Introduction
Primary malignant gliomas are diagnosed in 6–7 out
of 100,000 people each year. Despite advances in treatment
strategies that combine surgery with radiotherapy and chemotherapy,
these tumors remain one of the most fatal diseases (1). The genesis mechanisms of gliomas
remain unknown.
It was hypothesized in previous studies that the
development and prevalent resistance of brain tumor is attributed
to brain tumor stem cells (BTSCs), which are a small population of
tumor cells within the heterogeneous tumor mass with stem-like
characteristics (2). According to
this hypothesis, BTSCs sustain the long-term growth of brain tumor
and are responsible for tumor recurrence following conventional
treatments. Current treatments combining surgery and
chemoradiotherapy are not able to eliminate BTSCs (3–5);
therefore, new treatments are required to more efficiently target
BTSCs. It is necessary to thoroughly understand the molecular and
cellular mechanisms underlying the self-renewal and differentiation
of BTSCs.
OCT4, also known as OCT3 or POU5F1, is a member of
the POU (Pit, Oct, Unc) transcription factor family encoded by the
POU5F1 gene (6p2113) and was first identified in 1990
(6). The expression of OCT4 is
restricted to pluripotent stem cells and is downregulated when
differentiation is initiated during embryonic development. OCT4 is
regarded as a gatekeeper at early mammalian development (7) and regulates the self-renewal and
pluripotency of human embryonic stem cells (ESCs) (8,9). OCT4
plays an important role in maintaining cellular plasticity and
promoting the self-renewal and proliferation ability of stem cells
(7). In addition, OCT4-positive
cells identified in cancer may represent cancer stem cells (CSCs)
and account for the maintenance and propagation of tumors (10,11).
In primary glioma, OCT4 is highly expressed and increases in
parallel with glioma grading, indicating that the reactivation of
OCT4 or other stem cell genes may help to maintain the status of
BTSCs and enhance the malignancy of high-grade gliomas (12–14).
These findings provide evidence of OCT4 for the stem cell theory on
tumorigenesis. However, no study has reported the potential
mechanism of how OCT4 expression is upregulated with glioma grading
in primary gliomas.
Epigenetic regulation, which refers to the
alteration in the expression of heritable genes due to the changes
in non-DNA sequences, is one of the crucial mechanisms in the
control of gene transcription (15). DNA methylation is a mechanism
extensively investigated in this field. The methylation of the CpG
island at the promoter usually leads to the downregulation of gene
expression. Higher level methylation predicts lower transcription
activity (16,17). The aberrant DNA methylation pattern
can alter gene transcription that is etiologically linked to the
formation of cancer (18–20). It was believed that the epigenetic
regulation of the chromatin state plays an important role in the
control of OCT4 expression (21).
DNA methylation of OCT4 gene regulatory regions and histone
modifications contribute to the silencing of the OCT4 gene during
the differentiation and embryo development of ESCs in mouse and
human (22–24). In addition, OCT4 expression to the
germ line cells and the early embryo cells may also be mediated by
DNA methylation and the mouse ortholog OCT4 is more strongly
methylated in differentiated cells than in ESCs (25). OCT4 gene methylation has been
studied extensively, but little is known about its DNA methylation
at the promoter and exon in primary glioma and its function in
tumorigenesis.
Therefore, we hypothesized that DNA methylation may
be involved in the regulation of OCT4 expression during the
occurrence and development of primary glioma. In this study, we
evaluated the expression and the methylation status of OCT4 in 24
patients with primary gliomas and further analyzed their
relationship. In vitro, we maintained and treated glioma
cell lines with a demethylation reagent to further detect the OCT4
expression.
Materials and methods
Patient selection and sampling
All the investigations described in this study were
conducted after informed consent was obtained and in accordance
with an institutional review board protocol approved by the ethics
committee at the Affiliated Hospital of Nantong University. A total
of 24 patients with primary glioma were recruited from the
Department of Neurosurgery, Affiliated Hospital of Nantong
University, from January 2009 to December 2011. Pathological
findings were determined by more than 2 pathologists and classified
according to the WHO classification standard. There were 12 low-
(WHO Grade II) and 12 high-grade tumors (WHO Grade III and IV). The
glioma tissues were collected and fixed in 10% formaldehyde
followed by embedding in paraffin. In addition, a fraction of
samples was placed into liquid nitrogen (−70°C) for use.
Glioma cells and demethylation
treatment
Two human glioma cell lines, U87MG and U251MG, were
purchased from the Shanghai Cell Institute of the Chinese Academy
of Sciences. Both cell lines were maintained in Dulbecco’s Modified
Eagle’s Medium; Nutrient Mixture F-12 (DMEM/F12) supplemented with
10% heat-inactivated fetal bovine serum (FBS), 100 U/ml penicillin
and 100 μg/ml streptomycin (all from Gibco). Cells were incubated
at 37°C in a humidified atmosphere of 5% CO2.
Demethylation with 5-Aza-2′-deoxycytidine; when cell cultures
reached 50% confluence, they were treated with
5-Aza-2′-deoxycytidine (A3656; Sigma-Aldrich, St. Louis, MO, USA)
at the final concentration of 10 nM, respectively, for 3 days.
RNA isolation and real-time PCR
RNA expression levels of OCT4 were determined using
quantitative real-time PCR with GAPDH as positive controls. Total
mRNA was isolated from glioma specimens and cell lines using mRNA
Isolation Kit (Roche, UK) following the manufacturer’s
instructions. The concentration and purity of mRNA was determined
by ultraviolet spectrophotometry. Isolated mRNA (100 ng) from each
sample was transcribed to complementary DNA (cDNA) using a
First-Strand cDNA Synthesis Kit (Roche), which was then used as a
template for quantitative real-time PCR.
Primers used for OCT4 were:
5′-TATTCAGCCAAACGACCATCT-3′ (sense) and 5′-TCAGCTTCCTCCACCCACTT-3′
(antisense); for GAPDH, primers were: 5′-GGAAAGCTGTGGCGTGAT-3′
(sense) and 5′-AAGGTGGAAGAATGGGAGTT-3′ (antisense). The primers
were designed using Primer 5.0 software and manufactured by TIB
Molbiol. A 20 μl reaction, which included 2 μl of cDNA template, 2
μl of forward and reverse primer, 6 μl DEPC H2O and 10
μl SYBR-Green Mix (QPK-201, Toyobo), was conducted using the ABI
Prism 7500 Sequence Detection System (Applied Biosystems). PCRs of
each template were performed in duplicate in a 96-well plate. The
thermal cycling conditions included an initial denaturation step at
95°C for 5 min and 40 cycles at 95°C for 10 sec, at 59°C for 15 sec
and at 72°C for 20 sec. The relative fold-change 2−ΔΔCT
method was used to determine the relative quantitative gene
expression compared with GAPDH. The transcription level of target
genes observed in calibrating samples was treated as the basal
level and given the value 1.0. All PCR reactions were performed in
triplicate and a negative control was included that contained
primers without cDNA.
Western blot analysis
The samples were then homogenized in lysis buffer
(1% NP-40, 50 mmol/l Tris, pH 7.5, 5 mmol/l EDTA, 1% SDS, 1% sodium
deoxycholate, 1% Triton X-100, 1 mmol/l PMSF, 10 mg/ml aprotinin
and 1 mg/ml leupeptin; Sigma-Aldrich) and clarified by centrifuging
for 20 min in a microcentrifuge at 4°C. Following determination of
its protein concentration with the Bradford Assay (Bio-Rad, USA),
the resulting supernatant (50 μg of protein) was subjected to
SDS-polyacrylamide gel electrophoresis (PAGE). The separated
proteins were transferred to a polyvinylidine difluoride membrane
(Millipore Corp., USA) by a transfer apparatus at 350 mA for 2.5 h.
The membranes were first blocked and then incubated with the
primary antibody described above for 2 h at room temperature. After
washing three times, filters were incubated with horseradish
peroxidase-conjugated human anti-mouse or anti-rabbit antibodies
(Pierce) for 1 h at room temperature. Immunocomplexes were detected
with an enhanced chemiluminescence system (NEN Life Science
Products, USA). The western blotting experiments were repeated at
least three times.
Immunohistochemistry (IHC)
Each glioma was immunohistochemically examined for
OCT4 nuclear staining. Specimens were cut serially into 4-μm
sections. Sections were dewaxed in xylene and rehydrated in graded
ethanols. Endogenous peroxidase activity was blocked by immersion
in 0.3% methanolic peroxide for 30 min. Immunoreactivity was
enhanced by microwaving and incubating the tissue sections for 10
min in 0.1 mol/l citrate buffer. Primary rabbit polyclonal
antibodies OCT4 (1:200; Abcam, USA) were applied and incubated
overnight at 4°C. Secondary Goat Anti-Rabbit immunoglobulin
(Invitrogen Life Technologies, USA) was applied and incubated for 1
h at room temperature. Some sections were processed with
Tris-buffered saline rather than primary antibody and were used as
negative controls. Immunocomplexes were visualized by brown
pigmentation via a standard 3,3-diaminobenzidine (DAB) protocol.
Appropriate positive and negative controls were used. Ten
high-power fields were randomly chosen and at least 300 cells were
counted per field. Tumors were scored as a percentage of positive
cells. The staining procedures were repeated at least three
times.
DNA preparation and bisulfite genomic
sequencing
Genomic DNA was extracted from frozen tissues by
proteinase K digestion and the phenol-chloroform method (26). Sodium bisulfite treatment of the
extracted DNA was performed as previously described with some
modifications (27). In brief, 10
μg DNA in 50 μl TE was incubated with 5.5 μl of 0.3 M NaOH at 37°C
for 15 min and at 95°C for 2 min and then subjected to sodium
bisulfite chemical treatment (2.4 M sodium metabisulfite; 0.5 mM
hydroquinone, pH 5.0; both from Sigma-Aldrich). Following
incubation for 4 h at 55°C, the treated DNA was purified using the
SK1261 kit (Shenggong Biotechnology Co., China), desulfonated in
0.3 M NaOH and then neutralized to pH 7.0 using 3 M sodium acetate
(pH 5.2). The neutralized DNA was then purified using the SK1261
purification kit again, dissolved in TE buffer (pH 8.0).
The methylation status of the minimal promoter and
part of exon 1 region CpG islands of OCT4 was analyzed (23,28).
The primers were designed to amplify the promoter and exon 1 from
−234 to +46 for bisulfite genomic sequencing. The forward primer
was 5′-GGATTTGTATTGAGGTTTTGGAG-3′ and the reverse,
5′-TAACCCATCACCTCCACCAC-3′. Touchdown PCR was then carried out. An
initial denaturation at 98°C for 4 min was followed by five PCR
cycles of 94°C for 45 sec, 68°C for 45 sec and 72°C for 1 min. The
PCR was then completed with 35 cycles of 45 sec at 95°C, 45 sec at
58°C. The amplified products were gel-purified using the SK1261 kit
and then subjected to TA-cloning using pUC18-T vector (Shenggong
Biotechnology Co.). Ten clones for each case were selected for
sequencing using BigDye version 3.1 and analyzed on automated DNA
sequence analyzer (ABI Prism 3730; Applied Biosystems, Inc., Foster
City, CA, USA). The cytosine or thymine residues at the CpG sites
represented methylated or unmethylated status, respectively.
Statistical analysis
Statistical analysis was performed using SPSS 13.0
for Windows. The Student’s independent-sample t-test statistics was
used for statistical comparison. The correlation between
methylation status and mRNA expression level of OCT4 was tested by
Pearson’s correlation test. All statistical tests were calculated
in two-sided and a P-value <0.05 was considered to indicate a
statistically significant difference.
Results
OCT4 is highly expressed in primary
gliomas
The primary glioma samples were collected from 24
patients and placed in liquid nitrogen, followed by detection of
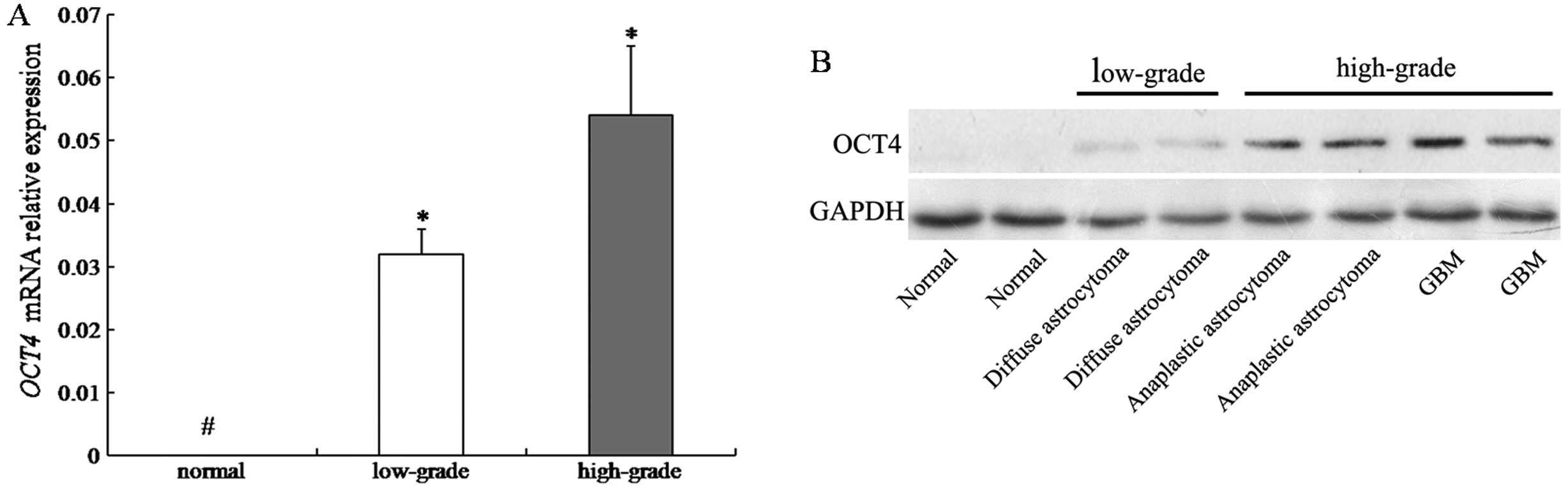
OCT4 mRNA expression by real-time PCR. Results revealed that
expression of OCT4 mRNA was upregulated in all 24 human glioma
samples, but no obvious expression was observed in the normal brain
samples. Densitometric evaluation of relative expression showed
that the level of OCT4 mRNA was significantly higher in
high-grade gliomas than in low-grade gliomas (P<0.05) (Fig. 1A). Western blot analysis confirmed
that the level of OCT4 protein was higher in primary high-grade
gliomas than in low-grade ones (Fig.
1B).
OCT4 protein is localized in the nuclei
of primary glioma cells
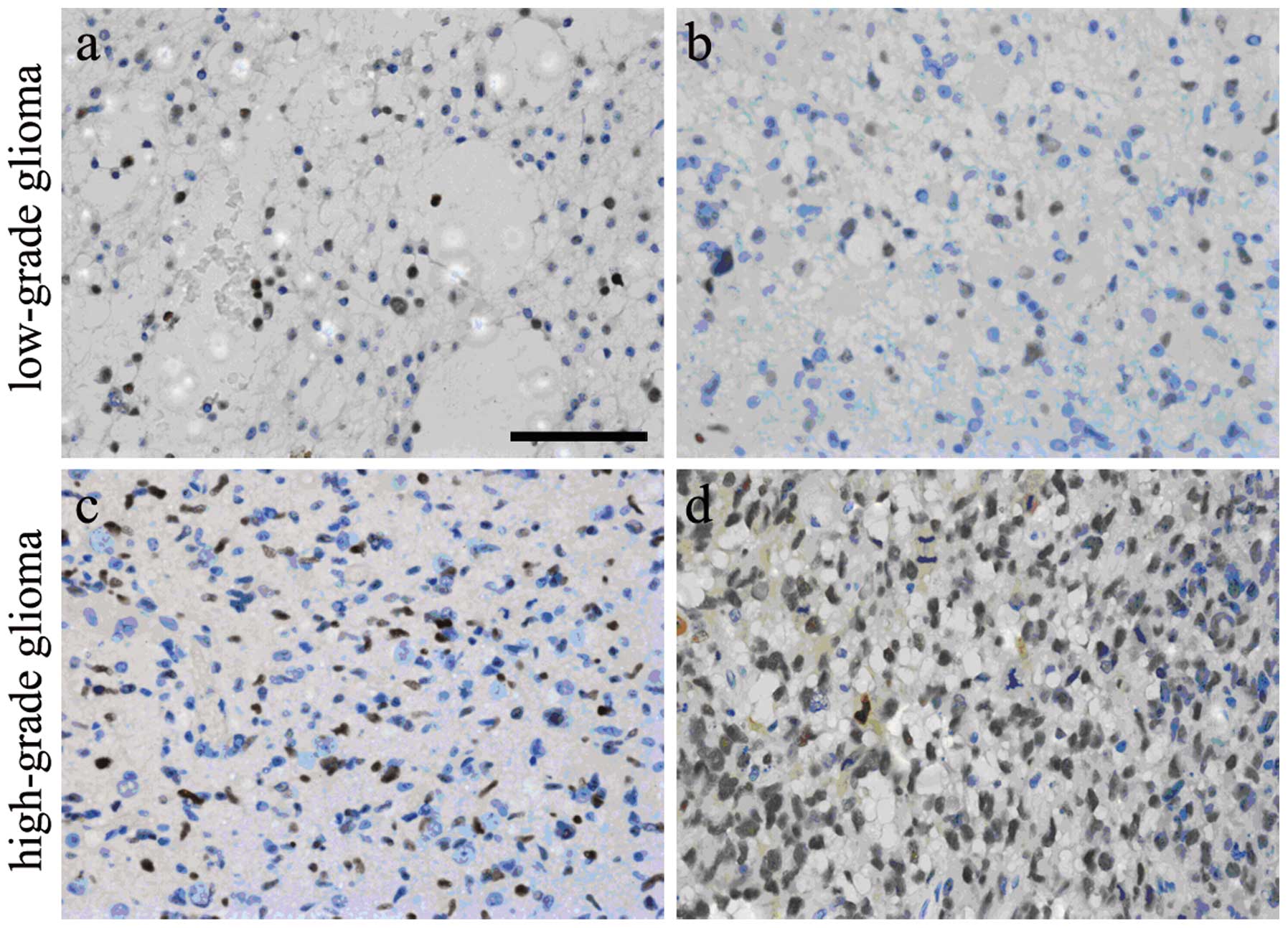
The primary glioma tissues were collected from 24
patients and paraffin-embedded sections were obtained for IHC,
followed by counting of OCT4-positive cells. OCT4 was expressed in
the nuclei of tumor cells and OCT4-positive cells were identified
in all glioma samples. The percentage of OCT4-positive cells ranged
from 3.79 to 79.67%. The nuclei of most high-grade glioma cells
(Fig. 2C and D) were more intensely
stained than those of low-grade tumors (Fig. 2A and B), indicating that OCT4
expression in the nuclei of glioma cells was in a grade-dependent
manner.
Hypomethylation of the OCT4 gene and
negative correlation with OCT4 expression in primary glioma
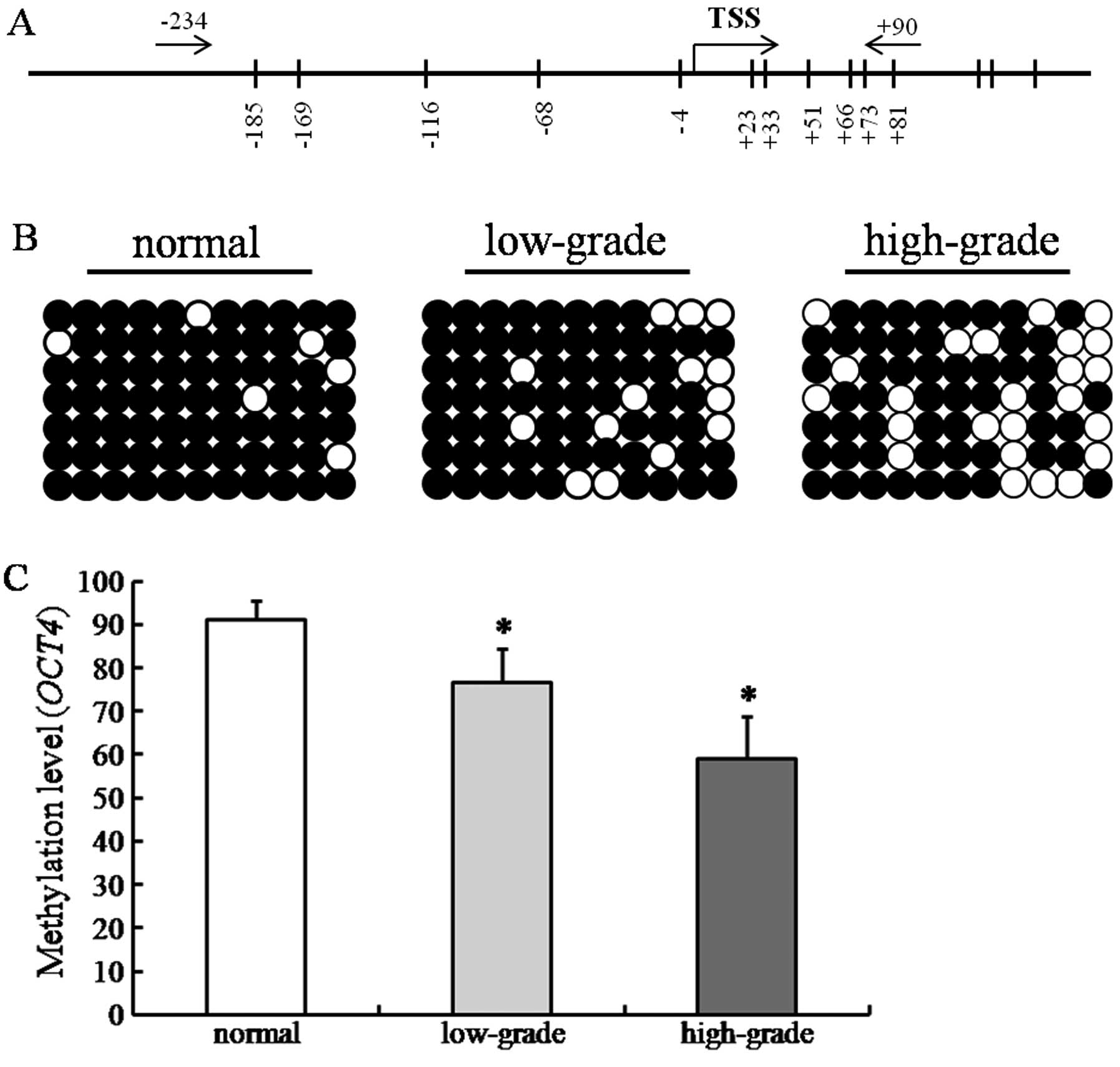
Bisulphate sequencing analysis was employed to
examine whether the OCT4 expression was correlated with the
hypomethylation of promoter and exon. A total of 11 CpG
dinucleotides were measured through BSP sequencing. The minimal
promoter region and the exon region contained 5 and 6 CpG
dinucleotides, respectively (Fig.
3A). DNA methylation of the OCT4 gene was found in all
samples. The CpG dinucleotides in the normal group were highly
methylated. However, the DNA methylation level was reduced with the
upgrading of the glioma malignancy (Fig. 3B). Statistical analysis showed that
DNA methylation level in primary gliomas was significantly lower
than that in the normal group (P<0.05). The percent of
methylation in high-grade gliomas was significantly lower than that
in low-grade ones (P<0.05)(Fig.
3C). When we compared the OCT4 expression profile
(Fig. 1), the DNA methylation level
was negatively correlated to the expression of OCT4 mRNA (r=
−0.156, P<0.05).
Upregulation of OCT4 in glioma cells
following demethylation
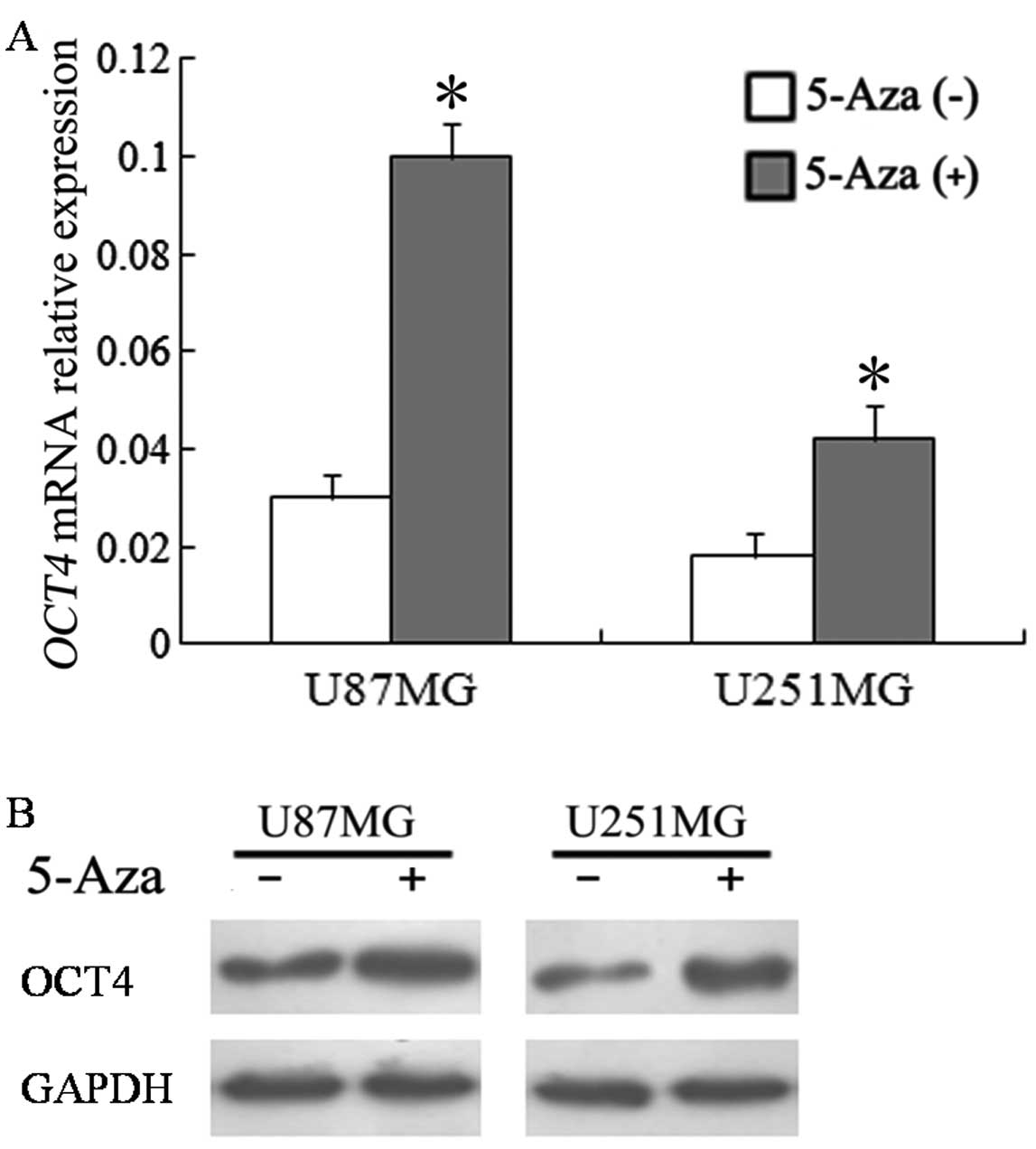
The glioma cell lines (U87MG and U251MG) were
treated with 5-Aza-dc, a demethylation reagent, for 72 h. The
results showed that OCT4 mRNA expression was upregulated 3.38-fold
in U87 cells and 2.35-fold in U251 cells (Fig. 4A). The changing tendency of protein
expression was similar to that of mRNA expression (Fig. 4B).
Discussion
It is considered that most human primary malignant
brain tumors contain BTSCs. These cells are the determinants for
the occurrence, development and recurrence of gliomas and for the
therapeutic response of brain tumor (29,30).
BTSCs provide a novel logical explanation for the drug resistance
and the high recurrence rate of primary malignant brain tumors.
Since BTSCs are similar to neural stem cells in molecular marker
expression and multi-lineage differentiation potential, BTSCs can
be studied indirectly in the absence of the default marker for
screening BTSCs. We hypothesize that the expression level of genes
related to stem-like cells (SLCs) can substitute the rate of SLCs
in glioma. SLC-related genes usually include Nes (nestin), CD133
(prominin), ABCG2 (ATP-binding cassette superfamily G member 2),
SOX2 (SRY-box containing gene 2), OCT4 (POU class 5 homeobox 1) and
Msi-1 (musashi-1). In the present study, we selected OCT4 as the
representative of SLC-related genes.
During the tumorigenesis of some CSCs in adults,
OCT4 expression is detectable since its level is consistent with
the number of CSCs and is closely related to the malignant degree,
development and prognosis of tumors (31). In this study, real-time PCR, western
blot and IHC analyses showed that both OCT4 mRNA and OCT4 protein
were highly expressed in primary gliomas and the levels were
grade-dependent. Our findings are consistent with previous studies
that high-level OCT4 increased the malignancy of ESC-derived tumors
(12–14,31,32).
Moreover, OCT4 expression is regarded as a ‘stem cell survival’
factor pattern and is restricted to pluripotent cells (7,33). Our
results also supported the BTSC theory of glioma, which suggested
that SLCs were present in neoplasm and the malignancy of glioma may
be related to the abundance of SLCs in the tumor.
Numerous studies indicate the DNA methylation of the
OCT4 gene at the gene regulatory region is a key regulatory factor
in OCT4 transcription (22–24,34–36).
Cantz et al(37) also argued
that the most appropriate method to determine OCT4 expression in
cells is to analyze the methylation status of the promoter.
Therefore, to explore whether the DNA methylation of OCT4 promoter
is involved in the changes of OCT4 expression in primary gliomas,
BSP sequencing was employed to measure the methylation of the
minimal promoter region and exon region (−234 ~ +46). We found that
the methylation levels of the OCT4 gene were different between
primary glioma and normal brain tissues. The methylation level in
tumor was markedly reduced as compared to the normal group and was
lower in high-grade gliomas than in low-grade ones. The results
showed that methylation status may regulate the transcription of
the OCT4 gene and were involved in glioma development. OCT4
promoter demethylation has already been reported in other types of
human cancer. It was also suggested that OCT4 promoter
demethylation contributes to tumorigenesis (38,39).
Since the demethylation of the OCT4 promter occurred predominantly
in high-grade glioma, OCT4 may contribute to the initiation and
progression of glioma and potentially serve as a biomarker for the
prognosis of human glioma.
Genomic hypomethylation can cause genome instability
and proto-oncogene formation, which leads to high expression
(40). Our study showed that the
expression of the OCT4 gene was significantly upregulated in glioma
samples, as compared with the normal brain samples. Notably, the
percentage of promoter methylation in the OCT4 gene was inversely
correlated with the expression levels of the OCT4 gene in glioma.
In vitro, 5-Aza-dc reactivated the expression of OCT4 in
glioma cells. These results suggested that aberrant promoter
hypomethylation of OCT4 plays a role in the reexpression of its
normal silence. Fanelli et al(41) reported that DNA hypomethylation in
GBMs is in part explained by a correlating decrease in expression
of de novo DNA methyltransferase DNMT3b. This global change
in methylation leads to the reactivation and remobilization of
repeat elements satellite 2, D4Z4 and Alu elements, resulting in
increased copy number alterations of genomic regions in proximity
to these elements (42). However,
with regard to OCT4, the potential specific mechanism of
hypomethylation in primary glioma requires further study.
In summary, OCT4 is epigenetically regulated by
methylation in primary glioma. The methylation status of the OCT4
gene can be regulated by a demethylation reagent. Our findings
provide evidence for the role of DNA methylation in primary glioma
and present a direction for developing more powerful strategies to
treat glioma in the clinic.
Acknowledgements
This study was supported by the Youth Fund of the
National Natural Science Foundation of China (81201975), the Youth
Fund of the Natural Science Foundation of Jiangsu Province
(BK2012224), the Natural Science Foundation of China Ministry of
Health (2010-2-025), the Natural Science Foundation of Jiangsu
Department of Health (H201124), the Six Major Human Resources
Project of Jiangsu Province (2011-WS-065; 2010-WS-038), the Natural
Science Foundation of Jiangsu Colleges and Universities Grant
(11KJB320010).
References
|
1
|
Bondy ML, Scheurer ME, Malmer B, et al:
Brain tumor epidemiology: consensus from the Brain Tumor
Epidemiology Consortium. Cancer. 113:1953–1968. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yuan X, Curtin J, Xiong Y, et al:
Isolation of cancer stem cells from adult glioblastoma multiforme.
Oncogene. 23:9392–9400. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Galli R, Binda E, Orfanelli U, et al:
Isolation and characterization of tumorigenic, stem-like neural
precursors from human glioblastoma. Cancer Res. 64:7011–7021. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Singh SK, Hawkins C, Clarke ID, et al:
Identification of human brain tumour initiating cells. Nature.
432:396–401. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hemmati HD, Nakano I, Lazareff JA, et al:
Cancerous stem cells can arise from pediatric brain tumors. Proc
Natl Acad Sci USA. 100:15178–15183. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schöler H, Ruppert S, Suzuki N, et al: New
type of POU domain in germ line-specific protein Oct-4. Nature.
344:435–439. 1990.PubMed/NCBI
|
|
7
|
Pesce M and Schöler HR: Oct-4: gatekeeper
in the beginnings of mammalian development. Stem Cells. 19:271–278.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Takeda J, Seino S and Bell GT: Human Oct3
gene family: cDNA sequences, alternative splicing, gene
organization, chromosomal location, and expression at low levels in
adult tissues. Nucleic Acids Res. 20:4613–4620. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Babaie Y, Herwig R, Greber B, et al:
Analysis of Oct4-dependent transcriptional networks regulating
self-renewal and pluripotency in human embryonic stem cells. Stem
Cells. 25:500–510. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Webster JD, Yuzbasiyan-Gurkan V, Trosko
JE, et al: Expression of the embryonic transcription factor Oct4 in
canine neoplasms: a potential marker for stem cell subpopulations
in neoplasia. Vet Pathol. 44:893–900. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chiou SH, Yu CC, Huang CY, et al: Positive
correlations of Oct-4 and Nanog in oral cancer stem-like cells and
high-grade oral squamous cell carcinoma. Clin Cancer Res.
14:4085–4095. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Du Z, Jia D, Liu S, et al: Oct4 is
expressed in human gliomas and promotes colony formation in glioma
cells. Glia. 57:724–733. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Holmberg J, He X, Peredo I, et al:
Activation of neural and pluripotent stem cell signatures
correlates with increased malignancy in human glioma. PloS One.
6:e184542011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ben-Porath I, Thomson MW, Carey VJ, et al:
An embryonic stem cell-like gene expression signature in poorly
differentiated aggressive human tumors. Nat Genet. 40:499–507.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ellis L, Atadja PW and Johnstone RW:
Epigenetics in cancer: targeting chromatin modifications. Mol
Cancer Ther. 8:1409–1420. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Baylin SB and Herman JG: DNA
hypermethylation in tumorigenesis: epigenetics joins genetics.
Trends Genet. 16:168–174. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ehrich M, Turner J, Gibbs P, et al:
Cytosine methylation profiling of cancer cell lines. Proc Natl Acad
Sci USA. 105:4844–4849. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jones PA: Epigenetics in carcinogenesis
and cancer prevention. Ann NY Acad Sci. 983:213–219. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Feinberg AP: Cancer epigenetics takes
center stage. Proc Natl Acad Sci USA. 98:392–394. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nagarajan RP and Costello JF: Molecular
epigenetics and genetics in neuro-oncology. Neurotherapeutics.
6:436–446. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shi G and Jin Y: Role of Oct4 in
maintaining and regaining stem cell pluripotency. Stem Cell Res
Ther. 1:392010. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Feldman N, Gerson A, Fang J, et al:
G9a-mediated irreversible epigenetic inactivation of Oct-3/4 during
early embryogenesis. Nat Cell Biol. 8:188–194. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Deb-Rinker P, Ly D, Jezierski A, et al:
Sequential DNA methylation of the Nanog and Oct-4 upstream regions
in human NT2 cells during neuronal differentiation. J Biol Chem.
280:6257–6260. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gidekel S and Bergman Y: A unique
developmental pattern of Oct-3/4 DNA methylation is controlled by a
cis-demodification element. J Biol Chem. 277:34521–34530. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hattori N, Nishino K, Ko YG, et al:
Epigenetic control of mouse Oct-4 gene expression in embryonic stem
cells and trophoblast stem cells. J Biol Chem. 279:17063–17069.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xue WC, Chan KY, Feng HC, et al: Promoter
hypermethylation of multiple genes in hydatidiform mole and
choriocarcinoma. J Mol Diagn. 6:326–334. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liao X, Siu MK, Chan KY, et al:
Hypermethylation of RAS effector related genes and DNA
methyltransferase 1 expression in endometrial carcinogenesis. Int J
Cancer. 123:296–302. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nordhoff V, Hübner K, Bauer A, et al:
Comparative analysis of human, bovine, and murine Oct-4 upstream
promoter sequences. Mamm Genome. 12:309–317. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Singh SK, Clarke ID, Terasaki M, et al:
Identification of a cancer stem cell in human brain tumors. Cancer
Res. 63:5821–5828. 2003.PubMed/NCBI
|
|
30
|
Singh SK, Clarke ID, Hide T and Dirks PB:
Cancer stem cells in nervous system tumors. Oncogene. 23:7267–7273.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sathomsumetee S, Reardon DA, Desjardins A,
et al: Molecularly targeted therapy for malignant glioma. Cancer.
110:13–24. 2007. View Article : Google Scholar
|
|
32
|
Gidekel S, Pizov G, Bergman Y and Pikarsky
E: Oct-3/4 is a dose-dependent oncogenic fate determinant. Cancer
Cell. 4:361–370. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yeom YI, Fuhrmann G, Ovitt CE, et al:
Germline regulatory element of Oct-4 specific for the totipotent
cycle of embryonal cells. Development. 122:881–894. 1996.PubMed/NCBI
|
|
34
|
Ben-Shushan E, Pikarsky E, Klar A and
Bergman Y: Extinction of Oct-3/4 gene expression in embryonal
carcinoma x fibroblast somatic cell hybrids is accompanied by
changes in the methylation status, chromatin structure, and
transcriptional activity of the Oct-3/4 upstream region. Mol Cell
Biol. 13:891–901. 1993.
|
|
35
|
Simonsson S and Gurdon J: DNA
demethylation is necessary for the epigenetic reprogramming of
somatic cell nuclei. Nat Cell Biol. 6:984–990. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tsuji-Takayama K, Inoue T, Ijiri Y, et al:
Demethylating agent, 5-azacytidine, reverses differentiation of
embryonic stem cells. Biochem Biophys Res Commun. 323:86–90. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Cantz T, Key G, Bleidissel M, et al:
Absence of OCT4 expression in somatic tumor cell lines. Stem Cells.
26:692–697. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang HJ, Siu MK, Wong ES, et al: Oct4 is
epigenetically regulated by methylation in normal placenta and
gestational trophoblastic disease. Placenta. 29:549–554. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hoffmann MJ, Müller M, Engers R and Schulz
WA: Epigenetic control of CTCFL/BORIS and OCT4 expression in
urogenital malignancies. Biochem Pharmacol. 72:1577–1588. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ehrlich M: DNA methylation in cancer: too
much, but also too little. Oncogene. 21:5400–5413. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Fanelli M, Caprodossi S, Ricci-Vitiani L,
et al: Loss of pericentromeric DNA methylation pattern in human
glioblastoma is associated with altered DNA methyltransferases
expression and involves the stem cell compartment. Oncogene.
27:358–365. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Cadieux B, Ching TT, VandenBerg SR and
Costello JF: Genome-wide hypomethylation in human glioblastomas
associated with specific copy number alteration,
methylenetetrahydrofolate reductase allele status, and increased
proliferation. Cancer Res. 66:8469–8476. 2006. View Article : Google Scholar
|