Introduction
We previously reported that vitamin B6
supplemented diet markedly reduces colon tumorigenesis in mice
exposed to azoxymethane (AOM) (1,2).
Consistently, increasing epidemiological evidence indicates vitamin
B6 acts as a protective factor against colon cancer
(3–6). We also reported that vitamin
B6 decreases oxidative stress, inflammation, cell
proliferation, epithelial cell damage, and angiogenesis, which may
lead to lower tumorigenesis (1,2,7–10).
Moreover, we recently found that high concentrations of pyridoxal
(PL) increase the expression of insulin-like growth factor-binding
protein 1 (IGFBP1), a putative tumor suppressor, in human hepatoma
(HepG2) cells via the upregulation of the ERK/c-Jun pathway
(11). However, the molecular
mechanisms involved in the antitumor effect of vitamin
B6 remain unclear.
Our preliminary experiment involving DNA microarray
analysis shows several genes are upregulated by 500 μM PL in human
colon carcinoma (HT29) cells. Among these upregulated genes, higher
p21 expression was confirmed by real-time PCR. It is well known
that p21 negatively regulates cell cycle progression and is an
antitumor factor (12). The gene
expression of p21 is tightly controlled by the tumor-suppressor
protein p53 (13). A recent study
shows that a high dose of pyridoxine (PN, 10 mM) induces
insulin-like growth factor-binding protein 3 (IGFBP3) mRNA
expression in MCF-7 cells in a p53-dependent manner (14). Therefore, the present study examined
the effect of vitamin B6 on p21 gene expression and p53
activation in cancer cells and the colon of mice.
Materials and methods
Materials
PL hydrochloride, PN hydrochloride, and pyridoxal
5′-phosphate (PLP) were obtained from Nacalai Tesque (Kyoto,
Japan), and pyridoxamine (PM) dihydrochloride was obtained from
Calbiochem (La Jolla, CA, USA). Human colorectal cancer (HT29)
cells, human epithelial colorectal adenocarcinoma (Caco2) cells,
human colon adenocarcinoma (LoVo) cells, human embryonic kidney
(HEK293T) cells, and human hepatoma (HepG2) cells were purchased
from the Health Science Research Resources Bank (Japan) and the
Japan Health Science Foundation (Japan). Dulbecco’s modified
Eagle’s medium (DMEM) was purchased from Sigma (St. Louis, MO,
USA). Anti-p-p53 antibody was obtained from Cell Signaling
Technology (USA). Anti-p53 antibody was obtained from Santa Cruz
Biotechnology (Santa Cruz, CA, USA). Anti-tubulin antibody was
obtained from Harlan Sera-Lab (UK).
Cell cultures and treatment
HT29, Caco2, LoVo, HEK293T, and HepG2 cells were
maintained in a DMEM supplemented with 10% fetal calf serum, 100
U/ml penicillin and 100 μg/ml streptomycin at 37°C in 5%
CO2. PL, PN, PM, or PLP was dissolved directly into the
culture medium and filtered through a Millex-HV (0.45 μm;
Millipore, Billerica, MA, USA).
Animals and diets
Four-week-old male ICR mice (Charles River, Japan)
were housed in groups of 3 in metal cages in a room with controlled
temperature (24±1°C) and a 12:12-h light/dark cycle (lights on from
800–2000 h) according to the Guide for the Care and Use of
Laboratory Animals established by Hiroshima University Animal
Research Committee. After one week of acclimation on commercial
stock diet (MF, Oriental Yeast, Tokyo, Japan), the mice were
divided into two groups (n=9 each) fed diets with different vitamin
B6 concentrations. The basal diet consisted of the
following components (g/kg diet): α-cornstarch, 302; casein, 200;
sucrose, 200; corn oil, 200; cellulose, 50; AIN-93G mineral
mixture, 35; AIN-93 vitamin mixture (PN-free), 10; and l-cysteine,
3. PN HCl (Nacalai Tesque) was added to the basal diet at 0 or 7
mg/kg diet (15). The level of PN
HCl/kg diet recommended in the AIN-93 diet is 7 mg (16). Animals had free access to food and
water ad libitum for 5 weeks.
mRNA analysis
Total RNA from HT29, Caco2, LoVo, HEK293T and HepG2
cells was isolated using TRIzol™ (Invitrogen, Carlsbad, CA, USA).
The Qiagen Midi kit was used to isolate total RNA from mouse
colons, which were subsequently prepared according to the standard
protocol. Total RNA (1 μg) was reverse-transcribed using the First
Strand cDNA Synthesis kit (Toyobo, Japan) according to the
manufacturer’s instructions. Real-time PCR was performed with a
StepOne™ Real-Time PCR System (Applied Biosystems, Japan) using
Thunderbird SYBR qPCR Mix (Toyobo, Japan). The human primer sets
for p21, p53, and GAPDH were purchased from Greiner Bio-One (Japan)
(p21: 5′-TGGAGACTCTCAGGGTCGAAA-3′ and 5′-CGGCGTTTGGAGTGGTAGA-3′;
p53: 5′-ATCTACTGGGACGGAACAGC-3′ and 5′-GTGAGGCTCCCCTTTCTTG-3′;
GAPDH: 5′-CAATGACCCCTTCATTGACC-3′ and 5′-TGGAAGATGGTGATGGGATT-3′).
The mouse primer sets for p21 and GAPDH were also purchased from
Greiner Bio-One (p21, 5′-AGTGTGCCGTTGTCTCTTCG-3′ and
5′-ACACCAGAGTGCAAGACAGC-3′; GAPDH: 5′-CATGGCCTTCCGTGTTCCTA-3′ and
5′-CCTGCTTCACCACCTTCTTGAT-3′). The cycling parameters were as
follows: initial step at 90°C for 1 min, followed by 40 cycles of
90°C for 15 sec and 60°C for 1 min. Relative gene expression levels
were calculated using the 2−ΔΔCt method normalized to
GAPDH expression levels, and fold differences in expression were
calculated relative to those of control samples.
Western blot analysis
Western blot analyses for p53 and p-p53 detection
were performed using HT29, LoVo, and HepG2 cell lysates. The cells
were grown to 70% confluence in 6-well plates. After PL treatment,
the cells were washed twice with PBS and subsequently lysed in RIPA
buffer [20 mM Tris-HCl (pH 7.4), 150 mM NaCl, 1 mM
MgCl2, and 1 mM CaCl2] with 1% Triton X-100.
Cell lysates were then centrifuged at 12,000 × g for 10 min to
pellet debris. For mouse colons, ~0.1 g tissue homogenized with
buffer [50 mM Tris-HCl (pH 7.4), 1% Triton X-100, 0.2% sodium
deoxycholate, 0.2% SDS, 1 mM EDTA, protease inhibitor cocktail (1
mM phenylmethysulfonyl fluoride, 5 μg/ml aprotinin and 5 μg/ml
leupeptin)] were added before homogenization. After homogenization,
samples were centrifuged at 12,000 × g for 10 min to pellet debris.
The total protein of cells or colons was assayed using a Bio-Rad
Protein Assay kit (Bio-Rad, Bath, UK).
To extract nuclear fractions, HT29, LoVo and HepG2
cells were collected and washed twice with ice-cold PBS. The
harvested cells were lysed in nuclear protein extraction buffer A
[10 mM HEPES (pH 7.9), 10 mM KCl, 0.1 mM EDTA and 0.1 mM EGTA with
50 μl 10% NP-40, 20 μl 0.1 M DTT, 5 μl 2 μg/ml aprotinin, and 5 μl
2 μg/ml leupeptin in 1 ml buffer added before use], and pipetted up
and down to disrupt cell clumps. The cell lysates were subsequently
centrifuged at 12,000 × g for 3 min at 4°C, and the supernatant was
removed. Then, 500 μl buffer A was added to the tube, which was
subsequently vortexed for 10 sec and centrifuged at 12,000 × g for
3 min at 4°C. The supernatant was subsequently discarded. The
pellet was lysed with 60 μl nuclear protein extraction buffer B [20
mM HEPES (pH7.9), 0.4 mM NaCl, 1 mM EDTA, and 1 mM EGTA with 10 μl
0.1 M DTT, 5 μl 2 μg/ml aprotinin, and 5 μl 2 μg/ml leupeptin in 1
ml buffer added before use] and incubated on ice for 15 min with
intermittent vortexing. The lysate was subsequently centrifuged at
12,000 × g for 10 min at 4°C, and the supernatant was collected as
the nuclear fraction. Protein concentration was measured with the
Bio-Rad Protein Assay kit.
Sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) sample buffer was added to the protein
pellets, and the samples were heated for 3 min at 95°C. Samples
were loaded (10 μg of total protein for whole-cell lysate or
nuclear fraction), and electrophoretically separated on 10%
polyacrylamide gels, and transferred to polyvinylidene difluoride
(PVDF) membranes. Western blotting was performed according to
standard protocols, and proteins were visualized using primary
antibodies against p53 (mouse monoclonal antibody, diluted
1:1,000), p-p53 (rabbit polyclonal antibody, diluted 1:1,000), and
tubulin (rat monoclonal antibody, diluted 1:1,000).
Statistical analysis
The SSRI Excel Statistics 2006 software (Japan) was
used in the statistical analysis. Data are presented as means ± SE.
Differences among the means of treatment groups were analyzed by
one-way ANOVA; significance was determined by Scheffe’s
multiple-range test. Student’s t-test was used for comparisons
between 2 groups. The level of significance for all tests was set
at P<0.05.
Results
Effect of vitamin B6 on p21
mRNA expression in cancer cells
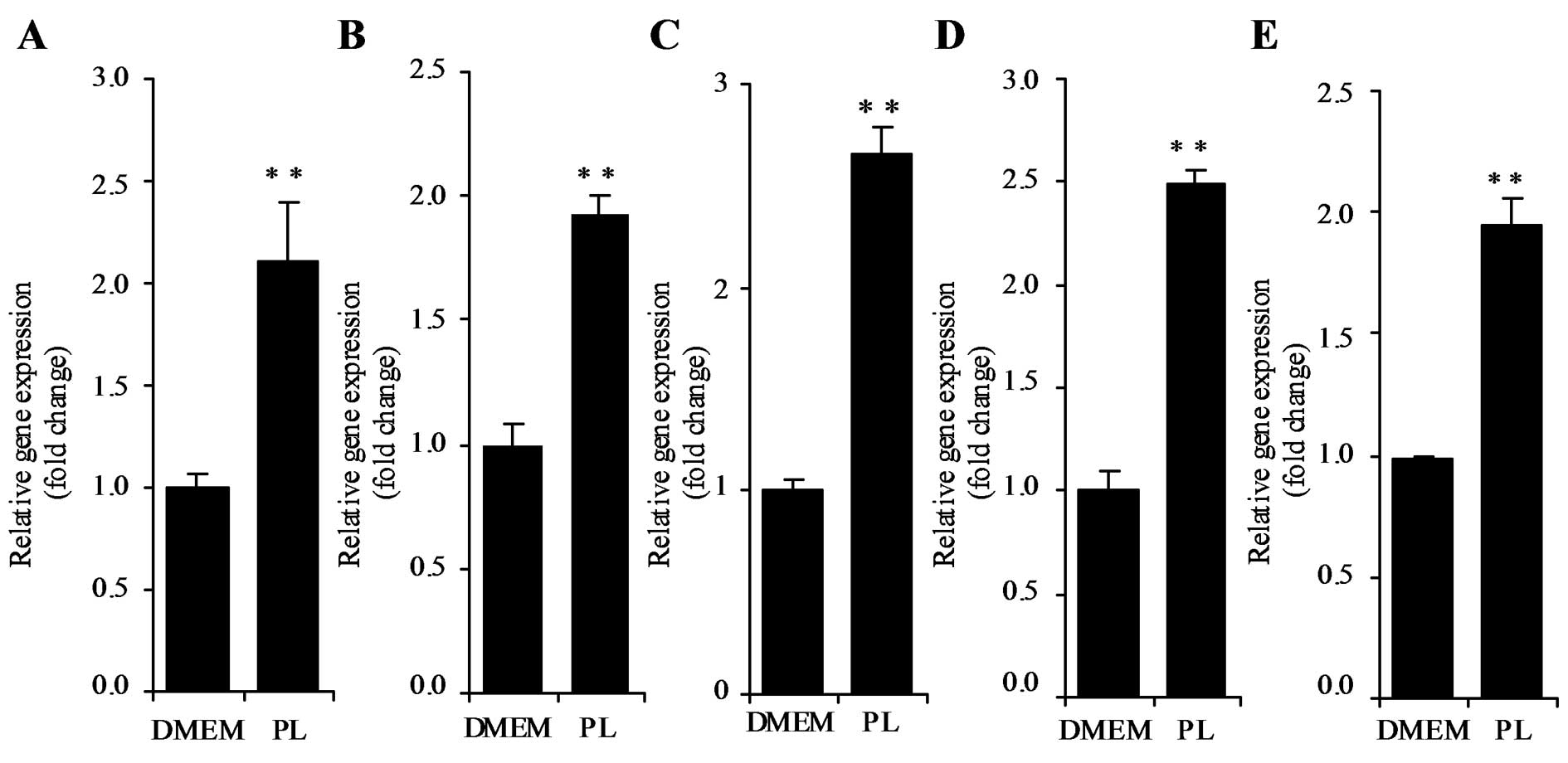
DNA microarray analysis revealed p21 mRNA expression
was upregulated in HT29 cells in response to treatment with 500 μM
PL for 24 h. Stimulation of p21 mRNA expression by PL in HT29 cells
was confirmed by real-time PCR (Fig.
1A). Caco2, LoVo, HEK293T and HepG2 cells were incubated in the
presence or absence of 500 μM PL for 24 h, and p21 mRNA levels were
examined by real-time PCR. As shown in Fig. 1B–E, p21 mRNA levels were
significantly increased by PL in Caco2, LoVo, HEK293T, and HepG2
cells (P<0.01).
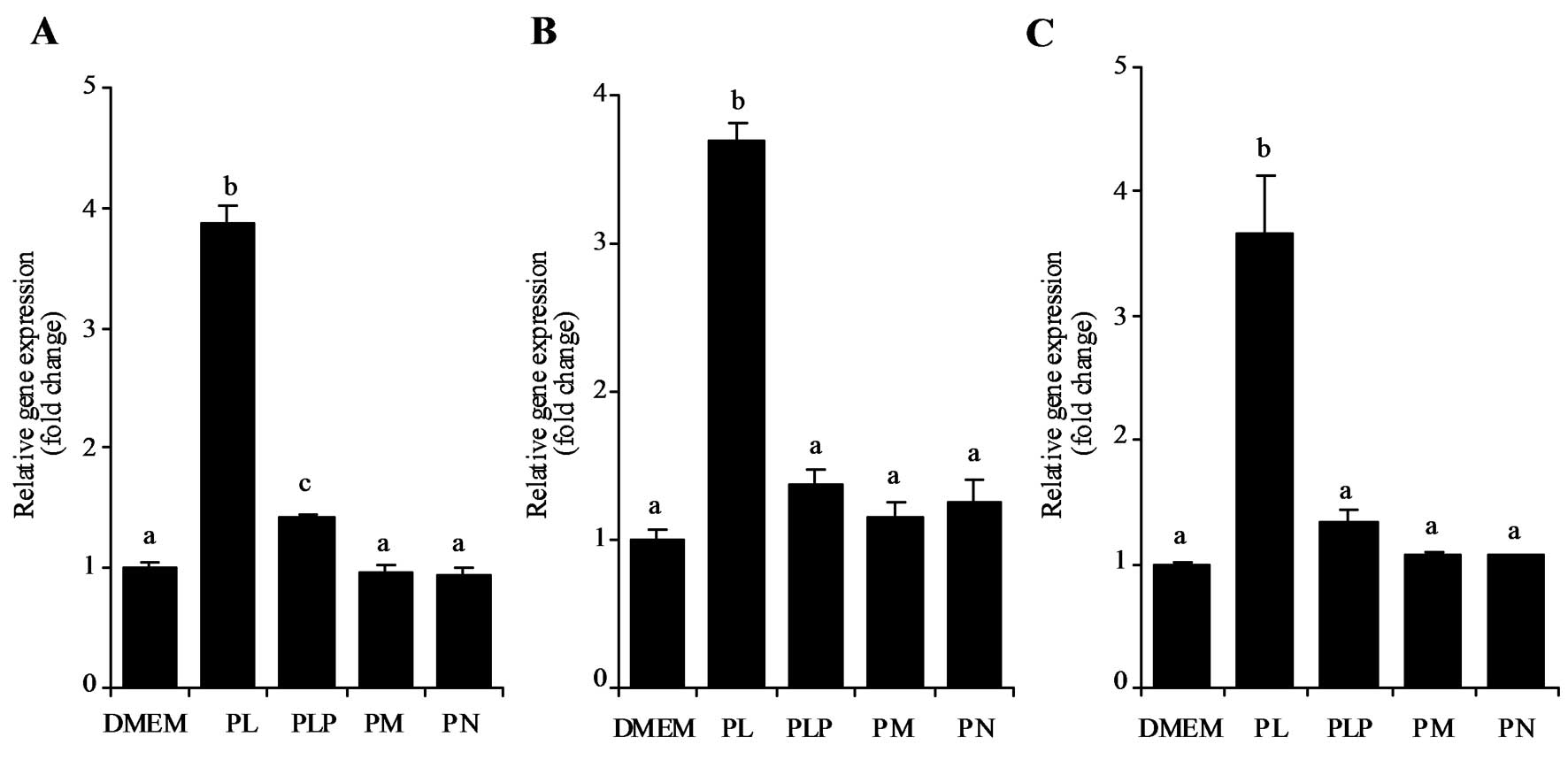
The effect of adding B6-vitamers
including PL, PM, PN, and PLP at 500 μM on p21 mRNA expression in
HT29, LoVo, and HepG2 cells were analyzed. The results indicate PL
significantly stimulated p21 mRNA expression (P<0.05) whereas
other B6-vitamers had no such effect in HT29 (Fig. 2A), LoVo (Fig. 2B), or HepG2 cells (Fig. 2C).
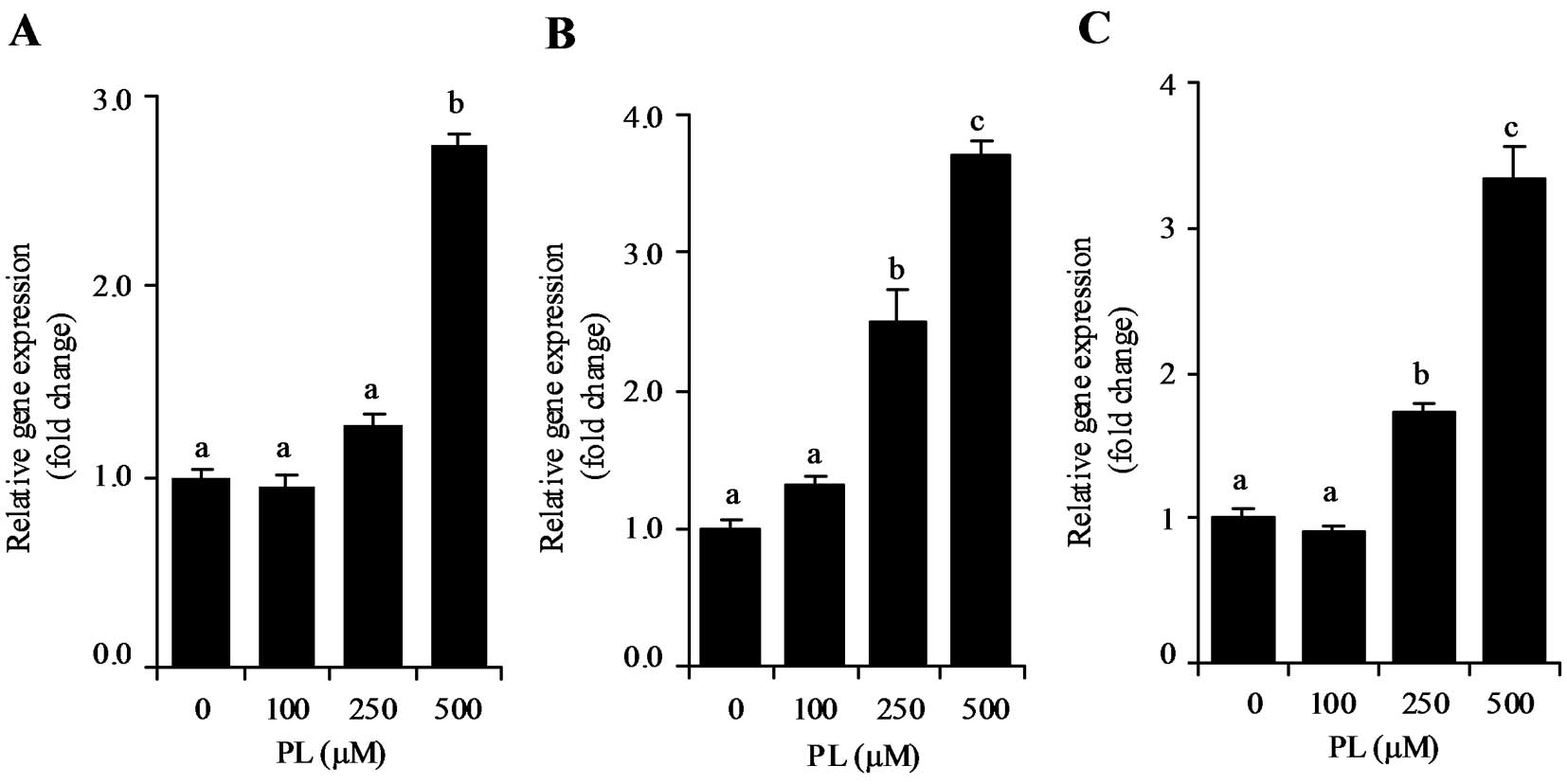
Time- and dose-dependent effect of PL on
p21 mRNA expression
We investigated the effect of different
concentrations of PL on p21 mRNA expression. After incubating HT29,
LoVo or HepG2 cells with 100, 250 or 500 μM PL for 24 h, p21 mRNA
levels increased dose-dependently from 100 to 500 μM (Fig. 3A–C). At 500 μM, PL significantly
stimulated p21 mRNA expression in HT29, LoVo and HepG2 cells
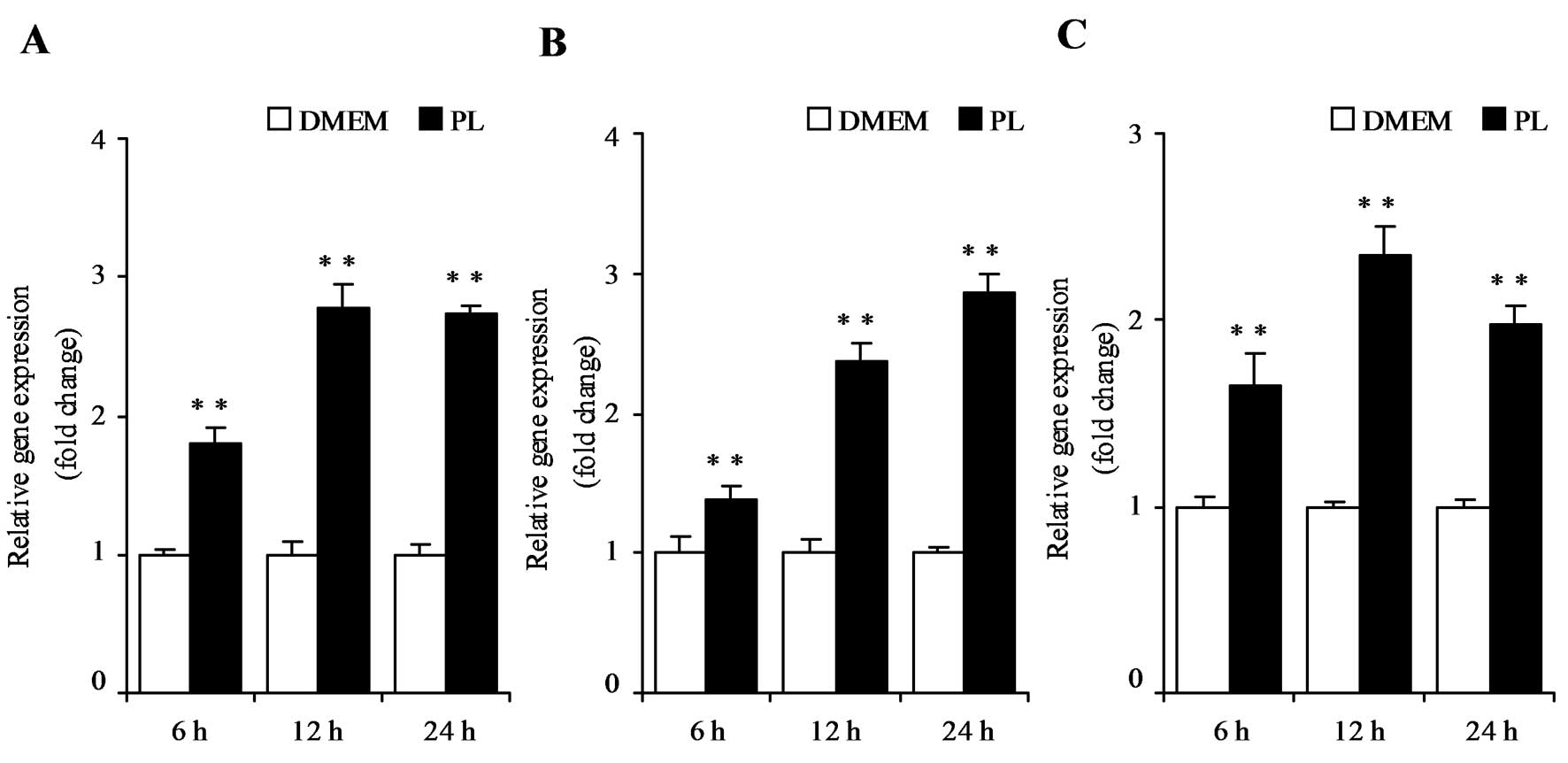
(P<0.05). To examine the time-dependent effect of PL, HT29, LoVo
or HepG2 cells were cultured with or without 500 μM PL for 6, 12 or
24 h. PL treatment increased p21 mRNA expression from 6 to 24 h in
these cells (P<0.01) (Fig.
4A–C).
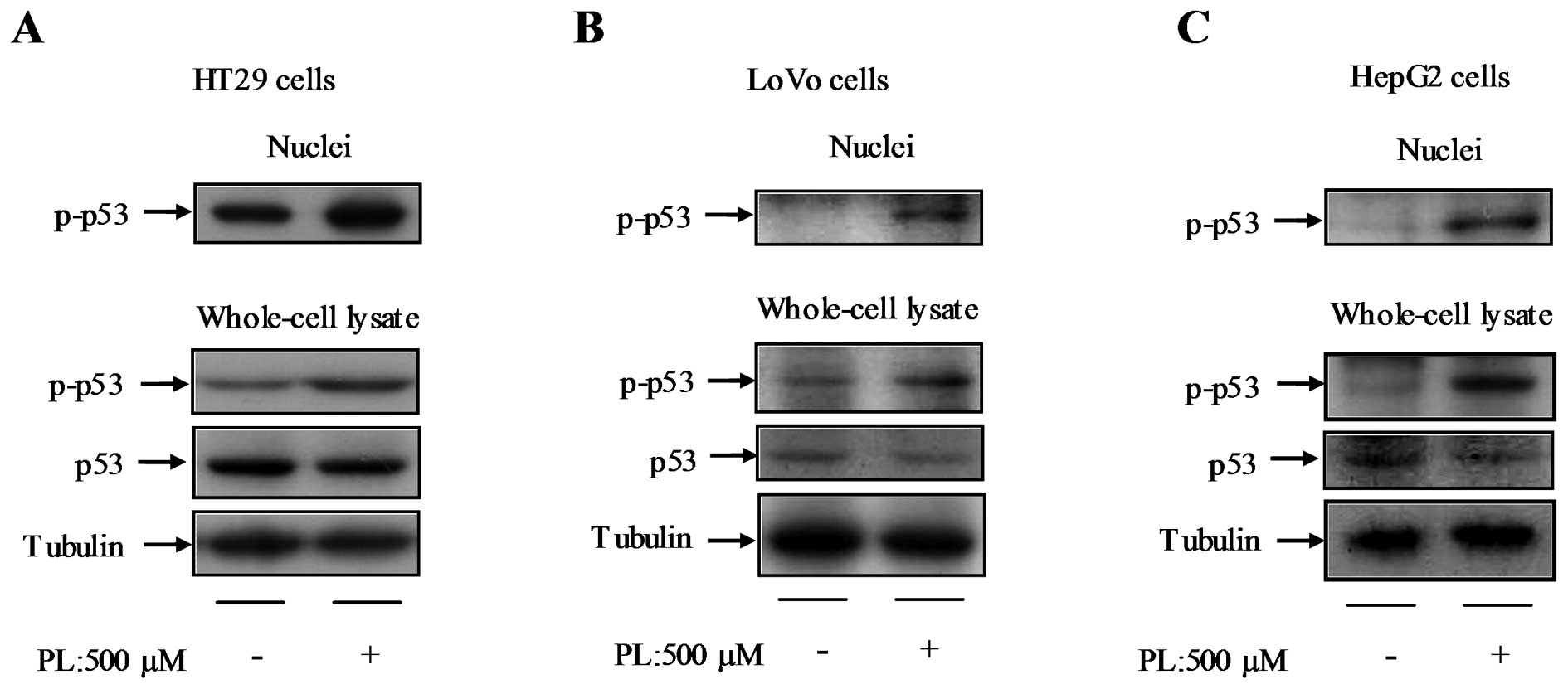
Effect of PL on p53 activation
As the transcription of p21 gene is tightly
controlled by p53, p53 gene expression was examined in HT29, LoVo,
and HepG2 cells in response to treatment with 500 μM PL for 24 h.
There was no significant difference in p53 mRNA expression between
the control and PL-treated cells (data not shown). In order to
understand the activation of the p53 pathway, p-p53 protein was
analyzed in HT29 (Fig. 5A), LoVo
(Fig. 5B), and HepG2 cells
(Fig. 5C). PL increased p-p53
protein expression in whole-cell lysates and the nuclei of these
cell lines, but did not increase the total protein level of p53
(Fig. 5A–C).
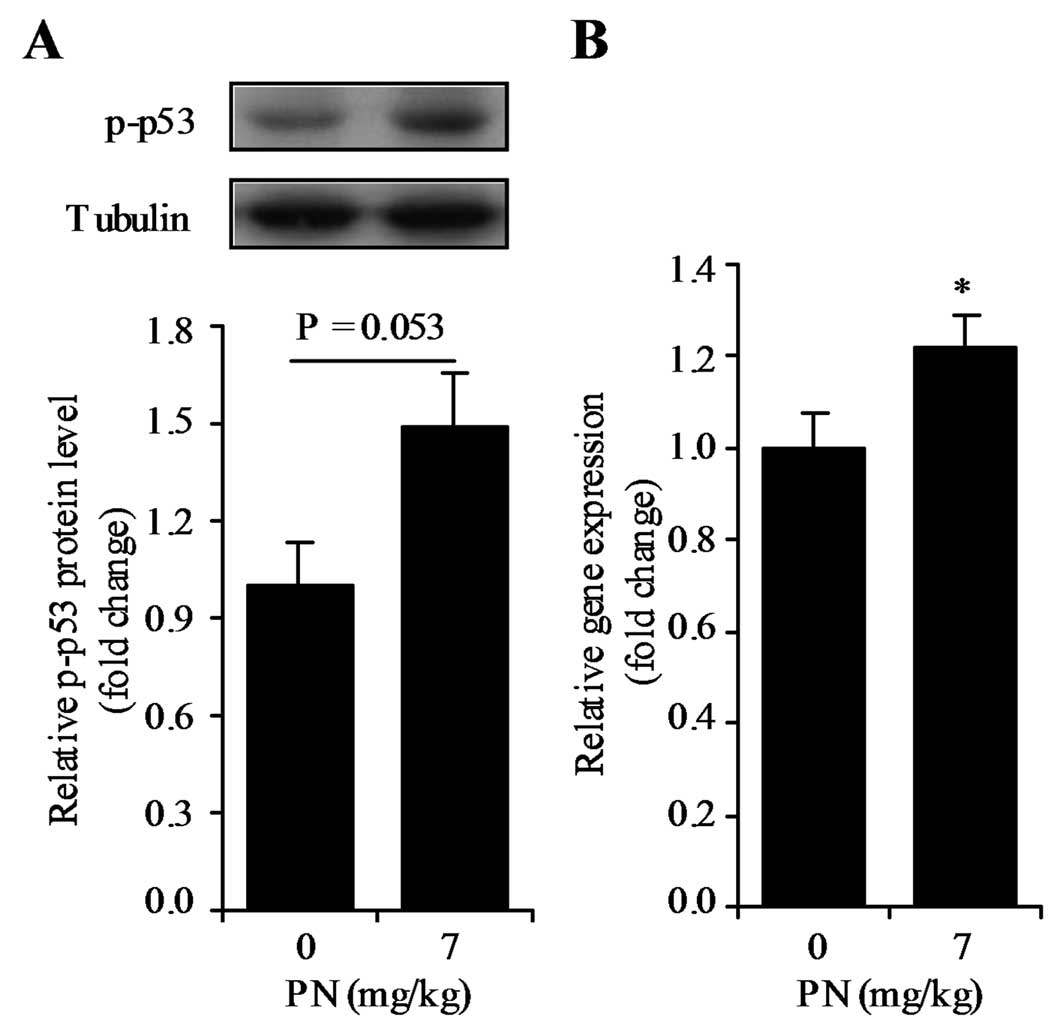
Effect of vitamin B6-deficient
diet on p-p53 protein level and p21 mRNA expression in the mouse
colon
To understand the role of vitamin B6 in
the upregulation of p21 mRNA expression in vivo, 2 groups of
mice fed diets with different vitamin B6 contents (i.e.,
0 and 7 mg PN HCl/kg) were used. There was a small, but significant
difference in final body weight between the mice fed the 0 and 7 mg
PN HCl/kg diets (41.1±1.0 g and 46.2±1.4 g, respectively)
(P<0.05). There was no significant difference in total food
intake between the 2 groups (data not shown). Mice fed the
PN-deficient diet tended to exhibit reduced p-p53 protein
expression (P=0.053) (Fig. 6A) and
significantly reduced p21 mRNA expression (P<0.05) (Fig. 6B) in the colon. There was no
association between final body weight and p-p53 protein expression
or p21 mRNA expression (P>0.05). This implies the effect of
dietary vitamin B6 may not be related to changes in body
weight.
Discussion
The results of the present study demonstrate PL
increases p21 mRNA expression in HT29, Caco2, LoVo, HEK293T and
HepG2 cells. This implies the upregulating effect of PL on p21 mRNA
expression might occur in a wide variety of cancer cells. On the
other hand, other B6-vitamers including PLP, PN and PM
had no such effect on p21 mRNA expression. We recently found that
PL remarkably increases IGFBP1 mRNA whereas no other
B6-vitamers have a similar effect (11). PL can freely pass through the cell
membrane, and only PL is reported to interact with the cell surface
of RAW264.7 cells cultured in culture medium treated with
B6-vitamers (i.e., PL, PM, PN and PLP) (17,18).
Therefore, the stimulation of p21 mRNA expression might be related
to the cell surface interaction and penetration of PL.
The results further indicate PL increases p-p53
protein levels in both the whole-cell lysate and nuclei of HT29,
LoVo and HepG2 cells. This implies PL activates p53. Concordant
with these cell culture experiments, mice fed the vitamin
B6-deficient diet exhibited decreased p21 mRNA
expression and tended to exhibit decreased phosphorylated p53
protein levels in the colon than mice fed the diet containing
adequate vitamin B6. Therefore, higher p21 expression by
vitamin B6 may be at least be partially mediated by the
activation of p53. Nakari et al show that a high dose of PN
(10 mM) inhibits the growth of MCF-7 cells and induces IGFBP3
expression in a p53-dependent manner (14). However, in the present study, 500 μM
PN did not increase p21 gene expression. Therefore, these findings
suggest PL rather than PN affects the p53/p21 pathway.
We previously suggested dietary vitamin
B6 supplementation suppresses colon tumorigenesis by
decreasing colon cell proliferation, inflammation and oxidative
stress in mice treated with AOM (1). Accumulating evidence from in
vitro and in vivo studies suggests p21 and p53 suppress
cell proliferation (2,11,19–25).
In addition, p21 protects against oxidative stress (26). Importantly, p53 and p21 are reported
to be anti-inflammation factors (27–29).
Furthermore, p21 is a negative regulator of macrophage activation;
in particular, it inhibits the lipopolysaccharide-dependent
stimulation of TNF-α and IL-1β (29,30).
Moreover, the inhibition caused by p21 inhibits the NF-κB activity
(29,30). Yanaka et al (7) reported the inhibitory effect of PL on
the lipopolysaccharide-dependent activation of NF-κB in
macrophages. In addition, activation of p53 is reported to lead to
cell cycle arrest, DNA repair, and genomic stability (31). Taken together, these findings raise
the question of whether increased p21 expression and p53 activation
by PL are associated with decreased cell proliferation, oxidative
stress and inflammation.
In conclusion, this study provides evidence of a
role of PL in the upregulation of p21 gene expression in HT29,
Caco2, LoVo, HEK293T and HepG2 cells. Furthermore, the p53 pathway,
which is responsible for controlling p21 mRNA transcription, is
activated by PL in cancer cells. Importantly, p21 mRNA levels were
higher in the colon of mice fed a diet with adequate vitamin
B6 than those fed a vitamin B6-deficient
diet. Thus, these findings may help us understand the antitumor
effect of vitamin B6 via the activation of p53 and
elevation of p21 mRNA.
Acknowledgements
This study was supported in part by a Grant-in-Aid
from the Ministry of Education, Culture, Sports, Science and
Technology of Japan.
Abbreviations:
|
AOM
|
azoxymethane
|
|
IGFBP1
|
insulin-like growth factor-binding
protein 1
|
|
IGFBP3
|
insulin-like growth factor-binding
protein 3
|
|
PL
|
pyridoxal
|
|
PN
|
pyridoxine
|
|
PM
|
pyridoxamine
|
|
PLP
|
pyridoxal 5′-phosphate
|
|
DMEM
|
Dulbecco’s modified Eagle’s medium
|
|
SDS-PAGE
|
sodium dodecyl sulphate-polyacrylamide
gel electrophoresis
|
|
PVDF
|
polyvinylidene difluoride
|
References
|
1
|
Komatsu SI, Watanabe H, Oka T, Tsuge H,
Nii H and Kato N: Vitamin B-6-supplemented diets compared with a
low vitamin B-6 diet suppress azoxymethane-induced colon
tumorigenesis in mice by reducing cell proliferation. J Nutr.
131:2204–2207. 2001.PubMed/NCBI
|
|
2
|
Komatsu S, Watanabe H, Oka T, Tsuge H and
Kato N: Dietary vitamin B6 suppresses colon tumorigenesis,
8-hydroxyguanosine, 4-hydroxynonenal, and inducible nitric oxide
synthase protein in azoxymethane-treated mice. J Nutr Sci
Vitaminol. 48:65–68. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ishihara J, Otani T, Inoue M, Iwasaki M,
Sasazuki S and Tsugane S; Japan Public Health Center-based
Prospective Study Group. Low intake of vitamin B-6 is associated
with increased risk of colorectal cancer in Japanese men. J Nutr.
137:1808–1814. 2007.PubMed/NCBI
|
|
4
|
Theodoratou E, Farrington SM, Tenesa A, et
al: Dietary vitamin B6 intake and the risk of colorectal cancer.
Cancer Epidemiol Biomarkers Prev. 17:171–182. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Larsson SC, Orsini N and Wolk A: Vitamin
B6 and risk of colorectal cancer: a meta-analysis of prospective
studies. JAMA. 303:1077–1083. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang XH, Ma J, Smith-Warner SA, Lee JE
and Giovannucci E: Vitamin B6 and colorectal cancer:
current evidence and future directions. World J Gastroenterol.
19:1005–1010. 2013.
|
|
7
|
Yanaka N, Koyama TA, Komatsu S, Nakamura
E, Kanda M and Kato N: Vitamin B6 suppresses NF-κB
activation in LPS-stimulated mouse macrophages. Int J Mol Med.
16:1071–1075. 2005.
|
|
8
|
Matsubara K, Mori M, Matsuura Y and Kato
N: Pyridoxal 5′-phosphate and pyridoxal inhibit angiogenesis in
serum-free rat aortic ring assay. Int J Mol Med. 8:505–508.
2001.
|
|
9
|
Komatsu S, Yanaka N, Matsubara K and Kato
N: Antitumor effect of vitamin B6 and its mechanisms.
Biochim Biophys Acta. 1647:127–130. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kayashima T, Tanaka K, Okazaki Y,
Matsubara K, Yanaka N and Kato N: Consumption of vitamin
B6 reduces colonic damage and protein expression of
HSP70 and HO-1, the anti-tumor targets, in rats exposed to
1,2-dimethylhydrazine. Oncol Lett. 2:1243–1246. 2011.
|
|
11
|
Zhang PP, Suidasari S, Hasegawa T, Yanaka
N and Kato N: High concentrations of pyridoxal stimulate the
expression of IGFBP1 in HepG2 cells through upregulation of the
ERK/c-Jun pathway. Mol Med Rep. 8:973–978. 2013.PubMed/NCBI
|
|
12
|
Harper JW, Adami GR, Wei N, Keyomarsi K
and Elledge SJ: The p21 Cdk-interacting protein Cip1 is a potent
inhibitor of G1 cyclin-dependent kinases. Cell. 75:805–816. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Deiry WS, Tokino T, Velculescu VE, et al:
WAF1, a potential mediator of p53 tumor suppression. Cell.
75:817–825. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nakari M, Kanouchi H and Oka T: High dose
of pyridoxine induces IGFBP-3 mRNA expression in MCF-7 cells and
its induction is inhibited by the p53-specific inhibitor
pifithrin-α. J Nutr Sci Vitaminol. 57:280–284. 2011.PubMed/NCBI
|
|
15
|
Masisi K, Suidasari S, Zhang P, Okazaki Y,
Yanaka N and Kato N: Comparative study on the responses of
concentrations of B6-vitamers in several tissues of mice
to the dietary level of pyridoxine. J Nutr Sci Vitaminol.
58:446–451. 2012.PubMed/NCBI
|
|
16
|
Reeves GP, Nielsen HF and Fahey CG Jr:
AIN-93 purified diets for laboratory rodents: Final report of the
American Institute of Nutrition ad hoc writing committee on
the reformulation of the AIN-76A rodent diet. J Nutr.
123:1939–1951. 1993.PubMed/NCBI
|
|
17
|
Kanouchi H, Shibuya M, Tsukamoto S,
Fujimura Y, Tachibana H, Yamada K and Oka T: Comparisons of uptake
and cell surface binding among pyridoxal, pyridoxine, and
pyridoxamine in RAW264.7 cells. Nutrition. 26:648–652. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sakurai T, Asakura T, Mizuno A and Matsuda
M: Absorption and metabolism of pyridoxamine in mice. I Pyridoxal
as the only form of transport in blood. J Nutr Sci Vitaminol.
37:341–348. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pasz-Walczak G and Kordek R: Comparative
evaluation of the expression of cell cycle regulating proteins:
cyclin D1, P53 and P21 (WAF1) in colorectal cancer. Pol J Pathol.
51:63–69. 2000.PubMed/NCBI
|
|
20
|
King ML and Murphy LL: Role of cyclin
inhibitor protein p21 in the inhibition of HCT116 human colon
cancer cell proliferation by American ginseng (Panax
quinquefolius) and its constituents. Phytomedicine. 17:261–268.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Volakaki AA, Lafkas D, Kassi E, Schally
AV, Papavassiliou AG and Kiaris H: Essential role of p21/waf1 in
the mediation of the anti-proliferative effects of GHRH antagonist
JMR-132. J Mol Endocrinol. 41:389–392. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang WC, Mathew J, Velcich A, et al:
Targeted inactivation of the p21(WAF1/cip1) gene enhances
Apc-initiated tumor formation and the tumor-promoting activity of a
Western-style high-risk diet by altering cell maturation in the
intestinal mucosal. Cancer Res. 61:565–569. 2001.PubMed/NCBI
|
|
23
|
Poole AJ, Heap D, Carroll RE and Tyner AL:
Tumor suppressor functions for the Cdk inhibitor p21 in the mouse
colon. Oncogene. 23:8128–8134. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang W, Velcich A, Lozonschi I, et al:
Inactivation of p21WAF1/cip1 enhances intestinal tumor
formation in Muc2−/− mice. Am J Pathol. 166:1239–1246.
2005.PubMed/NCBI
|
|
25
|
Zirbes TK, Baldus SE, Moenig SP, et al:
Prognostic impact of p21/waf1/cip1 in colorectal cancer. Int J
Cancer. 89:14–18. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Vitiello PF, Wu YC, Staversky RJ and
O’Reilly MA: p21(Cip1) protects against oxidative stress by
suppressing ER-dependent activation of mitochondrial death
pathways. Free Radic Biol Med. 46:33–41. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Komarova EA, Krivokrysenko V, Wang K, et
al: p53 is a suppressor of inflammatory response in mice. FASEB J.
19:1030–1032. 2005.PubMed/NCBI
|
|
28
|
Kawauchi K, Araki K, Tobiume K and Tanaka
N: Loss of p53 enhances catalytic activity of IKKbeta through
O-linked beta-N-acetyl glucosamine modification. Proc Natl Acad Sci
USA. 106:3431–3436. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Scatizzi JC, Mavers M, Hutcheson J, et al:
The CDK domain of p21 is a suppressor of IL-1beta-mediated
inflammation in activated macrophages. Eur J Immunol. 39:820–825.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Trakala M, Arias CF, García MI, et al:
Regulation of macrophage activation and septic shock susceptibility
via p21(WAF1/CIP1). Eur J Immunol. 39:810–819. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Vousden KH and Lane DP: p53 in health and
disease. Nat Rev Mol Cell Biol. 8:275–283. 2007. View Article : Google Scholar
|