1. Introduction
Glioma is one type of brain tumor that arises from
glial cells and accounts for over 30% of all primary central
nervous system tumors in the USA (1). Glioblastoma is the most common and
malignant subtype of glioma, which is categorized as grade IV
according to the classification of the World Health Organization
(WHO). The median survival time of glioblastoma patients is
approximately 14 months, in spite of aggressive surgery, radiation
and chemotherapy (2).
Nuclear factor erythroid 2-related factor 2 (Nrf2)
belongs to a subset of basic leucine-zipper (bZip) genes sharing a
conserved structural domain (3). It
is broadly expressed in tissues and can be activated in response to
a range of oxidative and electrophilic stimulation. The activity of
Nrf2 is primarily regulated by its inhibitor Kelch-like
ECH-associated protein 1 (Keap1) (4). When uncoupled from the Nrf2/Keap1
complex, Nrf2 is transported into the nucleus and modulates the
expression of antioxidant genes through interaction with the
antioxidant response element (ARE) (5). An increasing body of literature has
revealed alternative mechanisms of Nrf2 activation, including
phosphorylation of Nrf2 by various protein kinases, interaction
with other protein partners (p21, caveolin-1) and epigenetic
factors (microRNA-144, -28 and -200a and promoter methylation)
(6).
Recently, Nrf2 has been demonstrated as an important
regulator in different types of cancer. A dramatic increase in
research focusing on Nrf2 and the associated mechanisms in the
regulation of primary malignant brain tumors such as glioblastoma
has been carried out. High expression of Nrf2 in glioblastoma was
found to protects it from the killing effects of antitumor
therapies, and blocking of Nrf2 can inhibit glioblastoma. Thus,
Nrf2 is a potential new target with which to treat glioblastoma.
The mechanisms of the downregulation of Nrf2 in treating
glioblastoma contain two main aspects: direct and indirect means.
Direct mechanisms target tumor molecular pathways to overcome
resistance to chemotherapy and radiotherapy, to inhibit
proliferation, to block invasion and migration, to induce
apoptosis, to promote differentiation, to enhance autophagy and to
target glioblastoma stem cells (GSCs). Indirect mechanisms target
the reaction between glioblastoma cells and the surrounding
microenvironment, such as the perivascular, hypoxic and immune
microenvironments. In the present study, we review the function of
Nrf2 in the regulation of glioblastoma, and the associated
mechanisms concerning the downregulation of Nrf2 in treating
glioblastoma.
2. Direct mechanisms (Table I)
Overcoming resistance to chemotherapy and
radiotherapy
Standard treatment of glioblastoma currently
involves chemotherapy and radiotherapy. However, glioblastoma can
easily develop resistance to chemotherapy and radiotherapy. It has
been found that high expression of Nrf2 decreases the sensitivity
of glioblastoma cells to chemotherapy and radiotherapy.
Chemotherapy
There are a variety of tumors that develop strong
tolerance to chemotherapy, including glioblastoma (7). Recently, the role of Nrf2 in inducing
chemotherapy resistance has been reported in several types of
tumors (8). In glioblastoma, Nrf2
expression was found to be increased during drug resistance
(8). Temozolomide (TMZ) is an
alkylating agent which is commonly used for the treatment of
glioblastoma (9–11). TMZ treatment was found to induce
Nrf2 activation in the glioblastoma cell line U251 and
downregulation of Nrf2 expression increased TMZ-induced cell death
in U251 cells (12). In addition,
the silencing of Nrf2 also increased cell necrosis induced by
5-fluorouracil (5-FU), cisplatin, etoposide (13–15),
oxaliplatin (16) and doxorubicin
(ADM) (17,18). Blocking Nrf2 activation is a
potential method for enhancing chemotherapy sensitivity of
glioblastoma cells (19).
Nrf2 may induce the chemoresistance of glioblastoma
through stress response and a drug efflux mechanism (Fig. 1). The stress response mechanism
implies that Nrf2 transcription upregulates endogenous phase II
detoxifying enzymes, which may inactivate antitumor drugs by
modifying their structures (20).
In addition, activation of Nrf2 was also found to contribute to
drug efflux pathways (21).
ATP-binding cassette, subfamily G, member 2 (ABCG2) plays a crucial
role in the efflux of xenobiotics and drugs, and Nrf2-mediated
regulation of ABCG2 was found to increase the efflux of antitumor
drugs and decrease the effect of chemotherapy (21). However, research suggests that Nrf2
is not an independent molecule in chemoresistance. The possible
role of peroxiredoxin1 (Prx1) co-functioning with Nrf2 in
chemoresistance has been suggested (22).
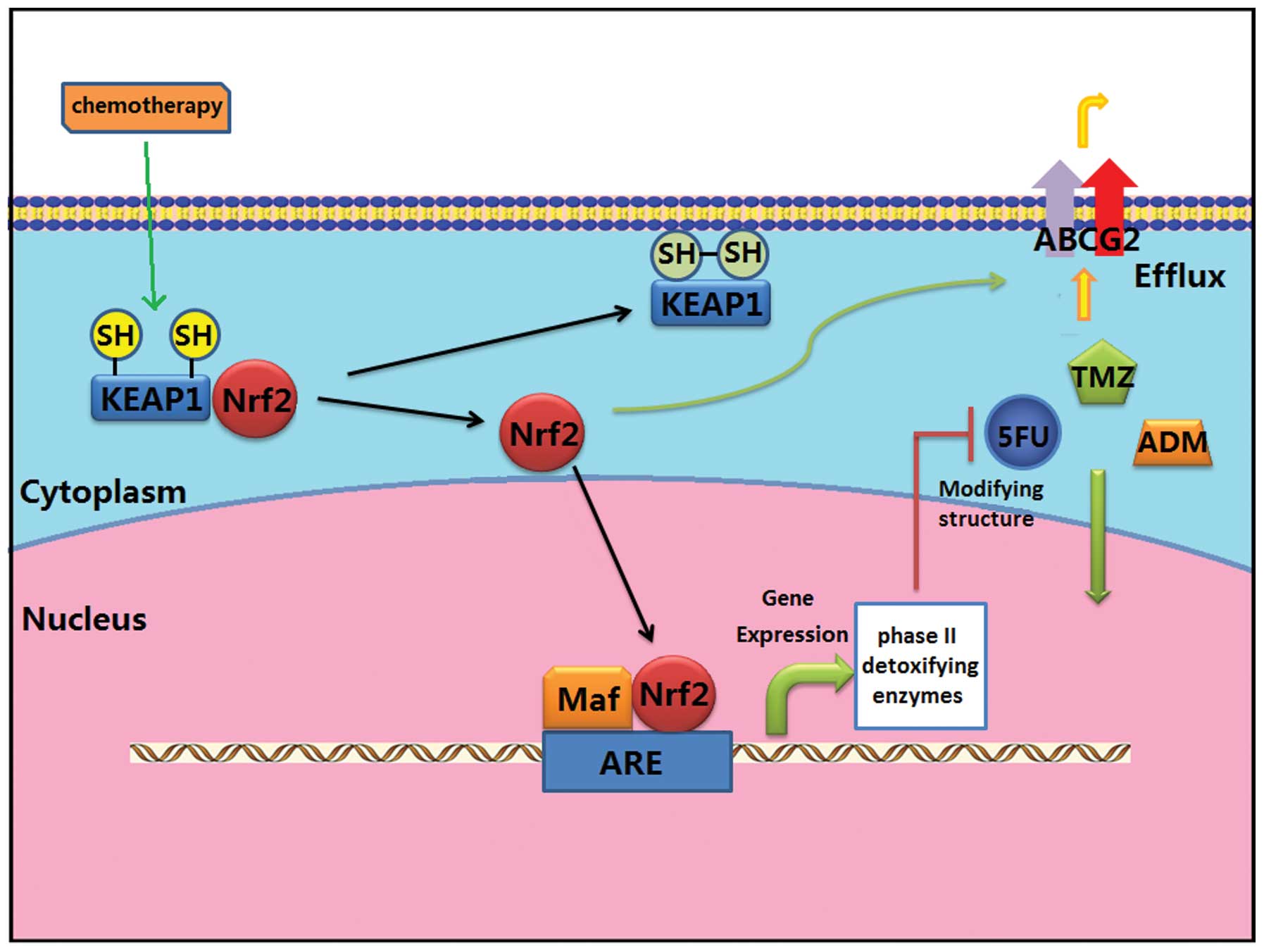 | Figure 1Mechanisms of chemoresistance of
glioblastoma induced by Nrf2. Nrf2 is activated by chemotherapy and
is transported into the nucleus. Nrf2 binds to the ARE region and
promotes the expression of phase II detoxifying enzymes, which
inactive antitumor drugs by modifying their structures. In
addition, Nrf2 upregulates the expression of ABCG2, increasing the
efflux of antitumor drugs. Nrf2, nuclear factor erythroid 2-related
factor 2; TMZ, temozolomide; ADM, doxorubicin; 5-FU,
5-fluorouracil; ARE, antioxidant response element; ABCG2,
ATP-binding cassette, subfamily G, member 2, Keap1, Kelch-like
ECH-associated protein 1. |
Radiotherapy
Radiotherapy is the foundation of therapy following
maximal surgical resection of glioblastoma (23,24).
However, glioblastoma displays high resistance to radiotherapy
(25). Low-dose radiation induces
Nrf2 activation reactively (12).
The role of Nrf2 in radioresistance has been investigated. Using a
genetically modified method to establish continuous activation of
Nrf2, Nrf2 was found to protect glioblastoma against ionizing
radiation toxicity, and Nrf2-inhibited tumor cells showed increased
sensitivity to γ-irradiation (26).
The Nrf2/ARE pathway regulates the radioresistance
of glioblastoma by modifying endogenous Nrf2 inhibitor and by
upregulating the downstream signal of Nrf2 (27). Radioresistance may involve the
loss-of-function mutations of the Nrf2 inhibitor Keap1, which
allows Nrf2 to be continuously transported to the nucleus (28). Other research has demonstrated that
Nrf2 induces radioresistance by regulating the function of the
major downstream molecule heme oxygenase-1 (HO-1) (29). Downstream activation of
Nrf2-ARE-dependent HO-1 was found to be important in the
maintenance of resistance to irradiation (12).
Inhibition of proliferation
Glioblastoma cells usually maintain a high rate of
proliferation. High expression of Nrf2 gives glioblastoma an
advantage for growth, and knockdown of Nrf2 was found to inhibit
the proliferation and growth of human glioblastoma cells (20,30,31).
The candidate mechanisms of Nrf2 in the regulation
of proliferation mainly include three means: i) upregulation of
downstream molecules of Nrf2; ii) cross-talk with other signaling
pathways; iii) and post-transcriptional regulation. Nrf2 can induce
the growth of tumor cells by increasing the expression of HO-1,
glutathione peroxidase-2 (GPx2) (32,33)
and CXCR3-B (34), which are
downstream molecules of Nrf2 and are important in the regulation of
the growth and proliferation of glioblastoma. The growth rate of
cancer cells is inhibited by downregulation of these molecules.
Nrf2 is also involved in regulating a variety of other signal
transduction pathways. Recently, studies have demonstrated that
Nrf2 can enhance cell proliferation by regulating epidermal growth
factor receptor (EGFR), Ki-67, Kras, and phosphoinositide-3-kinase
(PI3K)/Akt pathway, which are necessary for maintaining the
proliferation of glioblastoma (35–38).
Finally, Nrf2 may improve the accumulation of various
proliferation-related proteins by regulating the associated small
interfering RNA fraction. Recent studies have identified several
microRNAs (miRs) as post-translational targets of Nrf2 to regulate
proliferation. Studies have shown that NADPH and ribose are
essential for the cell proliferation in tumors (39,40),
and loss of Nrf2 was found to decrease the expression of the
redox-sensitive histone deacetylase HDAC4, resulting in increased
expression of miR-1, miR-200a and miR-206, which markedly impaired
NADPH production and ribose synthesis (41,42).
Blocking of invasion and migration
Glioblastoma can easily invade and migrate to
surrounding brain tissue. Nrf2 may facilitate the remodeling of the
tumor microenvironment making it advantageous for the autonomic
invasion and migration of cancer cells (43). Nrf2 acts as a master switch in these
processes by upregulating the expression of various invasion and
migration-related proteins (44).
The Nrf2/ARE pathway may regulate glioblastoma
invasion and migration through matrix metalloproteinases (MMPs) and
oxidative stress-related molecules. MMP activation could improve
the degradation of intercellular connections, which enables
glioblastoma cells to easily invade and migrate (45). Downregulation of the expression of
Nrf2 in the U251 glioblastoma cell line was found to inactivate
matrix metalloproteinase-9 (MMP-9) and to decrease the invasion and
migration of glioma (44).
Oxidative stress is another important mechanism involved in the
invasion and migration of glioblastoma. HO-1 is the downstream
molecule of Nrf2, which is important in regulating oxidative
stress. Inhibition of HO-1 can weaken the invasive and migratory
abilities of glioblastoma (46,47).
However, Thangasamy et al found that the Nrf2
inducer sulforaphane (SFN) can inhibit the expression of tyrosine
kinase receptor, recepteur d’origine nantais (RON), which can
mediate the invasion of carcinoma cells (48), indicating that Nrf2 may play a dual
role in regulating the invasiveness of tumors.
Induction of apoptosis
In most glioblastoma cells, apoptosis is inhibited
(49,50). It has been suggested that Nrf2 can
block the apoptotic death of cancer cells (51). Overexpression of Nrf2 was found to
significantly diminish apoptosis (52). Inhibition of the Nrf2 transcription
factor rendered cancer cells more susceptible to apoptosis
(53).
The Nrf2/ARE pathway may regulate apoptosis by
cross-linking with the B-cell lymphoma 2 (Bcl2), p53,
p38/mitogen-activated protein kinase (MAPK) and nuclear factor-κB
(NF-κB) pathways. Bcl2 is an important gene in tumor genesis and in
the anti-apoptosis process (54,55).
Following increased expression of Nrf2, the expression of caspases
3 was decreased and the apoptosis rate was reduced, accompanied by
the upregulated expression of Bcl-2/Bax. This indicates that Nrf2
regulates apoptosis through the Bcl2-related pathway (56,57).
p53 is important due to its anticancer function, and plays an
essential role in tumor apoptosis (58). Nrf2 also regulates the
tumor-suppressor p53 by influencing the degradation of p53. The
Nrf2 downstream molecule NQO1 interacts with p53 and induces its
degradation by the proteasome in a ubiquitin-independent manner
(59). In addition, Nrf2 also
attenuates the effect of the apoptosis inducer diamide in
glioblastoma by upregulating the activity of p38/MAPK and
inhibiting the NF-κB pathway (60,61).
Promotion of differentiation
Glioblastoma cells are usually in a poor stage of
differentiation and exhibit low maturity (62–64),
and differentiation therapy is required as a therapeutic strategy
for malignant tumors (65,66). Nrf2 induces the suppression of
differentiation by inhibiting a powerful differentiation inducer
1α, 25-dihydroxyvitamin D3 (1,25 D3) (67,68),
suggesting that Nrf2 plays an important role in the cooperative
suppression of cancer cell differentiation.
Nrf2 may regulate the differentiation of
glioblastoma through cross-talk with the Notch pathway and
upregulation of anti-redox molecules. The Notch pathway is
important for cell-cell communication, which involves genetic
regulatory mechanisms that control the cell differentiation process
(69). Nrf2 adaptive response
pathway could directly activate the Notch signal through
recruitment of the Notch intracellular domain (NICD) transcriptome
and restrain glioblastoma cells in a low state of differentiation
(70). In addition, high
accumulation of reactive oxygen species (ROS) can induce the
differentiation of cells (71).
Nrf2 was found to upregulate the anti-redox molecule GST to
eliminate ROS and reverse the differentiation induced by ROS
(71,72). It has been reported that neuronal
differentiation inducer retinoic acid (RA) increased Nrf2
expression reactively (73,74), and downregulation of Nrf2 improves
the efficiency of RA in inducing differentiation (73,74).
Enhancement of autophagy
Autophagy is a lysosomal degradation process.
Autophagy principally plays an adaptive role to protect organisms
against diverse pathological conditions (75,76).
Many studies have shed light on the importance of autophagy in
glioblastoma (77). Knockdown of
Nrf2 was found to regulate the autophagy induced by TMZ in the U251
glioblastoma cell line (78).
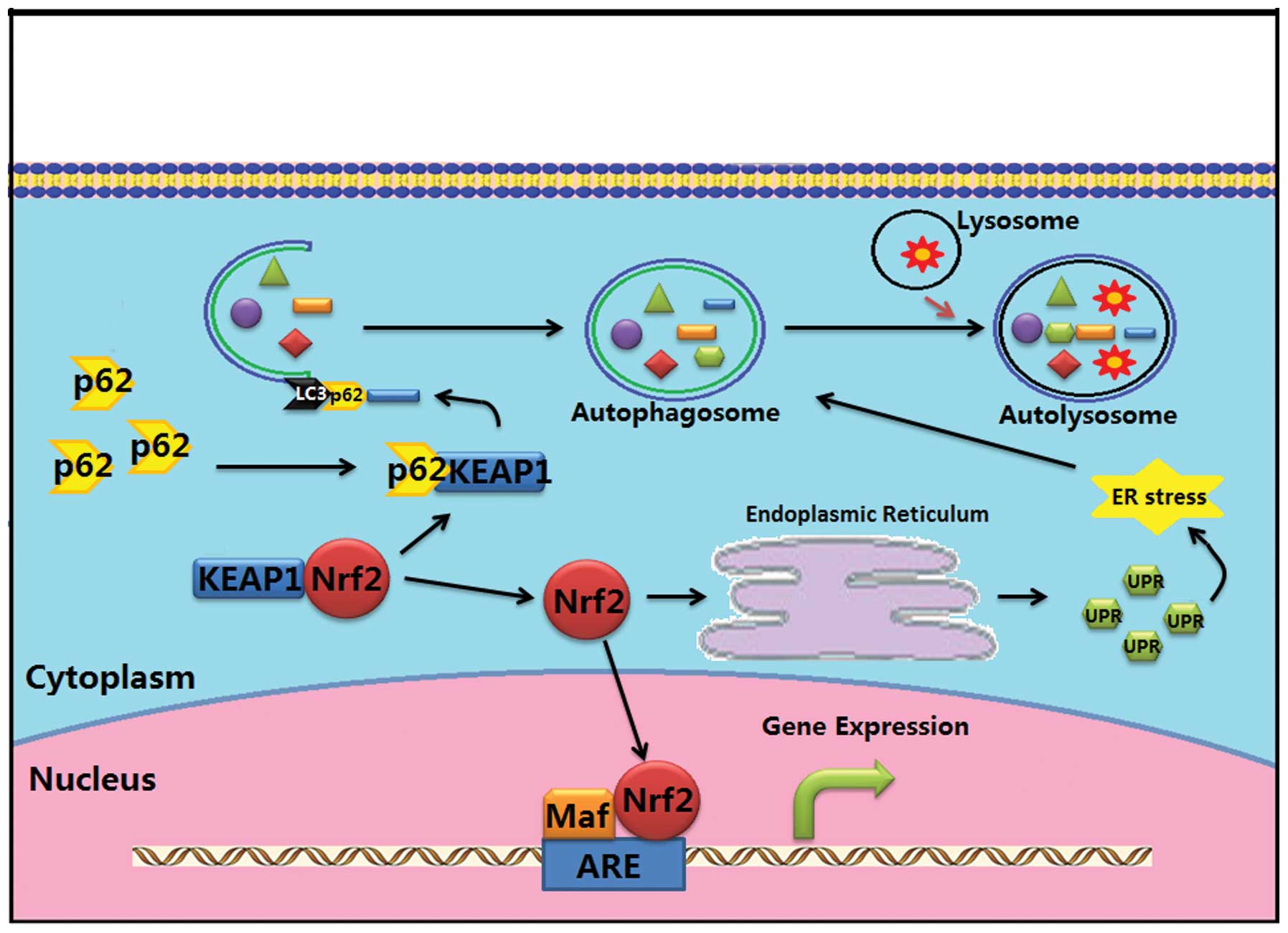
Nrf2 may regulate autophagy by altering the
P62/SQSTM1 system and endoplasmic reticulum (ER) stress reaction
(Fig. 2). The protein of p62, also
known as sequestosome 1 (SQSTM1), is one of the adaptors of
autophagy. It has been found to play a critical role in the
formation of cytoplasmic proteinaceous inclusion. Keap1 uncoupled
from the complex with Nrf2 can bind to the autophagy-adaptor
protein p62, and then interacts with LC3 and transports the
ubiquitin conjugate to the autophagosome for degradation (79–81).
ER stress is a cellular stress response which is activated in
response to an accumulation of the unfolded protein response (UPR).
High expression of Nrf2 can also induce autophagy by increasing ER
stress and by increasing ER-associated degradation (82).
Targeting GSCs
The glioma stem cell (GSC) hypothesis suggests that
neoplastic clones are maintained exclusively by a rare fraction of
cells with stem cell properties (83). The identification of brain
tumor-initiating cells established a new cellular target for more
effective therapies (84–86). Over the past decades, Nrf2 was found
to be pivotal in the maintenance of the stemness of human GSCs.
Knockdown of Nrf2 was found to inhibit the proliferation of GSCs,
and significantly reduce the expression of self-renewal-related
factors Bmi1, Sox2 and cyclin E (87).
Nrf2 may maintain the stemness of GSCs by
cross-linking with MAPK and p53 pathway, regulating HO-1 and
circulating cell-free DNA (cirDNA) (Fig. 3) (88). High expression of Nrf2 can regulate
the expression of MAPK and p53 in stem cells, which plays a
critical role in the self-renewal of GSCs, indicating that Nrf2 may
regulate self-renewal through MAPK and p53 pathway (89). Nrf2 downstream compound HO-1 is
important in maintaining the high proliferation of stem cells. The
HO-1 inducer cobalt protoporphyrin (CoPP) markedly improved stem
cell proliferation (90). Nrf2 also
plays an important role in regulating the reaction of stem cells to
cirDNA, which is a small fraction of DNA in the plasma and has been
found to be important in inhibiting the apoptosis of stem cells.
(91).
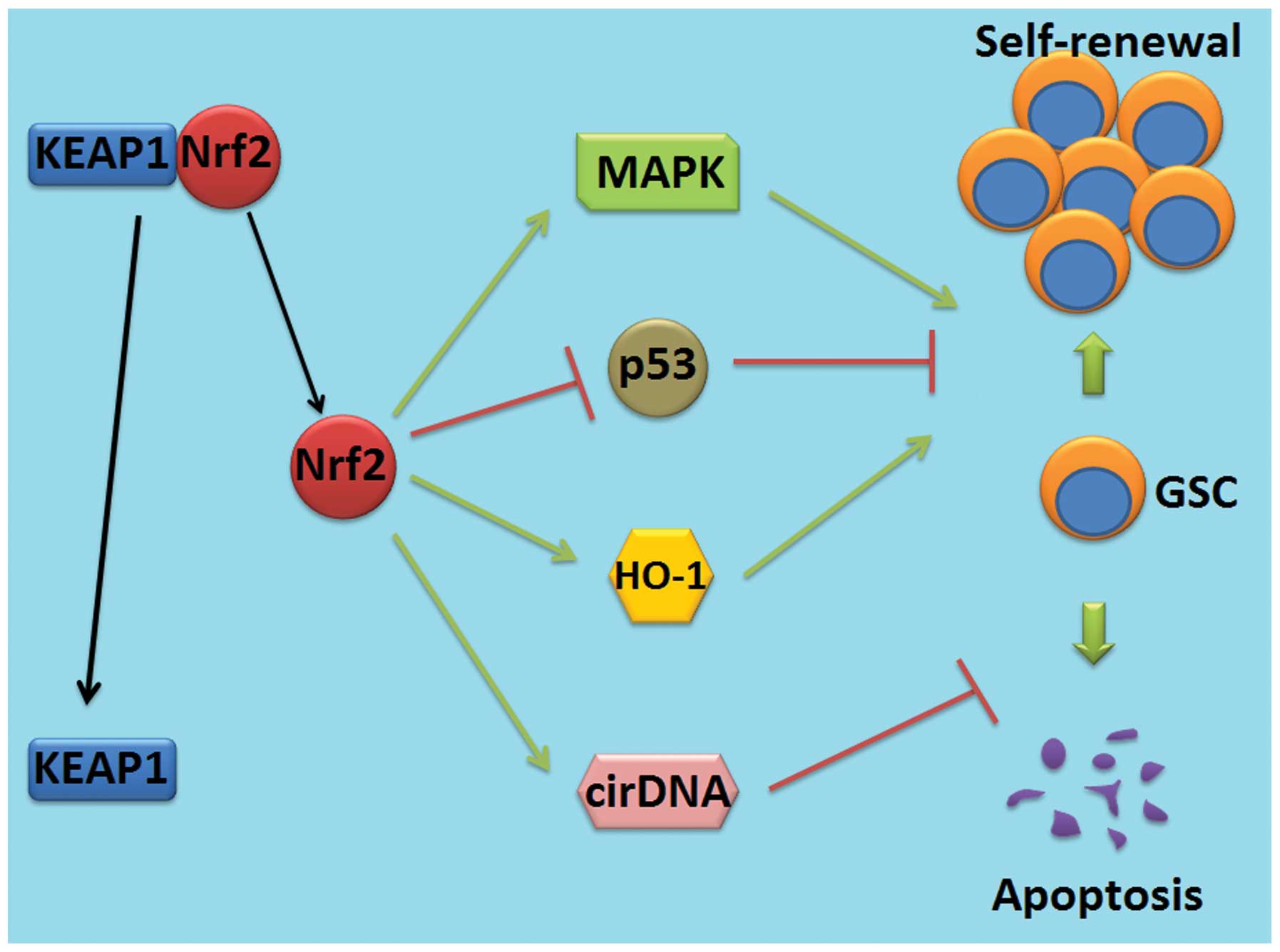 | Figure 3Role of Nrf2 in regulating the
function of GSCs in glioblastoma. After uncoupled from the
Nrf2/Keap1 complex, Nrf2 increases the expression of MAPK and
inhibits p53, along with the Nrf2 downstream compound HO-1,
maintaining the self-renewal of GSCs. Nrf2 also regulates the
reaction of GSCs to cirDNA, inhibiting the apoptosis of GSCs. Nrf2,
nuclear factor erythroid 2-related factor 2; MAPK,
mitogen-activated protein kinase; HO-1, heme oxygenase-1; cirDNA,
circulating cell-free DNA; GSCs, glioma stem cells, Keap1,
Kelch-like ECH-associated protein 1. |
3. Indirect mechanisms
The microenvironment is a functional unit enabling
complex and dynamic interactions with tumor cells (92). Glioblastoma cells are influenced by
non-malignant cells of the tumor microenvironment such as vascular
endothelial cells, fibroblasts and immune cells (93). The microenvironment serves as the
basis for indirect mechanisms of Nrf2 in the treatment of
glioblastoma. Indirect mechanisms include three main aspects of the
microenvironment: i) perivascular, ii) hypoxic and iii) immune
microenvironment (Table II).
 | Table IIIndirect mechanisms of the
downregulation of Nrf2 in the treatment of glioblastoma. |
Table II
Indirect mechanisms of the
downregulation of Nrf2 in the treatment of glioblastoma.
| Mechanisms | Factors and
associated molecules |
|---|
|
Microenvironment |
| Perivascular | HIF-1α, VEGF |
| Hypoxic | HIF-1α, HO-1 |
| Immune | Cytokines: IFN-γ,
IL-4, IL-5, IL-13
Immune cells: Th, microglia |
Perivascular microenvironment
Angiogenesis plays a key role in glioblastoma in
order to provide energy and maintain the development and
progression of glioblastoma. Glioblastoma cells develop a framework
to induce the angiogenesis around them (94,95).
Recent studies have begun to explore the role of Nrf2 in tumor
angiogenesis (96,97). In human glioblastoma cell line U251,
knockdown of Nrf2 was found to significantly decrease microvessel
density (MVD) and expression of small vessel marker CD31 (38).
Nrf2 may regulate angiogenesis through
hypoxia-inducible factor 1α (HIF-1α) and vascular endothelial
growth factors (VEGFs). As a main downstream molecule of Nrf2,
HIF-1α is one of the master regulators that orchestrate cellular
responses to hypoxia. Activation of HIF-1α can lead to the
activation of numerous perivascular compounds, such as
angiopoietin, endothelin-1, inducible nitric oxide synthase (iNOS),
adrenomedullin and erythropoietin. Blocking HIF-1α can inhibit the
angiogenesis effect of Nrf2 (98).
Another important inducer of vessels is VEGF. Nrf2 elevates VEGF
expression and improves the growth of the vascular endothelia in
tumors. Through a positive feedback loop, VEGF can also activate
Nrf2 in an ERK1/2-dependent manner and induce the production of
antioxidative enzymes (99).
Anti-angiogenesis effects of Nrf2 knockdown were documented in
chick chorioallantoic membrane assays and endothelial tube
formation assays (100).
Hypoxic microenvironment
Hypoxia and tumor genesis are closely related
(101). Glioblastoma has extensive
areas of hypoxia and displays high tolerance to a low concentration
of oxygen (102,103). Nrf2 has been identified as a
regulator of several genes involved in the hypoxic defense
response, such as HIF-1α (104).
In human glioblastoma, high expression of Nrf2 was significantly
correlated with high tolerance to a low concentration of oxygen,
less tumor necrosis on MRI and lower 1-year survival of patients
(105).
It is believed that Nrf2 regulates the hypoxia
resistance by HIF-1α and HO-1. HIF-1α is a downstream molecule of
Nrf2 and is one of the master regulators of hypoxia (98). In a CoCl2-induced hypoxia
model, blockage of Nrf2 suppressed the expression of HIF-1α, and
suppressed the migration and invasion of tumors in a hypoxic
microenvironment (106). HO-1 is
another important molecule for resistance to hypoxia. In a
6-hydroxydopamine (6-OHDA)-induced hypoxic model, Nrf2 activation
induced upregulation of HO-1, and mediated the cellular adaptive
survival response to a hypoxic microenvironment (107).
Immune microenvironment
Glioblastoma can escape from tumor
immunosurveillance and inactivate the reaction between tumors and
immune cells. The immune microenvironment surounding glioblastoma
plays an important role in these processes (108). In addition, Nrf2 was also found to
be a critical regulator of the immune reaction (109).
The Nrf2/ARE pathway may regulate tumor
immunosurveillance through regulation of the secretion of cytokines
and the function of immune cells. Nrf2 regulates the secretion of
many types of cytokines. Activation of Nrf2 was found to suppress
the production of interferon-γ (IFN-γ), while inducing the
production of T helper cells 2 (Th2), cytokines IL-4, IL-5, and
IL-13 (110). Nrf2 also regulates
the function of immune cells. In glioblastoma, T helper cells (Th)
play an important role in the adaptive immune system. Th helps the
activation of other immune cells by releasing T cell cytokines.
Nrf2 is a regulator of Th and activates CD4(+) T cells from
differentiating towards Th2, representing a novel regulatory
mechanism in CD4(+) T cells (111). Microglia act as the main form of
active immune defense in the central nervous system (CNS). Nrf2
also mediates immunoresistance by modifying the function of
microglia. Activation of the Nrf2/HO-1 pathway was found to
suppress BV2 microglial cells and immunology in the brain (112). Upregulation of Nrf2 suppressed
innate immune microglial cells in the CNS. Various small activators
of Nrf2/HO-1 such as carnosol, supercurcumin and dimethyl fumarate
are effective modulators of microglial-related immune responses
(112).
4. Conclusion
In the past decades, a marked increasing in research
has been carried out focusing on Nrf2 and its role in regulating
glioblastoma and the possibilities of the downregulation of Nrf2
for treating glioblastoma. Nrf2 plays an extensively role in the
regulation of glioblastoma; hence, downregulation of Nrf2 can
interfere with a variety of behaviors of glioblastoma and actions
of the microenvironment surrounding glioblastoma. Thus Nrf2 has
promising value as a therapeutic target for glioblastoma. However,
Nrf2 downregulation in most studies was obtained through RNA
interference or knockdown technology, rather than pharmaceutical
compounds, making targeted Nrf2 therapy somewhat difficult and less
appealing at this time from a translational perspective. Recently,
biochemists have identified the small molecule, ochratoxin A, as an
inhibitor of Nrf2 (113). Although
it is a toxin produced by Aspergillus ochraceus, the single
compound is a potential new strategy with which to inhibit Nrf2 in
glioblastoma. For these reasons, future studies should focus on
regulatory methods of Nrf2, which can be easily translated to the
clinical setting and be used safely.
Acknowledgements
This study was supported by grants from the National
Natural Science Foundation of China (nos. 81070974 and 81271377),
the Jiangsu Provincial Key Subject (no. X4200722), and Jinling
Hospital (no. 2010Q017).
References
|
1
|
Binello E and Germano IM: Targeting glioma
stem cells: a novel framework for brain tumors. Cancer Sci.
102:1958–1966. 2011. View Article : Google Scholar
|
|
2
|
Van Meir EG, Hadjipanayis CG, Norden AD,
et al: Exciting new advances in neuro-oncology: the avenue to a
cure for malignant glioma. CA Cancer J Clin. 60:166–193.
2010.PubMed/NCBI
|
|
3
|
Kensler TW, Wakabayashi N and Biswal S:
Cell survival responses to environmental stresses via the
Keap1-Nrf2-ARE pathway. Annu Rev Pharmacol Toxicol. 47:89–116.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Reuter S, Gupta SC, Chaturvedi MM and
Aggarwal BB: Oxidative stress, inflammation, and cancer: how are
they linked? Free Radic Biol Med. 49:1603–1616. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li H, Wang F, Zhang L, et al: Modulation
of Nrf2 expression alters high glucose-induced oxidative stress and
antioxidant gene expression in mouse mesangial cells. Cell Signal.
23:1625–1632. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bryan HK, Olayanju A, Goldring CE and Park
BK: The Nrf2 cell defence pathway: Keap1-dependent and -independent
mechanisms of regulation. Biochem Pharmacol. 85:705–717. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Livingston DM and Silver DP: Cancer:
crossing over to drug resistance. Nature. 451:1066–1067. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen Q, Li W, Wan Y, et al: Amplified in
breast cancer 1 enhances human cholangiocarcinoma growth and
chemoresistance by simultaneous activation of Akt and Nrf2
pathways. Hepatology. 55:1820–1829. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Stupp R, Mason WP, van den Bent MJ, et al:
Radiotherapy plus concomitant and adjuvant temozolomide for
glioblastoma. N Engl J Med. 352:987–996. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Newlands ES, Stevens MF, Wedge SR, et al:
Temozolomide: a review of its discovery, chemical properties,
pre-clinical development and clinical trials. Cancer Treat Rev.
23:35–61. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Friedman HS, Kerby T and Calvert H:
Temozolomide and treatment of malignant glioma. Clin Cancer Res.
6:2585–2597. 2000.PubMed/NCBI
|
|
12
|
Cong ZX, Wang HD, Zhou Y, et al:
Temozolomide and irradiation combined treatment-induced Nrf2
activation increases chemoradiation sensitivity in human
glioblastoma cells. J Neurooncol. 116:41–48. 2014. View Article : Google Scholar
|
|
13
|
Hu XF, Yao J, Gao SG, et al: Nrf2
overexpression predicts prognosis and 5-fu resistance in gastric
cancer. Asian Pac J Cancer Prev. 14:5231–5235. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jiang T, Chen N, Zhao F, et al: High
levels of Nrf2 determine chemoresistance in type II endometrial
cancer. Cancer Res. 70:5486–5496. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang XJ, Sun Z, Villeneuve NF, et al: Nrf2
enhances resistance of cancer cells to chemotherapeutic drugs, the
dark side of Nrf2. Carcinogenesis. 29:1235–1243. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen CC, Chu CB, Liu KJ, et al: Gene
expression profiling for analysis acquired oxaliplatin resistant
factors in human gastric carcinoma TSGH-S3 cells: the role of IL-6
signaling and Nrf2/AKR1C axis identification. Biochem Pharmacol.
86:872–887. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gao AM, Ke ZP, Shi F, et al: Chrysin
enhances sensitivity of BEL-7402/ADM cells to doxorubicin by
suppressing PI3K/Akt/Nrf2 and ERK/Nrf2 pathway. Chem Biol Interact.
206:100–108. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gao AM, Ke ZP, Wang JN, et al: Apigenin
sensitizes doxorubicin-resistant hepatocellular carcinoma
BEL-7402/ADM cells to doxorubicin via inhibiting PI3K/Akt/Nrf2
pathway. Carcinogenesis. 34:1806–1814. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim WD, Kim YW, Cho IJ, et al: E-cadherin
inhibits nuclear accumulation of Nrf2: implications for
chemoresistance of cancer cells. J Cell Sci. 125:1284–1295. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lau A, Villeneuve NF, Sun Z, et al: Dual
roles of Nrf2 in cancer. Pharmacol Res. 58:262–270. 2008.
View Article : Google Scholar
|
|
21
|
Singh A, Wu H, Zhang P, et al: Expression
of ABCG2 (BCRP) is regulated by Nrf2 in cancer cells that confers
side population and chemoresistance phenotype. Mol Cancer Ther.
9:2365–2376. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kim JH, Bogner PN, Ramnath N, et al:
Elevated peroxiredoxin 1, but not NF-E2-related factor 2, is an
independent prognostic factor for disease recurrence and reduced
survival in stage I non-small cell lung cancer. Clin Cancer Res.
13:3875–3882. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Alexander BM, Ligon KL and Wen PY:
Enhancing radiation therapy for patients with glioblastoma. Expert
Rev Anticancer Ther. 13:569–581. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Caruso C, Carcaterra M and Donato V: Role
of radiotherapy for high grade gliomas management. J Neurosurg Sci.
57:163–169. 2013.PubMed/NCBI
|
|
25
|
Frosina G: DNA repair and resistance of
gliomas to chemotherapy and radiotherapy. Mol Cancer Res.
7:989–999. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Singh A, Bodas M, Wakabayashi N, et al:
Gain of Nrf2 function in non-small-cell lung cancer cells confers
radioresistance. Antioxid Redox Signal. 13:1627–1637. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sharma PK and Varshney R:
2-Deoxy-D-glucose and 6-aminonicotinamide-mediated Nrf2 down
regulation leads to radiosensitization of malignant cells via
abrogation of GSH-mediated defense. Free Radic Res. 46:1446–1457.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang P, Singh A, Yegnasubramanian S, et
al: Loss of Kelch-like ECH-associated protein 1 function in
prostate cancer cells causes chemoresistance and radioresistance
and promotes tumor growth. Mol Cancer Ther. 9:336–346. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Na HK and Surh YJ: Oncogenic potential of
Nrf2 and its principal target protein heme oxygenase-1. Free Radic
Biol Med. 67:353–365. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
DeNicola GM, Karreth FA, Humpton TJ, et
al: Oncogene-induced Nrf2 transcription promotes ROS detoxification
and tumorigenesis. Nature. 475:106–109. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Taguchi K, Motohashi H and Yamamoto M:
Molecular mechanisms of the Keap1-Nrf2 pathway in stress response
and cancer evolution. Genes Cells. 16:123–140. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Brigelius-Flohé R, Müller M, Lippmann D
and Kipp AP: The yin and yang of nrf2-regulated selenoproteins in
carcinogenesis. Int J Cell Biol. 2012:4861472012.PubMed/NCBI
|
|
33
|
Lu SC: Regulation of glutathione
synthesis. Mol Aspects Med. 30:42–59. 2009. View Article : Google Scholar
|
|
34
|
Balan M and Pal S: A novel CXCR3-B
chemokine receptor-induced growth-inhibitory signal in cancer cells
is mediated through the regulation of Bach-1 protein and Nrf2
protein nuclear translocation. J Biol Chem. 289:3126–3137. 2014.
View Article : Google Scholar
|
|
35
|
Yamadori T, Ishii Y, Homma S, et al:
Molecular mechanisms for the regulation of Nrf2-mediated cell
proliferation in non-small-cell lung cancers. Oncogene.
31:4768–4777. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zou W, Chen C, Zhong Y, et al: PI3K/Akt
pathway mediates Nrf2/ARE activation in human L02 hepatocytes
exposed to low-concentration HBCDs. Environ Sci Technol.
47:12434–12440. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kong B, Qia C, Erkan M, et al: Overview on
how oncogenic Kras promotes pancreatic carcinogenesis by inducing
low intracellular ROS levels. Front Physiol. 4:2462013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ji XJ, Chen SH, Zhu L, et al: Knockdown of
NF-E2-related factor 2 inhibits the proliferation and growth of
U251MG human glioma cells in a mouse xenograft model. Oncol Rep.
30:157–164. 2013.PubMed/NCBI
|
|
39
|
Crosas-Molist E, Bertran E, Sancho P, et
al: The NADPH oxidase NOX4 inhibits hepatocyte proliferation and
liver cancer progression. Free Radic Biol Med. 69:338–347. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang Z, Li Y, Lv S and Tian Y: Inhibition
of proliferation and invasiveness of ovarian cancer C13*
cells by a poly(ADP-ribose) polymerase inhibitor and the role of
nuclear factor-κB. J Int Med Res. 41:1577–1585. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Singh A, Happel C, Manna SK, et al:
Transcription factor NRF2 regulates miR-1 and miR-206 to drive
tumorigenesis. J Clin Invest. 123:2921–2934. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Petrelli A, Perra A, Cora D, et al:
MicroRNA/gene profiling unveils early molecular changes and nuclear
factor erythroid related factor 2 (NRF2) activation in a rat model
recapitulating human hepatocellular carcinoma (HCC). Hepatology.
59:228–241. 2014. View Article : Google Scholar
|
|
43
|
Rachakonda G, Sekhar KR, Jowhar D, et al:
Increased cell migration and plasticity in Nrf2-deficient cancer
cell lines. Oncogene. 29:3703–3714. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Pan H, Wang H, Zhu L, et al: The role of
Nrf2 in migration and invasion of human glioma cell U251. World
Neurosurg. 80:363–370. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Deryugina EI, Bourdon MA, Luo GX, et al:
Matrix metalloproteinase-2 activation modulates glioma cell
migration. J Cell Sci. 110:2473–2482. 1997.PubMed/NCBI
|
|
46
|
Gan FF, Ling H, Ang X, et al: A novel
shogaol analog suppresses cancer cell invasion and inflammation,
and displays cytoprotective effects through modulation of NF-κB and
Nrf2-Keap1 signaling pathways. Toxicol Appl Pharmacol. 272:852–862.
2013.PubMed/NCBI
|
|
47
|
Zhang L, Wang N, Zhou S, et al: Propofol
induces proliferation and invasion of gallbladder cancer cells
through activation of Nrf2. J Exp Clin Cancer Res. 31:662012.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Thangasamy A, Rogge J, Krishnegowda NK, et
al: Novel function of transcription factor Nrf2 as an inhibitor of
RON tyrosine kinase receptor-mediated cancer cell invasion. J Biol
Chem. 286:32115–32122. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Evan GI and Vousden KH: Proliferation,
cell cycle and apoptosis in cancer. Nature. 411:342–348. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Johnstone RW, Ruefli AA and Lowe SW:
Apoptosis: a link between cancer genetics and chemotherapy. Cell.
108:153–164. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Bat-Chen W, Golan T, Peri I, et al:
Allicin purified from fresh garlic cloves induces apoptosis in
colon cancer cells via Nrf2. Nutr Cancer. 62:947–957. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Chen X, Liu J and Chen SY: Over-expression
of Nrf2 diminishes ethanol-induced oxidative stress and apoptosis
in neural crest cells by inducing an antioxidant response. Reprod
Toxicol. 42:102–109. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Arlt A, Sebens S, Krebs S, et al:
Inhibition of the Nrf2 transcription factor by the alkaloid
trigonelline renders pancreatic cancer cells more susceptible to
apoptosis through decreased proteasomal gene expression and
proteasome activity. Oncogene. 32:4825–4835. 2013. View Article : Google Scholar
|
|
54
|
Liston P, Fong WG and Korneluk RG: The
inhibitors of apoptosis: there is more to life than Bcl2. Oncogene.
22:8568–8580. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Thomadaki H and Scorilas A: BCL2
family of apoptosis-related genes: functions and clinical
implications in cancer. Crit Rev Clin Lab Sci. 43:1–67. 2006.
View Article : Google Scholar
|
|
56
|
Heasman SA, Zaitseva L, Bowles KM, et al:
Protection of acute myeloid leukaemia cells from apoptosis induced
by front-line chemotherapeutics is mediated by haem oxygenase-1.
Oncotarget. 2:658–668. 2011.PubMed/NCBI
|
|
57
|
Niture SK and Jaiswal AK: INrf2 (Keap1)
targets Bcl-2 degradation and controls cellular apoptosis. Cell
Death Differ. 18:439–451. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Attardi LD: The role of p53-mediated
apoptosis as a crucial anti-tumor response to genomic instability:
lessons from mouse models. Mutat Res. 569:145–157. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Rotblat B, Melino G and Knight RA: NRF2
and p53: Januses in cancer? Oncotarget. 3:1272–1283.
2012.PubMed/NCBI
|
|
60
|
Filomeni G, Piccirillo S, Rotilio G and
Ciriolo MR: p38MAPK and ERK1/2 dictate cell
death/survival response to different pro-oxidant stimuli via
p53 and Nrf2 in neuroblastoma cells SH-SY5Y. Biochem Pharmacol.
83:1349–1357. 2012.
|
|
61
|
Lee YM, Auh QS, Lee DW, et al: Involvement
of Nrf2-mediated upregulation of heme oxygenase-1 in
mollugin-induced growth inhibition and apoptosis in human oral
cancer cells. Biomed Res Int. 2013:2106042013.PubMed/NCBI
|
|
62
|
Sell S: Stem cell origin of cancer and
differentiation therapy. Crit Rev Oncol Hematol. 51:1–28. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Bollag W and Holdener EE: Retinoids in
cancer prevention and therapy. Ann Oncol. 3:513–526.
1992.PubMed/NCBI
|
|
64
|
Clarke N, Germain P, Altucci L and
Gronemeyer H: Retinoids: potential in cancer prevention and
therapy. Expert Rev Mol Med. 6:1–23. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Hansen LA, Sigman CC, Andreola F, et al:
Retinoids in chemoprevention and differentiation therapy.
Carcinogenesis. 21:1271–1279. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Leszczyniecka M, Roberts T, Dent P, et al:
Differentiation therapy of human cancer: basic science and clinical
applications. Pharmacol Ther. 90:105–156. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Bobilev I, Novik V, Levi I, et al: The
Nrf2 transcription factor is a positive regulator of myeloid
differentiation of acute myeloid leukemia cells. Cancer Biol Ther.
11:317–329. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Li K, Zhong C, Wang B, et al: Nrf2
expression participates in growth and differentiation of
endometrial carcinoma cells in vitro and in vivo. J Mol Histol.
45:161–167. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Capaccione KM and Pine SR: The Notch
signaling pathway as a mediator of tumor survival. Carcinogenesis.
34:1420–1430. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Wakabayashi N, Skoko JJ, Chartoumpekis DV,
et al: Notch-Nrf2 axis: regulation of Nrf2 gene expression
and cytoprotection by Notch signaling. Mol Cell Biol. 34:653–663.
2014.PubMed/NCBI
|
|
71
|
Kanzaki H, Shinohara F, Kajiya M and
Kodama T: The Keap1/Nrf2 protein axis plays a role in osteoclast
differentiation by regulating intracellular reactive oxygen species
signaling. J Biol Chem. 288:23009–23020. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Jayakumar S, Kunwar A, Sandur SK, et al:
Differential response of DU145 and PC3 prostate cancer cells to
ionizing radiation: role of reactive oxygen species, GSH and Nrf2
in radiosensitivity. Biochim Biophys Acta. 1840:485–494. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Wang XJ, Hayes JD, Henderson CJ and Wolf
CR: Identification of retinoic acid as an inhibitor of
transcription factor Nrf2 through activation of retinoic acid
receptor alpha. Proc Natl Acad Sci USA. 104:19589–19594. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Tan KP, Kosuge K, Yang M and Ito S: NRF2
as a determinant of cellular resistance in retinoic acid
cytotoxicity. Free Radic Biol Med. 45:1663–1673. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Levine B and Kroemer G: Autophagy in the
pathogenesis of disease. Cell. 132:27–42. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Mizushima N, Levine B, Cuervo AM and
Klionsky DJ: Autophagy fights disease through cellular
self-digestion. Nature. 451:1069–1075. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Kondo Y, Kanzawa T, Sawaya R and Kondo S:
The role of autophagy in cancer development and response to
therapy. Nat Rev Cancer. 5:726–734. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Zhou Y, Wang HD, Zhu L, et al: Knockdown
of Nrf2 enhances autophagy induced by temozolomide in U251 human
glioma cell line. Oncol Rep. 29:394–400. 2013.PubMed/NCBI
|
|
79
|
Fan W, Tang Z, Chen D, et al: Keap1
facilitates p62-mediated ubiquitin aggregate clearance via
autophagy. Autophagy. 6:614–621. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Kwon J, Han E, Bui CB, et al: Assurance of
mitochondrial integrity and mammalian longevity by the
p62-Keap1-Nrf2-Nqo1 cascade. EMBO Rep. 13:150–156. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Inami Y, Waguri S, Sakamoto A, et al:
Persistent activation of Nrf2 through p62 in hepatocellular
carcinoma cells. J Cell Biol. 193:275–284. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Digaleh H, Kiaei M and Khodagholi F: Nrf2
and Nrf1 signaling and ER stress crosstalk: implication for
proteasomal degradation and autophagy. Cell Mol Life Sci.
70:4681–4694. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Reya T, Morrison SJ, Clarke MF and
Weissman IL: Stem cells, cancer, and cancer stem cells. Nature.
414:105–111. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Singh SK, Hawkins C, Clarke ID, et al:
Identification of human brain tumour initiating cells. Nature.
432:396–401. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Singh SK, Clarke ID, Terasaki M, et al:
Identification of a cancer stem cell in human brain tumors. Cancer
Res. 63:5821–5828. 2003.PubMed/NCBI
|
|
86
|
Bao S, Wu Q, McLendon RE, et al: Glioma
stem cells promote radioresistance by preferential activation of
the DNA damage response. Nature. 444:756–760. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Zhu J, Wang H, Sun Q, et al: Nrf2 is
required to maintain the self-renewal of glioma stem cells. BMC
Cancer. 13:3802013. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Tsai JJ, Dudakov JA, Takahashi K, et al:
Nrf2 regulates haematopoietic stem cell function. Nat Cell Biol.
15:309–316. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Wang K, Zhang T, Dong Q, et al: Redox
homeostasis: the linchpin in stem cell self-renewal and
differentiation. Cell Death Dis. 4:e5372013. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Cai C, Teng L, Vu D, et al: The heme
oxygenase 1 inducer (CoPP) protects human cardiac stem cells
against apoptosis through activation of the extracellular
signal-regulated kinase (ERK)/NRF2 signaling pathway and cytokine
release. J Biol Chem. 287:33720–33732. 2012. View Article : Google Scholar
|
|
91
|
Loseva P, Kostyuk S, Malinovskaya E, et
al: Extracellular DNA oxidation stimulates activation of NRF2 and
reduces the production of ROS in human mesenchymal stem cells.
Expert Opin Biol Ther. 12(Suppl 1): S85–S97. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Hoelzinger DB, Demuth T and Berens ME:
Autocrine factors that sustain glioma invasion and paracrine
biology in the brain microenvironment. J Natl Cancer Inst.
99:1583–1593. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Joyce JA and Pollard JW:
Microenvironmental regulation of metastasis. Nat Rev Cancer.
9:239–252. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Bergers G and Benjamin LE: Tumorigenesis
and the angiogenic switch. Nat Rev Cancer. 3:401–410. 2003.
View Article : Google Scholar
|
|
95
|
Folkins C, Man S, Xu P, et al: Anticancer
therapies combining antiangiogenic and tumor cell cytotoxic effects
reduce the tumor stem-like cell fraction in glioma xenograft
tumors. Cancer Res. 67:3560–3564. 2007. View Article : Google Scholar
|
|
96
|
Zhou S, Ye W, Zhang M and Liang J: The
effects of nrf2 on tumor angiogenesis: a review of the possible
mechanisms of action. Crit Rev Eukaryot Gene Expr. 22:149–160.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Ashino T, Yamamoto M, Yoshida T and
Numazawa S: Redox-sensitive transcription factor Nrf2 regulates
vascular smooth muscle cell migration and neointimal hyperplasia.
Arterioscler Thromb Vasc Biol. 33:760–768. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Kaur B, Khwaja FW, Severson EA, et al:
Hypoxia and the hypoxia-inducible-factor pathway in glioma growth
and angiogenesis. Neuro Oncol. 7:134–153. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Kweider N, Fragoulis A, Rosen C, et al:
Interplay between vascular endothelial growth factor (VEGF) and
nuclear factor erythroid 2-related factor-2 (Nrf2): implications
for preeclampsia. J Biol Chem. 286:42863–42872. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Kim TH, Hur EG, Kang SJ, et al: NRF2
blockade suppresses colon tumor angiogenesis by inhibiting
hypoxia-induced activation of HIF-1α. Cancer Res. 71:2260–2275.
2011.PubMed/NCBI
|
|
101
|
Ji RC: Hypoxia and lymphangiogenesis in
tumor microenvironment and metastasis. Cancer Lett. 346:6–16. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Emara M and Allalunis-Turner J: Effect of
hypoxia on angiogenesis related factors in glioblastoma cells.
Oncol Rep. 31:1947–1953. 2014.PubMed/NCBI
|
|
103
|
Marampon F, Gravina GL, Zani BM, et al:
Hypoxia sustains glioblastoma radioresistance through
ERKs/DNA-PKcs/HIF-1α functional interplay. Int J Oncol.
44:2121–2131. 2014.PubMed/NCBI
|
|
104
|
Ji X, Wang H, Zhu J, et al: Knockdown of
Nrf2 suppresses glioblastoma angiogenesis by inhibiting
hypoxia-induced activation of HIF-1α. Int J Cancer. 135:574–584.
2014.PubMed/NCBI
|
|
105
|
Ji X, Wang H, Zhu J, et al: Correlation of
Nrf2 and HIF-1α in glioblastoma and their relationships to
clinicopathologic features and survival. Neurol Res. 35:1044–1050.
2013.
|
|
106
|
Shen H, Yang Y, Xia S, et al: Blockage of
Nrf2 suppresses the migration and invasion of esophageal squamous
cell carcinoma cells in hypoxic microenvironment. Dis Esophagus.
Sep 13–2013.(Epub ahead of print).
|
|
107
|
Lee C, Park GH and Jang JH: Cellular
antioxidant adaptive survival response to 6-hydroxydopamine-induced
nitrosative cell death in C6 glioma cells. Toxicology. 283:118–128.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Meisen WH and Kaur B: How can we trick the
immune system into overcoming the detrimental effects of oncolytic
viral therapy to treat glioblastoma? Expert Rev Neurother.
13:341–343. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Al-Huseini LM, Aw Yeang HX, Sethu S, et
al: Nuclear factor-erythroid 2 (NF-E2) p45-related factor-2 (Nrf2)
modulates dendritic cell immune function through regulation of p38
MAPK-cAMP-responsive element binding protein/activating
transcription factor 1 signaling. J Biol Chem. 288:22281–22288.
2013. View Article : Google Scholar
|
|
110
|
Thimmulappa RK, Lee H, Rangasamy T, et al:
Nrf2 is a critical regulator of the innate immune response and
survival during experimental sepsis. J Clin Invest. 116:984–995.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Rockwell CE, Zhang M, Fields PE and
Klaassen CD: Th2 skewing by activation of Nrf2 in CD4+ T
cells. J Immunol. 188:1630–1637. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Foresti R, Bains SK, Pitchumony TS, et al:
Small molecule activators of the Nrf2-HO-1 antioxidant axis
modulate heme metabolism and inflammation in BV2 microglia cells.
Pharmacol Res. 76:132–148. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Limonciel A and Jennings P: A review of
the evidence that ochratoxin A is an Nrf2 inhibitor: implications
for nephrotoxicity and renal carcinogenicity. Toxins. 6:371–379.
2014. View Article : Google Scholar : PubMed/NCBI
|