Introduction
Human fibrosarcoma is an aggressive type of tumor
composed of fibroblasts with variable collagen production and, in
classical cases, a herringbone architecture. This tumor is
relatively uncommon, accounting for only 1 to 3% of adult sarcomas
(1,2). It can develop in any fibrous tissue of
the body and strike at any age, but most commonly in middle-aged
and elderly adults. Generally, the incidence of fibrosarcoma is
equal among the genders.
Despite advances in chemotherapies and surgical
treatments, human fibrosarcoma has a higher recurrence rate
compared with these of other soft tissue sarcomas. The probability
of local recurrence relates to the completeness of excision, with
recurrence rates of 12–79% (3–5).
Fibrosarcomas metastasize to lungs and bone, especially the axial
skeleton, but rarely to lymph nodes. Metastasis is time- and
grade-dependent and occurs in 9–63% of patients. The 5-year
survival remains only 39–54% (3,5). Many
patients relapse and their tumors become drug-resistant after a
period of treatment, thus long-term patient survival rates have
remained poor (6).
It is well known that solid tumors consist of
heterogeneous cell populations that include cells with stem-like
properties, such as a high proliferation rate and invasive growth
potential (7,8). The cancer stem cell (CSC) theory
states that these cells exist as a minority population within the
tumor, possess stem cell properties of self-renewal and
multilineage differentiation and are the root cells from which
tumors are derived and sustained (9–11).
CSCs were initially identified and isolated from leukemia patients
(12). The discovery of leukemic
stem cells prompted further research into other types of cancer,
leading to the identification of CSCs in numerous solid tumors,
including cancers of the breast (13), brain (14), prostate (15) and other different types of sarcomas
(16–18). CSCs are postulated to be resistant
to standard cytotoxic chemotherapeutic agents both in vitro
and in vivo, and this resistance may be the major cause of
treatment failure, as the surviving reservoir of stem cells can
populate the tumor and lead to relapse. New therapies targeting
these cells, which are fundamental for tumor progression, can
significantly improve the clinical management of cancer (19). Therefore, identifying within tumors
subpopulations of cells exhibiting significant alterations in
proliferation, stem cell marker expression and behavior are
essential to understanding cancer pathology.
The cell surface glycoprotein CD133 is a pentaspan
transmembrane protein that was originally identified in
hematopoietic and neural stem cells (20). Interest in CD133 as a CSC marker has
grown dramatically since it was associated with a cancer-initiating
subpopulation in the brain (14)
and prostate (21). Moreover,
CD133+ cells have also been found in various sarcomas,
including Ewing’s sarcoma and osteosarcoma (22–26).
However, until now, it has not been determined whether
CD133+ cells with tumorigenic potential can be
prospectively isolated from human fibrosarcomas. In this study, a
population of HT1080 human fibrosarcoma cells expressing CD133 was
identified and fulfilled the criteria of being characterized as
CSCs both in vivo and in vitro. These cells may be
highly valuable targets for the therapy of recurrent and
chemoresistant disease.
Materials and methods
Cell culture
The HT1080 fibrosarcoma cell line was obtained from
the American Type Culture Collection (ATCC; Manassas, VA, USA) and
grown in Dulbecco’s modified Eagle’s medium (DMEM) with 10% fetal
bovine serum (FBS; HyClone, Logan, UT, USA) and 1%
penicillin/streptomycin (Quality Biological, Gaithersburg, MD,
USA), and maintained at 37°C (humidified, in 5% CO2).
The medium was replaced every 2–3 days, and the monolayer cultures
were trypsinized and recultured when reaching 70–80% confluency.
Single-cell suspensions for flow cytometry and cell sorting were
generated by trypsinization with a solution of 0.25% trypsin with 1
mM ethylenediaminetetraacetic acid (EDTA).
Magnetic-activated cell sorting (MACS)
and flow cytometric analysis
Cultured cells were trypsinized and resuspended in
MACS buffer [PBS without Ca2+ and Mg2+,
supplemented with 0.5% bovine serum albumin (BSA) and 2 mM EDTA] at
a concentration of 1×108 cells/300 μl. A minimum of
106 events was collected and acquired using CellQuest
software (Becton-Dickinson, Franklin Lakes, NJ, USA). For MACS,
single-cell suspensions were then incubated with FcR blocking
reagent (1 μl per million cells) and CD133 microbeads (1 μl per
million cells) for 30 min at 4°C. After washings, CD133+
cells were separated according to the manufacturer’s
recommendations by positive selection on LS columns, and the
CD133− cells were collected in the flow-through. Prior
to and following separation, samples were analyzed by
fluorescence-activated cell sorting (FACS) using CD133/2
phycoerythrin (293C) antibody and isotype control mouse IgG2b
phycoerythrin (1/10) in a FACSCalibur flow cytometer (BD
Biosciences, San Jose, CA, USA). The purity of CD133+
and CD133− cell populations was evaluated by standard
flow cytometric analysis. All reagents were purchased from Miltenyi
Biotech (Bergisch Bladbach, Germany).
Hoechst 33342 exclusion assay
HT1080 fibrosarcoma cells were resuspended at
1.0×106 cells/ml in pre-warmed DMEM culture medium and
divided into 2 portions. One portion was treated with 50 mM
verapamil (Sigma, St. Louis, MO, USA) as an inhibitor of the ABC
transporter, and the other was left untreated. Both portions were
incubated in DMEM culture medium with 5 μg/ml Hoechst 33342 (Sigma)
for 90 min at 37°C. Cells were washed in PBS after the incubation,
kept on ice for 5 min and analyzed for Hoechst 33342 efflux using a
BD FACS Aria II (BD Biosciences, Franklin Lakes, NJ, USA). The
Hoechst 33342 dye was excited at 357 nm, and the fluorescence was
analyzed at dual wavelengths (blue, 402–446 nm; red, 650–670
nm).
Sphere formation assay
Cells expressing CD133 were enriched using CD133
microbeads and a MACS device. To further improve the purity of the
CD133− fraction, a second sorting step was performed by
adding fresh CD133 microbeads to the CD133− cell
fraction and running a depletion protocol on the MACS. Single
CD133+ or CD133− cells were seeded into low
attachment 96-well plates (Corning Incorporated, Corning, NY, USA).
Visual inspection was performed the day after the initial plating
to confirm that each well contained a single cell. After 3–4 weeks,
spheres containing >50 cells were counted. Cells were inoculated
into N2/1% methylcellulose medium without serum. The N2 medium
consisted of 2X DMEM/F12 (Invitrogen Life Technologies, Carlsbad,
CA, USA) supplemented with progesterone (20 nM), putrescine (100
μM), sodium selenite (30 nM), transferrin (25 μg/ml), insulin (20
μg/ml), human recombinant epidermal growth factor (EGF; 10 ng/ml)
and basic fibroblast growth factor (bFGF; 10 ng/ml), mixed with an
equal volume of 2% methylcellulose. All reagents were purchased
from Sigma. Fresh aliquots of EGF and bFGF were added every other
day. After 3–4 weeks of culture, spherical colonies were quantified
using an Olympus CK2 inverted phase contrast microscope (Olympus,
Japan). The capacity for secondary sphere formation was also
investigated by repeated dissociation and re-culturing in
anchorage-independent, serum-starved conditions on ultra low
attachment plates.
Efficacy of chemotherapy drugs on
CD133+ and CD133− cells
The CD133+ and CD133− cells
were dissociated, seeded into 96-well microtiter plates (Corning
Incorporated) at a density of 2,000 cells/100 μl/well and allowed
to attach in DMEM/10% FBS for 8 h at 37°C. Cells were then exposed
to 10 μl of cisplatin (CDDP) or doxorubicin (DXR) (both from Sigma)
at different concentrations for 48 h. Viabilities of both
drug-treated and non-treated cells were measured by the MTS
colorimetric assay using the CellTiter 96® AQueous One
Solution Cell Proliferation assay kit (Promega Corporation,
Madison, WI, USA), according to the manufacturer’s instructions.
Drug resistance in CD133+ spheroids and adherent cells
was compared for a more detailed characterization of the stemness
properties of the CD133+ cell population. The
CD133+ cells were seeded into 96-well ultra low
attachment microplates (Corning Incorporated) in N2/1%
methylcellulose media at a density of 5,000 cells/100 μl/well for 2
weeks to allow sphere formation. CD133+ spheroids and
adherent cells were then treated with 10 μl of CDDP or DXR at
different concentrations for 48 h, and cell viability was assessed
by the MTS assay.
Protein isolation and western blot
analysis
Sorted cells were lysed in a buffer containing 50 mM
Tris-HCl (pH 7.4), 150 mM NaCl 1 mM EDTA, 1% NP-40, 0.1% SDS, 1%
Na-deoxycholate, 1 mM Na-vanadate, and protease inhibitors (5 mg/ml
pepstatin, 1 mM PMSF, 10 mg/ml leupeptin and 1 mM NaF) (Sigma) for
1 h in ice. After centrifugation at 13,000 × g for 10 min at 4°C,
the protein concentrations of the supernatants were measured by the
BCA protein assay kit (Pierce Biotechnology, Inc., Rockford, IL,
USA). Lysates were mixed (1:1) with Laemmli buffer (Sigma). Fifty
micrograms of protein was electrophoresed per lane in 10% SDS
polyacrylamide gels and transferred onto PVDF membranes (Sigma).
Membranes were blocked with nonfat milk for 1 h at room temperature
and incubated overnight at 4°C with the corresponding antibodies in
Tris-buffered saline (TBS) with 5% BSA (Sigma) and 0.1% Tween-20
(Bio-Rad Laboratories, Hercules, CA, USA). After being washed 3
times in TBS with 0.1% Tween-20, blots were incubated with
Ig-specific, peroxidase-conjugated secondary antibodies for the
appropriate species. Immunoreactive bands were detected with the
ECLPlus SuperSignal West Pico chemiluminescent substrate (Thermo
Scientific, Rockford, IL, USA) for 60 sec. Protein levels were
normalized with respect to the band density of
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) as an internal
control. Primary antibodies against Nanog, Oct3/4, Sox2, ABCG2,
c-Myc and Bmi-1 (all diluted 1:1,000) were purchased
from Cell Signaling Technology Inc. (Danvers, MA, USA). The
GAPDH-specific antibody (1:5,000) was purchased from Santa Cruz
Biotechnology, Inc. (Santa Cruz, CA, USA).
Real-time PCR
cDNA was obtained using Moloney murine leukemia
virus reverse transcriptase and RNase H minus (Promega
Corporation). Typically, 250 ng of template total RNA and 250 ng of
random hexamers were used per reaction. Real-time PCR amplification
was performed using the TaqMan Universal PCR Master Mix and
Assays-On-Demand gene expression products or Power SYBR-Green
Master Mix and specific PCR primers in an ABI Prism 7900 instrument
(Applied Biosystems, Carlsbad, CA, USA). Relative quantification of
each target, normalized to an endogenous control (cyclophilin A),
was performed using the comparative Ct method (Applied Biosystems).
Probes were used to detect c-Myc, Sox2, Oct3/4, Bmi-1, Nanog
and ABCG2. The primer sequences used for amplification of
the above genes are listed in Table
I. The GAPDH gene was used as an internal control for
normalizing template amounts.
 | Table IPrimers for Sox2, Oct3/4, Nanog,
c-Myc, Bmi-1 and ABCG2. |
Table I
Primers for Sox2, Oct3/4, Nanog,
c-Myc, Bmi-1 and ABCG2.
| Gene | Forward
(5′-3′) | Reverse
(5′-3′) |
|---|
| c-Myc |
GCATACATCCTGTCCGTCCA |
CAAGAGTTCCGTAGCTGTTCAAG |
| Sox2 |
ATCACCCACAGCAAATGACA |
CAAAGCTCCTACCGTACCACTA |
| Oct3/4 |
TATTCAGCCAAACGACCATCT |
TCAGCTTCCTCCACCCACTT |
| Bmi-1 |
CTCCACCTCTTCTTGTTTGC |
GATGACCCATTTACTGATGATTT |
| Nanog |
AGGCAAACAACCCACTTCT |
‘TCACACCATTGCTATTCTTCG |
| ABCG2 |
GATAAAGTGGCAGACTCCAAGGT |
CCAATAAGGTGAGGCTATCAAACA |
| GAPDH |
GGGAAACTGTGGCGTGAT |
GAGTGGGTGTCGCTGTTGA |
In vivo tumorigenicity studies
Groups of 6 female nude athymic mice (6- to 8-weeks
old) (BALB/c nu/nu; Vital River Laboratory Animal Center, Beijing,
China) were injected with either sorted or unsorted cells at
different concentrations subcutaneously into the right flank in a
1:1 mixture of Matrigel and Hank’s balanced salt solution (HBSS) in
a final volume of 300 μl. Engrafted mice were inspected biweekly
for tumor development by visual observation and palpation. Mice
were sacrificed by cervical dislocation when the tumor diameter
reached 15 mm or at 4 months post-transplantation. Xenograft tumors
were excised using aseptic techniques and processed for hematoxylin
and eosin (H&E) staining and anti-CD133 immunohistochemical
analysis. In addition, tumor samples were digested using
collagenase II (Sigma), and the CD133+ cells were
isolated and immediately re-injected into mice to generate
second-round tumors. All animal procedures were approved by the
Institutional Animal Care and Use Committee in Teaching and
Research of Harbin Medical University.
Histopathological and immunohistochemical
analysis
All samples of the mouse transplants were placed in
10% formalin immediately, processed with routine histological
procedures and embedded in paraffin. Serial sections (5-μm) were
cut and either stained with H&E for routine histological
observation under a light microscope or used for
immunohistochemical detection of CD133. After deparaffinization
(hydration), sections were treated sequentially with normal goat
serum, anti-CD133/2 antibody (Miltenyi; clone 293C3),
biotin-labeled goat anti-mouse IgG and avidin-biotin-peroxidase
complex (ABC). Peroxidase binding sites were detected by the
diaminobenzidine method. Sections were then counterstained with
hematoxylin for microscopic examination. The number and area of
CD133-positive foci >0.2 mm in diameter and the total area of
the examined sections were measured using an Olympus C5050Z digital
camera. Images were acquired digitally using MagnaFire Software
(Optronics) and processed in Adobe Photoshop.
Statistical analysis
All assays were repeated at least 3 times. Results
were reported as means ± standard deviation (SD) after analysis by
using SPSS 19.00 software. Differences with P-values <0.05 were
considered significant.
Results
Separation of CD133+ and
CD133− fractions from the HT1080 fibrosarcoma cell
line
CD133 is a phylogenetically conserved cell surface
molecule that was first described in neuro-endothelial progenitors
and recently has been proposed to be a selective marker for CSCs in
many cancer types. We first assessed the presence and size of the
cell populations expressing CD133 on the surface of HT1080
fibrosarcoma cells by standard flow cytometry. Analysis of this
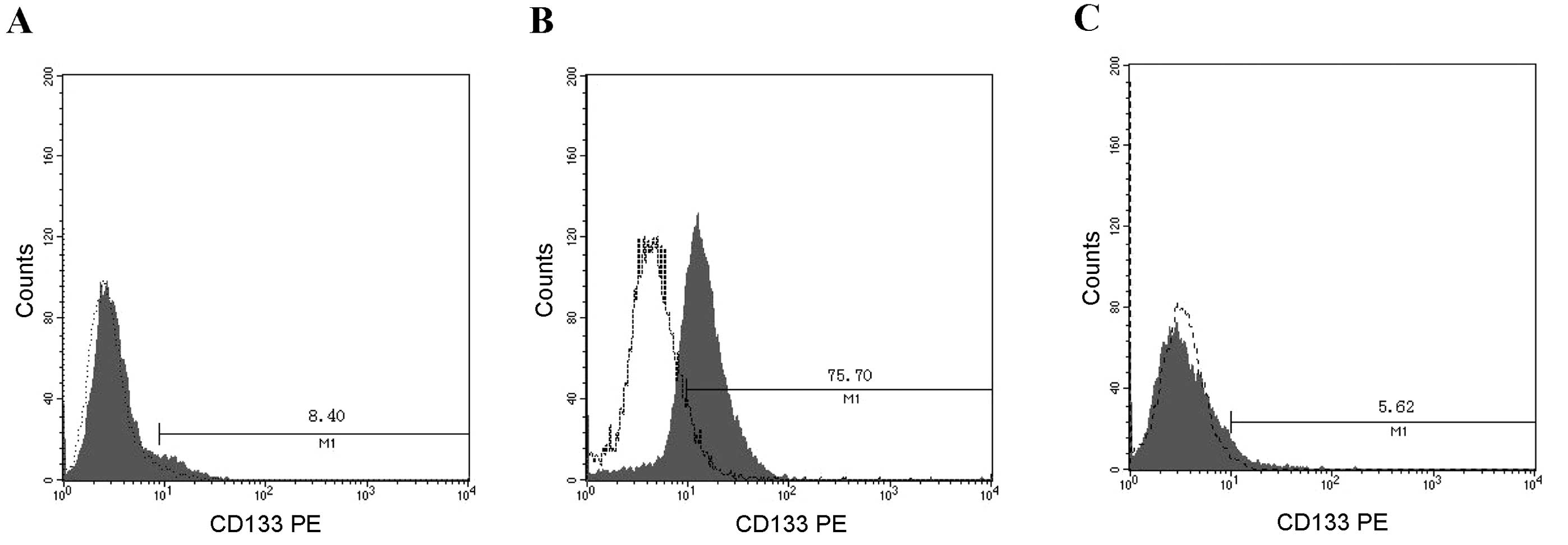
fibrosarcoma model cell line revealed that it consistently
contained a CD133+ subpopulation at a frequency of 8.40%
in HT1080 cells. Subsequently, the CD133+ and
CD133− fractions were separated by MACS sorting. As
shown in Fig. 1, a
CD133+ enriched population (75.70%) and a
CD133− cell population (94.38%) were obtained (Fig. 1).
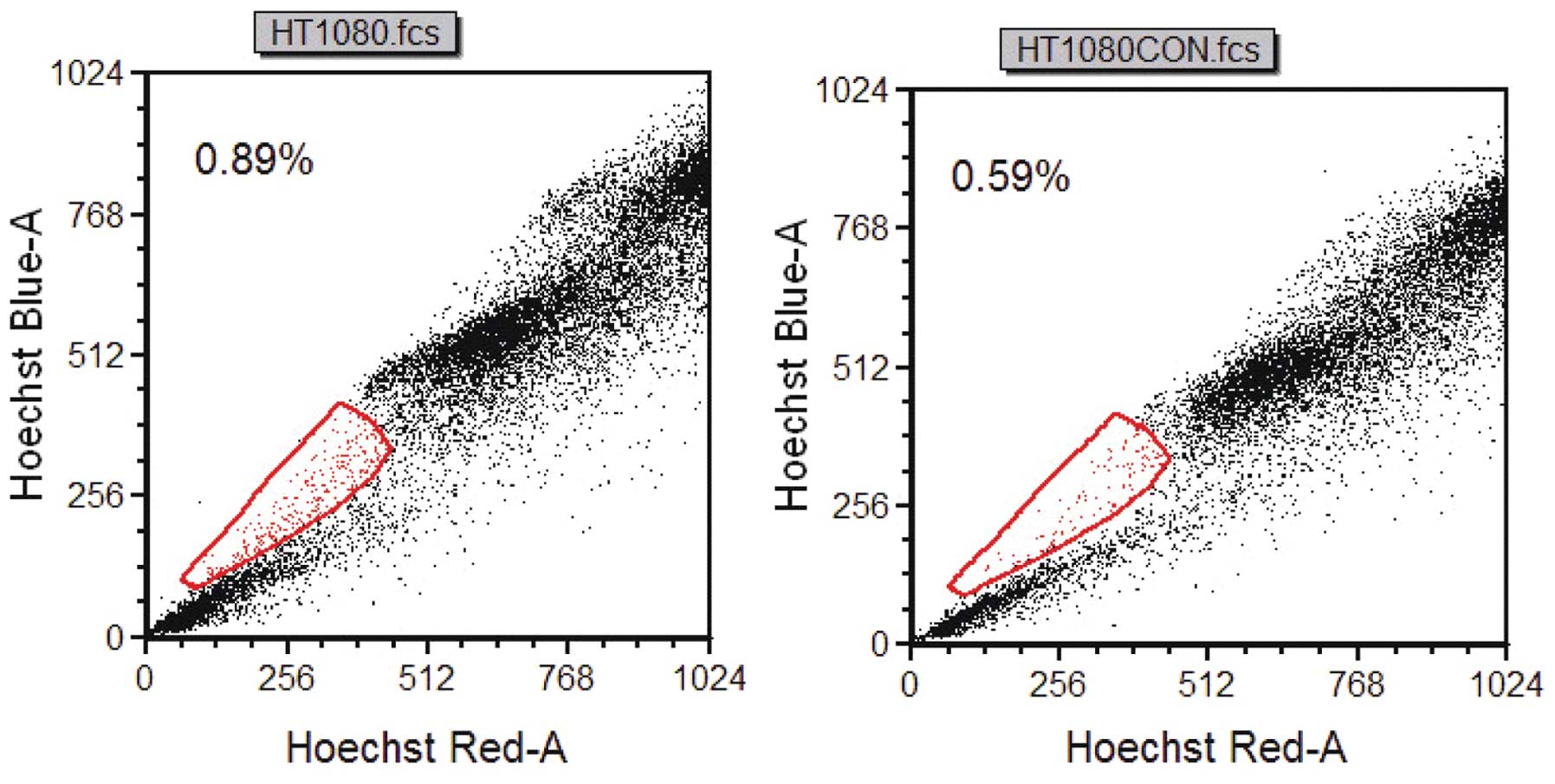
Side population
A very small subset (0.30%) of the HT1080 cell
displayed the characteristic profile of a side population. The side
population phenotype is considered the most significant attribute
of CSCs. The percentage of side population cells was markedly
diminished by treatment with verapamil, an inhibitor of the pumps
responsible for the exclusion of the Hoechst 33342 dye, indicating
that this population truly represented side population cells. Thus,
this result showed the detection of a side population in a
fibrosarcoma cell line (Fig.
2).
Functional characterization of
CD133+ cells
Although no in vitro assay can specifically
identify CSCs, some growth properties, such as spheroid formation,
can be assessed in culture as a potential characteristic of CSCs.
Therefore, the ability of freshly isolated CD133+ and
CD133− cells to form spheres in serum-free culture
conditions was tested. Because of the high likelihood of this assay
containing potential CD133+ contaminant cells in the
negative fraction, a second round of depletion was performed on the
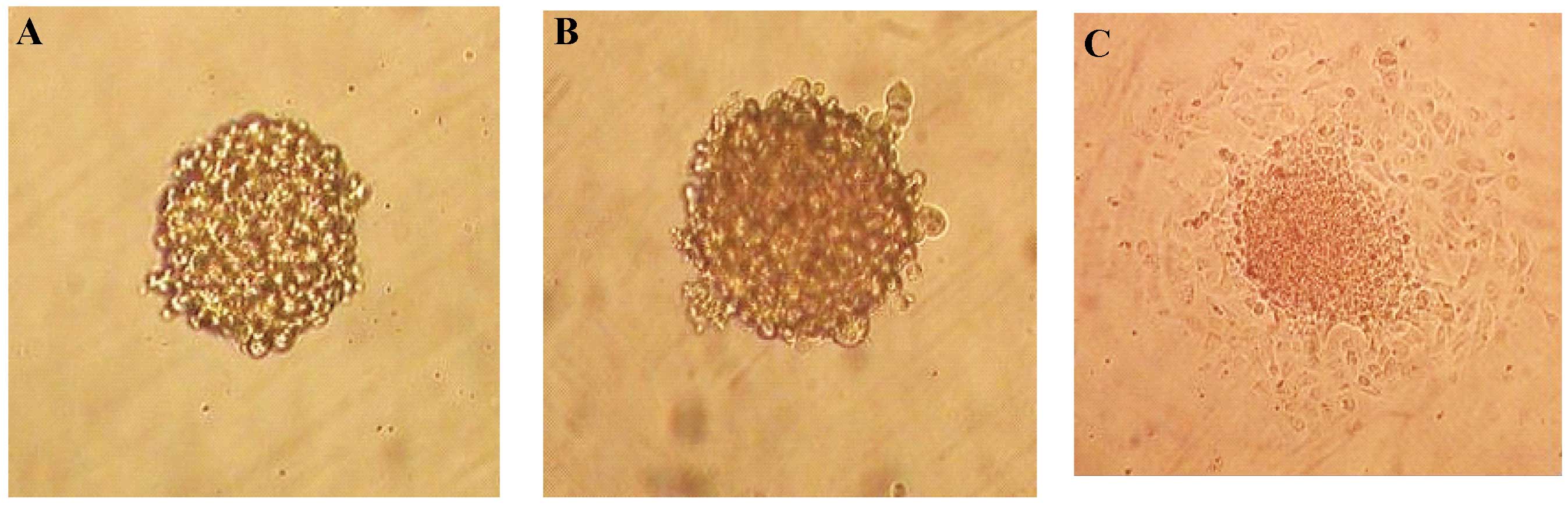
CD133− cells. Under serum-free conditions, the
CD133+ population proliferated as floating spheres
(Fig. 3A), whereas most of the
CD133− population did not survive. By re-culturing in
anchorage-independent, serum-starved conditions, cells dissociated
from the spheres were shown to be capable of repeatedly forming
spheres (Fig. 3B). When floating
spheres of CD133+ cells were seeded into
serum-containing medium, the cells migrated from the spheres within
a few hours and adhered to the bottom of the flasks, showing
spindle-like morphology similar to that of the original cell
culture (Fig. 3C). Single
CD133+ or CD133− cells were individually
seeded into ultra low adhesion 96-well plates that favor the
proliferation of undifferentiated cells. The number of spheres was
scored after 3–4 weeks in culture. Importantly, the
CD133+ fraction gave rise to an ~5–6-fold greater number
of spheres than the CD133− fraction, and this increase
was statistically significant (Table
II).
| Table IIShere-forming efficiency of
individual CD133+ and CD133− cells plated in
N2 medium for 3–4 weeks. |
Table II
Shere-forming efficiency of
individual CD133+ and CD133− cells plated in
N2 medium for 3–4 weeks.
|
CD133+ |
CD133− |
|---|
| Wells with
spheres | 64/384 | 11/384 |
| Sphere-forming
efficiency | 16.7% | 2.86% |
CD133+ cells have increased
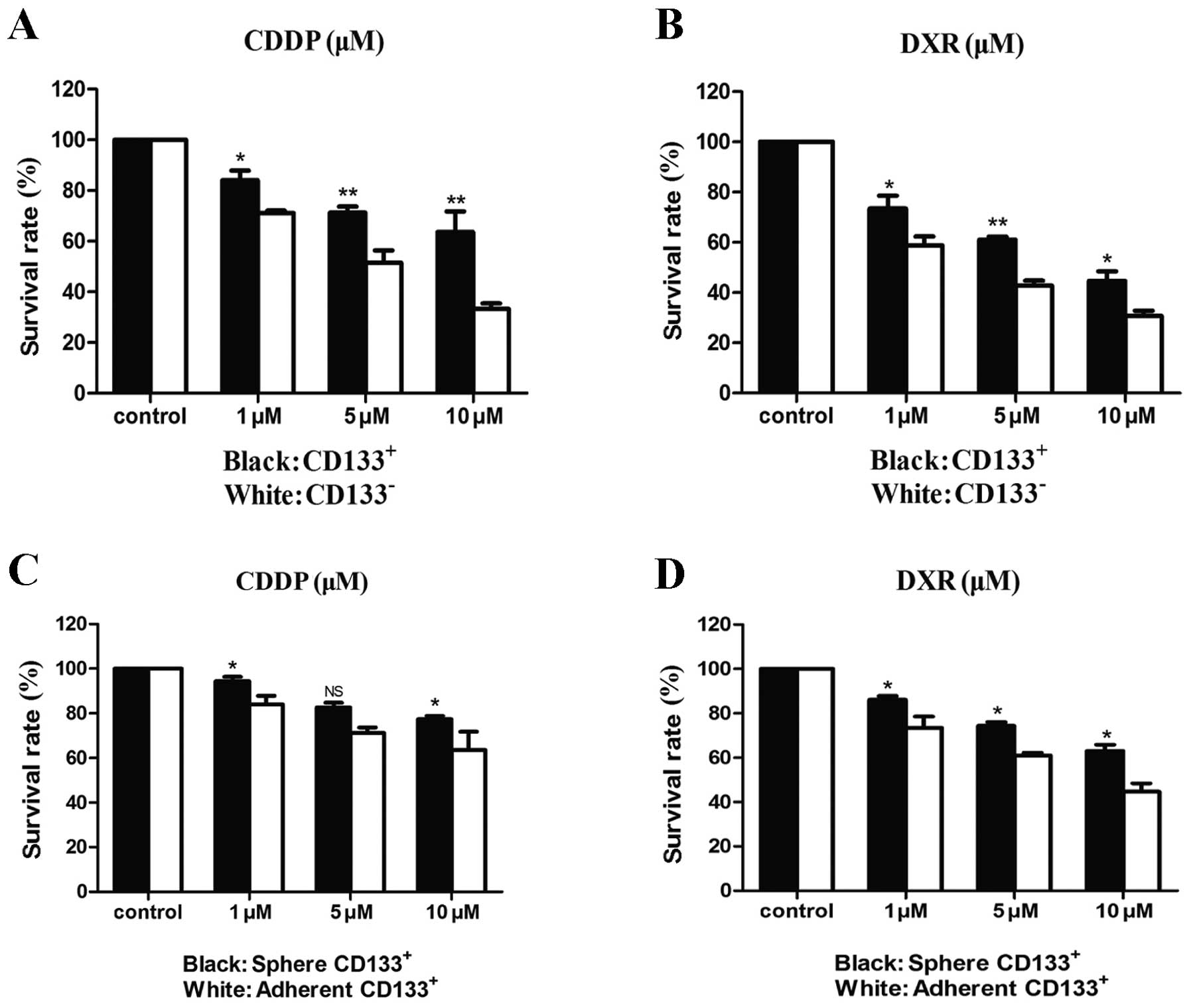
resistance to chemotherapeutic drugs
CSCs have been reported to be more resistant to
chemotherapy than other tumor cells. Therefore, viability was
examined for the sorted CD133+ and CD133−
cells that were exposed to increasing doses of cisplatin (CDDP) or
doxorubicin (DXR), drugs which are currently in use in the clinical
setting for the treatment of fibrosarcoma. As shown in Fig. 4, both CD133+ and
CD133− cells displayed a dose-dependent sensitivity to
these chemotherapeutic agents. However, the CD133+
population was significantly resistant to both CDDP and DXR in
comparison with the CD133− population. To better
understand how the stem cell properties of the CD133+
cell population could confer resistance to therapy, differences
between CD133+ spheroids in serum-free media and
CD133+ adherent cells grown in serum-containing media
were examined. The spheroids were found to be more resistant to
both chemotherapeutics in comparison with the adherent cells.
CD133+ cells are enriched in
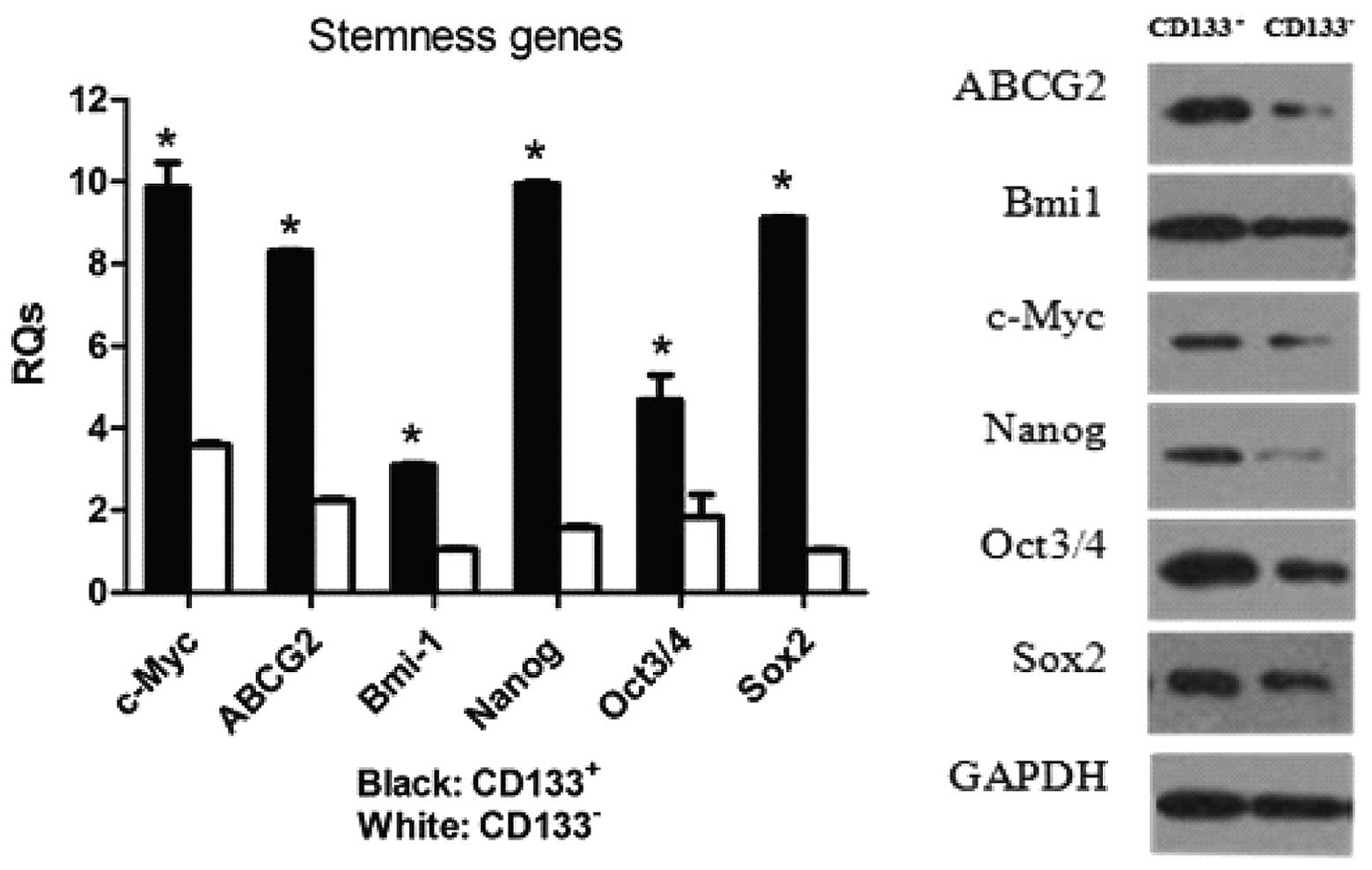
expression of ‘stem cell’ genes
Real-time PCR and western blotting were used to
examine whether the CD133+ cells were enriched in the
expression of genes that have been postulated to play key roles in
stem cell biology (Fig. 5).
Real-time PCR analysis revealed that the expression levels of genes
involved in the maintenance of stemness, namely Sox2 and
Nanog, were significantly higher in the CD133+
population in comparison with the CD133− population.
Moreover, expression levels of the Oct3/4, c-Myc,
Sox2 and Bmi-1 genes were consistently increased in
the CD133+ population compared with their
CD133− counterparts. A 6.3-fold increase in Nanog
mRNA and an 8.8-fold increase in Sox2 mRNA in
CD133+ cells compared to CD133− cells were
detected by real-time PCR. The level of c-Myc was also
upregulated in CD133+ cells in comparison with
CD133− cells, while Bmi-1 expression was nearly
the same in the 2 populations according to the western blotting
results. In addition, both gene and protein expression levels of
the ABCG2 drug transporter were significantly increased in
the CD133+ cells when compared with the levels in the
CD133− cells.
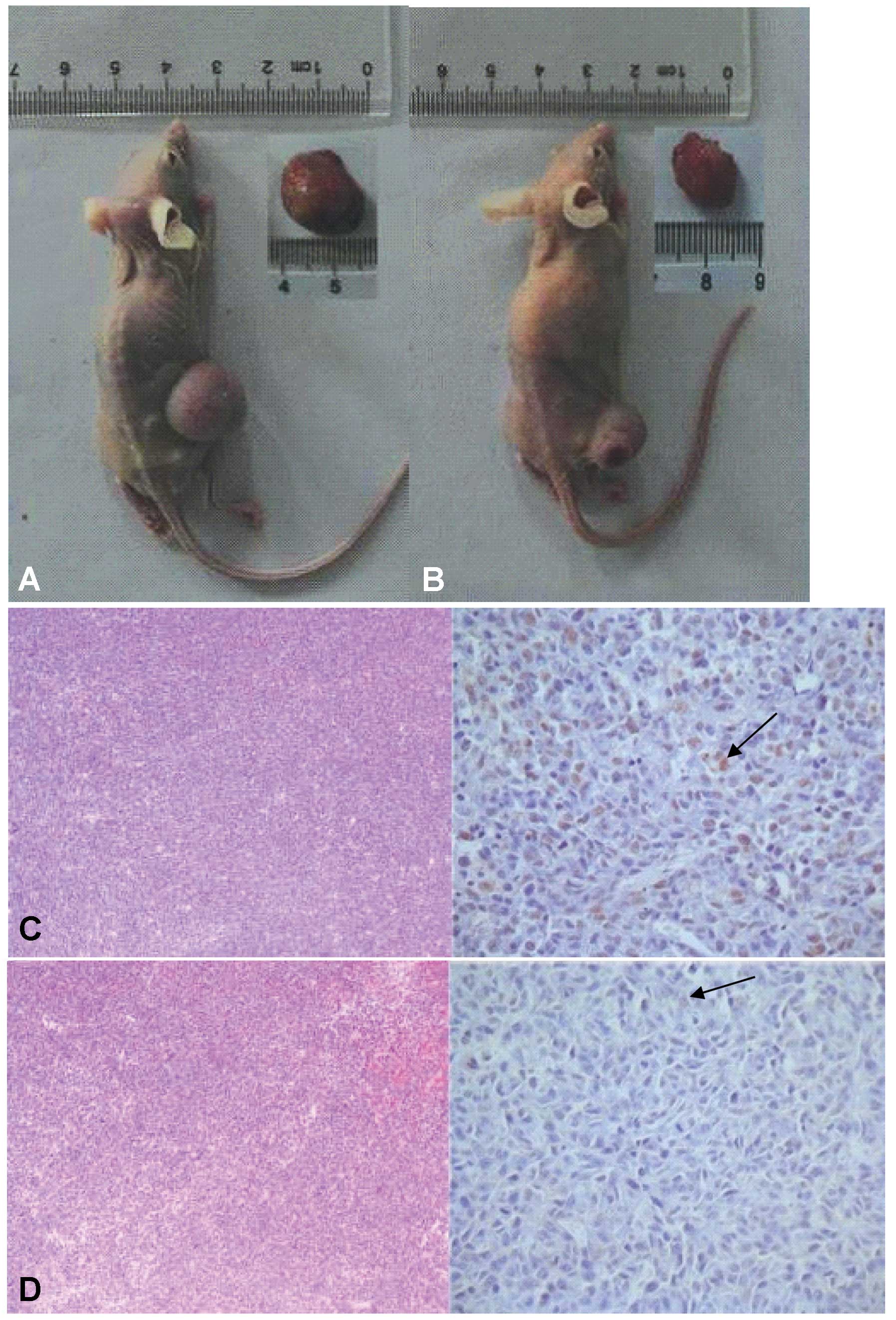
CD133+ cells exhibit
tumor-initiating activity in vivo
The definition of cancer-initiating cells relies
mainly on their functional properties such as self renewal. The
other defining characteristic of CSCs is recapitulating the
cellular heterogeneity of the original tumor. Therefore, the
tumorigenic capacity of HT1080-derived CD133+ and
CD133− cell populations was investigated. Cells were
MACS-sorted into CD133+ and CD133− fractions
and expanded in culture as independent populations for 3 to 5
passages. Subsequently, a range of CD133+ or
CD133− cells (200-106) were injected
subcutaneously into the right flank of individual BALB/c nude mice.
Unsorted cells were injected as a control. After 6–10 weeks, tumors
were detected in the flank region with an average size of 15 mm.
Tumor formation was noted after injection of as few as
103 CD133+ cells (Table III). By contrast, only 4 of 6 of
the mice injected with 106 CD133− cells and
none of the mice injected with 105 or fewer
CD133− cells developed a tumor, suggesting that this
population was relatively depleted of tumor-initiating cells.
Injections of unsorted cells also reliably resulted in tumor
formation, and 3 out of 6 mice injected with 105
unsorted cells developed a tumor. Importantly, tumors that arose
from CD133+ cells could be serially transplanted into
secondary recipients. Gross tumor appearance and histology were
similar for tumors arising from the unsorted cells,
CD133+ cells and CD133− cells.
Immunohistochemistry of these xenografts demonstrated that only a
small minority of tumor cells expressed a high level of CD133, even
in tumors that arose from implantation of purified
CD133+ cells (Fig. 6),
further supporting the hypothesis that they represent true CSCs
able to generate serially transplantable tumors containing mostly
CD133− non-stem cells. Together these results provide
preliminary in vivo evidence that the tumor-initiating
potential of CD133+ cells is greater than that of
CD133− cells and that CD133+ tumor-initiating
cells undergo asymmetric cell division during tumor growth to
generate both CD133+ and CD133− progeny.
| Table IIITumor initiating capacity of various
cell populations. |
Table III
Tumor initiating capacity of various
cell populations.
| Cell number
injected |
|---|
|
|
|---|
| 200 | 103 | 104 | 105 | 106 |
|---|
| Unsorted | | 0/6 | 0/6 | 3/6 | 5/6 |
|
CD133+ | 0/6 | 3/6 | 5/6 | 6/6 | |
|
CD133− | | 0/6 | 0/6 | 0/6 | 4/6 |
Discussion
Fibrosarcoma, a malignant tumor which prevailingly
affects adults, is associated with higher recurrence and metastasis
rates in comparison with other soft tissue sarcomas. The concept
that a small fraction of cancerous cells residing within each tumor
with stem cell properties, which are responsible for tumor
initiation, invasive growth and metastasis, is generally accepted.
CSCs are hypothesized to have a phenotype defined by their cell of
origin and retain the stem cell properties of self-renewal,
capacity to differentiate and drug resistance. Therefore, new
therapies targeting these cells may significantly improve the
clinical outcome of cancer patients. However, obtaining conclusive
evidence for the existence of CSCs has been a significant challenge
in cancer research. Recently, various studies using phenotypical
analyses have reported stem-like cell populations in numerous types
of cancers (8,10,12).
The presence of a CD133+ subpopulation with stem-like
features within human solid tumors has been documented both in
vitro and in vivo by many reports (14,21–25).
In the present study, the CD133 antigen was used to demonstrate
that the subpopulation expressing this marker within the stabilized
fibrosarcoma cell line HT1080 carries some characteristics of stem
cells, including the capacity to generate a heterogeneous
population, in vitro clonogenic activity and in vivo
tumorigenic activity. The CD133+ subset was found to be
8.40% of the total fibrosarcoma cells, consistent with the
assumption that CSCs comprise only a very small proportion of tumor
cells. In addition, a side population also was detected in the
HT1080 cell line for the first time in this study. The ability of
cancer cells to exclude small lipophilic molecules such as Hoechst
33342 depends on the presence of ABC (ATP-binding cassette)
transporters actively pumping out these molecules, and this
phenomenon has been exploited for the identification of putative
stem cells in a variety of human cancers (27–29).
The side population was found to comprise a much smaller fraction
(0.30%) of the total cells in this study, Therefore, this may be
considered to be an effective sorting approach for CSCs.
CD133+ cells in this study showed many
differences with respect to their negative counterparts both in
vitro and in vivo. In assays generally utilized to
assess indices of self-renewal, tumorigenicity and clonogenicity,
CD133+ cells displayed a stronger ability to form
spherical colonies in anchorage-independent, serum-starved culture
conditions. This finding was also supported by use of tumor
xenografts in vivo. Only 1,000 CD133+ cells were
needed to form fibrosarcomas, and these cells remained tumorigenic
in continuous passages, while up to 100,000 injected
CD133− cells could not form tumors in nude mice. The
results confirmed that the tumor-initiating potential of
CD133+ cells was much greater than that of the
CD133− cells.
More recent evidence has indicated that there is a
mutual genomic fingerprint between embryonic stem (ES) cells, adult
tissue stem cells and CSCs. Thus, relative expression levels of
genes that play an important role in the maintenance and promotion
of stem cell pluripotency and nuclear reprogramming, include
Sox2, Oct3/4, Nanog, c-Myc and Bmi-1 in
CD133+ and CD133− cells. The Sox
family of transcription factors plays a critical role in cell
differentiation and development. Sox2 was originally thought
to be the only Sox protein expressed in ES cells (30,31).
The Oct3/4 transcription factor and Nanog are
critically involved in self-renewal and the maintenance of
pluripotency in undifferentiated ES cells (32,33).
c-Myc is recognized as a dominant-acting oncogene that
encodes a transcription factor thought to regulate the G0-G1 cell
cycle transition. Bmi-1 is a member of the mammalian
polycomb group (Pc-G) gene family and has recently emerged as a
Myc-cooperating oncogene, which contributes to the proliferative
capacity and self-renewal of both normal and malignant stem cells
(34–36). All genes above were shown by
real-time PCR in this study to be distinctly overexpressed in the
CD133+ fraction. These data provide compelling evidence
that a CD133+ subpopulation with stem-like properties
exists in HT1080 cells.
To further investigate the CSC properties of
CD133+ cells, the sensitivity of cells to chemotherapy
drugs was examined. The results showed that CD133+ cells
were more resistant to standard cytotoxic chemotherapy in
comparison with the CD133− population. Advanced cancer
is generally initially responsive to standard chemotherapies, but
that response is almost inevitably followed by development of a
drug-resistant phenotype. One increasingly accepted hypothesis of
chemoresistance posits that standard therapies fail to target tumor
progenitors, which are believed to express normal stem cell
phenotypes, such as a low mitotic index, enhanced DNA repair and
expression of membrane efflux transporters. The ABC superfamily of
proteins contribute to multidrug resistance by pumping chemotherapy
drugs out of the cell (37,38). ABCG2, a member of this family
and a membrane transporter, is associated with the side population
phenotype. Thus, chemosensitizers are being developed for the
reversal of these transporters which play a prominent role in the
multidrug resistance mechanism of tumors. The expression of
ABCG2 was recently reported in many types of cancers, and it
was interpreted as a conserved feature of stem cells (39–41).
Consistent with that hypothesis, CD133+ cells were shown
here to express high levels of ABCG2, suggesting a
significant role for these cells in fibrosarcoma chemoresistance.
However, targeted therapy with an ABCG2 antagonist can only
partially inhibit the growth of CD133+ cells, perhaps
due to expression by CSCs of other drug-resistant proteins such as
ABCB1 and ABCC1(38,42,43).
Moreover, the CD133+ cell-derived spheres showed greater
resistance than the CD133+-adherent cells. These results
suggest that although CD133 is present in stem-like populations,
using it as a single marker to isolate these cells would be
insufficient. Further study is required to determine additional
definitive surface markers for fibrosarcoma stem cells, such as
ABCG2, that may be used in concert with CD133 to sort
CSCs.
The above results revealed that CD133 may be used as
a marker for isolation of fibrosarcoma CSCs from the human
fibrosarcoma cell line HT1080. The isolated CD133+ cell
subpopulation was demonstrated to have CSC characteristics. This
finding prompts further study to more precisely define sarcoma stem
cells and their mechanisms of drug resistance. A better
understanding of fibrosarcoma CSC biology will facilitate the
determination of prognosis and treatments. In particular,
expression studies of fibrosarcoma CSCs will help to identify more
specific diagnostic markers and therapeutic targets.
Acknowledgements
This research was supported by a grant from the
National Natural Science Foundation of China (no. 30772205 and no.
81072192) and a special financial grant from the China Postdoctoral
Science Foundation (no. 201104450).
References
|
1
|
Bahrami A and Folpe AL: Adult-type
fibrosarcoma: a reevaluation of 163 putative cases diagnosed at a
single institution over a 48-year period. Am J Surg Pathol.
34:1504–1513. 2010.PubMed/NCBI
|
|
2
|
Fisher C: The value of electron microscopy
and immunohistochemistry in the diagnosis of soft tissue sarcomas:
a study of 200 cases. Histopathology. 16:441–454. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pritchard DJ, Soule EH, Taylor WF and
Ivins JC: Fibrosarcoma - a clinicopathologic and statistical study
of 199 tumors of the soft tissues of the extremities and trunk.
Cancer. 33:888–897. 1974. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pritchard DJ, Sim FH, Ivins JC, et al:
Fibrosarcoma of bone and soft tissues of the trunk and extremities.
Orthop Clin North Am. 8:869–881. 1977.PubMed/NCBI
|
|
5
|
Scott SM, Reiman HM, Pritchard DJ and
Ilstrup DM: Soft tissue fibrosarcoma. A clinicopathologic study of
132 cases. Cancer. 64:925–931. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wibmer C, Leithner A, Zielonke N, et al:
Increasing incidence rates of soft issue sarcomas? A
population-based epidemiologic study and literature review. Ann
Oncol. 21:1106–1111. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Al-Hajj M and Clarke MF: Self-renewal and
solid tumor stem cells. Oncogene. 23:7274–7282. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hermann PC, Bhaskar S, Cioffi M and
Heeschen C: Cancer stem cells in solid tumors. Semin Cancer Biol.
20:77–84. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bansal N and Banerjee D: Tumor initiating
cells. Curr Pharm Biotechnol. 10:192–196. 2009. View Article : Google Scholar
|
|
10
|
O’Brien CA, Kreso A and Dick JE: Cancer
stem cells in solid tumors: an overview. Semin Radiat Oncol.
19:71–77. 2009.
|
|
11
|
Reya T, Morrison SJ, Clarke MF and
Weissman IL: Stem cells, cancer, and cancer stem cells. Nature.
414:105–111. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bonnet D and Dick JE: Human acute myeloid
leukemia is organized as a hierarchy that originates from a
primitive hematopoietic cell. Nat Med. 3:730–737. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Al-Hajj M, Wicha MS, Benito-Hernandez A,
et al: Prospective identification of tumorigenic breast cancer
cells. Proc Natl Acad Sci USA. 100:3983–3988. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Singh SK, Clarke ID, Terasaki M, et al:
Identification of a cancer stem cell in human brain tumors. Cancer
Res. 63:5821–5828. 2003.PubMed/NCBI
|
|
15
|
Collins AT, Berry PA, Hyde C, et al:
Prospective identification of tumorigenic prostate cancer stem
cells. Cancer Res. 65:10946–10951. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen SY, Huang YC, Liu SP, et al: An
overview of concepts for cancer stem cells. Cell Transplant.
20:113–120. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gibbs CP, Kukekov VG, Reith JD, et al:
Stem-like cells in bone sarcomas: implications for tumorigenesis.
Neoplasia. 7:967–976. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li C, Heidt DG, Dalerba P, et al:
Identification of pancreatic cancer stem cells. Cancer Res.
67:1030–1037. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ciavarella S, Milano A, Dammacco F and
Silvestris F: Targeted therapies in cancer. BioDrugs. 24:77–88.
2010. View Article : Google Scholar
|
|
20
|
Mizrak D, Brittan M and Alison M: CD133:
molecule of the moment. J Pathol. 214:3–9. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ceder JA, Jansson L, Ehrnström RA, et al:
The characterization of epithelial and stromal subsets of candidate
stem/progenitor cells in the human adult prostate. Eur Urol.
53:524–531. 2008. View Article : Google Scholar
|
|
22
|
Curley MD, Therrien VA, Cummings CL, et
al: CD133 expression defines a tumor initiating cell population in
primary human ovarian cancer. Stem Cells. 27:2875–2883.
2009.PubMed/NCBI
|
|
23
|
Rutella S, Bonanno G, Procoli A, et al:
Cells with characteristics of cancer stem/progenitor cells
expresses the CD133 antigen in human endometrial tumors. Clin
Cancer Res. 15:4299–4311. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Suvà ML, Riggi N, Stehle JC, et al:
Identification of cancer stem cells in Ewing’s sarcoma. Cancer Res.
69:1776–1781. 2009.
|
|
25
|
Tirino V, Desiderio V, d’Aquino R, et al:
Detection and characterization of CD133+ cancer stem
cells in human solid tumors. PLoS One. 3:e34692008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang J, Wang H, Li Z, et al: c-Myc is
required for maintenance of glioma cancer stem cells. PLoS One.
3:e37692008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hadnagy A, Gaboury L, Beaulieu R and
Balicki D: SP analysis may be used to identify cancer stem cell
populations. Exp Cell Res. 312:3701–3710. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Moserle L, Ghisi M, Amadori A and
Indraccolo S: Side population and cancer stem cells: therapeutic
implications. Cancer Lett. 288:1–9. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wu C and Alman BA: Side population cells
in human cancers. Cancer Lett. 268:1–9. 2008. View Article : Google Scholar
|
|
30
|
Kelberman D, de Castro SC, Huang S, et al:
SOX2 plays a critical role in the pituitary, forebrain, and
eye during human embryonic development. J Clin Endocrinol Metab.
93:1865–1873. 2008. View Article : Google Scholar
|
|
31
|
Maruyama M, Ichisaka T, Nakagawa M and
Yamanaka S: Differential roles for Sox15 and Sox2 in
transcriptional control in mouse embryonic stem cells. J Biol Chem.
280:24371–24379. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Medvedev SP, Shevchenko AI, Mazurok NA and
Zakiian SM: OCT4 and NANOG are the key genes in the system of
pluripotency maintenance in mammalian cells. Genetika.
44:1589–1608. 2008.(In Russian).
|
|
33
|
Tokuzawa Y, Kaiho E, Maruyama M, et al:
Fbx15 is a novel target of Oct3/4 but is dispensable for embryonic
stem cell self-renewal and mouse development. Mol Cell Biol.
23:2699–2708. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Brunk BP, Martin EC and Adler PN:
Drosophila genes posterior sex combs and suppressor two of zeste
encodes proteins with homology to the murine bmi-1 oncogene.
Nature. 353:351–353. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Iwama A, Oguro H, Negishi M, et al:
Enhanced self-renewal of hematopoietic stem cells mediated by the
polycomb gene product Bmi-1. Immunity. 21:843–851. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lessard J and Sauvageau G: Bmi-1
determines the proliferative capacity of normal and leukaemic stem
cells. Nature. 423:255–260. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Choi CH: ABC transporters as multidrug
resistance mechanisms and the development of chemosensitizers for
their reversal. Cancer Cell Int. 5:302005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Vasiliou V, Vasiliou K and Nebert DW:
Human ATP-binding cassette (ABC) transporter family. Hum Genomics.
3:281–290. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jin Y, Bin ZQ, Qiang H, et al: ABCG2 is
related with the grade of glioma and resistance to mitoxantone, a
chemotherapeutic drug for glioma. J Cancer Res Clin Oncol.
135:1369–1376. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liang Y, Russell I, Walworth C and Chen C:
Gene expression in stem cells. Crit Rev Eukaryot Gene Expr.
19:289–300. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Pascal LE, Oudes AJ, Petersen TW, et al:
Molecular and cellular characterization of ABCG2 in the prostate.
BMC Urol. 7:62007. View Article : Google Scholar
|
|
42
|
Takara K, Sakaeda T and Okumura K: An
update on overcoming MDR1 mediated multidrug resistance in cancer
chemotherapy. Curr Pharm Des. 12:273–286. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Sharom FJ: ABC multidrug transporters:
structure, function and role in chemoresistance. Pharmacogenomics.
9:105–127. 2008. View Article : Google Scholar : PubMed/NCBI
|