Introduction
Esophageal squamous cell carcinoma (ESCC) is one of
the most refractory malignant diseases worldwide, and more than 50%
of cases occur in China. Increasing rates of morbidity and
mortality of ESCC have been reported in recent years (1). Current treatment generally employs
surgical resection combined with chemotherapy and radiotherapy for
advanced ESCC. However, both the high recurrence rate and strong
systemic toxicity of the anticancer treatments led us to
investigate new agents to improve the therapeutic effect (2).
In recent years, plant food-derived phytochemical
anticancer strategies have been proposed to control various types
of carcinoma (3). Diallyl disulfide
(DADS), CH2=CH-CH2-SS-CH2CH=CH2, is one of the fat-soluble sulfur
compounds obtained from crushed garlic and represents 40–60% of
garlic essential oil (4). DADS has
a wide variety of biological activities including antifungal
(5), antibacterial (6), antiviral (7), antioxidant (8), antiplatelet and antithrombotic
properties (9). Moreover, DADS has
gained increasing attention due to its protective effects of cancer
development and anticancer effects against different types of
malignancies (10–13). Notably, the apoptotic effects of
DADS were even superior to those of chemotherapy agents such as
5-Fu and CTX in human breast cancer cells (14).
Apoptosis is an ordered and orchestrated cellular
process that occurs in physiological and pathological conditions,
which plays an important role in the treatment of cancer (15–17).
The exact mechanism of DADS on ESCC is still unclear. The ability
of DADS to control tumor growth may be due to its capacity to
interfere different pathways presented in the cell.
In the present study, we characterized the effects
of DADS on ESCC cell line ECA109. Our results support the use of
DADS as a potent anticancer drug by disrupting cell viability,
arresting G2/M cell cycle arrest and inducing apoptosis of ECA109
cells. These data suggest that DADS may be a suitable candidate for
the treatment of ESCC in the future.
Materials and methods
Materials
ECA109 human ESCC cell line and L02 human normal
liver cell line were purchased from the Chinese Academy of Shanghai
Institute of Cell Biology. DADS, dimethyl sulfoxide (DMSO),
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT),
bovine serum albumin (BSA), propidium iodide (PI), RNase A and
phosphate-buffered saline (PBS) were purchased from Sigma Aldrich
(USA). Fetal bovine serum (FBS), RPMI-1640 medium and 0.25% trypsin
digest were purchased from Hyclone Co. (USA). TRIzol reagent was
purchased from Invitrogen (USA). Annexin V conjugated to FITC
(Annexin V-FITC) apoptosis kit was purchased from Roche Technology
Co. (USA). PrimeScript™ RT Master Mix kit, SYBR® Premix
Ex Taq II were purchased from Takara Technology Company (Japan).
Enhanced chemiluminescence (ECL) kit was purchased from Amersham
Life Sciences (UK). Antibodies to ERK1/2, phospho-ERK1/2
(p-ERK1/2), MEK1, phospho-MEK1 (p-MEK1), and caspase-3, cleaved
caspase-3, cyclin B1, cdc2, phospho-cdc2 (p-cdc2), cdc25c, and PCNA
were purchased from Cell Signaling Technology Co. (USA). β-actin
antibody was purchased from Santa Cruz Biotechnology (USA).
Horseradish peroxidase (HRP)-coupled goat anti-mouse IgG and
anti-rabbit IgG (secondary antibody) were purchased from Santa Cruz
Biotechnology (USA).
Cell culture
Human ECA109 cell line and human L02 cell line were
cultured in RPMI-1640 supplemented with 10% FBS, 100 U/ml
penicillin G and 100 μg/ml streptomycin sulfate, at 37°C with 5%
CO2. The medium was replaced every other day. After
reaching 70–80% confluency, cells were digested with 0.25% trypsin
and subcultured. Exponentially growing cells were used for
assays.
Cell viability assay
Cell viability was determined by using MTT assay.
DADS was dissolved in PBS with less than 0.01% DMSO and prepared at
a concentration of 1,000 μg/ml. PBS with less than 0.01% DMSO was
used as the negative control. ECA109 and L02 cells were seeded in
96-well plates at 1×104 cells/well and incubated with
DADS (10–60 μg/ml). After incubation, 20 μl MTT (5 mg/ml) was added
to each well. Four hours later, the blue crystals were solubilized
with 150 μl DMSO. The absorbance was measured at 570 nm. The
following formula was used for the calculation: Cell viability
ratio = 1 - [(A value of the control - A value of the experimental
samples)/A value of the control] × 100%. Half maximal inhibitory
concentration (IC50) values were calculated by
SPSS® software. Each assay was performed in 5
replicates.
Cell cycle assay
A total of 5×105 ECA109 cells/plate in 6
cm plates were incubated with different concentrations of DADS (20,
40, 60 μg/ml). PBS with less than 0.01% DMSO was used as control.
Following 24 h treatment, cells were fixed with ice-cold 70%
ethanol overnight at −20°C, then stained with 5 μl PI at room
temperature for 30 min in the dark. Finally, cell cycle analysis
was performed using Flow Cytometer, and data were analyzed with
Cell Quest Pro® software. The experiments were repeated
three times.
Apoptosis assay
The apoptotic ratios of cells were determined with
the Annexin V-FITC apoptosis detection kit. A total of
5×105 ECA109 cells/plate in 6 cm plates were incubated
with different concentrations of DADS (20, 40, 60 μg/ml). Cells
treated with PBS with less than 0.01% DMSO were used as control.
The cells were trypsinized, washed twice with PBS, resuspended in
100 μl of binding buffer at a concentration of 1×106
cells/ml, incubated with 5 μl Annexin V-FITC and 10 μl PI for 15
min at room temperature in the dark, and added with 400 μl binding
buffer. Apoptosis analysis was performed by Flow Cytometer, and
data were analyzed with Cell Quest Pro® software. The
experiments were repeated at least three times.
Real-time polymerase chain reaction
(PCR)
Cells were treated with serially diluted
concentrations of DADS (20, 40, 60 μg/ml) and harvested for 24 h.
PBS with less than 0.01% DMSO was used as control. Total RNA was
extracted by the TRIzol method and the quantity of RNA was assessed
by spectrophotometry. cDNA was obtained by reverse transcription
with 1 μg total RNA using PrimeScript™ RT Master Mix kit. Real-time
PCR was performed with the SYBR-Premix Ex Taq™ II Perfect Real Time
kit. All reactions were performed on an ABI QPCR System. The
individual values were normalized to the β-actin control, and the
ratio of the relative expression levels was calculated. The
specific primers are shown in Table
I. The samples were run in triplicate.
 | Table IPrimers for real-time PCR
analysis. |
Table I
Primers for real-time PCR
analysis.
| Gene | Forward
sequence | Reverse
sequence |
|---|
| β-actin |
5′-TGGCACCCAGCACAATGAA-3′ |
5′-CTAAGTCATAGTCCGCCTAGAAGCA-3′ |
| p21 |
5′-AACATGTTGAGCTCTGGCATAGAAG-3′ |
5′-GCATGGGTTCTGACGGACA-3′ |
| p53 |
5′-TCAGCATCTTATCCGAGTGGAA-3′ |
5′-TGTAGTGGATGGTGGTACAGTCA-3′ |
| Bcl-2 |
5′-GTGTGGAGAGCGTCAACC-3′ |
5′-CTTCAGAGACAGCCAGGAG-3′ |
| Bax |
5′-ATGGGCTGGACATTGGACTTC-3′ |
5′-TGGTGAGTGAGGCGGTGAG-3′ |
Western blot assay
ECA109 cells were prepared by serially diluted
concentrations of DADS (20, 40, 60 μg/ml). Cells treated with PBS
with less than 0.01% DMSO were used as control. After incubation
for 24 h, cells were harvested to obtain protein concentrations by
RIPA lysis buffer. Proteins (50 μg) were separated by 8–12% sodium
dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE),
transferred to polyvinylidene difluoride (PVDF) membranes, then
probed with appropriate primary antibodies. HRP-conjugated goat
anti-mouse antibody or anti-rabbit antibody (1:10,000) was used as
the secondary antibody. Antigen-antibody complex signals were
visualized with BeyoECL Plus. The densitometric analysis was
performed by Image J® software. The experiments were
repeated three times.
The following primary antibodies were used:
anti-cyclin B1 (1:1,000), anti-cdc2 (1:1,000), anti-p-cdc2 (1:500),
anti-cdc25c (1:1,000), anti-ERK1/2 (1:1,000), anti-p-ERK1/2
(1:300), anti-caspase-3 (1:1,000), anti-cleaved caspase-3 (1:500),
anti-MEK1 (1:1,000), anti-p-MEK1 (1:500) and anti-β-actin (1:1,000)
antibodies.
Quantification and statistical
analysis
Quantitative data are expressed as the mean ± SD
from at least three independent experiments. The two-tailed
Student’s t-test was performed for paired samples, one-way analysis
of variance (ANOVA) or two-factor factorial ANOVA was used for
multiple groups. Results were considered to indicate a
statistically significant difference at the P<0.05 level.
Statistical analyses were performed by SPSS 17.0 statistical
software.
Results
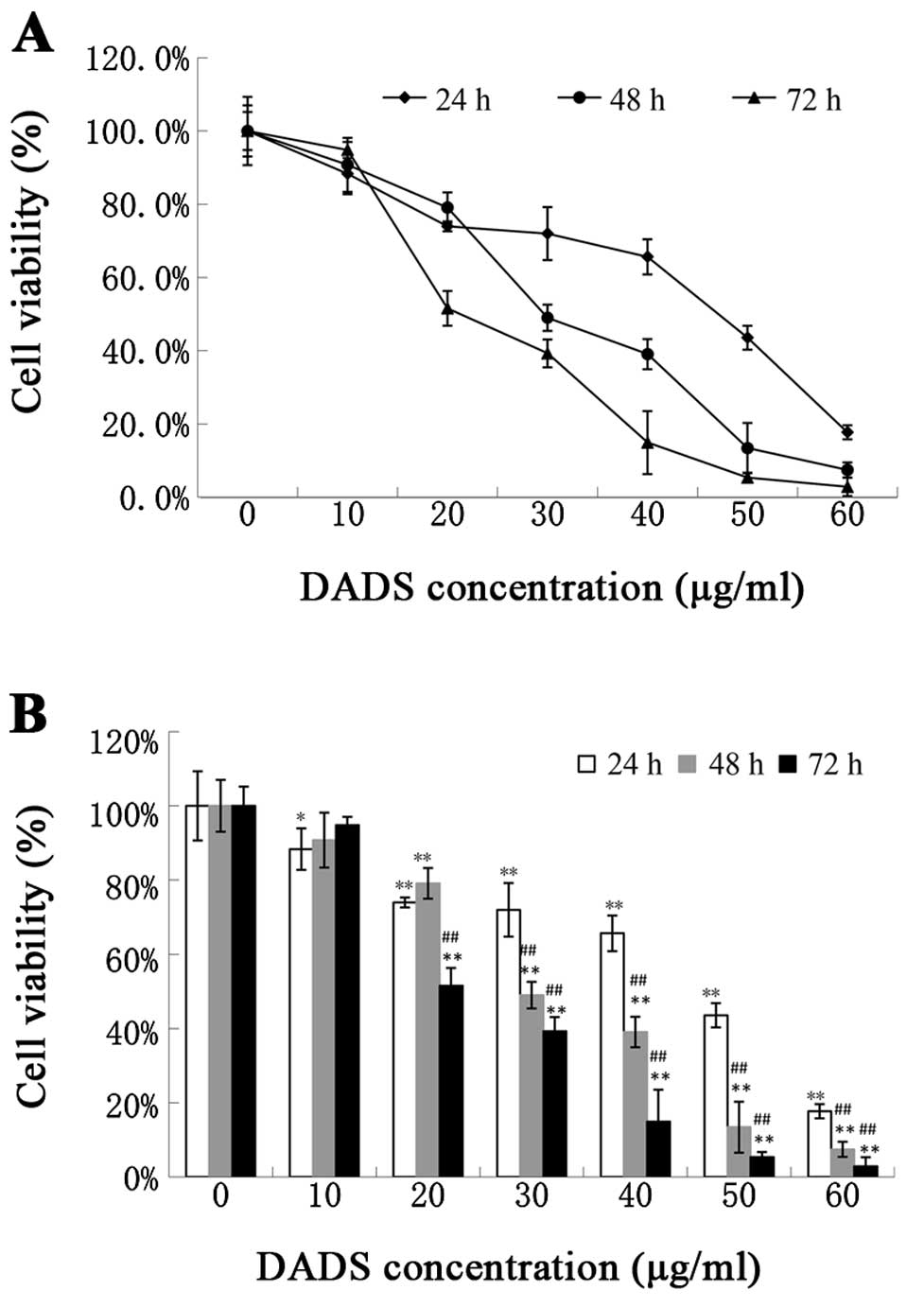
DADS inhibits cell viability
The MTT assay was used to detect the viability
effects of DADS at various concentrations (10–60 μg/ml) on ECA109
and L02 cells. Our data showed that DADS clearly inhibited cell
viability at the concentrations of 10–60 μg/ml following exposure
for 24 h (P<0.05, Fig. 1), 20–60
μg/ml for 48 and 72 h (P<0.01, Fig.
1) compared with the control group. The IC50 was
49.02±4.78, 33.14±5.02 and 22.74±4.05 μg/ml for 24, 48 and 72 h
DADS treatment respectively. The 72-h treatment group had an
apparent difference compared with the 24-h treatment group
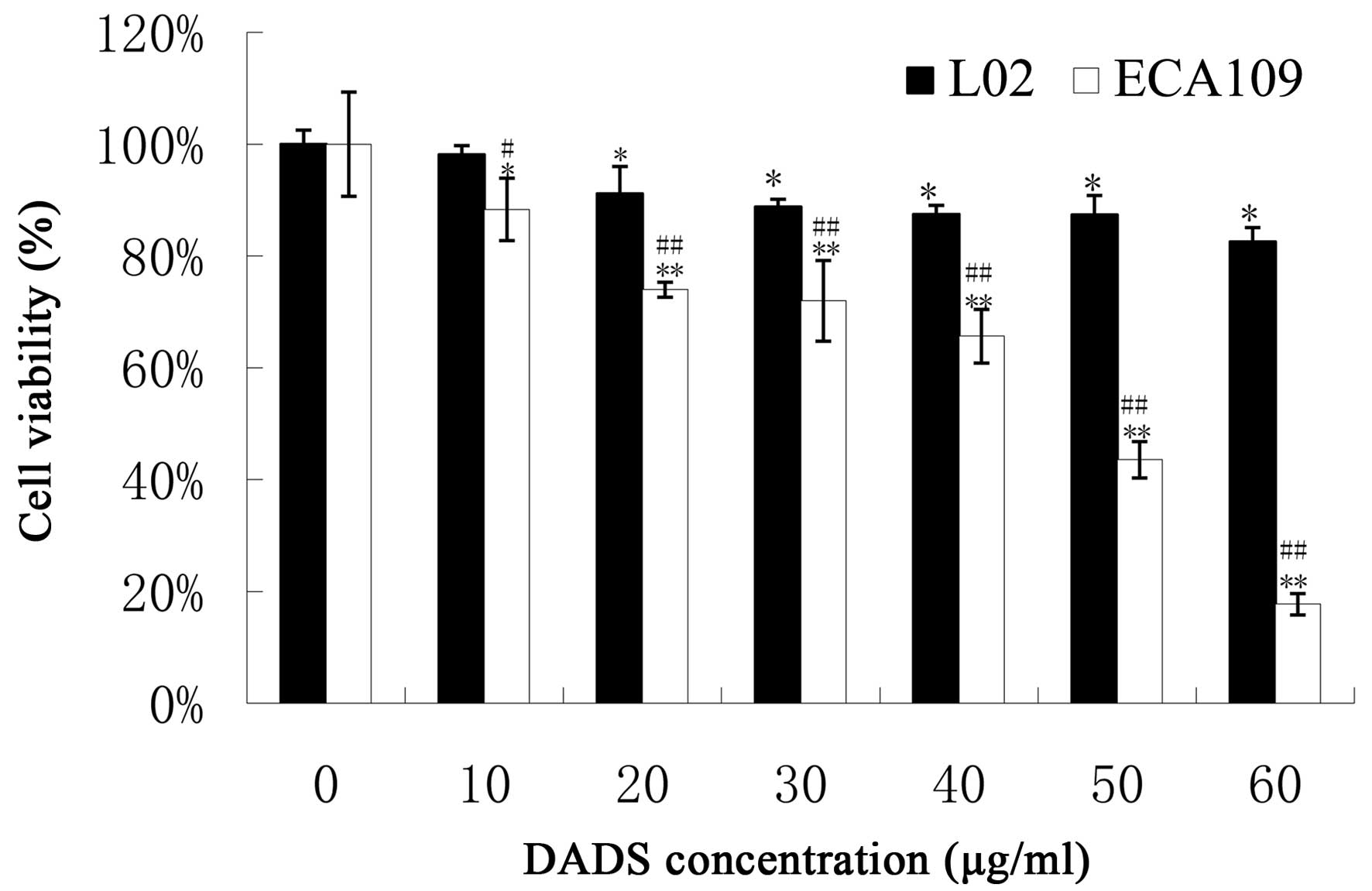
(P<0.05, Fig. 1). DADS had less
of an influence on L02 normal cells than on ECA109 esophageal
carcinoma cells (P<0.05, Fig.
2). These results indicate that DADS significantly inhibited
ECA109 cell viability in a dose- and time-dependent manner, and had
a lower effect on normal cells.
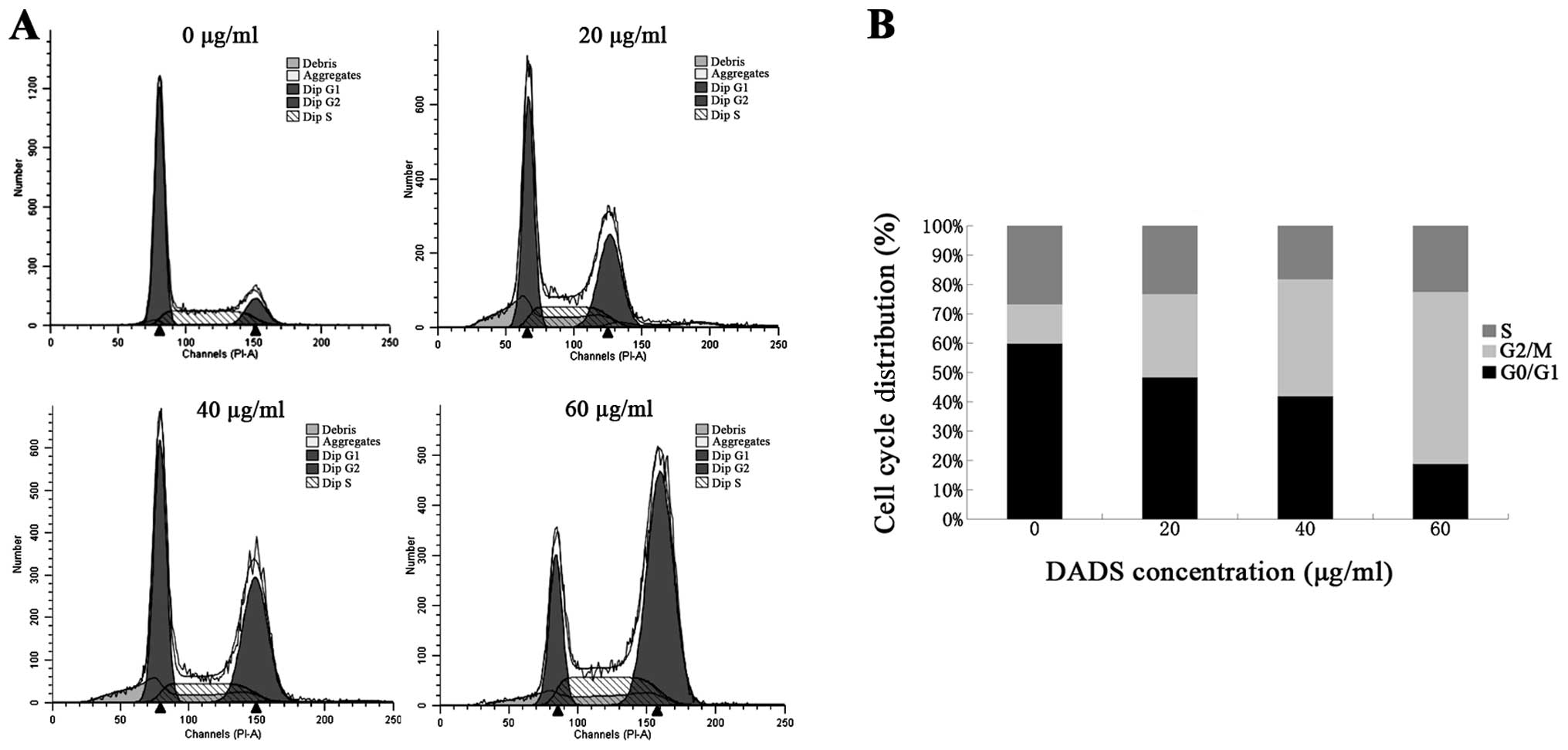
DADS induces G2/M phase cell cycle
arrest
Cell cycle distribution of ECA109 cells treated with
different doses of DADS for 24 h was analyzed by flow cytometry.
Results showed that the percentage of cells in the DADS groups,
particularly at the 20, 40, 60 μg/ml concentrations, significantly
increased at the G2/M phase in a dose-dependent manner compared
with the control group. The percentage of cells at the G2/M phase
increased from 13.43±1.25 to 58.65±3.79%, and decreased from
60.01±3.13 to 18.97±1.37% and from 26.56±2.02 to 22.38±2.56%, at
the G0/G1 and S phase, respectively (Table II). These results suggest that DADS
induced cell cycle arrest at the G2/M phase (Fig. 3), which may be one of the reasons
for the viability inhibition and apoptosis induction of ECA109
cells.
| Table IIEffect of DADS on the cell cycle of
ECA109 cells. |
Table II
Effect of DADS on the cell cycle of
ECA109 cells.
| Groups | N | G0/G1 | G2/M | S |
|---|
| Control | 3 | 60.01±3.13 | 13.43±1.25 | 26.56±2.02 |
| 20 μg/ml | 3 | 48.51±2.65a | 28.43±2.56a | 23.06±2.45b |
| 40 μg/ml | 3 | 42.08±1.12a | 39.90±2.06a | 18.02±1.24b |
| 60 μg/ml | 3 | 18.97±1.37a | 58.65±3.79a | 22.38±2.56b |
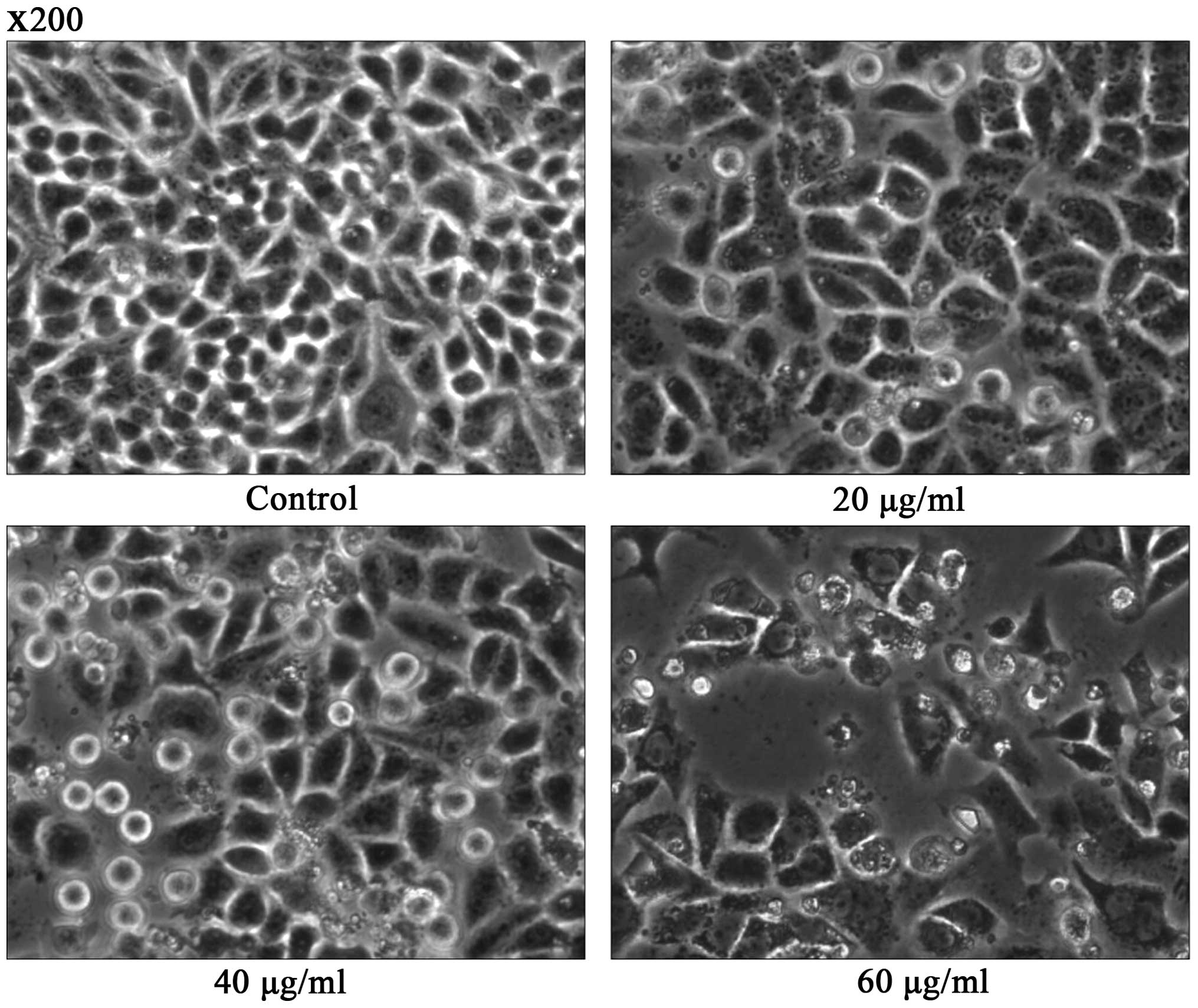
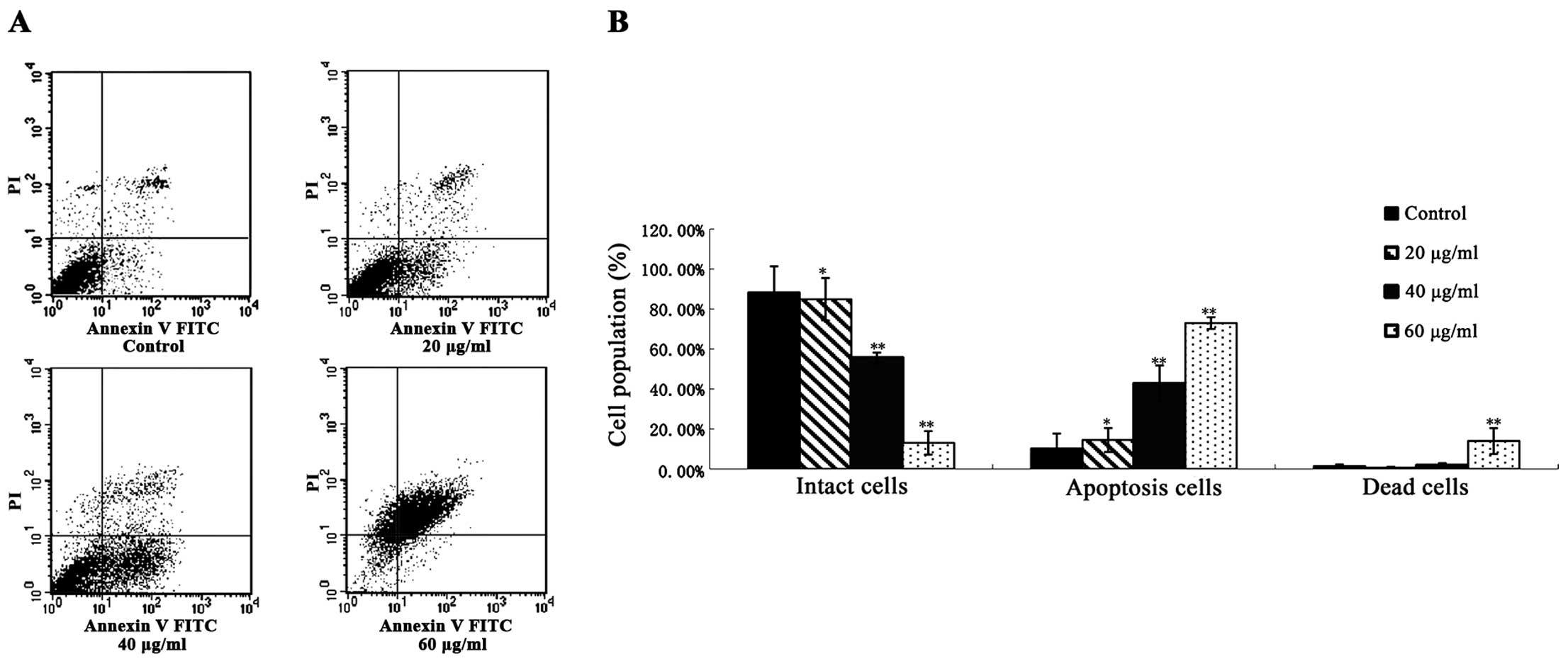
DADS induces cell apoptosis
ECA109 cells were examined by phase contrast
microscopy after incubation with DADS at different concentrations
(20, 40, 60 μg/ml) for 24 h. Cells treated with PBS were used as
control. The control group cells showed a typical polygonal and
intact appearance, whereas the DADS-treated cells displayed
dose-dependent changes in cell shape, such as membrane blebbing,
formation of apoptotic bodies, cellular shrinkage, poor adherence
and floating shapes (Fig. 4).
The suppression of cancer cell growth correlates
with apoptosis. We explored the apoptosis rate of ECA109 cells
impacted with different concentrations of DADS for 24 h with
Annexin V-FITC and PI staining observed by flow cytometry. The rate
of apoptosis in the control and DADS groups (20, 40 and 60 μg/ml)
for 24 h was 10.26±1.05, 14.48±0.99, 42.68±2.08 and 72.96±3.43%
respectively (P<0.05, Fig. 5).
These results demonstrate that DADS induced apoptosis of ECA109
cells in a dose-dependent manner.
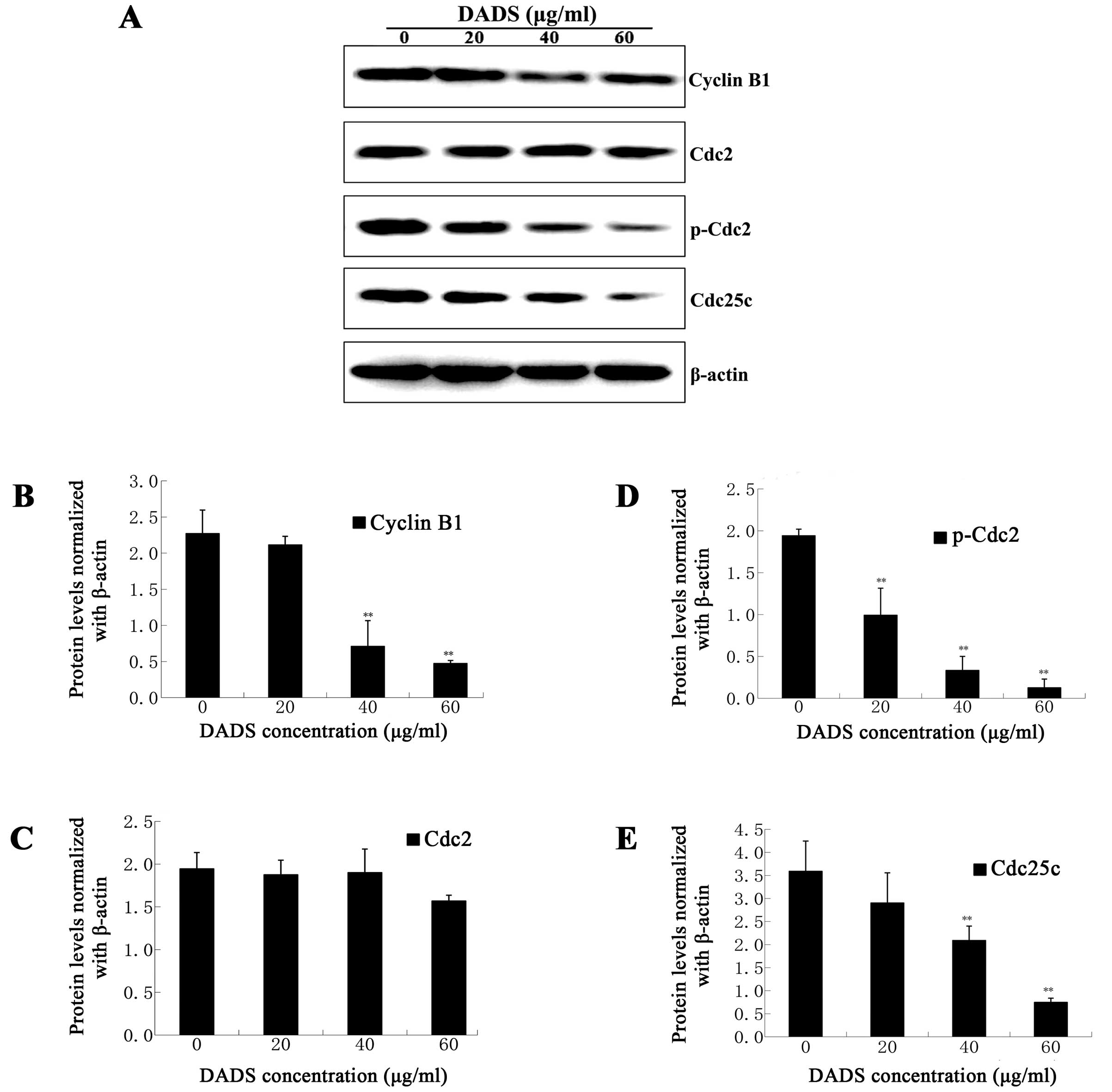
DADS inhibits G2/M phase-associated
protein expression of cyclin B1, cdc2, p-cdc2 and cdc25c
Since DADS induced G2/M phase arrest in ECA109
cells, we evaluated the expression of proteins that regulate the
G2/M phase transition by western blot assay. Cyclin B1, cdc2, and
cdc25c are important proteins related to the G2/M phase. G2/M phase
is controlled by a complex formed cyclin B1 and cdc2, and the
complex is regulated by cdc25c. Our data showed that cdc2 protein
levels had no apparent change (P>0.05, Fig. 6), and the protein levels of cyclin
B1 and cdc25c decreased in a dose-dependent manner with significant
inhibition occurring at DADS concentrations of 40 and 60 μg/ml
(P<0.01, Fig. 6). Moreover, the
protein levels of p-cdc2 decreased in a dose-dependent manner, with
significant inhibition occurring at DADS concentrations of 20, 40
and 60 μg/ml (P<0.01, Fig. 6).
These results indicate that DADS reduced the expression of cyclin
B1, p-cdc2, and cdc25c to induce G2/M phase arrest of ECA109
cells.
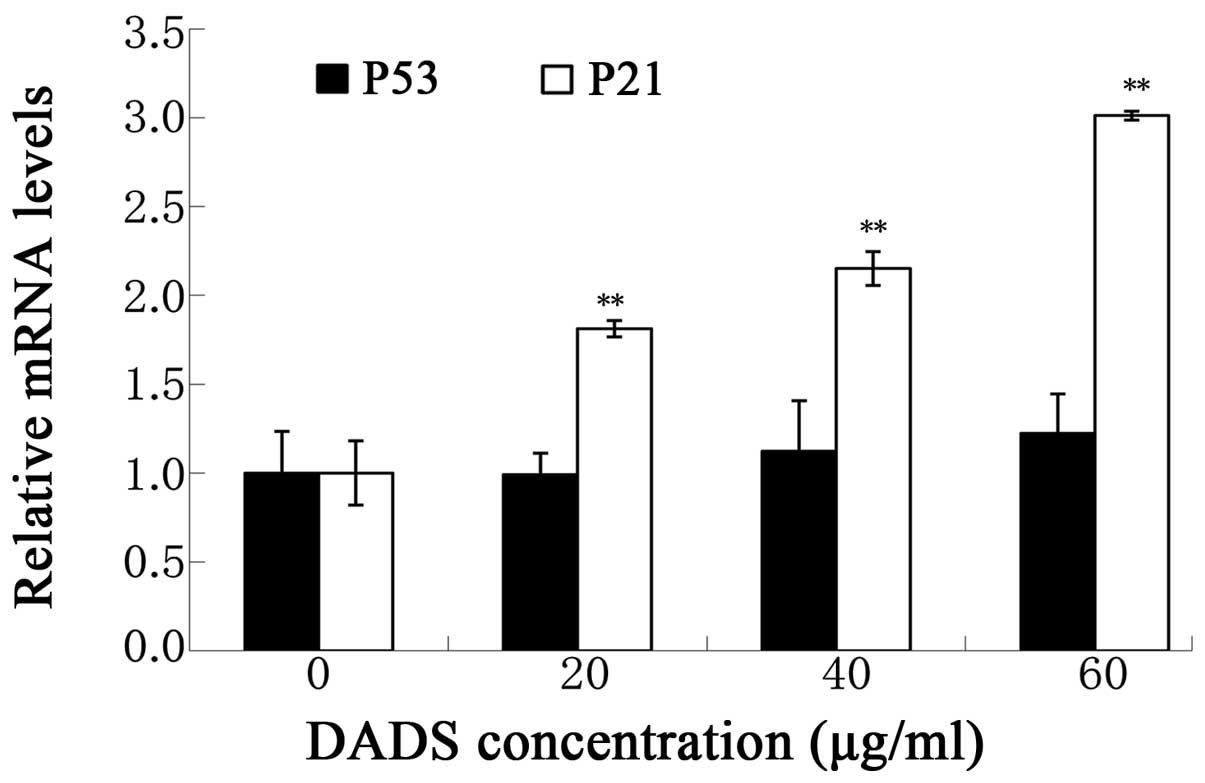
Effects of DADS on the p53/p21
pathway
The p53/p21 pathway is involved in the cell
apoptosis progression and plays an important role in the cell cycle
arrest of G2/M phase. Therefore, we used real-time PCR assay to
investigate the G2/M phase relative molecular pathway. Our data
showed that DADS (20, 40 and 60 μg/ml) upregulated mRNA levels of
p21 (P<0.01, Fig. 7), p53
(P<0.05, Fig. 7) compared with
the relative control. The upregulated levels of p21 mRNA expression
were accompanied by the increase of G2/M arrest. Based on these
results, we suggest that DADS-increased G2/M phase arrest and
apoptosis rate of ECA109 cells might be partly affected by the
p53/p21 signaling pathway.
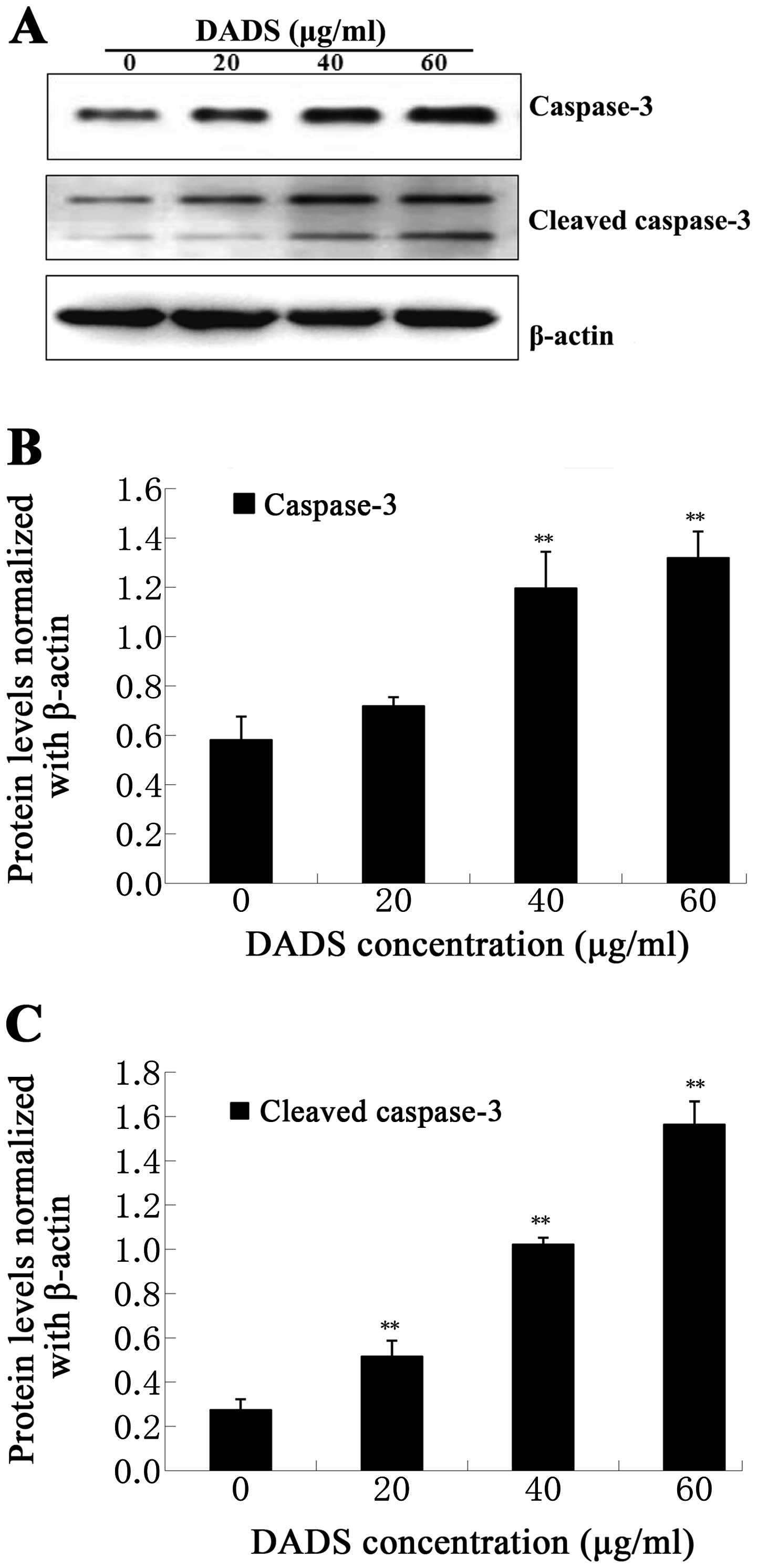
Protein expression of caspase-3 and
cleaved caspase-3 by western blot analysis
As the imbalance of apoptotic signals is closely
related to the initiation of the apoptotic program, we analyzed the
protein expression levels of caspase-3 and cleaved caspase-3
proteins by western blotting. These proteins were blotted against
the corresponding antibodies as mentioned above in the ECA109
cells, following 24 h of treatment with DADS (20, 40 and 60 μg/ml).
Our results showed that the protein levels of caspase-3 increased
at DADS doses of 40 and 60 μg/ml (P<0.01, Fig. 8), and the expression levels of
cleaved caspase-3 fragment (17 kDa, 19 kDa) were clearly
upregulated in a dose-dependent manner, particularly at 20, 40 and
60 μg/ml doses (P<0.01, Fig. 8).
Therefore, this study suggests that DADS-induced apoptosis of
ECA109 cells is mediated by effector caspase-3.
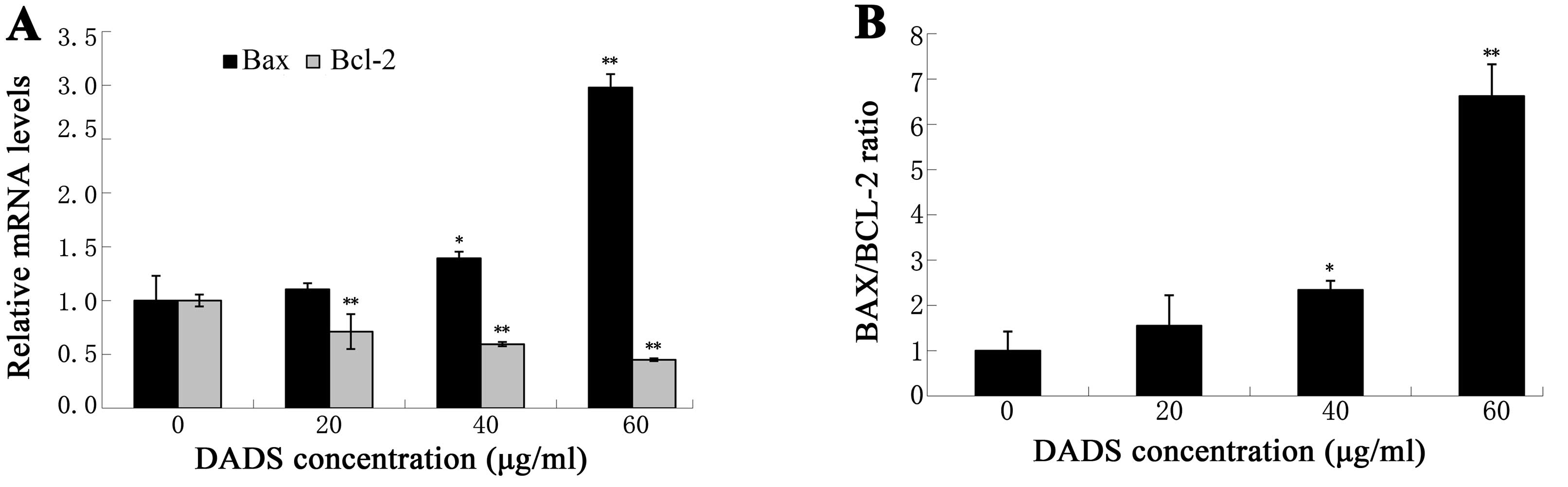
Effects of DADS on Bax, Bcl-2 expression
levels and Bax/Bcl-2 ratio
DADS-induced apoptosis of ECA109 cells prompted an
examination of certain apoptosis regulatory proteins, such as Bax
and Bcl-2. The expression levels of Bax mRNA significantly
increased in 40 and 60 μg/ml (P<0.05, Fig. 9), whereas the expression levels of
Bcl-2 mRNA were clearly decreased in 20, 40 and 60 μg/ml
(P<0.01, Fig. 9). The Bax/Bcl-2
ratio was significantly increased in the presence of DADS at
concentrations of 40 and 60 μg/ml (P<0.05, Fig. 9). Collectively, the data suggest
that DADS induced apoptosis through upregulation of Bax mRNA,
downregulation of Bcl-2 mRNA and a shift of Bax/Bcl-2 ratio in a
dose-dependent manner.
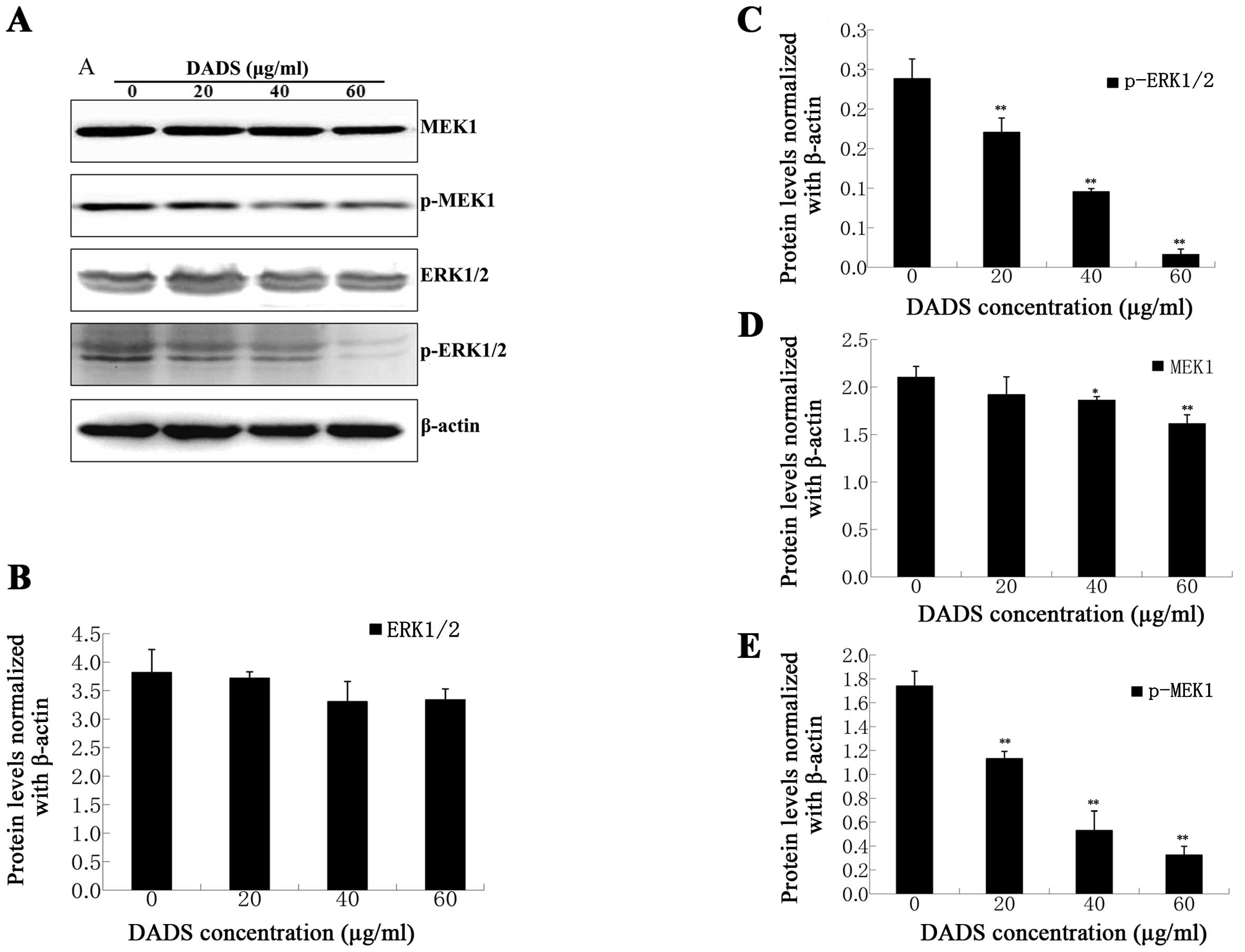
Effects of DADS on the MEK-ERK
pathway
Western blot analysis was used to evaluate the
effect of DADS on MEK1, ERK1/2 and their phosphorylation levels in
ECA109 cells. Our data showed that the protein levels of MEK1 and
ERK1/2 had no obvious change, whereas the expression levels of
p-MEK1 and p-ERK1/2 were clearly decreased in the 20, 40 and 60
μg/ml DADS groups compared with the control (P<0.01, Fig. 10). As shown in Fig. 10, DADS inhibited p-MEK1 and
p-ERK1/2 in a dose-dependent manner. Based on these results, we
suggest that DADS-related apoptosis in ECA109 cells might be
influenced by the MEK-ERK signaling pathway.
Discussion
Diallyl disulfide (DADS) is a lipid-soluble organic
compound from garlic. Scientific investigations have shown that
DADS can reduce the risk of cardiovascular disease and diabetes,
stimulate the immune system, protect against infections, and show
significant protection against different types of malignancies
(4,18).
By MTT assay, DADS showed dose- and time-dependent
anti-viability effects on ESCC cell lines (Figs. 1 and 2). Moreover, we further demonstrated that
DADS had much lower cytotoxicity in L02 normal liver cells compared
with ECA109 carcinoma cells (P<0.01). The viability inhibitory
properties of DADS may be attributable to its induction of cell
cycle arrest and apoptosis cell death, which is consistent with
previous findings (10–13).
G2/M phase cell cycle arrest can reduce
proliferation and induce apoptosis by inhibiting the segregation of
damaged chromosomes during mitosis (15). DADS has been demonstrated to induce
G2/M cell cycle arrest in several cancer cell lines such as colon
cancer (11), breast cancer
(12) and human gastric cancer
cells (13). In the present study,
we demonstrated that the percentage of ECA109 cells in DADS
treatment groups significantly increased at the G2/M phase in a
dose-dependent manner (Fig. 3). The
G2/M phase arrest induced by DADS showed an increase in the number
of ECA109 cells from 13.43 to 58.65%, as well as apoptotic cells
from 10.2 to 72.96% (Table II),
suggesting that the block in the G2/M phase results in triggering
the apoptotic program.
Apoptosis is a morphologically and biochemically
distinct mode of cell death that plays major roles during
carcinogenesis, cancer treatment and toxic cell killing (19). Apoptosis induction involves
different apoptotic genes and enzymes depending on the cell type of
different tumors (16). DADS has
the ability to induce apoptosis of human tumor cells including
those of colon, gastric, prostate and breast origin (10,11,18,20–22).
Our study confirmed the apoptotic effects of DADS on ECA109 cells
by using several methods. First, under light microscope, we
observed that morphological changes of apoptosis were induced by
DADS. Typical morphological characteristics of apoptotic ECA109
cells, such as membrane blebbing, cell shrinkage and formation of
apoptotic bodies, were observed in the 40, 60 μg/ml DADS dose group
(Fig. 4). Second, we detected the
apoptosis rate by the double-staining of Annexin V-FITC and PI. Our
flow cytometry analysis showed that DADS promoted ECA109 apoptosis
in a dose-dependent manner (Fig.
5).
DADS controlled tumor growth and death. Although the
mechanism for DADS-induced apoptosis has been studied in several
cellular systems, it remains a controversial issue. As the balance
and interplay of multiple signaling networks involved in various
signaling contexts, it is not surprising that DADS regulation is
complex. Therefore, the results of the present study indicate the
noticeable changes in molecular signals of ECA109 cells affected by
DADS.
G2/M phase progression is regulated by cdc2 kinases
and cyclin B1. In addition, cdc2 activation depends on the
dephosphorylation of Tyr15 by cdc25c. Otherwise, p53 and the
p53-responsive gene, p21, can suppress cyclin B1 and cdc2
expression by inhibiting either cdc2 kinase activity or blocking
the interaction of cyclin B1-cdc2 complexes with their substrates,
leading to G2/M-phase cell cycle arrest (23–27).
Furthermore, the MEK-ERK pathway has crosstalk with p53/p21, and
Bax is a key target of the p53 transcription process in aspects of
cell apoptosis (26). In the case
of our study, the data showed that DADS downregulated cyclin B1,
cdc2, p-cdc2 and cdc25c expression (Fig. 7) and upregulated p21 expression of
ECA109 cells in a dose-dependent manner (Fig. 6). Thus, we found that upregulation
of the p53/p21 pathway could be due to blocking the interaction of
cyclin B1-cdc2 complexes, and downregulation of cdc25c consequently
reduced the activity of cdc2. Therefore, we suggest that
alterations in G2/M phase-associated protein levels and p53/p21
signaling pathway may be the mechanism underlying the growth
inhibition and G2/M phase arrest.
Bax is a key target of the p53 transcription process
in aspects of cell apoptosis (26).
Anti-apoptotic protein Bcl-2 could bind to and inactivate Bax and
other pro-apoptotic proteins to inhibit apoptosis. Therefore,
Bax/Bcl-2 ratio plays a pivotal role in apoptosis (28). Moreover, caspase-3 plays an
important role as the central effector for initiation of apoptosis
(16). Our results suggest that
DADS promoted cell death by downregulating Bcl-2 mRNA expression
and upregulating the Bax expression in a dose-dependent manner,
thus increasing the Bax/Bcl-2 ratio (Fig. 9), leading to a pro-apoptotic process
via caspase-3 in ECA109 cells (Fig.
8). Based on these observations, the data indicate that the
increase of Bax/Bcl-2 and activation of caspase-3 are involved in
the DADS-induced apoptosis mechanism.
The sequential phosphorylation of MAPK kinase1/2
(MEK1/2) makes the activation of extracellular signal-regulated
kinase1/2 (ERK1/2). Then, ERK1/2 activation leads to
phosphorylation of a variety of transcription factors and results
in proliferation and differentiation, protecting cells against
apoptosis (29). MEK-ERK signaling
cascade may be affected by the activation of the p53/p21 pathway
(26). Although DADS was reported
to rapidly and potently inhibit the phosphorylation of ERK1/2 to
induce apoptosis in human leukemia cell line HL-60 (21), other studies showed that DADS
activated ERK1/2 in human non-small cell lung carcinoma H1299 cell
death (20). Our results showed
that both p-MEK1 and p-ERK1/2 were inhibited following DADS
treatment in a dose-dependent manner (Fig. 10), suggesting that the MEK-ERK
pathway plays a role in maintaining the regulation of apoptosis by
DADS in ECA109 cells.
In summary, the present study confirmed that DADS
inhibits ESCC cell viability with only a slight effect on normal
cells. To the best of our knowledge, this is the first report to
identify mechanisms of the antitumor properties of DADS in human
ESCC cells. DADS has been shown to arrest cancer cells at the G2/M
phase via the modulation of cell-cycle related proteins, which is
associated with the reduction of cyclin B1, cdc2, p-cdc2, cdc25c
expression. Furthermore, DADS controlled cellular apoptosis by
activating caspase-3, upregulating Bax/Bcl-2 ratio and
downregulating the MEK-ERK signaling pathway. Moreover, activation
of the p53/p21 pathway is involved in the process of inhibiting
cell differentiation, arresting G2/M phase and inducing apoptosis.
In brief, DADS regulates ESCC cells via multiple networks involved
in various signaling contexts. Therefore, this study suggests that
DADS may be a promising anticancer therapeutic for ESCC in the near
future.
Acknowledgements
This study was supported by funds from the Science
and Technology Program of Shaanxi Province (nos. 2010K01-141 and
2011K13-02-05), the Important Clinic Project of the Chinese
Ministry of Health (no. 2007353). This study was also supported by
the Office of Oncology Research (Zong-Fang Li). The authors are
grateful to Dr Guleng B for his technical assistance.
References
|
1
|
Cui X, Zhao Z, Liu D, et al: Inactivation
of miR-34a by aberrant CpG methylation in Kazakh patients with
esophageal carcinoma. J Exp Clin Cancer Res. 33:20–33. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Orditura M, Galizia G, Fabozzi A, et al:
Preoperative treatment of locally advanced esophageal carcinoma.
Int J Oncol. 43:1745–1753. 2013.PubMed/NCBI
|
|
3
|
Matés JM, Segura JA, Alonso FJ and Márquez
J: Anticancer antioxidant regulatory functions of phytochemicals.
Current Medicinal Chemistry. 18:2315–2338. 2011.PubMed/NCBI
|
|
4
|
Zhou Y, Su J, Shi L, Liao Q and Su Q: DADS
downregulates the Rac1-ROCK1/PAK1-LIMK1-ADF/cofilin signaling
pathway, inhibiting cell migration and invasion. Oncol Rep.
29:605–612. 2013.PubMed/NCBI
|
|
5
|
Alam M, Zubair S, Farazuddin M, et al:
Development, characterization and efficacy of niosomal diallyl
disulfide in treatment of disseminated murine candidiasis.
Nanomedicine. 9:247–256. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Maldonado PD, Chánez-Cárdenas ME and
Pedraza-Chaverrí J: Aged garlic extract, garlic powder extract,
S-allylcysteine, diallyl sulfide and diallyl disulfide do not
interfere with the antibiotic activity of gentamicin. Phytother
Res. 19:252–254. 2005. View
Article : Google Scholar
|
|
7
|
Lissiman E, Bhasale AL and Cohen M: Garlic
for the common cold. Cochrane Database Syst Rev.
3:CD0062062012.
|
|
8
|
Lee IC, Kim SH, Baek HS, et al: The
involvement of Nrf2 in the protective effects of diallyl disulfide
on carbon tetrachloride-induced hepatic oxidative damage and
inflammatory response in rats. Food Chem Toxicol. 63:174–185. 2014.
View Article : Google Scholar
|
|
9
|
Truong D, Hindmarsh W and O’Brien PJ: The
molecular mechanisms of diallyl disulfide and diallyl sulfide
induced hepatocyte cytotoxicity. Chem Biol Interact. 180:79–88.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Arunkumar R, Sharmila G, Elumalai P, et
al: Effect of diallyl disulfide on insulin-like growth factor
signaling molecules involved in cell survival and proliferation of
human prostate cancer cells in vitro and in silico approach through
docking analysis. Phytomedicine. 19:912–923. 2012. View Article : Google Scholar
|
|
11
|
Song JD, Lee SK, Kim KM, et al: Molecular
mechanism of diallyl disulfide in cell cycle arrest and apoptosis
in HCT-116 colon cancer cells. J Biochem Mol Toxicol. 23:71–79.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nkrumah-Elie YM, Reuben JS, Hudson AM, et
al: The attenuation of early benzo (a) pyrene-induced carcinogenic
insults by diallyl disulfide (DADS) in MCF-10A cells. Nutr Cancer.
64:1112–1121. 2012. View Article : Google Scholar
|
|
13
|
Ling H, Wen L, Ji XX, et al: Growth
inhibitory effect and Chk1-dependent signaling involved in G2/M
arrest on human gastric cancer cells induced by diallyl disulfide.
Braz J Med Biol Res. 43:271–278. 2010. View Article : Google Scholar
|
|
14
|
Jun Z, Suzuki M, Xiao J, Wen J, Talbot SG,
Li GC and Xu M: Comparative effects of natural and synthetic
diallyl disulfide on apoptosis of human breast-cancer MCF-7 cells.
Biotechnol Appl Biochem. 52:113–119. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang XH, Zou ZQ, Xu CW, Shen YZ and Li D:
Myricetin induces G2/M phase arrest in HepG2 cells by inhibiting
the activity of the cyclin B/cdc2 complex. Mol Med Rep. 4:273–277.
2011.PubMed/NCBI
|
|
16
|
Sankari SL, Masthan KM, Babu NA,
Bhattacharjee T and Elumalai M: Apoptosis in cancer - an update.
Asian Pac J Cancer Prev. 13:4873–4878. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Czepukojc B, Baltes AK, Cerella C, et al:
Synthetic polysulfane derivatives induce cell cycle arrest and
apoptotic cell death in human hematopoietic cancer cells. Food Chem
Toxicol. 64:249–257. 2014. View Article : Google Scholar
|
|
18
|
Tang H, Kong Y, Guo J, et al: Diallyl
disulfide suppresses proliferation and induces apoptosis in human
gastric cancer through Wnt-1 signaling pathway by upregulation of
miR-200b and miR-22. Cancer Lett. 340:72–81. 2013. View Article : Google Scholar
|
|
19
|
Haneji T, Hirashima K, Teramachi J and
Morimoto H: Okadaic acid activates the PKR pathway and induces
apoptosis through PKR stimulation in MG63 osteoblast-like cells.
Int J Oncol. 42:1904–1910. 2013.PubMed/NCBI
|
|
20
|
Hui C, Jun W, Ya LN and Ming X: Effect of
Allium sativum (garlic) diallyl disulfide (DADS) on human
non-small cell lung carcinoma H1299 cells. Trop Biomed. 25:37–45.
2008.
|
|
21
|
Tan H, Ling H, He J, Yi L, Zhou J, Lin M
and Su Q: Inhibition of ERK and activation of p38 are involved in
diallyl disulfide induced apoptosis of leukemia HL-60 cells. Arch
Pharm Res. 31:786–793. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Altonsy MO, Habib TN and Andrews SC:
Diallyl disulfide-induced apoptosis in a breast-cancer cell line
(MCF-7) may be caused by inhibition of histone deacetylation. Nutr
Cancer. 64:1251–1260. 2012. View Article : Google Scholar
|
|
23
|
Duong HQ, Hwang JS, Kim HJ, Seong YS and
Bae I: BML-275, an AMPK inhibitor, induces DNA damage, G2/M arrest
and apoptosis in human pancreatic cancer cells. Int J Oncol.
41:2227–2236. 2012.PubMed/NCBI
|
|
24
|
Paul P, Rajendran SK, Peuhu E, Alshatwi
AA, Akbarsha MA, Hietanen S and Eriksson JE: Novel action modality
of the diterpenoid anisomelic acid causes depletion of E6 and E7
viral oncoproteins in HPV-transformed cervical carcinoma cells.
Biochem Pharmacol. 89:171–184. 2014. View Article : Google Scholar
|
|
25
|
Zhang Z, Wang CZ, Du GJ, et al: Genistein
induces G2/M cell cycle arrest and apoptosis via ATM/p53-dependent
pathway in human colon cancer cells. Int J Oncol. 43:289–296.
2013.PubMed/NCBI
|
|
26
|
Li B, Zhao J, Wang CZ, Searle J, He TC,
Yuan CS and Du W: Ginsenoside Rh2 induces apoptosis and
paraptosis-like cell death in colorectal cancer cells through
activation of p53. Cancer Lett. 301:185–192. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ding L, Huang Y, Du Q, et al: TGEV
nucleocapsid protein induces cell cycle arrest and apoptosis
through activation of p53 signaling. Biochem Biophys Res Commun.
445:497–503. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Takahashi H, Chen MC, Pham H, et al:
Baicalein, a component of Scutellaria baicalensis, induces
apoptosis by Mcl-1 downregulation in human pancreatic cancer cells.
Biochim Biophys Acta. 1813:1465–1474. 2011.
|
|
29
|
Lv C, Sun W, Sun H, Wei S, Chen R, Wang B
and Huang C: Asperolide A, a marine-derived tetranorditerpenoid,
induces G2/M arrest in human NCI-H460 lung carcinoma cells, is
mediated by p53-p21 stabilization and modulated by Ras/Raf/MEK/ERK
signaling pathway. Mar Drugs. 11:316–331. 2013. View Article : Google Scholar
|