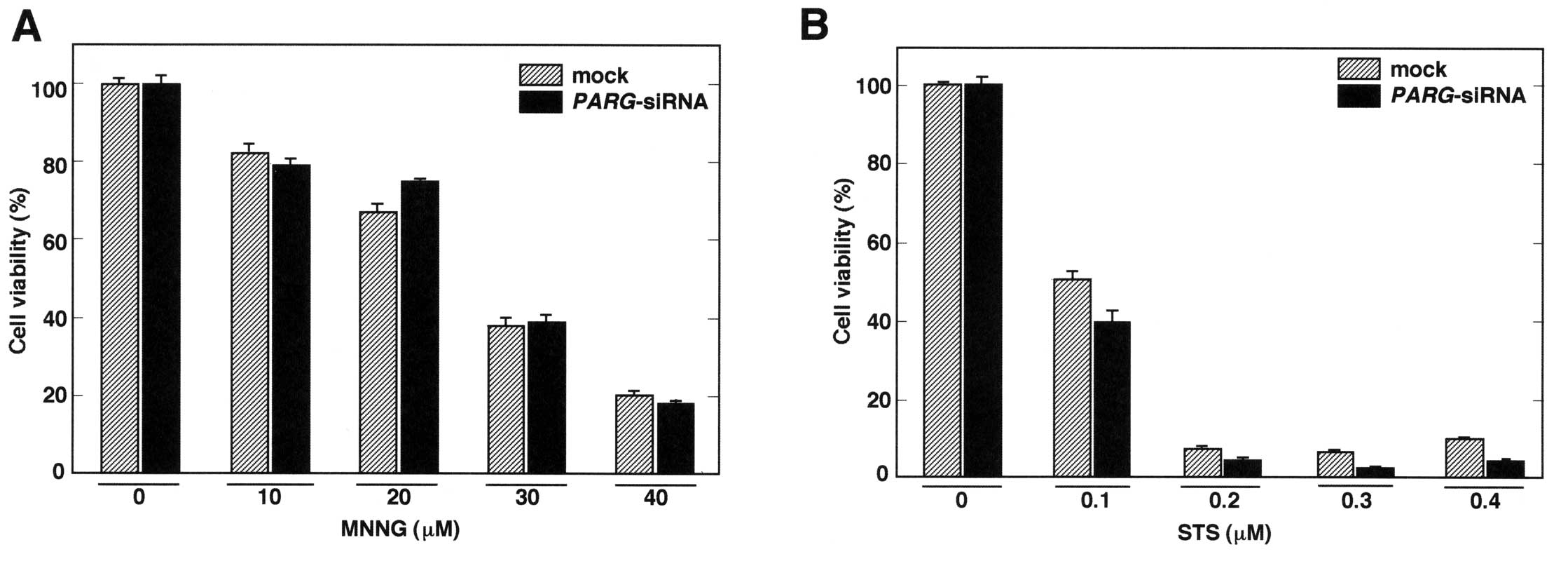
Subsequently, the effect of MNNG and STS on cell
viability was analyzed following the transfection of
PARG-siRNA. Although 40 μM of MNNG treatment caused severe
damage to the cells, transfection of PARG-siRNA did not
further increase cell death (Fig.
4A). On the other hand, PARG-siRNA strongly impaired
cell viability when cells were treated with 0.3 and 0.4 μM of STS
(Fig. 4B). These results suggest
that PARG-siRNA treatment enhances cell death in conditions
that cause PARP1 cleavage.
Discussion
Several experiments concerning expression of the
PARG gene that have utilized PARG-siRNA or
PARG-shRNA overexpressing systems have been reported
(27). Constitutive suppression of
PARG gene expression by introducing shRNA into HeLa S3 cells
caused accumulation of poly(ADP-ribose) and led to enhancement of
mitotic catastrophe by X-ray irradiation (28). It was reported that poly(ADP-ribose)
accumulation occurred after a PARG shRNA expression vector
was stably introduced into A549 cells, a lung adenocarcinoma cell
line (29). Of note, these
PARG-suppressed A549 cells exhibited reduced PARP activity. The
decrease in poly(ADP-ribose) was reported in mouse 3T3 embryonic
fibroblasts, PARG-Δ2,3 cells, which express PARG60 but not
full-length PARG110 (30). In the
present study, a PARG-siRNA pool that contains sequences
located upstream of exon 5 was transfected into HeLa S3 cells.
Therefore, PARG-siRNA used here may selectively reduce
full-length PARG expression but may not affect PARG60. In addition,
the suppressing effect by the introduction of 25–100 nM of the
PARG-siRNA was approximately 50% of the control (Fig. 1A). This low efficiency might be
caused by the reduced poly(ADP-ribose) level in the cells (Fig. 1B).
It has been suggested that poly(ADP-ribosyl)ation of
Sp1 impairs activation of the PARP1 promoter (16). It has also been reported that PARP1
binds to its own promoter to suppress transcription (31). Since PARP1 physically interacts with
Sp1 (16), both may cooperatively
regulate transcription of the PARP1 gene. In our study, the
introduction of PARG-siRNA into HeLa S3 cells caused
suppression of the PARP1 promoter activity and led to
subsequent reduction of its gene/protein expression. These data
suggest a possible mechanism that prevents cells from accumulating
excess poly(ADP-ribose) by suppressing PARP1 promoter
activity. It is noteworthy that inverted repeats or putative
cruciform-like structures are found both in the
PARP1(31) and
PARG(21) promoters. Since
PARP1 has been suggested to recognize and bind to the
cruciform-like structures of DNA (31), the PARG promoter is possibly
one of the targets for PARP1. Moreover, the PARP1 promoter
harbors overlapped TTCC motifs as 5′-GCGGGTT
CCGTGGGCGTTCCCGCGG-3′ (Fig.
2B and C). The duplicated GGAA (TTCC) motifs, which are also
found in the human PARG promoter region (21), have been suggested to be one of the
determinants of TSS in TATA-less promoters with responses to
various stimuli, including cytokine and differentiation inducing
signals (32–34). In addition, previous reports
suggested that PARP1 and PARG interact with each other (14) and coordinately regulate global
patterns of gene expression, and that the binding of PARP1 to the
promoters in the genome is dependent on the presence of PARG
(27). Collectively, these
observations indicate that the location of PARP1 with PARG on these
promoters may play an important role in the regulation of
poly(ADP-ribose) metabolism at the transcriptional level. This
might be relevant to the cooperative function of PARP1 and PARG to
repair single-strand break of DNA (35).
In this study, we showed that the introduction of
PARG-siRNA enhances sensitivity to STS but not to MNNG
(Fig. 4). Since PARP1 cleavage was
detected in STS-treated cells (Fig.
3), the introduction of PARG-siRNA may cause elevation
of cell damage by reducing PARP1 expression and lead to complete
loss of native PARP1, and induce cell death by apoptosis. On the
other hand, cell death induced by MNNG was not further affected by
the PARG-siRNA, suggesting that the signals induced by the
reduction of PARP1 have been adequately saturated by the MNNG
treatment. However, this possibility remains to be elucidated in
future analyses.
In conclusion, the data presented in this study
provide a new theory that PARP1 gene expression is regulated
by the amount of PARG transcripts, suggesting a mechanism to
avoid accumulation of excess amounts of poly(ADP-ribose) in a cell
by regulating their transcription. Since poly(ADP-ribose)
metabolism also controls NAD+ levels, local changes in
the concentrations of NAD+ might affect transcription of
the PARP1 and PARG genes. Markedly, in accordance
with the development of PARG inhibitors (36), the use of the PARG-siRNA may
contribute to a treatment of cancer with a reduced dose of
anti-cancer drugs or ionizing radiation.
Acknowledgements
The authors thank T. Sato and A. Asanuma for their
technical assistance. This study was supported in part by a
Research Fellowship from the Research Center for RNA Science, RIST,
Tokyo University of Science.
References
|
1
|
Schreiber V, Dantzer F, Ame JC and de
Murcia G: Poly(ADP-ribose): novel functions for an old molecule.
Nat Rev Mol Cell Biol. 7:517–528. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tanuma S and Kanai Y:
Poly(ADP-ribosyl)ation of chromosomal proteins in the HeLa S3 cell
cycle. J Biol Chem. 257:6565–6570. 1982.PubMed/NCBI
|
|
3
|
Lönn U and Lönn S: Accumulation of
10-kilobase DNA replication intermediates in cells treated with
3-aminobenzamide. Proc Natl Acad Sci USA. 82:104–108.
1985.PubMed/NCBI
|
|
4
|
Durkacz BW, Omidiji O, Gray DA and Shall
S: (ADP-ribose)n participates in DNA excision repair. Nature.
283:593–596. 1980. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kreimeyer A, Wielckens K, Adamietz P and
Hilz H: DNA repair-associated ADP-ribosylation in vivo.
Modification of histone H1 differs from that of the principal
acceptor proteins. J Biol Chem. 259:890–896. 1984.PubMed/NCBI
|
|
6
|
Tanuma S, Yagi T and Johnson GS:
Endogenous ADP ribosylation of high mobility group proteins 1 and 2
and histone H1 following DNA damage in intact cells. Arch Biochem
Biophys. 237:38–42. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chang P, Jacobson MK and Mitchison TJ:
Poly(ADP-ribose) is required for spindle assembly and structure.
Nature. 432:645–649. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kraus WL and Lis JT: PARP goes
transcription. Cell. 113:677–683. 2003. View Article : Google Scholar
|
|
9
|
d’Adda di Fagagna F, Hande MP, Tong WM,
Lansdorp PM, Wang ZQ and Jackson SP: Functions of poly(ADP-ribose)
polymerase in controlling telomere length and chromosomal
stability. Nat Genet. 23:76–80. 1999.PubMed/NCBI
|
|
10
|
Zampieri M, Passananti C, Calabrese R,
Perilli M, Corbi N, De Cave F, Guastafierro T, Bacalini MG, Reale
A, Amicosante G, Calabrese L, Zlatanova J and Caiafa P: Parp1
localizes within the Dnmt1 promoter and protects its unmethylated
state by its enzymatic activity. PloS One. 4:e47172009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Simbulan-Rosenthal CM, Rosenthal DS, Hilz
H, Hickey R, Malkas L, Applegren N, Wu Y, Bers G and Smulson ME:
The expression of poly(ADP-ribose) polymerase during
differentiation-linked DNA replication reveals that it is a
component of the multiprotein DNA replication complex.
Biochemistry. 35:11622–11633. 1996. View Article : Google Scholar
|
|
12
|
Uchiumi F, Ikeda D and Tanuma S: Changes
in the activities and gene expressions of poly(ADP-ribose)
glycohydrolases during the differentiation of human promyelocytic
leukemia cell line HL-60. Biochim Biophys Acta. 1676:1–11. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Andrabi SA, Kim NS, Yu SW, Wang H, Koh DW,
Sasaki M, Klaus JA, Otsuka T, Zhang Z, Koehler RC, Hurn PD, Poirier
GG, Dawson VL and Dawson TM: Poly(ADP-ribose) (PAR) polymer is a
death signal. Proc Natl Acad Sci USA. 103:18308–18313. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Keil C, Gröbe T and Oei SL: MNNG-induced
cell death is controlled by interactions between PARP-1,
poly(ADP-ribose) glycohydrolase, and XRCC1. J Biol Chem.
281:34394–34405. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
García S, Gómez-Reino JJ and Conde C:
Poly(ADP-ribose) polymerase suppression protects rheumatoid
synovial fibroblast from Fas-induced apoptosis. Rheumatology
(Oxford). 48:483–489. 2009.PubMed/NCBI
|
|
16
|
Zaniolo K, Desnoyers S, Leclerc S and
Guérin SL: Regulation of poly(ADP-ribose) polymerase-1 (PARP-1)
gene expression through the post-translational modification of Sp1:
a nuclear target protein of PARP-1. BMC Mol Biol. 8:962007.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Oei SL, Griesenbeck J, Schweiger M, Babich
V, Kropotov A and Tomilin N: Interaction of the transcription
factor YY1 with human poly(ADP-ribosyl) transferase. Biophem
Biophys Res Commun. 240:109–111. 1997.PubMed/NCBI
|
|
18
|
Hassa PO and Hottiger MO: A role of poly
(ADP-ribose) polymerase in NF-kappaB transcriptional activation.
Biol Chem. 380:953–959. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yokoyama Y, Kawamoto T, Mitsuuchi Y,
Kurosaki T, Toda K, Ushiro H, Terashima M, Sumimoto H, Kuribayashi
I, Yamamoto Y, Maeda T, Ikeda H, Sagara Y and Shizuta Y: Human
poly(ADP-ribose) polymerase gene. Cloning of the promoter region.
Eur J Biochem. 194:521–526. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Meyer RG, Meyer-Ficca ML, Jacobson EL and
Jacobson MK: Human poly(ADP-ribose) glycohydrolase (PARG) gene and
the common promoter sequence it shares with inner mitochondrial
membrane translocase 23 (TIM23). Gene. 314:181–190. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Uchiumi F, Sakakibara G, Sato J and Tanuma
S: Characterization of the promoter region of the human
PARGgene and its response to PU.1 during differentiation of
HL-60 cells. Genes Cells. 13:1229–1248. 2008.PubMed/NCBI
|
|
22
|
Satoh MS and Lindahl T: Role of
poly(ADP-ribose) formation in DNA repair. Nature. 356:356–358.
1992. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Benjamin RC and Gill DM: ADP-ribosylation
in mammalian cell ghosts. Dependence of poly(ADP-ribose) synthesis
on strand breakage in DNA. J Biol Chem. 255:10493–10501.
1980.PubMed/NCBI
|
|
24
|
Zhou B, Ikejima T, Watanabe T, Iwakoshi K,
Idei Y, Tanuma S and Uchiumi F: The effect of 2-deoxy-D-glucose on
Werner syndrome RecQ helicase gene. FEBS Lett. 583:1331–1336. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Uchiumi F, Enokida K, Shiraishi T, Masumi
A and Tanuma S: Characterization of the promoter region of the
human IGHMBP2(Sμbp-2) gene and its response to TPA in HL-60
cells. Gene. 463:8–17. 2010.PubMed/NCBI
|
|
26
|
Uchiumi F, Watanabe T and Tanuma S:
Characterization of various promoter regions of the human DNA
helicase-encoding genes and identification of duplicated
ets(GGAA) motifs as an essential transcription regulatory
element. Exp Cell Res. 316:1523–1534. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Frizzell KM, Gamble MJ, Berrocal JG, Zhang
T, Krishnakumar R, Cen Y, Sauve AA and Kraus WL: Global analysis of
transcriptional regulation by poly(ADP-ribose) polymerase-1 and
poly(ADP-ribose) glycohydrolase in MCF-7 human breast cancer cells.
J Biol Chem. 284:33926–33938. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Amé JC, Fouquerel E, Gauthier LR, Biard D,
Boussin FD, Dantzer F, de Murcia G and Schreiber V:
Radiation-induced mitotic catastrophe in PARG-deficient cells. J
Cell Sci. 122:1990–2002. 2009.PubMed/NCBI
|
|
29
|
Erdélyi K, Bai P, Kovács I, Szabó E,
Mocsár G, Kakuk A, Szabó C, Gergely P and Virág L: Dual role of
poly(ADP-ribose) glycohydrolase in the regulation of cell death in
oxidatively stressed A549 cells. FASEB J. 23:3553–3563.
2009.PubMed/NCBI
|
|
30
|
Gao H, Coyle DL, Meyer-Ficca M, Meyer RG,
Jacobson EL, Wang ZQ and Jacobson MK: Altered poly(ADP-ribose)
metabolism impairs cellular responses to genotoxic stress in a
hypomorphic mutant poly(ADP-ribose) glycohydrolase. Exp Cell Res.
313:984–996. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Soldatenkov VA, Chasovskikh S, Potaman VN,
Trofimova I, Smulson ME and Dritschilo A: Transcriptional
repression by binding of poly(ADP-ribose) polymerase to promoter
sequences. J Biol Chem. 277:665–670. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Uchiumi F, Miyazaki S and Tanuma S: The
possible functions of duplicated ets(GGAA) motifs located
near transcription start sites of various human genes. Cell Mol
Life Sci. 68:2039–2051. 2011.PubMed/NCBI
|
|
33
|
Uchiumi F, Larsen S and Tanuma S:
Biological systems that control transcription of DNA repair-and
telomere maintenance-associated genes. DNA Repair-New Research
Directions. ISBN: 980-953-307-746-3Chen C: InTech-Open Access
Publisher, Inc; Rijeka, Croatia: (In press).
|
|
34
|
Uchiumi F, Larsen S, Masumi A and Tanuma
S: The implications of duplicated GGAA-motifs located in the human
interferon regulated genes (ISGs). Introduction to Sequence and
Genome Analysis. ISBN: 980-14775549-1-3(iConcept ed). iConcept
Press Ltd; Hong Kong: (In press).
|
|
35
|
Fisher AE, Hochegger H, Takeda S and
Caldecott KW: Poly(ADP-ribose) polymerase 1 accelerates
single-strand break repair in concert with poly(ADP-ribose)
glycohydrolase. Mol Cell Biol. 27:5597–5605. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Okita N, Ashizawa D, Ohta R, Abe H and
Tanuma S: Discovery of novel poly(ADP-ribose) glycohydrolase
inhibitors by a quantitative assay system using dot-blot with
anti-poly(ADP-ribose). Biochem Biophys Res Commun. 392:485–489.
2010. View Article : Google Scholar : PubMed/NCBI
|