Introduction
Breast cancer is the leading cause of cancer-related
death in women and the third most common tumor worldwide (1). While significant treatment advances
have been achieved, all breast cancer patients face the risk of
metastasis to other organs, which is the main cause of death from
breast cancer (2,3). The etiology of breast cancer is a
complex combination of both environmental and genetic factors;
thus, the determination of genetic changes provides a new way to
investigate the etiology of such a complex genetic disease
(4).
The KISS1 gene was originally identified as a
metastasis suppressor gene in human melanoma and breast carcinoma
cells (5). The KISS1 gene
encodes a 145-amino acid peptide that is cleaved into an amidated
C-terminal 54-amino acid product, kisspeptin or metastin. Shorter
fragments of kisspeptin-54, kisspeptin-14, kisspeptin-13, and
kisspeptin-10 (KP-10), also bind to GPR54 (6,7).
KISS1 transfection was found to lead to reduced invasion and
migration of endometrial carcinoma (8) and breast cancer cells (9). Treatment with KP-10 showed similar
effects in cancer cell lines including breast and endometrial
carcinoma cells (10). However, the
mechanisms of KP-10 in breast cancer cells remain unclear.
Epithelial-mesenchymal transition (EMT) is an
essential developmental process through which cells of epithelial
origin lose cell-cell contacts and cell polarity and acquire
mesenchymal phenotypes (11). More
and more evidence shows that EMT is the key means through which
cancer cells acquire more highly mobile potentials to migrate and
metastasize to distant sites during tumor progression (12).
In the present study, we sought to examine the
association between KP-10 and EMT in human breast cancer cells. We
demonstrated that KP-10 regulates EMT in breast cancer cells in
vitro and in vivo. It is anticipated that the results of
this study will provide novel insight into potential strategies for
the treatment of breast cancer.
Materials and methods
Cell culture
MDA-MB-231 and MDA-MB-157 cell lines were purchased
from the American Type Culture Collection (Rockville, MD, USA) and
maintained in Dulbecco’s modified Eagle’s medium (DMEM)
supplemented with 10% fetal calf serum (FCS) and antibiotics (100
μM penicillin and 100 μM streptomycin). Human umbilical vein
endothelial cells (HUVECs) were purchased from the Shanghai
Institute of Biochemistry (Shanghai, China) and maintained in
RPMI-1640 medium supplemented with 10% FCS and antibiotics (100 μM
penicillin and 100 μM streptomycin). Cells were maintained in a
humidified cell incubator with 5% CO2 at 37°C.
MTT assay
Viability of cells was determined using the
3-(4,5-dimethylthiazolyl-2)-2,5-diphenyltetrazolium bromide (MTT)
assay (Sigma-Aldrich, Carlsbad, CA, USA). MDA-MB-231 cells,
MDA-MB-157 cells or HUVECs were plated at a density of
5×103 cells/cm2, cultured until 60%
confluency was reached and treated with various concentrations of
KP-10 (Tyr-Asn-Trp-Asn-Ser-Phe-Gly-Leu-Arg-Phe) (ChinaPeptides Co.,
Ltd., Biomart.cn, Shanghai, China) (0, 20, 40, 60, 80 and 100 ng)
for 48 h. The cells were treated with 0.5 mg/ml MTT for 4 h and
lysed with dimethyl sulfoxide (DMSO). Absorbance rates were
measured at 550–560 nm using a microplate reader (Bio-Rad,
Hercules, CA, USA).
Cell apoptosis assay
Apoptosis was determined using an apoptosis
detection kit (KeyGen, Nanjing, China). Briefly, cells were
collected, washed twice in ice-cold phosphate-buffered saline
(PBS), and then resuspended in binding buffer at a density of
1×106 cells/ml. Cells were simultaneously incubated with
fluorescein-labeled Annexin V and propidium iodide (PI) for 20 min.
The mixture was then analyzed on a FACSCalibur (BD Biosciences,
Baltimore, MD, USA). Annexin V-FITC generated signals were detected
on FL1 at a wavelength of 525 nm, and PI signals were monitored
using a detector reserved for phycoerythrin emission (FL2, 575 nm).
Data were analyzed using CellQuest software from BD.
Transwell migration assay
The migration assay was performed using a Boyden
chamber (8-μm pore size polycarbonate membrane; Cell Biolabs, San
Diego, CA, USA). Cells were resuspended in FBS-free DMEM to a
concentration of 3×105 cells/ml. The upper chamber was
loaded with 200 μl of cell suspension and the lower chamber was
loaded with 600 μl of DMEM containing 10% FBS. The filter was fixed
in 4% paraformaldehyde (Sigma) and stained with crystal violet (CV;
Beyotime, Shanghai, China). The cells on the upper side of the
filter were wiped off using a cotton swab. The cells that migrated
to the undersurface of the membrane were counted using a light
microscope. Ten microscopic fields (x400) were randomly selected to
count the cells.
In vitro wound healing assay
Cells were grown in a 6-well dish. A confluent
monolayer of cells was scratched with a 200-μl pipette tip to
simulate a wound. Cells were washed twice with PBS and then
supplemented with medium and incubated for 4 h at 37°C. Cell
migration into the wounded area was monitored microscopically.
Images were captured at the interface of the unwounded and wounded
areas.
FLIPR Calcium 4 assay
Cell monolayers were plated in black, clear-bottomed
96-well plates overnight. Cells were then incubated in serum-free
media at 37°C for 1 h prior to stimulation with FLIPR Calcium 4
reagent (Molecular Devices, Sunnyvale, CA, USA). Cells were then
incubated for 1 h at 37°C while ligands were prepared in a separate
flat-bottomed 96-well plates. Cells were then tested for intensity
of fluorescence on the NOVOstar (BMG Labtech, UK) over 60 sec with
ligands being added 1:10 at 11 sec.
Gelatin zymography
Fifty micrograms of protein was applied to 10%
polyacrylamide gels with 1% gelatin incorporated as a substrate for
gelatinolytic proteases. After running the gel the SDS was removed
by washing twice in 2.5% Triton X-100 for 30 min. The gels were
incubated overnight in zymography development buffer containing 50
mM Tris-HCl (pH 7.4), 2 mM NaN3 and 5 mM
CaCl2. After development, the gels were stained for 3 h
in 45% methanol/10% glacial acetic acid containing 1% (w/v)
Coomassie blue R-250 and subsequently partially destained with the
same solution without dye. The gelatinolytic activity of each MMP
was evaluated as a clear band against the blue-stained gelatin
background.
Animals
The present study was conducted using protocols
approved by the Animal Care and Use Committee of China Medical
University. NOD/SCID mice were allowed a standard rat diet and
water ad libitum, and maintained on a 10-/14-h light/dark
cycle. MDA-MB-231 or MDA-MB-157 cells (1×108) were
injected into NOD/SCID mice by subcutaneous injection. After the
tumor diameters reached 3–5 mm, the mice were divided randomly into
6 groups (MDA-MB-231, MDA-MB-231 + PBS, MDA-MB-231 + KP-10,
MDA-MB-157, MDA-MB-157 + PBS and MDA-MB-157 + KP-10). Tumor growth
was monitored every 5 days after the first week of inoculation. All
mice were sacrificed at day 60. All of the tumor tissues were
collected, fixed in 4% formaldehyde and embedded in paraffin for
immunostaining.
Immunostaining
For immunohistochemical staining of CD31, endogenous
peroxidase activity was blocked in 4-μm tumor sections with 3%
hydrogen peroxide for 30 min. Antigen retrieval was performed in
citrate buffer (10 mM, pH 6.0) for 30 min at 95°C in a pressure
cooker. The CD31 antibody (Sigma) was incubated with the sections
at 1:500 overnight at 4°C. The sections were then incubated with a
biotinylated secondary antibody for 1 h at room temperature,
followed by incubation with a streptavidin-horseradish peroxidase
(HRP) complex (Beyotime, Beijing, China) for 60 min at room
temperature. The bound antibody was visualized with
3,3′-diaminobenzidine tetrahydrochloride (DAB; Beyotime). Sections
were also counterstained with hematoxylin (Beyotime).
Chick embryo chorioallantoic membrane
(CAM) angiogenesis assay
According to the method of Hong et al
(13), the fertilized eggs were
incubated at 37°C for 8 days and then divided into 2 groups (PBS
and KP-10). After incubation for 3 more days, the CAM of each live
egg was harvested and placed individually in a 6-well plate. Images
of the blood vessels of each CAM were captured using Motic Images
Plus® (MediaCybernetics®, Bethesda, MD, USA).
The blood vessel area as a percentage of the total CAM area, blood
vessel length and number of branch points were quantified using
Image Pro® analysis software
(MediaCybernetics®).
Western blot analysis
Proteins were resolved by 10% SDS-PAGE, transferred
to a nitrocellulose membrane, and detected using the following
antibodies: E-cadherin (sc-7870), vimentin (sc-6260), N-cadherin
(sc-7939) and β-actin (sc-47778) (all from Santa Cruz
Biotechnology, Santa Cruz, CA, USA). Immunostaining was detected
using an enhanced chemiluminescence (ECL) system (Amersham
Biosciences, Westborough, MA, USA).
Statistical analysis
Data are presented as the means ± standard deviation
(SD). Differences between groups were analyzed using the Student’s
t-test for continuous variables. The Kaplan-Meier estimator was
used to compare different patient groups, and P-values were
calculated using the log-rank (Mantel-Cox) test. Statistical
analysis was performed using GraphPad 5.0 software (GraphPad
Software, La Jolla, CA, USA) and significance was established at
P<0.05.
Results
Effects of KP-10 on breast cancer cells
in vitro
The inhibitory growth effects of KP-10 on breast
cancer MDA-MB-231 and MDA-MB-157 cells were determined by MTT
assay. This inhibition was dependent on the concentration of KP-10.
The IC50 values of KP-10 for MDA-MB-231 and MDA-MB-157
cells were 62.5 and 63.3 nM, respectively (P<0.05; Fig. 1A). The apoptotic ratio of
KP-10-treated MDA-MB-231 or MDA-MB-157 cells was 5–15 times higher
compared with the ratio of the respective control cells (P<0.05;
Fig. 1B). The mobility of breast
cancer cells was also inhibited by KP-10. The Transwell assay
showed that less cells after KP-10 treatment migrated to the lower
side of the membrane than the number of cells in the untreated ones
(P<0.05; Fig. 1C). The
wound-healing assay showed that the migration of MDA-MB-231 or
MDA-MB-157 cells after KP-10 treatment was significant reduced when
compared with that in the untreated cells (P<0.05; Fig. 1D). We also found that the activity
of MMP-2 and MMP-9 was inhibited by KP-10 in the MDA-MB-231 and
MDA-MB-157 cells (Fig. 1E).
Furthermore, both MDA-MB-231 and MDA-MB-157 cells exhibited a
marked increase in [Ca2+] in response to treatment with
KP-10 (Fig. 1F). Notably, KP-10 had
similar effects on HUVECs (Fig.
1).
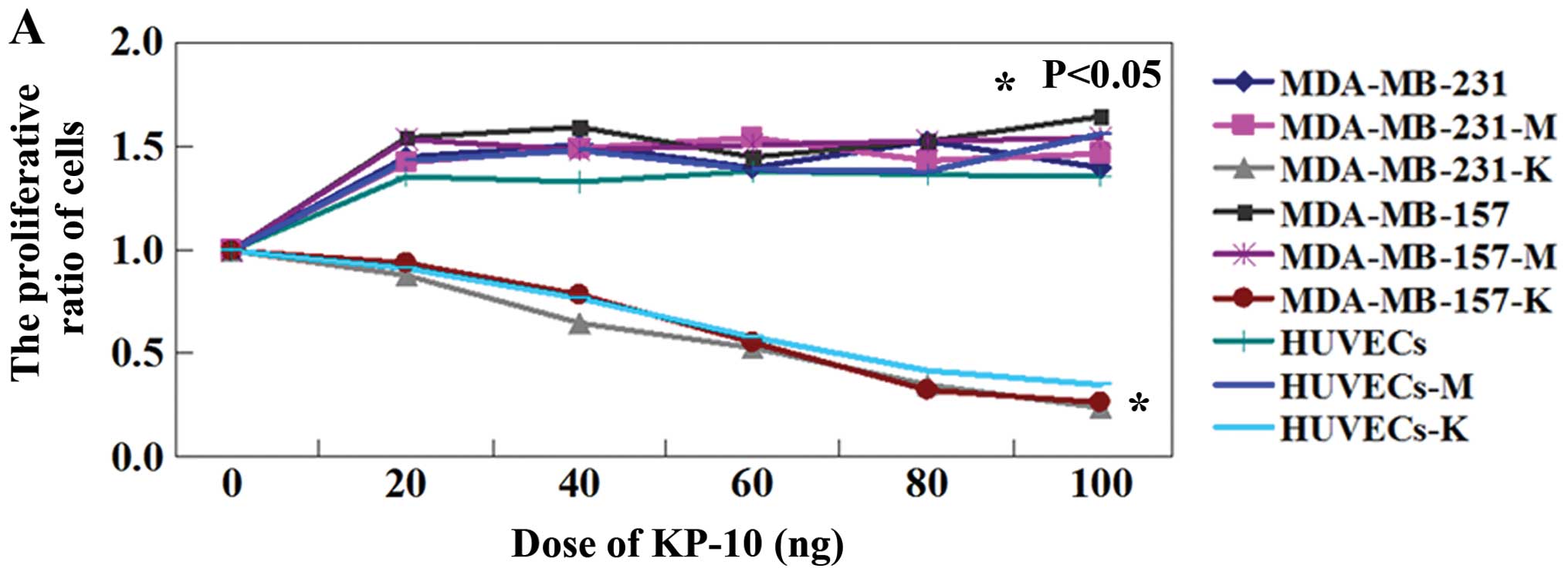 | Figure 1Antitumor activities of KP-10 in
vitro. (A) Growth curves of MDA-MB-231 cells, MDA-MB-157 cells
and HUVECs treated with different concentrations of KP-10. (B) The
proportion of apoptotic cells was determined by double-staining
with Annexin V/FITC and PI. (C) The mobility of the cells was
determined using a Transwell assay. MDA-MB-231, untreated
MDA-MB-231 cells; MDA-MB-231-M, MDA-MB-231 cells treated with PBS;
MDA-MB-231-K, MDA-MB-231 cells treated with KP-10; MDA-MB-157,
untreated MDA-MB-157 cells; MDA-MB-157-M, MDA-MB-157 cells treated
with PBS; MDA-MB-157-K, MDA-MB-157 cells treated with KP-10;
HUVECs, untreated HUVECs; HUVECs-M, HUVECs treated with PBS;
HUVECs-K, HUVECs treated with KP-10. KP-10, kisspeptin-10; HUVECs,
human umbilical vein endothelial cells. (D) Wound closure was
quantified in the regions flanked by dotted lines. (E)
Gelatinolytic activity of secreted MMP-2 and MMP-9 in the breast
cancer cells was analyzed by zymography. (F) The concentration of
Ca2+ was measured using FLIPR Calcium 4 assay.
MDA-MB-231, untreated MDA-MB-231 cells; MDA-MB-231-M, MDA-MB-231
cells treated with PBS; MDA-MB-231-K, MDA-MB-231 cells treated with
KP-10; MDA-MB-157, untreated MDA-MB-157 cells; MDA-MB-157-M,
MDA-MB-157 cells treated with PBS; MDA-MB-157-K, MDA-MB-157 cells
treated with KP-10; HUVECs, untreated HUVECs; HUVECs-M, HUVECs
treated with PBS; HUVECs-K, HUVECs treated with KP-10. KP-10,
kisspeptin-10; HUVECs, human umbilical vein endothelial cells. |
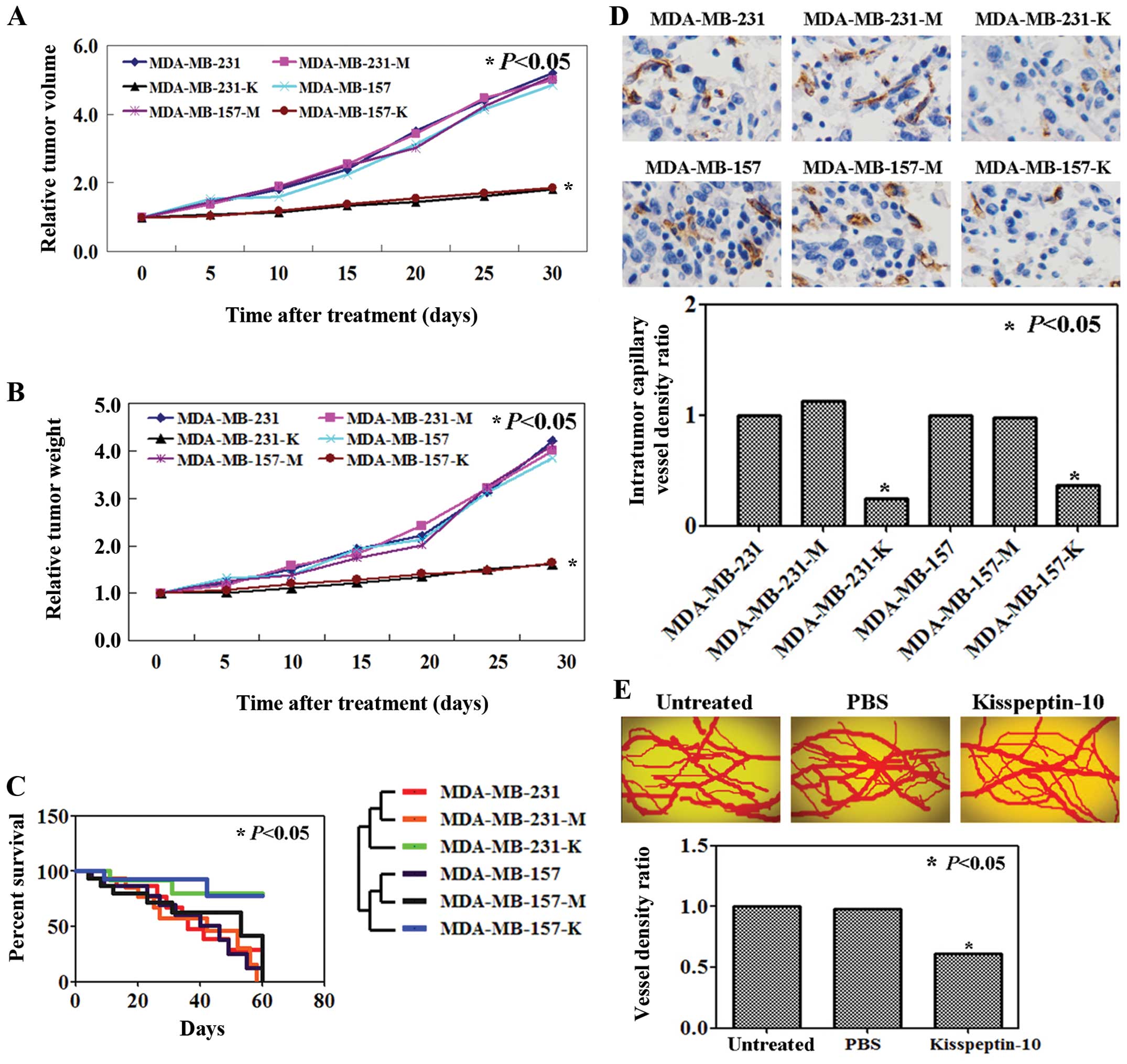
Antitumor activity of KP-10 in a breast
cancer mouse model
The antitumor properties of KP-10 were evaluated
using mouse breast cancer models. Both tumor volume and the weight
of the KP-10-treated tumors were lower compared to the untreated
ones (P<0.05; Fig. 2A and B).
KP-10 also increased the survival rate of the tumor-bearing mice
(P<0.05; Fig. 2C). In addition,
the blood vessel density of the tumors from the mice in the
KP-10-treated group was significantly reduced compared to that in
the untreated group using immunostaining for CD31 expression
(P<0.05; Fig. 2D).
We evaluated the inhibitory effect of KP-10 on in
vivo angiogenesis by using a chick embryo chorioallantoic
membrane angiogenesis assay. The results showed that KP-10
inhibited neovascularization in the chick CAM model (P<0.05;
Fig. 2E).
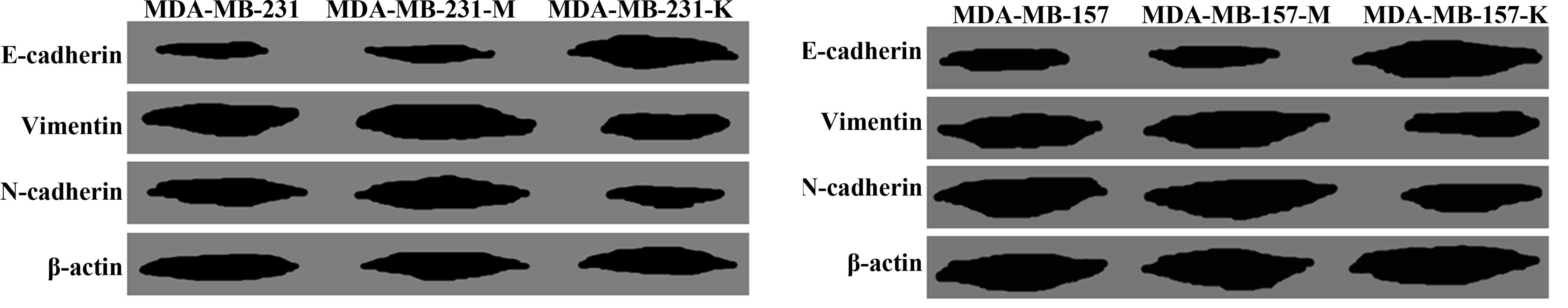
KP-10 induces EMT in breast cancer
cells
Based on the antitumor roles of KP-10 in breast
cancer cells, we carried out western blotting to identify the
mechanism of KP-10. Compared with the untreated cells,
KP-10-treated MDA-MB-231 and MDA-MB-157 cells showed a higher
expression level of epithelial marker E-cadherin, while lower
expression levels of mesenchymal markers vimentin and N-cadherin
were noted (Fig. 3).
Discussion
In the present study, we found that EMT is involved
in the inhibition of migration in breast cancer cells caused by
KP-10. The metastasis suppressor activity of KPs has been
identified in numerous types of cancers, including brain (14), gastric (15), bladder (16) and ovarian (17). Consistent with previous studies, we
also noted a metastasis suppressor role of KP-10 in breast cancer
cells. Some studies have shown that KP-induced metastatic
suppression is related with the suppression of matrix
metalloproteinase (MMP)-9 activity (5,18,19).
Our finding showed that KP-10 increased MMP-9 and MMP-2 activity
and subsequently inhibited cancer cell migration and invasion. A
significant feature of MDA-MB-231 and MDA-MB-157 cell lines is that
KP-10 can stimulate intracellular calcium release. Roseweir et
al (20) found that a similar
effect of KP-10 could induce Ca2+ mobilization in
extravillious trophoblast-derived cells, HTR8SV. However, they did
not find IP3 release during this process. We will investigate the
mechanism of KP-10-induced Ca2+ release in future
studies. KP has been shown to inhibit angiogenesis in tumors and
the placenta (21). Cho et
al (22) found that KP-10
inhibited the migration and invasion of HUVECs. Usui et al
(23) also found that KP-10
inhibited neovascularization in ischaemic limb muscle and inhibited
the proliferation of HUVECs. In the present study, we confirmed
that KP-10 inhibited the migration of breast cancer cells and
HUVECs. Furthermore, we used chick embryo chorioallantoic membrane
angiogenesis assay to confirm the angiogenesis inhibitory role of
KP-10 in vivo.
EMT is a process by which epithelial cells change to
a mesenchymal phenotype and is a crucial step in the initiation of
the metastatic spread of many tumor cells into distal organs
(24). EMT is characterized by a
switch from E-cadherin to N-cadherin expression (25). The loss or reduction in E-cadherin
expression is a key event in the initiation of EMT (26). We here showed that KP-10 was
significantly associated with high expression of E-cadherin, yet
adversely with low N-cadherin expression.
Collectively, the present study revealed that KP-10
suppressed breast cancer and endothelial cell growth both in
vivo and in vitro. KP-10 is a novel regulator of EMT and
metastasis in breast cancer cells.
Acknowledgements
We are indebted to Su-Ning Zhang for his helpful
criticism of the manuscript and excellent technical assistance.
References
|
1
|
Jemal A, Bray F, Center MM, et al: Global
cancer statistics. CA Cancer J Clin. 61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Culhane AC and Quackenbush J: Confounding
effects in ‘a six-gene signature predicting breast cancer lung
metastasis’. Cancer Res. 69:7480–7485. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chiang AC and Massagué J: Molecular basis
of metastasis. N Engl J Med. 359:2814–2823. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Theodoropoulos GE, Saridakis V, Karantanos
T, et al: Toll-like receptors gene polymorphisms may confer
increased susceptibility to breast cancer development. Breast.
21:534–538. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Makri A, Pissimissis N, Lembessis P, et
al: The kisspeptin (KiSS-1)/GPR54 system in cancer biology. Cancer
Treat Rev. 34:682–692. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Muir AI, Chamberlain L, Elshourbagy NA, et
al: AXOR12, a novel human G protein-coupled receptor, activated by
the peptide KiSS-1. J Biol Chem. 276:28969–28975. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kotani M, Detheux M, Vandenbogaerde A, et
al: The metastasis suppressor gene KiSS-1 encodes kisspeptins, the
natural ligands of the orphan G protein-coupled receptor GPR54. J
Biol Chem. 276:34631–34636. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jiang T, Zhang SL, Lin B, et al:
Expression and clinical significance of KiSS-1 and GPR54 mRNA in
endometrial carcinoma. Zhonghua Zhong Liu Za Zhi. 27:229–231.
2005.(In Chinese). PubMed/NCBI
|
|
9
|
Song GQ and Zhao Y: Different therapeutic
effects of distinct KISS1 fragments on breast cancer in vitro and
in vivo. Int J Oncol. 43:1219–1227. 2013.PubMed/NCBI
|
|
10
|
Olbrich T, Ziegler E, Türk G, et al:
Kisspeptin-10 inhibits bone-directed migration of GPR54-positive
breast cancer cells: evidence for a dose-window effect. Gynecol
Oncol. 119:571–578. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lim J and Thiery JP:
Epithelial-mesenchymal transitions: insights from development.
Development. 139:3471–3486. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Scheel C and Weinberg RA: Phenotypic
plasticity and epithelial-mesenchymal transitions in cancer and
normal stem cells? Int J Cancer. 129:2310–2314. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hong J, Zhang Z, Lv W, et al: Icaritin
synergistically enhances the radiosensitivity of 4T1 breast cancer
cells. PLoS One. 8:e713472013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zohrabian VM, Nandu H, Gulati N, et al:
Gene expression profiling of metastatic brain cancer. Oncol Rep.
18:321–328. 2007.PubMed/NCBI
|
|
15
|
Dhar DK, Naora H, Kubota H, et al:
Downregulation of KiSS-1 expression is responsible for tumor
invasion and worse prognosis in gastric carcinoma. Int J Cancer.
111:868–872. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cebrian V, Fierro M, Orenes-Piñero E, et
al: KISS1 methylation and expression as tumor stratification
biomarkers and clinical outcome prognosticators for bladder cancer
patients. Am J Pathol. 179:540–546. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hata K, Dhar DK, Watanabe Y, Nakai H and
Hoshiai H: Expression of metastin and a G-protein-coupled receptor
(AXOR12) in epithelial ovarian cancer. Eur J Cancer. 43:1452–1459.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li N, Wang HX, Zhang J, Ye YP and He GY:
KISS-1 inhibits the proliferation and invasion of gastric carcinoma
cells. World J Gastroenterol. 18:1827–1833. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Navenot JM, Fujii N and Peiper SC:
Activation of Rho and Rho-associated kinase by GPR54 and KiSS1
metastasis suppressor gene product induces changes of cell
morphology and contributes to apoptosis. Mol Pharmacol.
75:1300–1306. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Roseweir AK, Katz AA and Millar RP:
Kisspeptin-10 inhibits cell migration in vitro via a receptor-GSK3
beta-FAK feedback loop in HTR8SVneo cells. Placenta. 33:408–415.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ramaesh T, Logie JJ, Roseweir AK, et al:
Kisspeptin-10 inhibits angiogenesis in human placental vessels ex
vivo and endothelial cells in vitro. Endocrinology. 151:5927–5934.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cho SG, Yi Z, Pang X, et al:
Kisspeptin-10, a KISS1-derived decapeptide, inhibits tumor
angiogenesis by suppressing Sp-1-mediated VEGF expression and
FAK/Rho GTPase activation. Cancer Res. 69:7062–7070. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Usui S, Iso Y, Sasai M, et al:
Kisspeptin-10 induces endothelial cellular senescence and impaired
endothelial cell growth. Clin Sci. 127:47–55. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nakajima S, Doi R, Toyoda E, et al:
N-cadherin expression and epithelial-mesenchymal transition in
pancreatic carcinoma. Clin Cancer Res. 10:4125–4133. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Werling AM, Doerflinger Y, Brandner JM, et
al: Homo- and heterotypic cell-cell contacts in Merkel cells and
Merkel cell carcinomas: heterogeneity and indications for cadherin
switching. Histopathology. 58:286–303. 2011. View Article : Google Scholar : PubMed/NCBI
|