Introduction
Prostate cancer (PCa) is one of the most common
lethal malignant diseases in males in the United States (1). Disease confined to the prostate is
treatable, while metastatic PCa eventually leads to accelerated
disease progression and mortality (2). Thus, to further improve patient
survival, it is essential to further understand the molecular and
cellular mechanisms of PCa development and to establish novel
therapeutic strategies to target PCa progression and
metastasis.
microRNAs (miRNAs) are a class of highly conserved,
small non-coding, single-stranded RNAs, 17–25 nucleotides in
length, that regulate gene expression via post-transcriptional
inhibition of protein synthesis (3). These small RNAs negatively regulate
approximately one-third of human coding genes by binding to
complementary sites in the 3′-untranslated region (3′-UTR) of their
target genes (4). miRNAs may also
play critical roles in the pathogenesis of PCa. It is anticipated
that identification and characterization of PCa-related miRNAs may
shed light on understanding the molecular mechanisms of disease
development and are therefore crucial for the development of novel
therapies.
The miR-200 family (including miR-200a, miR-200b,
miR-200c and miR-141) is a cluster of miRNAs highly correlated with
epithelial-mesenchymal transition (EMT), wherein miR-200b was
identified as a critical regulator of tumor invasion, metastasis
and chemosensitivity (5).
Increasing evidence suggests that miR-200b acts as a
tumor-suppressive miRNA and is a critical regulator of EMT, cancer
stem cell (CSC) maintenance, apoptosis and cancer chemosensitivity
(6,7). However, the functional role and
mechanism of involvement of miR-200b during the development and
pathogenesis of PCa remain to be determined.
B-cell-specific Moloney murine leukemia virus
insertion site 1 (Bmi-1) is a member of the polycomb (PcG) family
of epigenetic gene silencers. Bmi-1 is frequently overexpressed in
several types of human cancers and may play an important role in
carcinogenesis (8–11). Ectopic expression of Bmi-1 can
directly promote EMT and malignancy in human nasopharyngeal
epithelial cells, whereas silencing endogenous Bmi-1 expression can
reverse EMT (12). Bmi-1 also plays
a critical role in the maintenance of stem cells (13) and is a crucial regulator of
self-renewal in adult prostate cells (14). However, the regulatory mechanism of
Bmi-1 and its role in PCa cells require further research.
In the present study, we hypothesized that miR-200b
may influence the biological behaviors of PCa cells by inhibiting
the expression of Bmi-1. Our results revealed that upregulation of
miR-200b effectively downregulated the expression of Bmi-1 and also
inhibited cell proliferation and cell migration and promoted
chemosensitivity. These findings provide evidence that miR-200b
acts as a tumor-suppressor gene and suggest that miR-200b may be a
potential new strategy to prevent prostate progression by targeting
the oncogene Bmi-1. Moreover, we demonstrated that silencing of
Bmi-1 revealed its function as an oncogene in PCa.
Materials and methods
Tissue specimens
Human PCa specimens were obtained from 30 patients
who underwent radical prostatectomies at the Department of Urology,
Shanghai First People’s Hospital, School of Medicine, Shanghai
Jiaotong University between March 2011 and December 2012. Fifteen
benign prostatic hyperplasia (BPH) tissue samples used as the
control were obtained by transurethral resection of the prostate
(TURP). All of the samples were confirmed by pathological
examination and stored in liquid nitrogen for miRNA analysis.
Formalin-fixed, paraffin-embedded samples for immunohistochemistry
from 60 PCa tissues and 30 BPH tissues obtained by radical
prostatectomy and TURP, respectively, were from the Tissue Paraffin
Block Bank of Pathology in our hospital. The Institutional Review
Board of Shanghai First People’s Hospital approved all experimental
procedures, and patient consent was obtained before tissue
collection.
Cell lines and cell culture
Human PCa cell lines LNCaP, PC3 and DU145 were
obtained from the Shanghai Cell Bank, the Chinese Academy of
Sciences, and the benign hyperplastic epithelial cell line BPH-1
was developed in our laboratory. All cell lines were maintained in
RPMI-1640 medium (Gibco-BRL, Rockville, MD, USA) supplemented with
50 U/ml penicillin, 50 mg/ml streptomycin and 10% fetal bovine
serum (Gibco-BRL) in a humidified atmosphere at 37°C in 5%
CO2.
Real-time quantitative RT-PCR
Total RNA from PCa cell lines was extracted using
TRIzol reagent (Invitrogen Life Technologies, Carlsbad, CA, USA).
Real-time quantitative RT-PCR was carried out using the PrimeScript
Reverse Transcription System and SYBR Premix Ex Taq™ II kit
(Takara, Dalian, China) according to the manufacturer’s
instructions. The primer sequences used were as follows: Bmi-1
(forward primer, 5′-TGGACTGACAAATGCTGGAG-3′ and reverse primer,
5′-GGCAAACAAGAAGAGGTGGA-3′); internal control GAPDH (forward
primer, 5′-TCGACAGTCAGCCGCATCTTCTTT-3′ and reverse primer,
5′-ACCAAATCCGTTGACTCCGACCTT-3′). PCR conditions included
denaturation at 94°C for 2 min, followed by 40 cycles of 5 sec at
94°C, 30 sec at 60°C and 30 sec at 72°C.
Expression of mature miR-200b was assayed using
stem-loop RT followed by real-time PCR analysis. Bulge-Loop™ miRNA
qRT-PCR Primer Set and miRNA qRT-PCR Control Primer Set (RiboBio,
Guangdong, China) were used for quantitative real-time PCR analysis
of miR-200b and U6 small nuclear RNA, respectively. PCR conditions
included denaturation at 95°C for 20 sec, followed by 40 cycles of
10 sec at 95°C, 20 sec at 60°C and 10 sec at 70°C. U6 expression
was used as an internal control for miR-200b expression. The
relative expression fold change of miRNAs and mRNAs was calculated
using the 2−ΔΔCt method (15). All experiments were performed in
triplicates.
Transient transfections
PC3 and DU145 cells seeded at 1.5×105
cells/well in 6-well plates were transfected with miR-200b mimic
and a scramble control (RiboBio) performed at a concentration of 50
nM using Lipofectamine 2000 reagent (Invitrogen Life Technologies)
according to the manufacturer’s instructions. Transfection
efficiency was confirmed by qRT-PCR. For the silencing experiments,
PC3 and DU145 cells were transfected with small interfering RNAs
(siRNAs) targeting Bmi-1 (siRNA-Bmi-1) and their negative controls
(siRNA-NC) (RiboBio) using Lipofectamine 2000 reagent according to
the manufacturer’s instruction. The sequences of the siRNA were
identical to those described in a previous study (16). Forty-eight hours after transfection,
cell proliferation and migration were assayed.
Cell Counting Kit-8 (CCK-8) and EdU
analysis
Cell proliferation was assessed using the CCK-8
assay (Dojindo, Kumamoto, Japan). The transfected PC3 and DU145
cells were plated with 100 μl culture mediun in 96-well plates at a
density of 3,000 cells/well. After 24, 48 and 72 h, 10 μl of CCK-8
reagent (5 mg/ml) was added to each well, and incubation was
carried out at 37 °C for 2 h. Viable cells were evaluated by
absorbance measurements at 450 nm. Each assay was performed in 6
replicates for 3 independent experiments. The impact of miR-200b on
PC3 and DU145 cell proliferation was also assessed by the EdU cell
proliferation assay kit, according to the manufacturer’s
instructions. The assays were performed as recommended by the
manufacturer of the EdU detection kit Apollo 488 (RiboBio).
Migration assay
The migratory capacity of the PCa cells following
miR-200b and siRNA-Bmi-1 or NC transfection was assessed using a
Corning Transwell assay according to the manufacturer’s protocol. A
total of 0.5×105 PCa cells in 200 μl of serum-free
medium were seeded into the upper chamber of the system. Lower
chambers were filled with 0.75 ml of complete medium. After 24 h of
incubation, the cells in the upper chamber were removed with a
cotton swab, and the transmigrated cells were fixed in methanol and
stained with crystal violet. Stained cells were counted by
photographing 5 fields/membrane.
Chemosensitivity assay
Rates of sensitivity to drugs were determined by the
cell proliferation reagent CCK-8. Forty-eight hours after
transfection, cells were digested and plated in 96-well plates at a
density of 3,000 cells/well. After an overnight incubation, the
cells were treated with docetaxel (10 nM). After 72 h, 10 μl of
CCK-8 reagent was added to each well and the plate was incubated at
37°C for 2 h. Viable cells were evaluated by absorbance
measurements at 450 nm. Each assay was performed in 6 replicates in
3 independent experiments.
Immunofluorescence staining
BPH-1, LNCaP, PC3 and DU145 cells were grown on
glass coverslips, fixed with 4% paraformaldehyde, permeabilized
with 0.5% Triton X-100 and then blocked with 5% bovine serum
albumin in phosphate-buffered saline (PBS). The coverslips were
then exposed to the primary antibody, anti-Bmi-1, overnight at 4°C,
followed by secondary antibodies. Images were captured using a
laser scanning confocal fluorescence microscope (LSM-510; Carl
Zeiss, Jena, Germany).
Western blot analysis
Cells were washed twice with cold PBS and
homogenized in ice-cold RIPA buffer (Beyotime, Jiangsu, China)
containing phosphatase and protease inhibitors. Total protein was
separated by denaturing SDS-polyacrylamide gel electrophoresis and
transferred electrophoretically onto PVDF membranes. Membranes were
incubated with the primary antibodies for rabbit anti-Bmi-1, rabbit
anti-E-cadherin, vimentin, P16 and rabbit anti-GAPDH (Cell
Signaling Technology, Inc., Beverly, MA, USA) at 4°C overnight and
subsequently with horseradish peroxidase-conjugated secondary
antibody. Signals were visualized with an ECL chemiluminescence kit
(Boster, Wuhan, China) and exposed to X-ray film.
Immunohistochemical staining of
tissues
Immunohistochemistry was conducted on archived
paraffin-embedded formalin-fixed tissues of the study patients.
Bmi-1 immunohistochemistry was performed according to the
avidin-biotin-peroxidase complex method, with primary antibodies
specific against human Bmi-1 (Cell Signaling Technology, Inc.) used
at a 1:400 dilution. The secondary antibody (Long Island Biotech,
Shanghai, China) was applied at a dilution of 1:400. Bmi-1
immunoreactivity was assessed by the intensity of the positive
reaction (no staining, 0; low staining, 1; medium staining, 2;
strong staining, 3). Immunohistochemical evaluation was performed
in a blinded manner by two independent pathologists.
Statistical analyses
Statistical analyses were performed using the
Statistical Package for the Social Sciences software version 17.0
(SPSS, Inc., Chicago, IL, USA). Data are presented as means ± SD
from at least 3 independent experiments. The two-tailed
t-test and χ2 tests were used to assess
statistically significant differences. A P-value <0.05 was
considered to indicate a statistically significant result.
Results
miR-200b is reduced in PCa when compared
with normal prostate cells and tissues
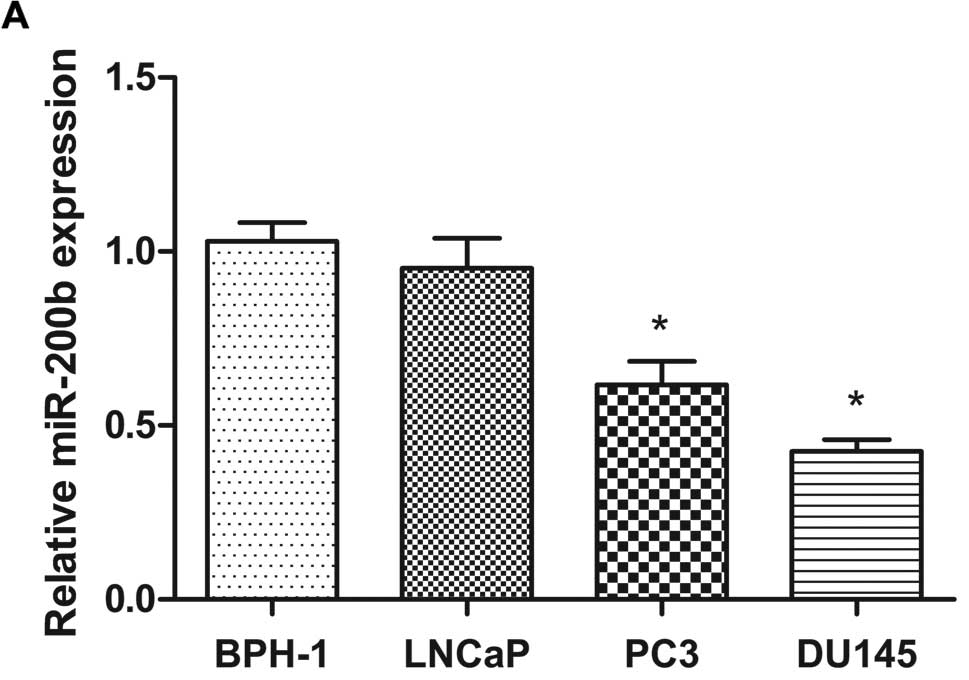
We determined the expression levels of miR-200b in
human prostate cell lines, which included prostate carcinoma cell
lines (LNCaP, PC3 and DU145) and the benign hyperplastic epithelial
cell line BPH-1 by real-time qPCR analysis. We found that miR-200b
was expressed at significantly lower levels in the PC3 and DU145
cells when compared with that in the non-malignant BPH-1 cells,
while there was no difference between the BPH-1 and LNCaP cells
(Fig. 1A). We next investigated the
clinical relevance of miR-200b. Its expression levels were analyzed
by real-time qPCR in PCa tissue and BPH tissue samples. miR-200b
expression was significantly downregulated in the cancer tissues
when compared with that in the BPH tissue samples (P<0.05)
(Fig. 1B).
miR-200b suppresses cell proliferation,
migration and improves chemosensitivity to docetaxel
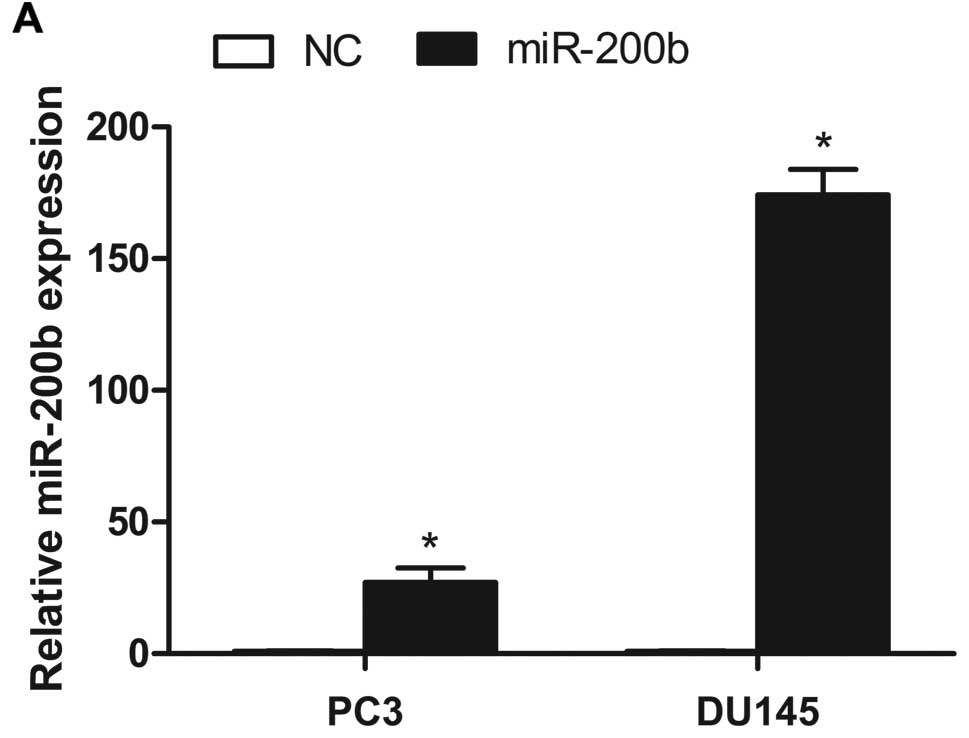
We performed gain-of-function studies using miR-200b
mimics, and miRNA transfection efficiency was determined by
real-time qRT-PCR. After PC3 and DU145 cells were transfection with
50 nM miR-200b mimics and negative control, the expression of
miR-200b was significantly upregulated (Fig. 2A). The effects of ectopic expression
of miR-200b on cell proliferation in PC3 and DU145 cells were
examined by CCK-8 assay. Transfection with miR-200b mimics resulted
in a significant decrease in cell growth of the PCa cell lines
(P<0.05) (Fig. 2B). We further
investigated the effect of miR-200b on cell proliferation using an
EdU incorporation assay. Fewer EdU-positive cells were observed in
the miR-200b mimic-transfected cells when compared to the
NC-transfected cells (Fig. 2C).
These data indicate that miR-200b has a vital role in reducing the
growth of PCa cells.
A Transwell assay was introduced to investigate
whether miR-200b regulates tumor migration. PC3 and DU145 cells
were transfected with either miR-200b mimics or the negative
control. PCa cells transfected with miR-200b exhibited a decrease
in cell motility (Fig. 2D). Our
results are consistent with a previous study that miR-200b inhibits
EMT and cancer cell migration (17). Moreover, miR-200b-treated PC3 and
DU145 cells showed higher chemosensitivity to 10 nM docetaxel than
the NC-treated cells (Fig. 2B),
demonstrating that upregulation of miR-200b improved PC3 and DU145
cell chemosensitivity to docetaxel.
miR-200b inhibits Bmi-1 and increases
E-cadherin expression in PCa cells
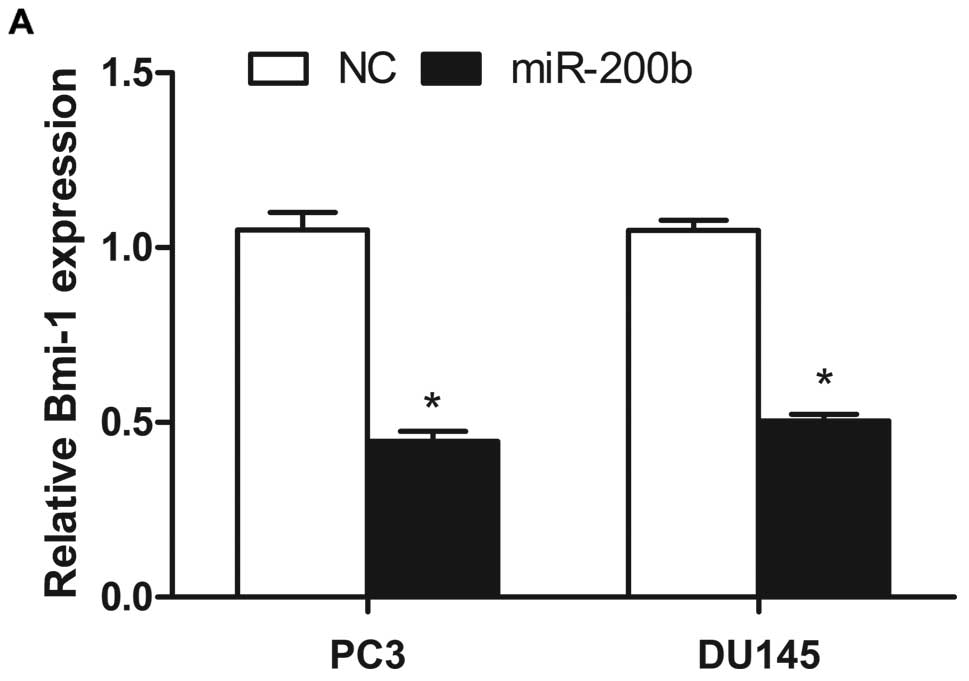
Real-time qRT-PCR demonstrated that Bmi-1 mRNA
expression was decreased in PC3 and DU145 cells after transfection
with miR-200b mimics when compared with the negative control
(Fig. 3A). Furthermore, the protein
expression level of Bmi-1 was also markedly reduced in the PCa cell
lines after miR-200b transfection (Fig.
3B). These results indicate that Bmi-1 may be a target of
miR-200b in PC3 and DU145 cells. Furthermore, enforced expression
of miR-200b in PC3 and DU145 cells resulted in increased expression
of E-cadherin and reduced the expression of vimentin when compared
with the control cells (Fig.
3C).
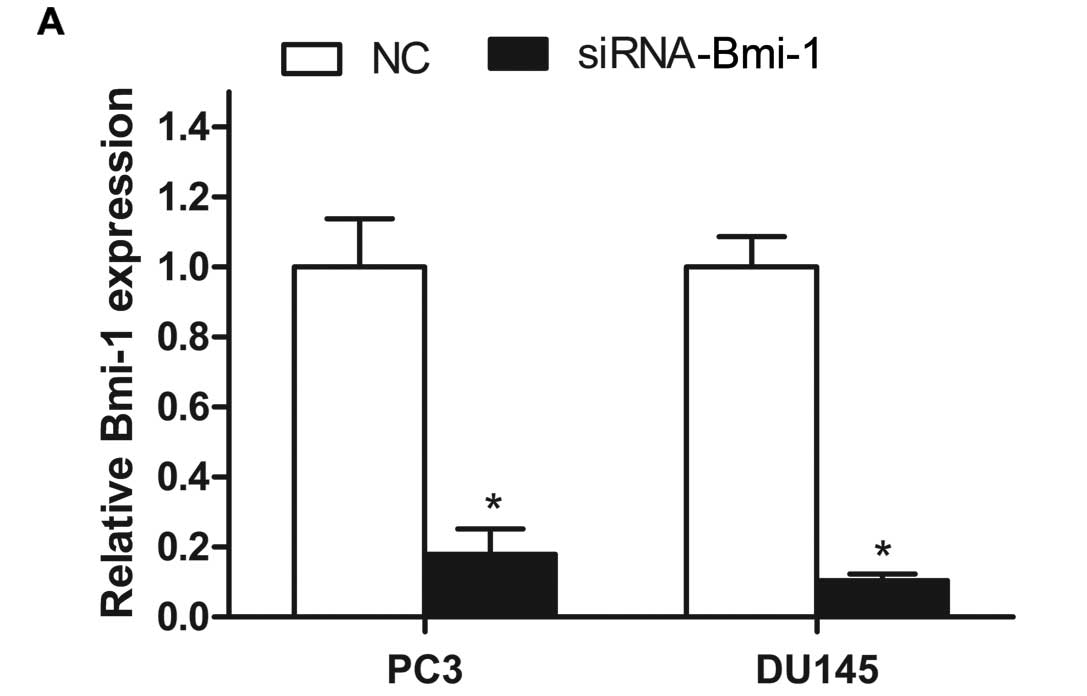
Bmi-1 is efficiently inhibited by
siRNA
To explore the biological functions of Bmi-1 in PC3
and DU145 cells, we transfected these two cell lines with
siRNA-Bmi-1. The efficiency of siRNA-Bmi-1 inhibition was measured
by qRT-PCR and western blotting. The Bmi-1 mRNA (Fig. 4A) and protein (Fig. 4D) levels were downregulated in the
PC3 and DU145 cells following transfection with Bmi-1 siRNA. These
data showed that the expression of Bmi-1 was efficiently silenced
by Bmi-1 siRNA in the PCa cells. We selected the Bmi-1 siRNA to
further study the functional consequences of downregulation of
Bmi-1 expression on cell growth and migration.
Bmi-1 modulates PCa cell proliferation,
motility and chemosensitivity
Transfection of PC3 and DU145 cells with siRNA-Bmi-1
significantly decreased cell proliferation when compared with the
corresponding negative controls (Fig.
4B). Since previous studies have shown that ectopic Bmi-1
suppresses the expression of tumor-suppressor p16 (18), in the present study the p16 protein
level was determined by western blotting. Downregulation of Bmi-1
expression by siRNA-Bmi-1 transfection significantly upregulated
p16 protein expression (Fig. 4D).
The Transwell chamber migration assay revealed that the motility of
DU145 and PC3 cells was dramatically hampered by the ablation of
Bmi-1 (Fig. 4C). Knockdown of Bmi-1
protein led to the upregulation of E-cadherin and downregulation of
vimentin at the protein levels (Fig.
4D). Moreover, siRNA-Bmi-1-treated PCa cells showed higher
chemosensitivity to 10 nM docetaxel when compared to the NC-treated
cells (Fig. 4B), suggesting that
Bmi-1 silencing enhances docetaxel activity. Previous studies have
shown that Bmi-1 knockdown decreased expression of stem cell-like
markers and chemoresistance markers (19). In the present study, we also found
that the mRNA expression levels of stemness genes CD44 and OCT4
were downregulated in the siRNA-Bmi-1-treated cells when compared
with the control siRNA cells (Fig.
4E). These results suggest that the suppression of Bmi-1
decreases cell proliferation and motility and improves
chemosensitivity to docetaxel.
Bmi-1 is overexpressed in PCa
To investigate differential expression between
prostate cancer cell lines and the normal prostate cells, we
measured the Bmi-1 protein level in 3 different human PCa cell
lines (LNCaP, PC3 and DU145) and the benign hyperplastic epithelial
cell line BPH-1 using western blot assay (Fig. 5A). Immunofluorescence staining also
showed that expression of Bmi-1 was higher in the PCa cell lines
than that in the BPH-1 cells (Fig.
5B). We then examined Bmi-1 expression in 30 cases of BPH and
60 malignant prostate tissue samples. The staining intensity was
generally increased in the malignant cells. The Bmi-1 antibody
resulted in a strong, predominantly nuclear staining in the
malignant epithelia, whereas normal epithelial cells showed
significantly lower staining intensities when compared with the PCa
tissue (P<0.05, χ2 tests) (Fig. 5C). These data showed that Bmi-1
plays a vital role as an oncogene in vitro and in
vivo.
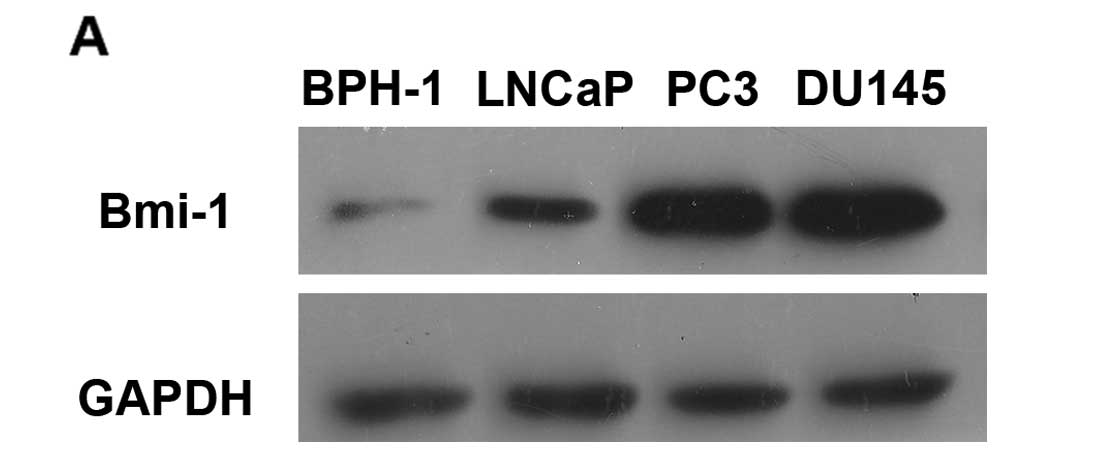 | Figure 5Expression of Bmi-1 in prostate
cancer cell lines and patient tissue samples. (A) The expression of
Bmi-1 protein levels in BPH-1, LNCaP, PC3 and DU145 cell lines.
GAPDH was used as an internal control. (B) Immunofluorescence
staining in human prostate adenocarcinoma cell lines LNCaP, PC3 and
DU145 and human prostate epithelium cell line BPH-1 showed that
Bmi-1 was overexpressed in different degrees in the prostate cancer
cell lines, when compared with the BPH-1 cell line. (C) Different
expression levels of Bmi-1 in normal prostate and prostate cancer
tissues. a, Nonmalignant epithelia showing low staining; b,
malignant cells showing strong staining. c, Scoring of the staining
intensities with the anti-Bmi-1 antibody indicates that Bmi-1
protein expression was significantly increased in PCa when compared
with the BPH tissue specimens (original magnification, ×400).
P<0.05 (χ2 tests). Bmi-1, B-cell-specific Moloney
murine leukemia virus insertion site 1; BPH, benign prostatic
hyperplasia. |
Discussion
miRNAs have recently been described as important
players in human cancer and their role as therapeutic targets has
been proposed. The expression of miRNAs was found to be markedly
deregulated in prostate cancer (PCa), strongly suggesting that
miRNAs are involved in the initiation and progression of this
disease (20). Recent reports
indicate that miR-200b is highly correlated with
epithelial-mesenchymal transition (EMT) and is identified as a
critical regulator of tumor invasion, metastasis and
chemosensitivity (6,21). The miR-200 family was found to
directly target the mRNA of the E-cadherin transcriptional
repressors ZEB1 and ZEB2. Ectopic expression of miR-200 reduced
cancer cell motility by upregulation of E-cadherin protein levels.
Conversely, inhibition of miR-200 reduced E-cadherin expression,
increased expression of vimentin and induced EMT (22). Moreover, recent studies have
discovered that miR-200b and miR-1 inhibit EMT via a Slug-dependent
mechanism and tumorigenesis via a Slug-independent mechanism
(23). In the present study, we
confirmed that miR-200b expression levels were underexpressed in
advanced prostate cancer cell lines and in PCa clinical specimens.
Consistent with our results, miR-200b has frequently been found to
be downregulated in cancers, including gastric (24), lung cancer (25), renal cell carcinoma (26) and bladder cancer (27), as well as PCa (28). The downregulation of miR-200b in PCa
indicates that it may play a vital role in PCa tumor development
and progression as a potential tumor-suppressor.
In PCa cells, miR-200b has an impact on pathways
governing cell proliferation, drug sensitivity and cell migration.
We found that overexpression of miR-200b in PCa cells suppressed
cell growth and cell migration. Moreover, PC3 and DU145 cells
showed a higher sensitivity to 10 nM docetaxel when cells were
treated with miR-200b mimics, when compared to the negative
control. These findings demonstrate that miR-200b may have
significant future clinical implications.
In addition, we further explored the mechanism by
which miR-200b affects proliferation, migration and sensitivity to
docetaxel in PCa cells. We used target prediction databases
(miRanda, Targetscan and PicTar) to search for genes regulated by
miR-200b and found that miR-200b may target Bmi-1. Recent research
has revealed that miR-200b functions as a tumor-suppressor gene and
that Bmi-1 may be the target of miR-200b in human tongue cancer
(29).
Previous studies have suggested that Bmi-1 acts as
an oncogene and plays a crucial role in cell proliferation,
transformation, EMT, self-renewal of stem cells and cancer
initiation and chemoresistance in various human tumors (19,30,31).
We discovered that Bmi-1 was overexpressed in PCa tumor tissues
when compared with that in the BPH normal tissues. Bmi-1
overexpression was also observed in PCa cell lines when compared
with the BPH-1 normal prostate epithelium cells. Consistent with
our findings, previous publications also reported that Bmi-1 was
overexpressed in PCa (32,33), and its overexpression was associated
with a high Gleason score and increased risk of recurrence after
prostatectomy (34). In the present
study, we showed that rescue of miR-200b expression led to
downregulation of Bmi-1 mRNA and protein levels in PCa cells. These
results suggest that miR-200b may directly target the 3′-UTR of
Bmi-1 mRNA and act as a potential tumor-suppressor in PCa.
We demonstrated that knockdown of Bmi-1 in PC3 and
DU145 cells significantly reduced cell proliferation and
upregulated the p16 tumor-suppressor. There is an enormous body of
evidence suggesting that Bmi-1 regulates cell proliferation and
senescence through suppressing p16 (35). Previous studies have discovered that
targeting Bmi-1 improves the outcome of docetaxel therapy in animal
models bearing chemoresistant prostatic tumors (32). Crea et al (36) identified that Bmi-1 silencing
impairs antioxidant defense and sensitizes PCa cells to docetaxel.
The present study also found that inhibition of Bmi-1 enhanced the
antitumor activity of docetaxel in PCa cells. These results
indicate that Bmi-1 may be exploited as a potential molecular
target for therapeutics to treat chemoresistant tumors. Recent
studies have demonstrated that many metastatic cancers undergo EMT
resulting in enhanced cell motility and invasion (37). The hallmark of EMT is the loss of
adherens protein E-cadherin and acquisition of mesenchymal markers
such as vimentin (38). A recent
study showed that Bmi-1 is regulated by Twist1 in HNSCC and is
essential in EMT (39). Yet, the
role of Bmi-1 in PCa cell migration has not been thoroughly
explored. The present study is consistent with the literature
reported. Following inhibition of the expression of Bmi-1, the
migration ability of PCa cells was reduced. Here we also provided
evidence that Bmi-1 is correlated with E-cadherin and vimentin
protein expression in PCa cells.
In summary, our data suggest that Bmi-1 is expressed
at a high level in PCa. miR-200b plays a pivotal role in PCa at
least in part via downregulation of the oncogene Bmi-1 and may
serve as a therapeutic target by which to inhibit PCa cell
proliferation and migration and enhance chemosensitivity.
Acknowledgements
The present study was supported by a grant from the
National Natural Science Foundation of China (no. 81072114).
References
|
1
|
Siegel R, Ward E, Brawley O and Jemal A:
Cancer statistics, 2011: the impact of eliminating socioeconomic
and racial disparities on premature cancer deaths. CA Cancer J
Clin. 61:212–236. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Damber JE and Aus G: Prostate cancer.
Lancet. 371:1710–1721. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bartel DP: MicroRNAs: genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Meister G, Landthaler M, Patkaniowska A,
Dorsett Y, Teng G and Tuschl T: Human Argonaute2 mediates RNA
cleavage targeted by miRNAs and siRNAs. Mol Cell. 15:185–197. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Feng B, Wang R and Chen LB: Review of
miR-200b and cancer chemosensitivity. Biomed Pharmacother.
66:397–402. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhu W, Xu H, Zhu D, et al: miR-200bc/429
cluster modulates multidrug resistance of human cancer cell lines
by targeting BCL2 and XIAP. Cancer Chemother Pharmacol. 69:723–731.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wellner U, Schubert J, Burk UC, et al: The
EMT-activator ZEB1 promotes tumorigenicity by repressing
stemness-inhibiting microRNAs. Nat Cell Biol. 11:1487–1495. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Guo BH, Feng Y, Zhang R, et al: Bmi-1
promotes invasion and metastasis, and its elevated expression is
correlated with an advanced stage of breast cancer. Mol Cancer.
10:102011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bhattacharya R, Nicoloso M, Arvizo R, et
al: MiR-15a and MiR-16 control Bmi-1 expression in ovarian cancer.
Cancer Res. 69:9090–9095. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Song W, Tao K, Li H, et al: Bmi-1 is
related to proliferation, survival and poor prognosis in pancreatic
cancer. Cancer Sci. 101:1754–1760. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
He X, Dong Y, Wu CW, et al: MicroRNA-218
inhibits cell cycle progression and promotes apoptosis in colon
cancer by downregulating BMI1 polycomb ring finger oncogene. Mol
Med. 18:1491–1498. 2013.PubMed/NCBI
|
|
12
|
Song LB, Li J, Liao WT, et al: The
polycomb group protein Bmi-1 represses the tumor-suppressor PTEN
and induces epithelial-mesenchymal transition in human
nasopharyngeal epithelial cells. J Clin Invest. 119:3626–3636.
2009. View
Article : Google Scholar
|
|
13
|
Molofsky AV, Pardal R, Iwashita T, Park
IK, Clarke MF and Morrison SJ: Bmi-1 dependence distinguishes
neural stem cell self-renewal from progenitor proliferation.
Nature. 425:962–967. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lukacs RU, Memarzadeh S, Wu H and Witte
ON: Bmi-1 is a crucial regulator of prostate stem cell self-renewal
and malignant transformation. Cell Stem Cell. 7:682–693. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Onder TT, Kara N, Cherry A, et al:
Chromatin-modifying enzymes as modulators of reprogramming. Nature.
483:598–602. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Korpal M, Lee ES, Hu G and Kang Y: The
miR-200 family inhibits epithelial-mesenchymal transition and
cancer cell migration by direct targeting of E-cadherin
transcriptional repressors ZEB1 and ZEB2. J Biol
Chem. 283:14910–14914. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fan C, He L, Kapoor A, et al: Bmi1
promotes prostate tumorigenesis via inhibiting p16INK4A
and p14ARF expression. Biochim Biophys Acta.
1782:642–648. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dong P, Kaneuchi M, Watari H, et al:
MicroRNA-194 inhibits epithelial to mesenchymal transition of
endometrial cancer cells by targeting oncogene BMI-1. Mol Cancer.
10:992011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Schaefer A, Jung M, Mollenkopf HJ, et al:
Diagnostic and prognostic implications of microRNA profiling in
prostate carcinoma. Int J Cancer. 126:1166–1176. 2010.PubMed/NCBI
|
|
21
|
Kong D, Banerjee S, Ahmad A, et al:
Epithelial to mesenchymal transition is mechanistically linked with
stem cell signatures in prostate cancer cells. PLoS One.
5:e124452010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Park SM, Gaur AB, Lengyel E and Peter ME:
The miR-200 family determines the epithelial phenotype of cancer
cells by targeting the E-cadherin repressors ZEB1 and ZEB2. Genes
Dev. 22:894–907. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu YN, Yin JJ, Abou-Kheir W, et al: MiR-1
and miR-200 inhibit EMT via Slug-dependent and tumorigenesis via
Slug-independent mechanisms. Oncogene. 32:296–306. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tang H, Kong Y, Guo J, et al: Diallyl
disulfide suppresses proliferation and induces apoptosis in human
gastric cancer through Wnt-1 signaling pathway by up-regulation of
miR-200b and miR-22. Cancer Lett. 340:72–81. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pacurari M, Addison JB, Bondalapati N, et
al: The microRNA-200 family targets multiple non-small cell lung
cancer prognostic markers in H1299 cells and BEAS-2B cells. Int J
Oncol. 43:548–560. 2013.PubMed/NCBI
|
|
26
|
Yoshino H, Enokida H, Itesako T, et al:
Epithelial-mesenchymal transition-related microRNA-200s regulate
molecular targets and pathways in renal cell carcinoma. J Hum
Genet. 58:508–516. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Köhler CU, Bryk O, Meier S, et al:
Analyses in human urothelial cells identify methylation of miR-152,
miR-200b and miR-10a genes as candidate bladder cancer biomarkers.
Biochem Biophys Res Commun. 438:48–53. 2013.
|
|
28
|
Cao Q, Mani RS, Ateeq B, et al:
Coordinated regulation of polycomb group complexes through
microRNAs in cancer. Cancer Cell. 20:187–199. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sun L, Yao Y, Liu B, et al: MiR-200b and
miR-15b regulate chemotherapy-induced epithelial-mesenchymal
transition in human tongue cancer cells by targeting BMI1.
Oncogene. 31:432–445. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kang MK, Kim RH, Kim SJ, et al: Elevated
Bmi-1 expression is associated with dysplastic cell transformation
during oral carcinogenesis and is required for cancer cell
replication and survival. Br J Cancer. 96:126–133. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Siddique HR and Saleem M: Role of BMI1, a
stem cell factor, in cancer recurrence and chemoresistance:
preclinical and clinical evidences. Stem Cells. 30:372–378. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Siddique HR, Parray A, Tarapore RS, et al:
BMI1 polycomb group protein acts as a master switch for growth and
death of tumor cells: regulates TCF4-transcriptional factor-induced
BCL2 signaling. PLoS One. 8:e606642013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wolters T, Vissers KJ, Bangma CH, Schröder
FH and van Leenders GJ: The value of EZH2, p27kip1,
BMI-1 and MIB-1 on biopsy specimens with low-risk prostate cancer
in selecting men with significant prostate cancer at prostatectomy.
BJU Int. 106:280–286. 2010.PubMed/NCBI
|
|
34
|
van Leenders GJ, Dukers D, Hessels D, et
al: Polycomb-group oncogenes EZH2, BMI1, and RING1 are
overexpressed in prostate cancer with adverse pathologic and
clinical features. Eur Urol. 52:455–463. 2007.PubMed/NCBI
|
|
35
|
Jacobs JJ, Kieboom K, Marino S, DePinho RA
and van Lohuizen M: The oncogene and Polycomb-group gene
bmi-1 regulates cell proliferation and senescence through
the ink4a locus. Nature. 397:164–168. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Crea F, Duhagon Serrat MA, Hurt EM, Thomas
SB, Danesi R and Farrar WL: BMI1 silencing enhances docetaxel
activity and impairs antioxidant response in prostate cancer. Int J
Cancer. 128:1946–1954. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sánchez-Tilló E, Lázaro A, Torrent R, et
al: ZEB1 represses E-cadherin and induces an EMT by recruiting the
SWI/SNF chromatin-remodeling protein BRG1. Oncogene. 29:3490–3500.
2010.PubMed/NCBI
|
|
39
|
Yang MH, Hsu DS, Wang HW, et al: Bmi1 is
essential in Twist1-induced epithelial-mesenchymal transition. Nat
Cell Biol. 12:982–992. 2010. View
Article : Google Scholar : PubMed/NCBI
|