Introduction
Osteosarcoma is the most common primary bone
malignant tumor, affecting primarily children and young adolescents
(1). Despite the advances in
multimodality treatments, the combination of neoadjuvant
chemotherapy plus surgery have increased the survival rates
markedly from 20 to 65% since the 1980s, but the progress has been
considerably slow over the past 20 years (2,3).
Therefore, an understanding of the molecular mechanisms of
osteosarcoma is one of the most important issues for treatment. New
therapeutic strategies are necessary to increase the survival rates
in patients with osteosarcoma (4).
In recent years, there has been a global trend
toward the importance of naturally occurring phytochemicals in
plants for the prevention and treatment of human diseases. Several
of these phytochemicals have shown potential as cancer
chemopreventive or therapeutic agents in the human body (5). Most of these bioactive phytochemicals
exert their cancer chemotherapeutic activity by blocking cell cycle
progression and triggering apoptotic cell death (5). Therefore, the induction of apoptosis
in cancer cells is an important indicator of the cancer treatment
response when employing a bioactive substance to reduce and control
human mortality due to cancer (6,7).
Apoptosis, which is a major form of programmed cell
death, plays an important role in regulating tissue development and
homeostasis in eukaryotes (8).
Apoptosis can occur via a death receptor-dependent extrinsic or a
mitochondria-dependent intrinsic pathway, and apoptosis is induced
by a treatment with chemotherapeutic agents (9,10).
Diferuloylmethane (curcumin), which is a constituent
of turmeric powder derived from the rhizome of Curcuma
longa, has anti-inflammatory, antimicrobial, anti-oxidative,
immunomodulating and anti-atherogenic properties (11,12).
Furthermore, many studies have reported that curcumin has
chemopreventive and antiproliferative activity in many human
cancers (12,13). Of particular interest is that
curcumin is also pharmacologically safe as it is a naturally
occurring compound (14). However,
natural curcumin has limited use due to its poor intestinal
absorption and low bioavailability (15). On the other hand, it is an excellent
compound for the design of more effective analogs. One monoketone
analog in particular, diphenyldifluoroketone (EF-24), is effective
in anticancer screens (16). EF-24
was reported to inhibit the growth of human breast tumor xenografts
in a mouse model with relatively low toxicity and at a much lower
dose than that of curcumin (16).
Although it was shown to reduce the cancer cell viability, its
action mechanisms remain to be determined. In addition, the effects
of EF-24 and curcumin on osteosarcoma are unclear.
The present study examined the effects of EF-24 and
curcumin on cell growth and the mechanism of cell death elicited by
EF-24 and curcumin in Saos2 human osteogenic sarcoma cells. The
study reports for the first time that the induction of apoptotic
cell death by EF-24 is associated with the activation of caspases
in osteogenic sarcoma cells.
Materials and methods
Materials
Saos2 human osteogenic sarcoma cells were provided
by the American Type Culture Collection (ATCC, Rockville, MD, USA).
EF-24 and curcumin were supplied by Sigma (St. Louis, MO, USA).
Anti-cleaved caspase-3/-7/-8/-9, poly(ADP-ribose) polymerase
(PARP), Fas, Bid, Bax, Bcl-2, p53 and β-actin antibodies were
purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA).
The cell-permeable fluorogenic substrate
PhiPhiLux-G1D2 was purchased from
OncoImmunin, Inc. (Gaithersburg, MD, USA). All other analytical
reagents purchased were of analytical grade.
Cell cultures
The Saos2 human osteogenic sarcoma cells were grown
in DMEM supplemented with 10% fetal bovine serum (FBS) (17). The cells were maintained as
monolayers in plastic culture plates at 37°C in a humidified
atmosphere containing 5% CO2.
MTT assay
The Saos2 cells were seeded at a concentration of
5×103 cells/well in 24-well plates. After 24-h growth,
the cells were treated with EF-24 or curcumin at various
concentrations for 24 h. The cell viability was assessed using an
MTT assay.
Nuclear staining with DAPI
Nuclear staining with DAPI
(40,60-diamidino-2-phenylindole) was performed to examine the level
of apoptosis. The Saos2 cells were cultured in 24-well plates at a
seeding density of 5×103 cells/well. After 24-h growth,
the cells were treated with 10 μM EF-24 or 30 μM curcumin for 24 h.
The treated Saos2 cells were fixed with 1% paraformaldehyde for 30
min at room temperature and washed twice with PBS. The cells were
permeated with ice-cold ethanol for 5 min at room temperature and
washed twice with PBS. The fixed cells were stained with DAPI (300
nM) for 5 min at room temperature in the dark, washed twice with
PBS and examined by fluorescent inverted microscopy (IX71; Olympus,
Tokyo, Japan).
Determination of caspase activation
The activity of caspase-3/-7 was determined using
the cell-permeable fluorogenic substrate,
PhiPhiLux-G1D2, according to the
manufacturer’s instructions. The cells were treated with 0 and 10
μM EF-24 or 0 and 30 μM curcumin for 24 h and incubated with
PhiPhiLux-G1D2. The activity of caspase-3/-7
was examined by fluorescence microscopy (IX71; Olympus).
Annexin V-fluorescein isothiocyanate
(V-FITC), propidium iodide (PI) and flow cytometric analysis
The Saos2 cells were cultured in 100-mm
tissue-culture dishes at a density of 1×106 cells/dish
for 24 h and were treated with 10 μM EF-24 or 30 μM curcumin for 24
h. The cells were washed twice in phosphate-buffered saline and
resuspended in the binding buffer (BD Biosciences, San Diego, CA,
USA). Annexin V-FITC and 7-amino-actinomycin D (BD Biosciences)
were added to the cells, which were then incubated in the dark for
15 min, and resuspended in 400 μl of binding buffer. The cells were
analyzed using a fluorescence activated cell sorting Calibur flow
cytometer (Becton-Dickinson, San Jose, CA, USA). Data analysis was
performed using standard CellQuest software (Becton-Dickinson).
Immunoblotting
The Saos2 cells were treated with EF-24 or curcumin
for 24 h. Immunoblotting was performed using a minor modification
of the method described elsewhere (18). Anti-cleaved caspase-3/-7/-8/-9,
PARP, Fas, Bid, tBid, Bax, Bcl-2 and p53 antibodies (Cell Signaling
Technology) were used as the primary antibodies.
Data analysis
All experiments were performed at least in
triplicate. The results are presented as the mean ± SEM. The
statistical significance was analyzed using a Student’s t-test for
the two group comparison and one-way analysis of the variance for
the multi-group comparisons. A P-value <0.05 was considered to
indicate a statistically significant difference.
Results
Cytotoxic effects of EF-24 and curcumin
on Saos2 cells
The Saos2 cells were treated with EF-24 and curcumin
at various concentrations for 24 h, and were analyzed by MTT assay.
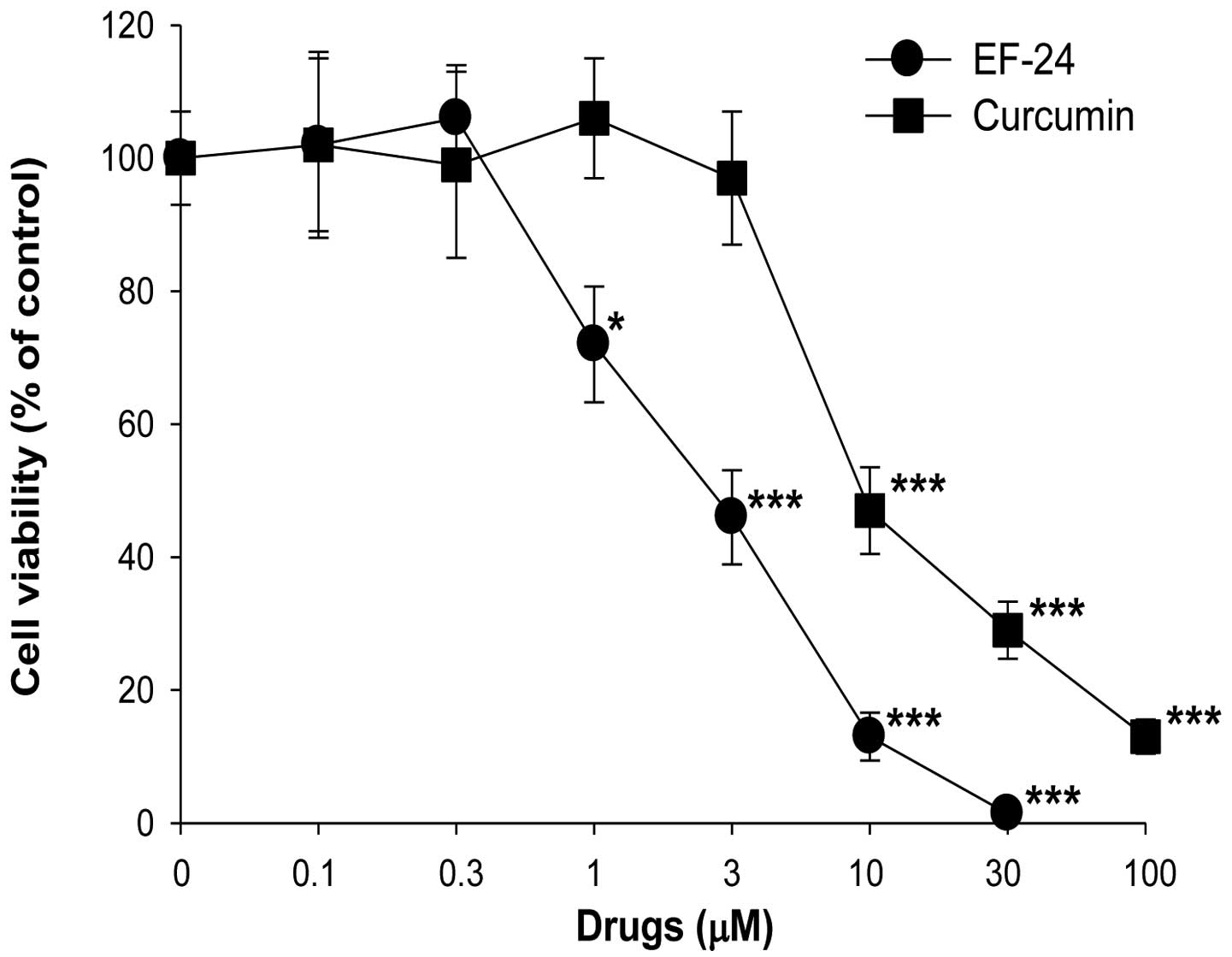
As shown in Fig. 1, treatment of
curcumin at doses from 0.1 to 3 μM did not significantly affect the
cell viability of the Saos2 cells, but curcumin at 10, 30 and 100
μM reduced Saos2 cell viability. When the cells were treated with
EF-24 for 24 h, EF-24 inhibited the growth of cells in a
dose-dependent manner, suggesting that EF-24 induces Saos2 cell
death (Fig. 1). The IC50
values of EF-24 and curcumin on Saos2 cell viability after a 24-h
treatment were 2.7±0.3 and 9.7±1.4 μM, respectively. The apparent
potency of EF-24 was >3-fold higher that of curcumin. More
importantly, the effects were observed at an EF-24 concentration
<3 μM, a dose at which curcumin had no significant effects on
cell proliferation, indicating the higher potency of EF-24.
Changes in the nuclear morphology by
EF-24 and curcumin
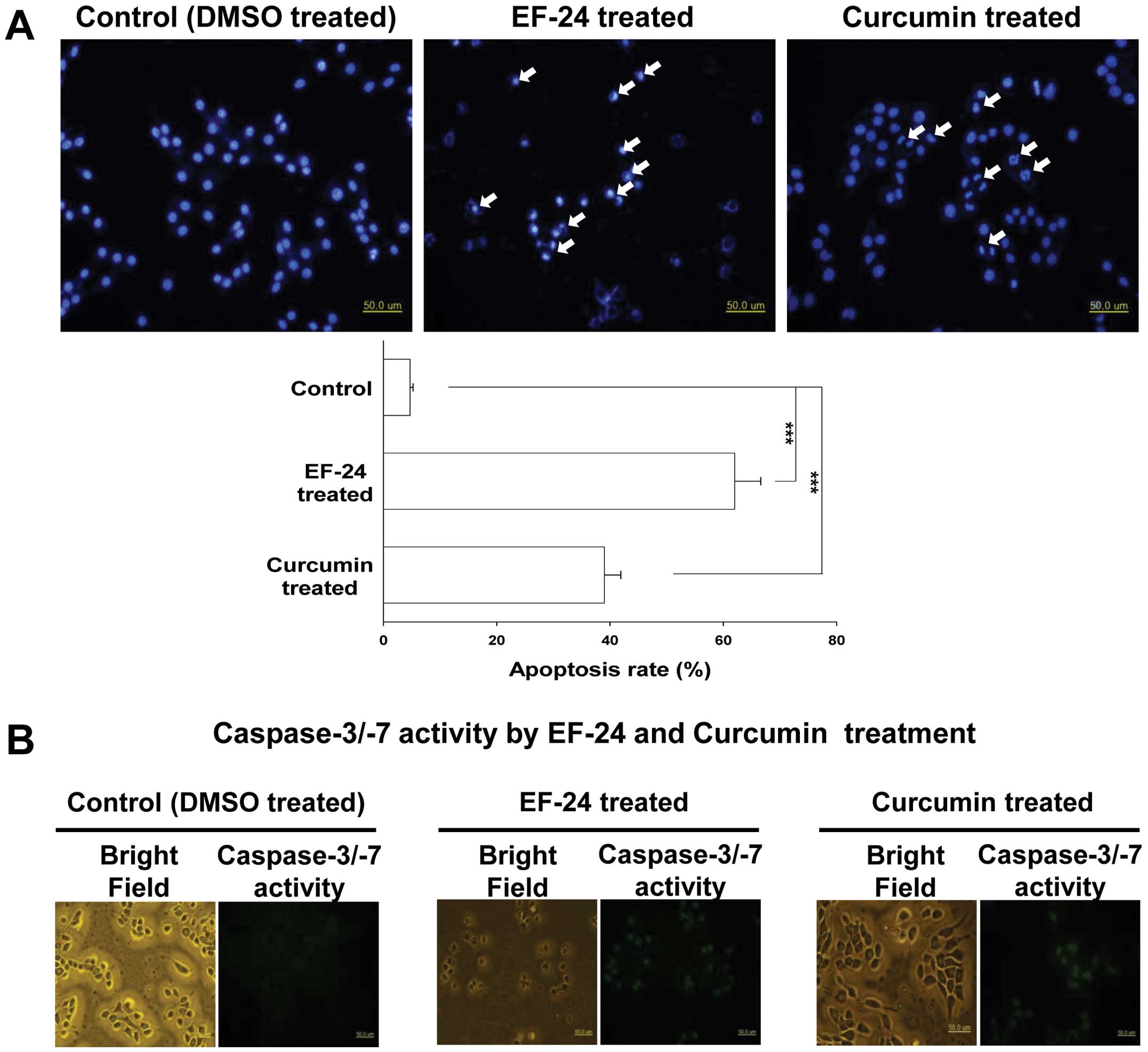
The nuclear morphological changes were assessed by
DAPI staining. As shown in Fig. 2A,
the nuclei of the control Saos2 cells (control) had a normal
regular and oval shape. Treatment with 10 μM EF-24 or 30 μM
curcumin for 24 h resulted in nuclear condensation and
fragmentation, which are the characteristics of apoptosis (Fig. 2A, upper panel). As quantified in
Fig. 2A (lower panel), EF-24 and
curcumin increased the apoptotic rate of Saos2 cells significantly
to 62.0±4.6 and 39.1±2.9%, respectively.
Activation of caspase-3/-7 by EF-24 and
curcumin
The activation of caspase-3/-7 in the EF-24- or
curcumin-treated Saos2 cells was confirmed by fluorescence
microscopy using a fluorogenic substrate. As shown in Fig. 2B, both the EF-24 and curcumin
treatment led to the activation of caspase-3/-7 in the living Saos2
cells.
Apoptosis induction of Saos2 cells by
EF-24 and curcumin
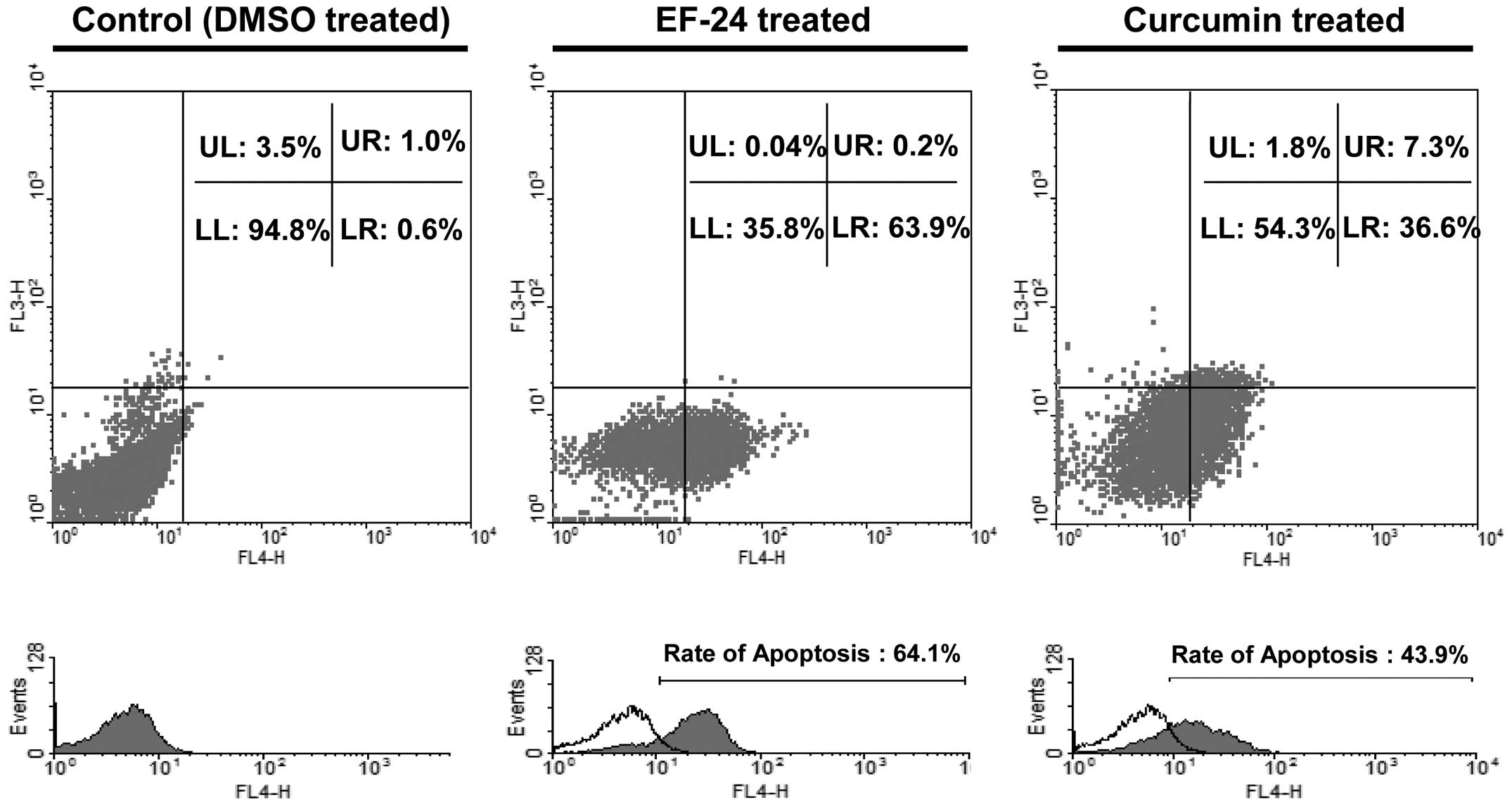
To determine if EF-24- or curcumin-induced cell
death is associated with the induction of apoptosis, the Saos2
cells were stimulated with 10 μM EF-24 or 30 μM curcumin for 24 h
and then co-stained with Annexin V-FITC, an apoptotic marker, and
PI, a necrotic marker. The percentage of Annexin V-FITC-positive
cells at the stage of apoptosis was increased to 64.1 and 43.9% by
EF-24 and curcumin, respectively, compared to the control (Fig. 3).
Activation of caspases by EF-24 and
curcumin
The levels of cleaved caspase-3/-7/-9 were examined
by immunoblotting as caspase-3/-7/-9 are effector caspases of
apoptotic cell death (19).
Treatment with 1 and 10 μM EF-24 or 1, 10 and 30 μM curcumin for 24
h promoted the proteolytic cleavage of procaspase-3/-7/-9 in Saos2
cells, with increases in the amount of cleaved caspase-3/-7/-9
(Fig. 4A).
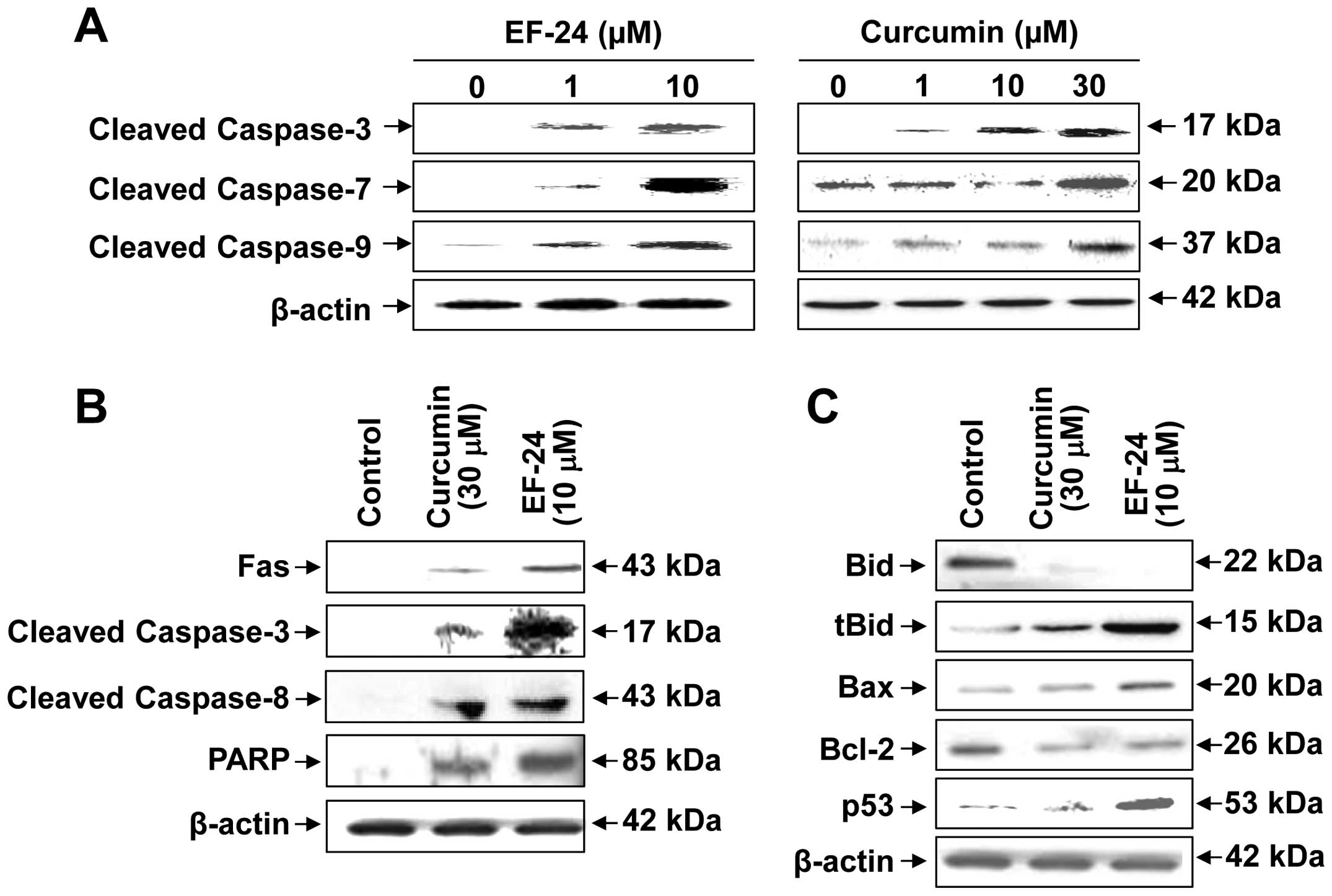 | Figure 4EF-24 or curcumin-induced apoptosis
mediated by expression and activation of caspases in Saos2 cells.
(A) Proteolytic cleavage of caspase-3/-7/-9 by EF-24 or curcumin
treatment. The activity of cleaved caspase-3/-7/-9 by EF-24 or
curcumin was measured in Saos2 cells. The cells were treated with
0, 1 and 10 μM EF-24 or 0, 1, 10 and 30 μM curcumin for 24 h. The
cell lysate was prepared and analyzed by immunoblotting as
described in Materials and methods. (B) Activation of the extrinsic
apoptosis signaling pathway by EF-24 and curcumin. The activity of
extrinsic apoptosis signaling pathway in Saos2 cells by EF-24 or
curcumin was measured. The Saos2 cells were stimulated with 0 and
10 μM EF-24 or 0 and 30 μM curcumin for 24 h, harvested, and lysed
using a cell lysis buffer. (C) Activation of p53 and regulation of
Bid, tBid, Bax and Bcl-2 levels by EF-24 and curcumin. The
activities of p53, Bid, tBid, Bax and Bcl-2 by EF-24 or curcumin
were measured in Saos2 cells. |
Apoptosis mediated via Fas/PARP axis by
EF-24 and curcumin
To determine how EF-24 or curcumin induce the
apoptosis of Saos2 cells, immunoblotting was performed to measure
the expression of the apoptotic genes at the protein level. As
shown in Fig. 4B, Fas, which is an
apoptotic ligand that triggers the extrinsic apoptotic pathway in
Saos2 cells (20,21), was induced significantly by either
EF-24 or curcumin. Subsequently, the cleaved caspase-3 and -8,
which are the downstream targets of Fas, were induced by EF-24 or
curcumin. Cleaved PARP was increased by EF-24 or curcumin compared
to the control.
Apoptosis-related signal pathways by
EF-24 and curcumin
The level of several proteins that are highly
relevant to understanding the apoptotic signaling pathways in Saos2
cells by EF-24 or curcumin was measured by immunoblot analysis. The
level of Bid and Bcl-2 protein expressions in Saos2 cells
stimulated with 10 μM EF-24 or 30 μM curcumin for 24 h decreased
(Fig. 4C). On the other hand, the
treatment of Saos2 cells with EF-24 or curcumin increased the level
of truncated-Bid (tBid), Bax and p53 protein expression (Fig. 4C).
Discussion
Recent studies have shown that the chemicals derived
from natural materials can elicit chemopreventive and therapeutic
effects (5,22). This effect was reported to alter
many factors associated with the cell cycle and to induce apoptotic
cell death (5,23). Finding new anticancer agents that
can kill cancerous cells with minimal toxicity is critical.
Curcumin has been extracted from the dried ground rhizome of the
perennial herb, Curcuma longa. Several studies have
suggested that curcumin induces cell cycle arrest and apoptosis in
a variety of cancer cells (12,13).
On the other hand, the design of more effective analogs is needed
due to the poor intestinal absorption and low bioavailability of
curcumin (15). EF-24, a monoketone
analog of curcumin, is efficacious in anticancer screens and was
reported to inhibit the growth of human breast tumor xenografts in
a mouse model (16). Although EF-24
can reduce cancer cell viability, the mechanisms of action are
unclear, and the effects of EF-24 and curcumin on osteosarcoma have
not been established. Osteosarcoma is the most common type of
malignant bone tumor, in which the neoplastic mesenchymal cells
show evidence of osteoid production (18). The present study examined the
cytotoxic effects of EF-24, and the mechanism of cell death
exhibited by EF-24 in Saos2 human osteogenic sarcoma cells was
assessed. The results suggest that EF-24, a novel curcumin analog,
possesses considerable promise as an anti-osteosarcoma
therapeutic.
An MTT assay showed that the treatment of curcumin
from 0.1 to 3 μM did not significantly affect the cell viability of
Saos2 cells, but curcumin at 10, 30 and 100 μM reduced Saos2 cell
viability (Fig. 1). EF-24 inhibited
the growth of Saos2 cells in a concentration-dependent manner
(Fig. 1). This corresponded to the
results of EF-24 and curcumin, which have anticancer effects via
the suppression of cancer cell growth in many types of cancer cells
(12,13). In addition, the apparent potency of
EF-24 was >3-fold higher that of curcumin. This suggests that
EF-24 and curcumin are cytotoxic to osteosarcoma cells with EF-24
having higher potency. In addition, these results suggest that it
has potential value for anticancer drug discovery.
Apoptosis is an important way of maintaining
cellular homeostasis between cell division and cell death (8). The induction of apoptosis in cancer
cells is a useful strategy for anticancer drug development
(24). Therefore, many studies have
screened plant-derived compounds for their effects on apoptosis
(5). In the present study,
treatment with EF-24 and curcumin induced nuclear condensation and
fragmentation and the activation of the caspase-3/-7 in living
Saos2 cells, suggesting apoptotic cell death (Fig. 2). These results indicate that EF-24
and curcumin inhibit the growth of Saos2 cells by activating cell
apoptosis.
Curcumin and EF-24 were evaluated for their
apoptosis-inducing activities by the flow cytometry of Saos2 cells
stained with Annexin V-FITC and PI. Exposure of the membrane
phospholipid, phosphatidylserine, to the external cellular
environment is one of the earliest markers of apoptotic cell death
(25,26). Annexin V is a calcium-dependent
phospholipid binding protein with high affinity to
phosphatidylserine expressed on the cell surface (25). PI does not enter a cell with an
intact cell-membrane and has been used to differentiate among early
apoptotic (Annexin V-positive, PI-negative), late apoptotic
(Annexin V-, PI-double positive) and necrotic cell death (Annexin
V-negative, PI-positive) (25). In
the present study, the ratio of early apoptotic Saos2 cells (lower
right) was increased after a treatment with 10 μM EF-24 (from 0.6
to 63.9%) or 30 μM curcumin (from 0.6 to 36.6%) (Fig. 3). These results showed that most of
the cytotoxic activity of curcumin and EF-24 against Saos2 cells is
due to the induction of apoptotic cell death.
The activation of a family of intracellular cysteine
proteases, known as caspases, plays an important role in the
initiation and execution of apoptosis induced by a range of stimuli
(27). Among the caspases
identified in mammalian cells, caspase-3/-7/-8/-9 can serve as the
effector caspases of apoptotic cell death (27). Caspase-3/-7/-8/-9 are synthesized as
inactive proenzymes, which require proteolytic activation to
cleaved enzymes (of sizes 17, 20, 43 and 37 kDa, respectively)
(27). The results of the present
study showed that the amount of cleaved caspase-3/-7/-8/-9 in Saos2
cells was increased after the EF-24 or curcumin treatment (Fig. 4A and B). These results suggest that
EF-24 and curcumin induce apoptotic cell death through both the
mitochondria-mediated intrinsic pathway and death receptor-mediated
extrinsic pathway by the activation of caspases-3/-7/-8/-9 in Saos2
cells.
The factor associated suicide ligand (Fas), which is
an important regulatory factor of apoptosis, initiates the death
receptor-mediated extrinsic apoptotic pathway through the
activation of caspase-8, -3 and PARP, sequentially, after binding
with the receptor FasR spanned on the surface of the target cells
(20,21). In the present study, the expression
of Fas was upregulated significantly by EF-24 or curcumin in Saos2
cells (Fig. 4B). Subsequently, Fas
upregulated by EF-24 or curcumin triggers a caspase cascade, which
then results in the activation of apoptotic factors, including
cleaved caspase-8 and -3. Finally, activated caspase-3 cleaved its
major substrate, PARP, leading to consequent apoptosis. Therefore,
these results suggest that EF-24- or curcumin-induced apoptosis in
Saos2 cells is mediated by the death receptor-mediated extrinsic
apoptotic pathway via the Fas/PARP axis.
Next, we assessed the effects of EF-24 and curcumin
on the expression of p53, Bid, tBid, Bax and Bcl-2 in Saos2 cells.
The molecular biological pathways underlying the inhibition of
cancerous growth typically involve tumor suppressors, such as p53
(8,28–30).
The level of p53 is consistent in normal cells and the protein
becomes phosphorylated during cellular stress, which then interacts
with mouse double minute 2 (MDM2), resulting in apoptosis (8,28–31).
In the present study, the level of p53 was higher in the Saos2
cells stimulated with EF-24 or curcumin than in the control
(Fig. 4C), indicating that p53 may
mediate the EF-24- or curcumin-induced apoptosis of Saos2 cells.
The pro-apoptotic proteins, Bid, tBid and Bax, and the
anti-apoptotic mitochondrial protein, Bcl-2, are important
regulators of cytochrome c release from the mitochondria
(32,33). The Bcl-2 family is localized to the
mitochondrial membrane and modulates apoptosis by permeabilizing
the mitochondrial membrane, leading to the release of cytochrome
c (34). In the present
study, treatment of Saos2 cells with EF-24 or curcumin decreased
the level of Bid and increased the level of tBid (Fig. 4C). Caspase-8 was reported to
truncate Bid, and tBid could activate the mitochondrial pathway
(35). In addition, treatment of
Saos2 cells with EF-24 or curcumin decreased the level of Bcl-2 but
increased the level of Bax (Fig.
4C). The Bax/Bcl-2 ratio is one of the indices of the intrinsic
mechanism of apoptosis in the mitochondria (36). EF-24 or curcumin-induced apoptosis
appears to involve Bax/Bcl-2 signal transduction since EF-24 and
curcumin increased this ratio in Saos2 cells. Therefore, EF-24 and
curcumin are suggested to induce apoptosis in Saos2 cells involving
both the mitochondrial- and death receptor-signal transduction
pathways. On the other hand, the mechanisms of apoptosis induced by
EF-24 and curcumin in Saos2 cells are not fully understood. Further
studies are required to examine the precise cellular and molecular
mechanisms of apoptosis induced by EF-24 and curcumin.
In conclusion, these in vitro results suggest
that EF-24 and curcumin inhibit cell proliferation and induce
apoptotic cell death in Saos2 human osteogenic sarcoma cells
through both the mitochondria-mediated intrinsic pathway and the
death receptor-mediated extrinsic pathway. Moreover, EF-24 was
>3-fold more potent than curcumin. Overall, EF-24 can be a model
compound for the further development of natural product-derived
anti-osteosarcoma agents. On the other hand, to elaborate on this
nascent possibility, a further study of its activity including
in vivo and the purification of bioactive compounds is
currently being conducted.
Acknowledgements
The present study was supported by a research fund
from Chosun University, 2013.
References
|
1
|
Daiy H, Huangy Y, Li Y, Meng G, Wang Y and
Guo QN: TSSC3 overexpression associates with growth inhibition,
apoptosis induction and enhances chemotherapeutic effects in human
osteosarcoma. Carcinogenesis. 33:30–40. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fuchs B and Pritchard DJ: Etiology of
osteosarcoma. Clin Orthop Relat Res. 397:40–52. 2002. View Article : Google Scholar
|
|
3
|
Gibbs CP Jr, Weber K and Scarborough MT:
Malignant bone tumors. Instr Course Lect. 51:413–428. 2002.
|
|
4
|
Zhao Q, Wang C, Zhu J, Wang L, Dong S,
Zhang G and Tian J: RNAi-mediated knockdown of cyclooxygenase2
inhibits the growth, invasion and migration of Saos2 human
osteosarcoma cells: a case control study. J Exp Clin Cancer Res.
30:262011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mukherjee AK, Basu S, Sarkar N and Ghosh
AC: Advances in cancer therapy with plant based natural products.
Curr Med Chem. 8:1467–1486. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Smets LA: Programmed cell death
(apoptosis) and response to anti-cancer drugs. Anticancer Drugs.
5:3–9. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Paschka AG, Butler R and Young CYF:
Induction of apoptosis in prostate cancer cell lines by the green
tea component, (−)-epigallocatechin-3-gallate. Cancer Lett.
130:1–7. 1998.
|
|
8
|
Kaufmann SH and Hengartner MO: Programmed
cell death: alive and well in the new millennium. Trends Cell Biol.
11:526–534. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kaufmann SH and Earnshaw WC: Induction of
apoptosis by cancer chemotherapy. Exp Cell Res. 256:42–49. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Reed JC: Apoptosis-regulating proteins as
targets for drug discovery. Trends Mol Med. 7:314–319. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Banerjee M, Tripathi LM, Srivastava VM,
Puri A and Shukla R: Modulation of inflammatory mediators by profen
and curcumin treatment during chronic inflammation in rat.
Immunopharmacol Immunotoxicol. 25:213–224. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sahu RP, Batra1 S and Srivastava SK:
Activation of ATM/Chk1 by curcumin causes cell cycle arrest and
apoptosis in human pancreatic cancer cells. Br J Cancer.
100:1425–1433. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wahl H, Tan L, Griffith K, Choi M and Liu
JR: Curcumin enhances Apo2L/TRAIL-induced apoptosis in
chemoresistant ovarian cancer cells. Gynecol Oncol. 105:104–112.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Goel A, Kunnumakkara AB and Aggarwal BB:
Curcumin as ‘Curecumin’: from kitchen to clinic. Biochem Pharmacol.
75:787–809. 2008.
|
|
15
|
Anand P, Kunnumakkara AB, Newman RA and
Aggarwal BB: Bioavailability of curcumin: problems and promises.
Mol Pharm. 4:807–818. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Adams BK, Cai J, Armstrong J, et al: EF24,
a novel synthetic curcumin analog, induces apoptosis in cancer
cells via a redox-dependent mechanism. Anticancer Drugs.
16:263–275. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Komatsu M, Kuroda M, Wang Y, et al:
Manganese superoxide dismutase overexpression changes plating
efficiency bidirectionally according to change in redox for SaOS2
human osteosarcoma cell line. Int J Oncol. 26:853–862. 2005.
|
|
18
|
Kim SG, Kim HH, Kim HK, et al:
Differential expression and functional characterization of system L
amino acid transporters in human normal osteoblast cells and
osteogenic sarcoma cells. Anticancer Res. 26:1989–1996. 2006.
|
|
19
|
Hu W and Kavanagh JJ: Anticancer therapy
targeting the apoptotic pathway. Lancet Oncol. 4:721–729. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Herrnring C, Reimer T, Jeschke U, et al:
Expression of the apoptosis-inducing ligands FasL and TRAIL in
malignant and benign human breast tumors. Histochem Cell Biol.
113:189–194. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li HJ, Wang CY, Mi Y, et al: FasL-induced
apoptosis in bovine oocytes via the Bax signal. Theriogenology.
80:248–255. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cheng YL, Lee SC, Lin SZ, et al:
Anti-proliferative activity of Bupleurum scrozonerifolium in
A549 human lung cancer cells in vitro and in vivo. Cancer Lett.
222:183–193. 2005.PubMed/NCBI
|
|
23
|
Tian Z, Chen S, Zhang Y, et al: The
cytotoxicity of naturally occurring styryl lactones. Phytomedicine.
13:181–186. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hauser PJ, Han Z, Sindhwani P and Hurst
RE: Sensitivity of bladder cancer cells to curcumin and its
derivatives depends on the extracellular matrix. Anticancer Res.
27:737–740. 2007.PubMed/NCBI
|
|
25
|
Kikuchi T, Pan X, Ishii K, et al:
Cytotoxic and apoptosis-inducing activities of
12-O-Acetylazedarachin B from the fruits of Melia
azedarach in human cancer cell lines. Biol Pharm Bull.
36:135–139. 2013.PubMed/NCBI
|
|
26
|
Martin SJ, Reutelingsperger CP, McGahon
AJ, Rader JA, van Schie RCAA, LaFace DM and Green DR: Early
redistribution of plasma membrane phosphatidylserine is a general
feature of apoptosis regardless of the initiating stimulus:
inhibition by overexpression of Bcl-2 and Abl. J Exp Med.
182:1545–1556. 1995. View Article : Google Scholar
|
|
27
|
Liu X, Zou H, Slaughter C and Wang X: DFF,
a heterodimeric protein that functions downstream of caspase-3 to
trigger DNA fragmentation during apoptosis. Cell. 89:175–184. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ryu DS, Lee HS, Lee GS and Lee DS: Effects
of the ethylacetate extract of Orostachys japonicus on
induction of apoptosis through the p53-mediated signaling pathway
in human gastric cancer cells. Biol Pharm Bull. 35:660–665.
2012.PubMed/NCBI
|
|
29
|
Lavin MF and Gueven N: The complexity of
p53 stabilization and activation. Cell Death Differ. 13:941–950.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Heath-Engel HM, Chang NC and Shore GC: The
endoplasmic reticulum in apoptosis and autophagy: role of the BCL-2
protein family. Oncogene. 27:6419–6433. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Fisher DE: The p53 tumor suppressor:
critical regulator of life & death in cancer. Apoptosis.
6:7–15. 2001. View Article : Google Scholar
|
|
32
|
Kluck RM, Bossy-Wetzel E, Green DR and
Newmeyer DD: The release of cytochrome c from mitochondria: a
primary site for Bcl-2 regulation of apoptosis. Science.
275:1132–1136. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kluck RM, Esposti MD, Perkins G, et al:
The pro-apoptotic proteins, Bid and Bax, cause a limited
permeabilization of the mitochondrial outer membrane that is
enhanced by cytosol. J Cell Biol. 147:809–822. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yu CS, Huang AC, Lai KC, Huang YP, Lin MW,
Yang JS and Chung JG: Diallyl trisulfide induces apoptosis in human
primary colorectal cancer cells. Oncol Rep. 28:949–954.
2012.PubMed/NCBI
|
|
35
|
Luo X, Budihardjo I, Zou H, Slaughter C
and Wang X: Bid, a Bcl2 interacting protein, mediates cytochrome c
release from mitochondria in response to activation of cell surface
death receptors. Cell. 94:481–490. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Oltvai ZN, Milliman CL and Korsmeyer SJ:
Bcl-2 heterodimerizes in vivo with a conserved homolog, Bax, that
accelerates programmed cell death. Cell. 74:609–619. 1993.
View Article : Google Scholar : PubMed/NCBI
|