Introduction
The human POU5F1 gene, located on chromosome
6p21.3, can generate at least three transcripts (OCT4A, OCT4B and
OCT4B1 mRNA) and four protein isoforms (Oct4A, Oct4B-265,
Oct4B-190, and Oct4B-164) through alternative splicing and
alternative translation initiation (1). Oct4A was found to be expressed in
unfertilized oocytes, early embryos, primordial germ cells and
different embryonic stem cell (ESC) and embryonal carcinoma cell
(ECC) lines (2). Oct4A has been
established as a master regulator in the maintenance of
self-renewal and pluripotency of ESCs and ECCs (2–4) and is
an indispensable transcription factor in the generation of induced
pluripotent stem cells (iPSCs) (5–7). Oct4B
and Oct4B1 isoforms (including Oct4B-265, 190 and 164), differ from
Oct4A in the N-transactivation domain but share identical POU and
C-transactivation domains with Oct4A. The Oct4B and Oct4B1 isoforms
are considered unable to sustain ESC self-renewal but may play a
role in the response to cell stresses (1,2,8). Apart
from multiple transcriptional and translational products of
POU5F1, six POU5F1 pseudogenes, including processed
and non-processed types, that produce transcripts highly homologous
to OCT4A mRNA have been reported (9). Some of these pseudogenes detected in
certain cancer cell lines and cancer tissues are involved in the
regulation of POU5F1 gene activity and are correlated with
poor prognosis of human cancer (10–12).
Notably, POU5F1P1 (also known as POU5F1B, OCT4-pg1,
OTF3C or OTF3P1), a processed pseudogene located on
human chromosome 8q24.21 and the best documented pseudogene of
POU5F1, may encode a protein exhibiting 95% homology with
the amino acid sequence of Oct4A (13). Of note, Breyer et al
(14) found that OCT4-pg1
expression was positively correlated with POU5F1 expression
in normal prostate tissue and prostate tumors, indicating a
coordinative regulation between the two genes.
However, whether POU5F1 is expressed and
Oct4A protein is present in normal and cancer adult tissues and
cells remains highly controversial, mainly because the RT-PCR
primers and anti-Oct4 antibodies employed did not distinguish Oct4A
from other Oct4 isoforms and its pseudogene products (15–19).
By employing OCT4A-specific primers and OCT4A-specific restriction
digestion of PCR fragments, and by confirming the PCR products with
DNA sequencing, Jez et al (20) found that OCT4A transcription was
undetectable in normal adult human dermal fibroblasts but was
significantly induced when the cells were treated with hypoxia and
FGF2, leading to a so-called ‘regeneration-competent’ state. Thus,
it is possible that OCT4A gene in differentiated cells may
be re-activated under certain conditions.
In addition to the diversity of the transcription
and translation processes of POU5F1 gene, Oct4 protein was
also subjected to multiple post-translational modifications (PTMs)
such as sumoylation, ubiquitination and phosphorylation, all of
which critically regulate its functions (21–24).
Recently, we (25) and other
authors (26,27) reported that human Oct4A can be
phosphorylated directly by Akt at threonine 235 (T235, equivalent
to mouse T228). This site-specific phosphorylation resulted in the
stabilization of Oct4A, and the levels of phosphorylated Oct4A
(Oct4-pT235) correlated with the resistance to apoptosis and the
tumorigenic potential of ECCs (25). As an extension of that study, we
examined the expression of Oct4 and Oct4-pT235 in human somatic
cancer cell lines and tissues using combinatory approaches. The
results suggested that Oct4A or its homolog and Akt-phosphorylated
Oct4-pT235 are present in human cancer cells, and that the Akt-Oct4
regulatory circuit was enhanced in neurosphere cells, thereby
promoting the self-renewal and survival of these stem-like cancer
cells.
Materials and methods
Cell lines and culture
293T, NCCIT, U87, SW837, MCF-7 and HepG2 cells were
obtained from the American Type Culture Collection (ATCC,
Rockville, MD, USA). U251 cells were obtained from Shanghai Bogoo
Biotechnology, Co., Ltd. (Shanghai, China). HCCLM3 cells were
purchased from the Cell Bank of the Chinese Academic of Sciences,
(Shanghai, China). L3.6pl cells, derived from human pancreatic
carcinoma (28), were a gift from
Professor M.H. Wang (Cancer Biology Research Center, School of
Pharmacy, Texas Technical University Health Sciences Center,
Amarillo, TX, USA). Most cells were cultured in DMEM (21063-029,
Invitrogen, Carlsbad, CA, USA), SW837 cells were maintained in
RPMI-1640 medium (11835-030, Invitrogen) and L3.6pl cells were
cultured in MEM (51200-038, Invitrogen), all supplemented with 10%
heat-inactivated fetal bovine serum (FBS) (10099, Gibco, Carlsbad,
CA, USA) and 1% (v/v) penicillin/streptomycin (PS) (15140-148,
Gibco). The cells were cultured at 37°C in a humidified 5%
CO2 incubator (3111, Thermo Fisher Scientific, Waltham,
MA, USA).
Reagents and antibodies
B-27 supplement minus Vitamin A (12587-010) and
basic fibroblast growth factor (bFGF) (PHG0266) were obtained from
Gibco. Epidermal growth factor (EGF) (E5036), leukemia inhibitory
factor (LIF) (L5283) and DMSO (D5879) were purchased from
Sigma-Aldrich (St. Louis, MO, USA). The anti-CK19 (ab52625) was
obtained from Abcam (MA, Cambridge, USA). ITE was chemically
synthesized by KNC Laboratories Co., Ltd. (Tokyo, Japan) (29). The sources of other reagents were
previously described (25).
RT-PCR
Total cell RNAs were extracted using TRIzol reagent
(15596-026, Life Technologies, Carlsbad, CA, USA) and the reverse
transcription reaction was performed using a PrimeScript RT reagent
kit with gDNA eraser (RR047A, Takara, Mountain View, CA, USA). The
‘Total OCT4’ transcripts were amplified by polymerase chain
reaction (PCR) in a C1000 Thermal Cycler (Bio-Rad, Hercules, CA,
USA) using the primers: forward: 5′-GTGGAGGAAGCTGACAACAA-3′ and
reverse: 5′-ATTCTCCAGGTTGCCTCTCA-3′, and the amplified fragment was
120 bp in length. The thermal profile was as follows: 94°C for 2
min; 35 cycles with denaturation at 95°C for 50 sec, annealing at
63°C for 30 sec, and extension at 72°C for 30 sec; with a final
extension at 72°C for 5 min. The ‘OCT4A’ transcripts were amplified
using the primers (30): forward:
5′-CTTCTCGCCCCCTCCAGGT-3′ and reverse: 5′-AAATAG
AACCCCCAGGGTGAGC-3′. The amplified fragment was 496 bp in length,
and the thermal profile was as follows: Denaturation at 94°C for 30
sec, annealing at 64°C for 30 sec, and extension at 72°C for 35 sec
for 35 cycles, with a final extension at 72°C for 10 min. The PCR
products were then subjected to 2% agarose gel electrophoresis with
ethidium bromide (15585011, Life Technologies).
DNA sequencing
A primer pair (forward: 5′-CGGGACA CCTGGCTTCGGAT-3′
and reverse: 5′-CTCAGGCTGAGA GGTCTCCA-3′; with an amplified
fragment of 284/285 bp in length) designated as ‘OCT4A+pg134
primers’, targeting the transcripts of OCT4A, OCT4-pg1, OCT4-pg3
and OCT4-pg4, was designed to investigate the transcription and
proportion of OCT4A and its major pseudogenes in human cancer
cells. The primers were synthesized by Sangon Biotech Shanghai Co.,
Ltd. (Shanghai, China). The thermal profile used was: 94°C for 3
min; 35 cycles with denaturation at 94°C for 30 sec, annealing at
66°C for 30 sec, and extension at 72°C for 45 sec; at the end of 35
cycles an additional extension step of 5 min at 72°C was added. The
PCR products were separated on a 2% agarose gel by electrophoresis
and the positive band was excised and extracted from the gel using
the SanPrep Column DNA Gel Extraction kit (SK8132, Sangon Biotech
Shanghai Co., Ltd.) and then cloned to pEASY-T1 Simple Cloning
Vector (CT111, Beijing TransGen Biotech Co., Ltd., Beijing, China).
The recombinants were then transfected into DH5α-competent cells
(CD201, Beijing TransGen Biotech Co., Ltd.). Ampicillin-resistant
clones were selected for sequencing by Sangon Biotech Shanghai Co.,
Ltd. The obtained DNA sequences were analyzed with the Nucleotide
BLAST program and the Lasergene software package (DNA Star Inc.,
Madison, WI, USA).
Western blot analysis
Cultured cells with or without treatment were lysed,
and the whole cell lysates were separated by SDS-PAGE, blotted onto
nitrocellulose membranes, and probed with the indicated antibodies,
as previously described (25).
GAPDH was used as an internal control.
Lentiviral vector construction, viral
production and viral infection
Lentiviral vector construction and viral production
were carried out as described previously (25). For viral infection, U87 cells were
plated and cultured overnight, and the culture media were replaced
with viral supernatants supplemented with polybrene (AL-118,
Sigma-Aldrich) at a final concentration of 8 μg/ml. In most cases,
the multiplicity of infection (MOI) was estimated to be between 0.5
and 2. After 8–10 h, the viral supernatants were replaced with
fresh culture medium to allow further growth until use.
Immunofluorescence microscopy and
immunohistochemistry
Immunofluorescence microscopy was performed as
previously described (25). For
immunohistochemistry, surgically resected specimens were collected
from patients with liver cancer or glioma prior to therapeutic
treatment. Non-cancerous liver or brain tissue specimens were
obtained from individuals who had died in accidents. Tissue
collection or autopsies were conducted at the First Affiliated
Hospital, School of Medicine, Zhejiang University. Written informed
consent was obtained from the patients or their relatives and the
study was conducted with the approval of the Ethics Committee of
the First Affiliated Hospital, School of Medicine, Zhejiang
University. Immunohistochemical staining was carried out with
paraffin-embedded specimens. Briefly, the sections were cut,
de-waxed in 100% xylene twice, rehydrated in a graded alcohol
series (100, 95, 80 and 70%, for 5 min each), rinsed with water
three times, and incubated in 0.3% H2O2 in
methanol for 20 min to inactivate endogenous peroxidase activity.
The sections were then boiled in a microwave oven in 0.01 M citrate
buffer solution for 10 min. Each section was incubated with 3%
bovine serum albumin (BSA, 9048-46-8, Sigma-Aldrich) for 30 min,
followed by incubation with the rabbit anti-Oct4-pT235 antibody
(25) diluted in 1% BSA (1:500)
overnight at 4°C. The sections were washed with PBS three times,
for 5 min each. After incubation with peroxidase-conjugated
anti-rabbit secondary antibody (7074, Cell Signaling Technology,
Beverly, MA, USA) for 60 min, each section was washed with PBS for
three 5-min washes as described above. The sections were visualized
by an Olympus IX81 microscope with an Olympus IX-TVAD camera
following development in 3,3′-diaminobenzidine tetrahydrochloride
(DAB) solution and counterstaining by hematoxylin for 1 min and
sealed by neutral gum.
Mouse xenograft tumor model and
immunohistochemistry
BALB/c nude mice were purchased from the Shanghai
Experimental Animal Centre, Chinese Academy of Science and
maintained as previously described (25). Animal experiments were conducted in
accordance with the Guide for the Care and Use of Animals for
Research Purposes, and were approved by the Committee of Animal
Ethics, Zhejiang University.
The HCCLM3 cell orthotopic xenograft model was
established as previously described (31). ITE treatment was performed two weeks
after implantation. Each treatment group consisted of three
tumor-bearing mice. Vehicle (DMSO) or ITE in the vehicle at 80
mg/kg body weight was administered to the mice by intraperitoneal
injection once daily for 15 consecutive days. The mice were then
sacrificed. Following tumor excision from the euthanized mice, a
portion of the tumor tissue was fixed in 10% formalin for
subsequent histological examination as previously described
(31). Immunohistochemistry was
performed as described above using anti-CK19 (diluted at 1:250) and
anti-Oct4-pT235 (diluted at 1:200) antibodies.
Neurosphere and WST-1 assays
The neurosphere assay was performed as previously
described (32). Typical
neurospheres formed after cells were cultured in NSC medium for 5
days. Cell proliferation was determined by the WST-1 Cell
Proliferation kit (05015944001, Roche Diagnostics, Indianapolis,
IN, USA) according to the manufacturer’s instructions.
DNA microarray
The service for DNA microarray analysis was provided
by the Shanghai Biotechnology Corp. (Shanghai, China). Briefly, RNA
was extracted from U87 parental and neurosphere cells using the
TRIzol method according to the manufacturer’s instructions. Reverse
transcription to the first-strand cDNA was primed with T7 oligo(dT)
primer to synthesize cDNA containing a T7 promoter sequence. The
second-strand cDNA synthesis converted the single-stranded cDNA
into a double-stranded DNA (dsDNA) template for transcription.
Hybridization was carried out by the GeneChip Hybridization, Wash,
and Stain kit (affymetrix: 900720). The arrays were scanned at 570
nm with a confocal scanner from Affymetrix. Analysis of the arrays
was performed using the Partek GS 6.5. Normalization of the array
was performed using a robust multiarray analysis (RMA). A p-value
cut off of 0.05 was used to filter genes that were significantly
expressed between the two samples. A fold change of >1.5 was
used as a criterion for differential gene expression. Histograms of
upregulated genes were prepared using GraphPad Prism 5.0 software
(GraphPad Software, Inc., La Jolla, CA, USA).
Statistical analysis
Statistical analyses were carried out using the SPSS
19.0 statistical software package (IBM Corp., New York, NY, USA).
Quantitative data were presented as means ± SD of three independent
experiments. The statistical significance was evaluated using the
two-tailed unpaired Student’s t-test. P<0.05 was considered
statistically significant.
Results
Oct4 and Akt-phosphorylated Oct4-pT235
are detected in human cancer cell lines
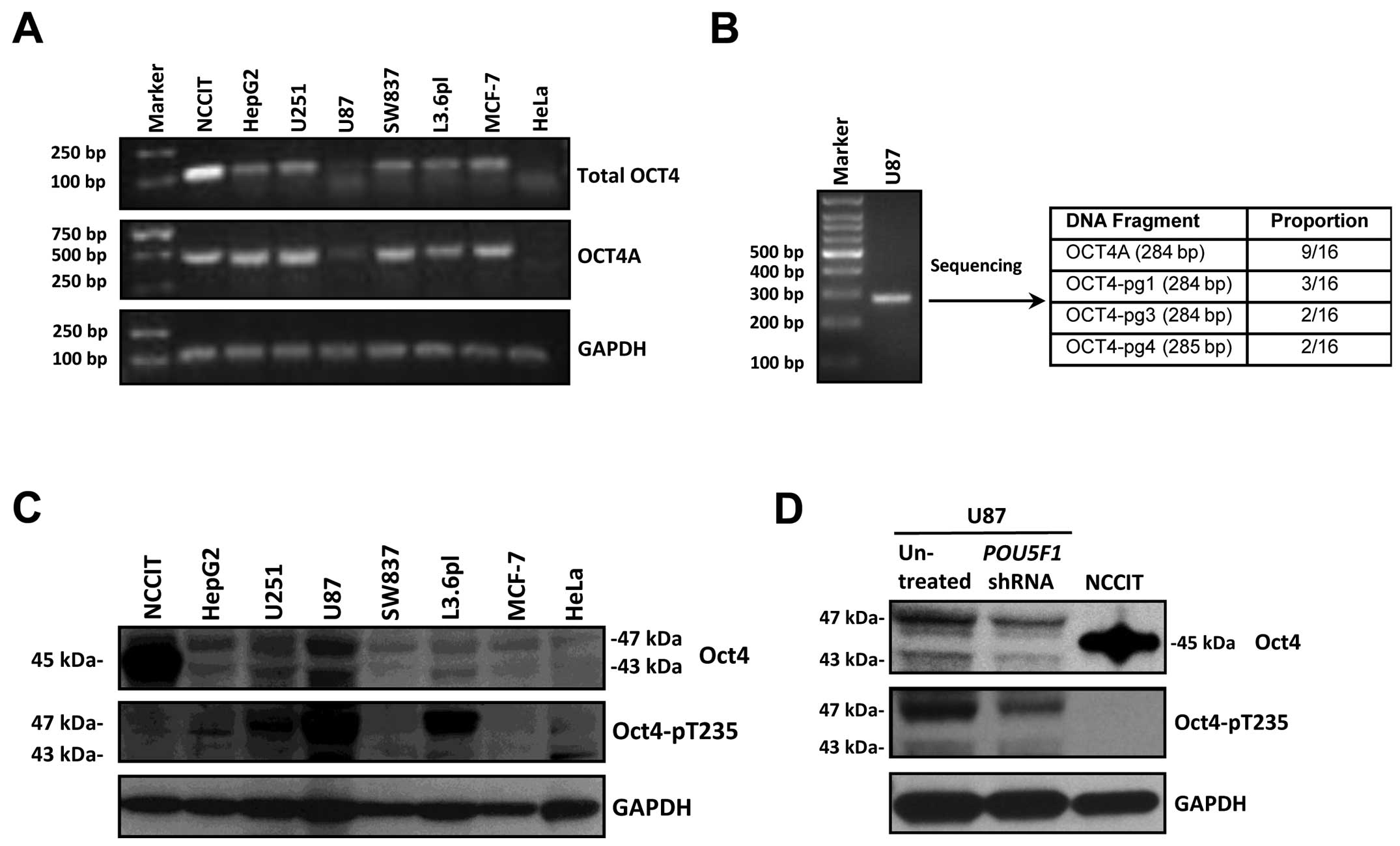
Using a pair of primers that can amplify all known
OCT4 isoforms and POU5F1 pseudogenes (designated as ‘Total
OCT4’) (25), we detected the mixed
OCT4 transcripts in a variety of human cancer cell lines by RT-PCR
(Fig. 1A). The same set of samples
was amplified by the OCT4A-specific primers that presumably exclude
major known POU5F1 pseudogenes as described by Atlasi et
al (30). A clear band with
anticipated size (496 bp) was detected in all the tested cancer
cell lines with the exception of HeLa cells (Fig. 1A). Furthermore, a single band was
visible when the U87 cell sample was amplified with the primers
that target the transcripts of OCT4A and three pseudogenes
(OCT4-pg1, OCT4-pg3, OCT4-pg4), and DNA sequencing confirmed the
presence of all four types of transcripts in U87 cells (Fig. 1B). The anti-Oct4A antibody which
identified a strong band of 45 kDa in NCCIT cells detected two
major bands in most human cancer cell lines that ran at 47 and 43
kDa, respectively, with the 47 kDa band being stronger in most
cases (Fig. 1C). When the
electrophoresis conditions were set to optimal, the major 47 kDa
band was further resolved into two bands. Using an antibody that
can specifically recognize Akt-phosphorylated Oct4 (Oct4-pT235), we
detected strong bands in several but not all cell lines that
coincided with the 47 kDa band(s) (Fig.
1C). Since U87 cells exhibited the strongest signals for the 47
kDa Oct4 band(s) and the Oct4-pT235 band, we conducted most of the
subsequent studies with this cell line. To determine whether any of
the above detected protein bands were truly derived from Oct4, we
compared the bands in U87 cells treated with or without a
POU5F1 shRNA (25) that can
target the 3′UTR of POU5F1 (including OCT4A, OCT4B and
OCT4B1) and OCT4-pg1. Of note, both the 47 and 43 kDa bands
recognized by either anti-Oct4 or anti-Oct4-pT235 were
significantly reduced by the POU5F1 shRNA, while other
non-specific bands remained unchanged (Fig. 1D). Since the reduction of the 47 kDa
band by POU5F1 shRNA was more obviously revealed by
anti-Oct4-pT235, it is likely to be the main Oct4 variant that is
phosphorylated by Akt in U87 cells. Taken together, our data
suggested that Oct4A or its homolog protein (most likely Oct4-pg1)
was present in certain human cancer cell lines such as glioblastoma
cells and it is phosphorylated by Akt at T235.
Akt-phosphorylated Oct4-pT235 is
predominantly localized in the nucleus of cancer cells
To determine the intracellular localization of Oct4
in human cancer cells, we examined Oct4 and pOct-T235 in the U87
and U251 glioblastoma cell lines by immunofluorescence microscopy,
using the anti-Oct4 and anti-pOct-T235 employed in the above
western blot analysis and in our previous study (25), with 293T and NCCIT cells being the
negative and positive controls, respectively. The nuclei were
counterstained with Hoechst 33342. As expected, there was no
discernable Oct4 or Oct4-pT235 immunofluorescent signal in 293T
cells.
Consistent with results of previous studies
(25), Oct4 was predominantly
localized in the nuclei of NCCIT cells and the immunofluorescent
signals were relatively strong (Fig.
2A). By contrast, the Oct4 signals in U87 and U251 cells were
much weaker but still discernable. Notably, the immunofluorescence
intensity of Oct4-pT235 exhibited an opposite pattern where the
signals seen in the two glioblastoma cell lines were much stronger
than those in NCCIT cells although all the signals were mainly
detected in the nucleus (Fig. 2A).
To further confirm the presence and the subcellular localization of
Oct4-pT235 in human cancer cells, we then conducted
immunohistochemical analysis of Oct4-pT235 in tissue specimens from
normal and liver cancer or glioblastoma patients. The results
showed that Oct4-pT235 was present at certain regions in the human
liver cancer and glioblastoma specimens but rarely in normal liver
and brain specimens (Fig. 2B), and
consistent with the immunofluorescence data, Oct4-pT235 was
predominantly localized in the nucleus of cells in those cancer
tissues. Collectively, our data indicated that Oct4A or its homolog
protein is present in the nucleus of certain human cancer cells and
a significant portion of this Oct4 is phosphorylated at T235 by
Akt.
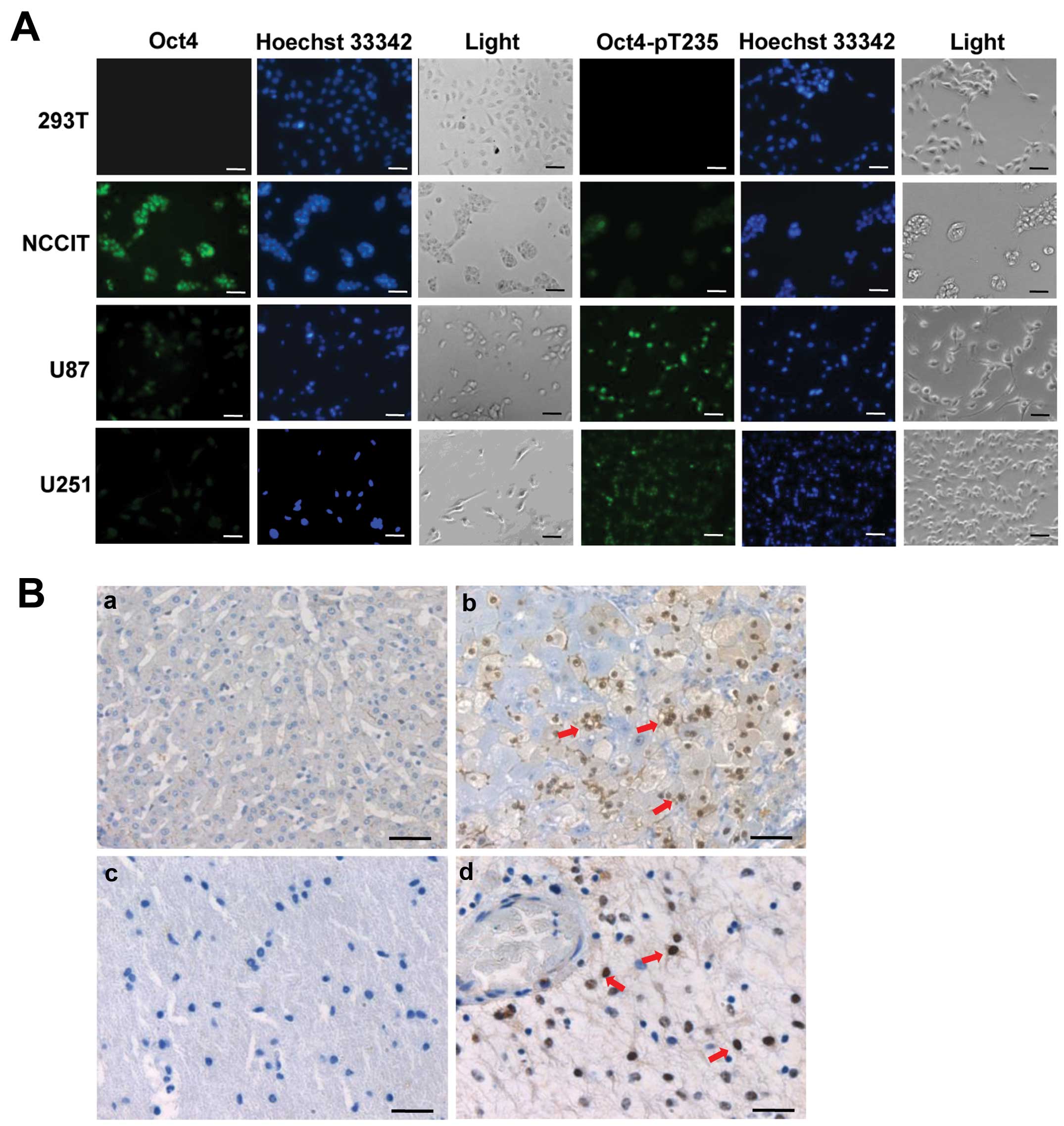 | Figure 2Oct4-pT235 is predominantly localized
in the nucleus of cancer cells. (A) U87 and U251 cells were
immunostained with anti-Oct4 (green) or anti-Oct4-pT235 (green),
respectively, and counterstained with Hoechst 33342 (blue). 293T
and NCCIT cells were used as the negative and positive controls,
respectively. For all panels, original magnification, ×200; scale
bar, 50 μm. (B) Human cancerous and non-cancerous tissues were
subjected to immunohistochemical staining with anti-Oct4-pT235 (DAB
staining and hematoxylin counterstaining). (a and b) Oct4-pT235
staining of human non-cancerous and cancerous liver sections,
respectively. Original magnification, ×200; scale bar, 50 μm. (c
and d) Oct4-pT235 staining of human non-cancerous brain tissue and
glioma sections, respectively. Original magnification, ×200; scale
bar, 50 μm. |
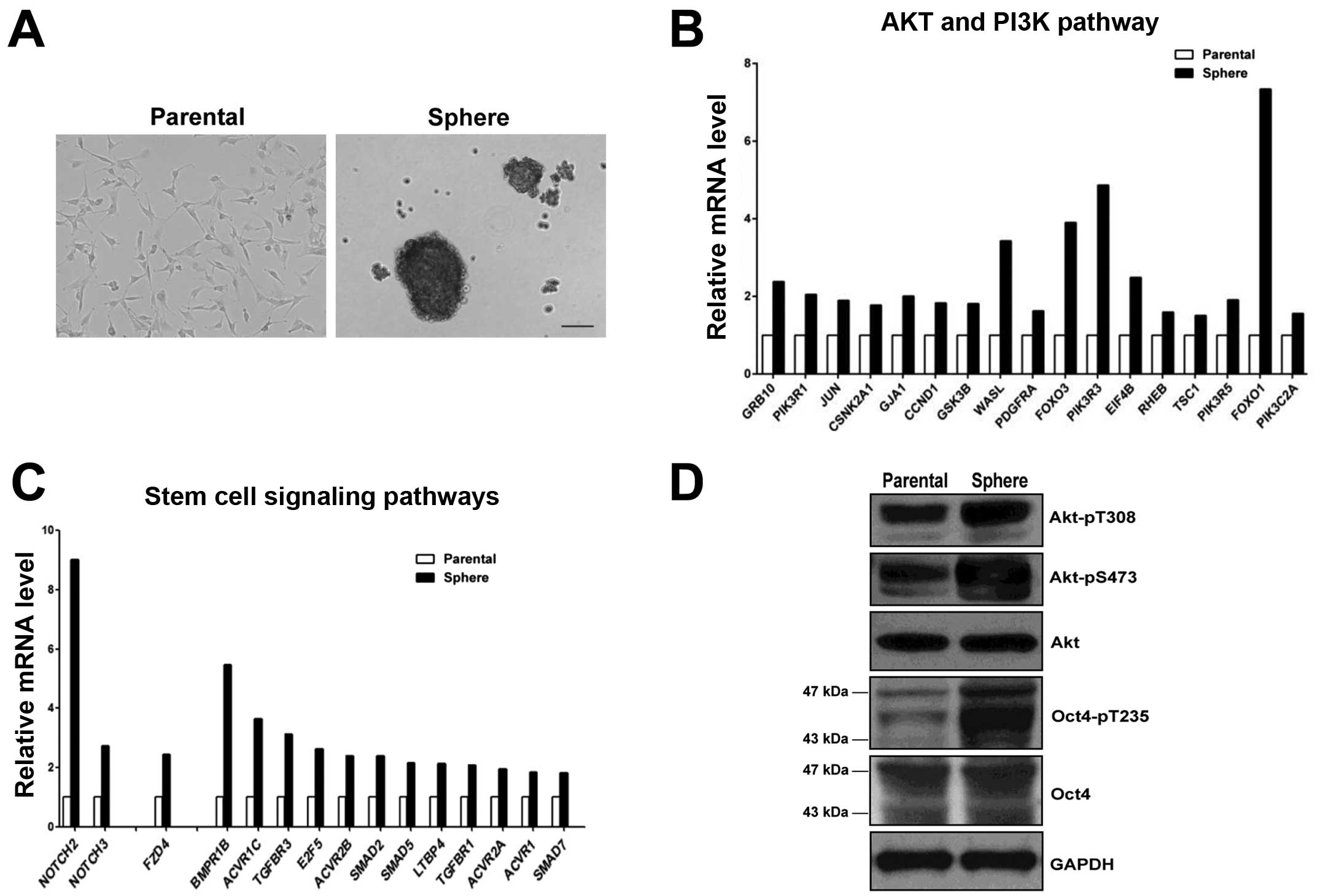
Level of Akt-phosphorylated Oct4-pT235 is
increased in glioblastoma stem-like cells
It has been established that when glioblastoma cells
such as U87 cells are cultured in a neural stem cell (NSC) medium
to induce the formation of neurospheres, a number of stem cell
markers are upregulated and therefore such neurospheres are
considered to be rich in glioblastoma stem-like cells (32,33).
We obtained the U87 neurospheres that exhibited the
identical morphological features as those reported in the
literature (Fig. 3A). A genome-wide
DNA microarray analysis was conducted to comprehensively compare
the transcriptional profiles between U87 parental and sphere cells.
A total of 4,642 genes were found to be upregulated in U87
neurosphere cells. Among them, some were associated with the
PI3K-Akt pathway (Fig. 3B), and a
number of stem-cell related pathways such as the Notch, Wnt and
TGFβ pathways (Fig. 3C). We then
compared the protein levels of Akt and Oct4 between U87 parental
cells and corresponding neurosphere cells by western blot analysis.
While there was no discernable change in the total Akt level in U87
neurosphere cells, the levels of Akt-pT308 and particularly
Akt-pS473 were markedly enhanced. Similarly, although the total
Oct4 level was similar between U87 parental and sphere cells, the
Oct4-pT235 level in sphere cells was significantly higher than that
in parental cells (Fig. 3D).
Collectively, our data showed that the PI3K-Akt signaling pathway
and the major stem cell self-renewal pathways were upregulated in
glioblastoma stem-like cells, and this was associated with an
increased Oct4-pT235 level in those cells.
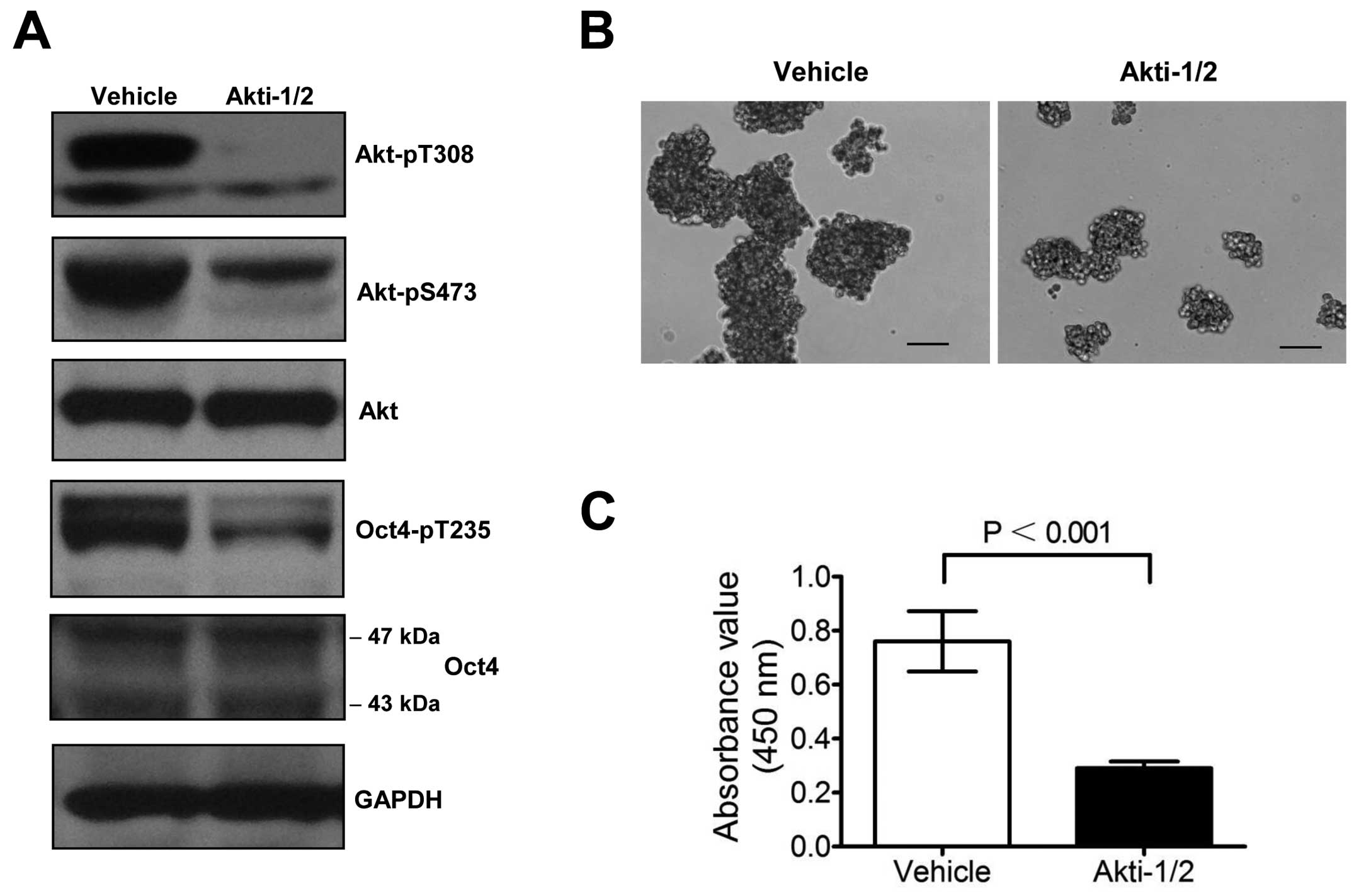
Inhibition of Akt activity decreases the
Oct4-pT235 level and attenuates the proliferation of glioblastoma
stem-like cells
We previously demonstrated that Oct4-pT235 promotes
the self-renewal and survival of ECCs (25). In the present study, we found that,
Akti-1/2, a specific inhibitor of Akt, significantly reduced the
level of Oct4-pT235 in U87 neurosphere cells while concomitantly
decreasing the Akt-pT308 and Akt-pS473 levels (Fig. 4A). Notably, the sizes of
Akti-1/2-treated U87 neurospheres were much smaller than those of
vehicle-treated neurospheres (Fig.
4B), and the proliferation of Akti-1/2-treated neurosphere
cells were markedly attenuated (Fig.
4C).
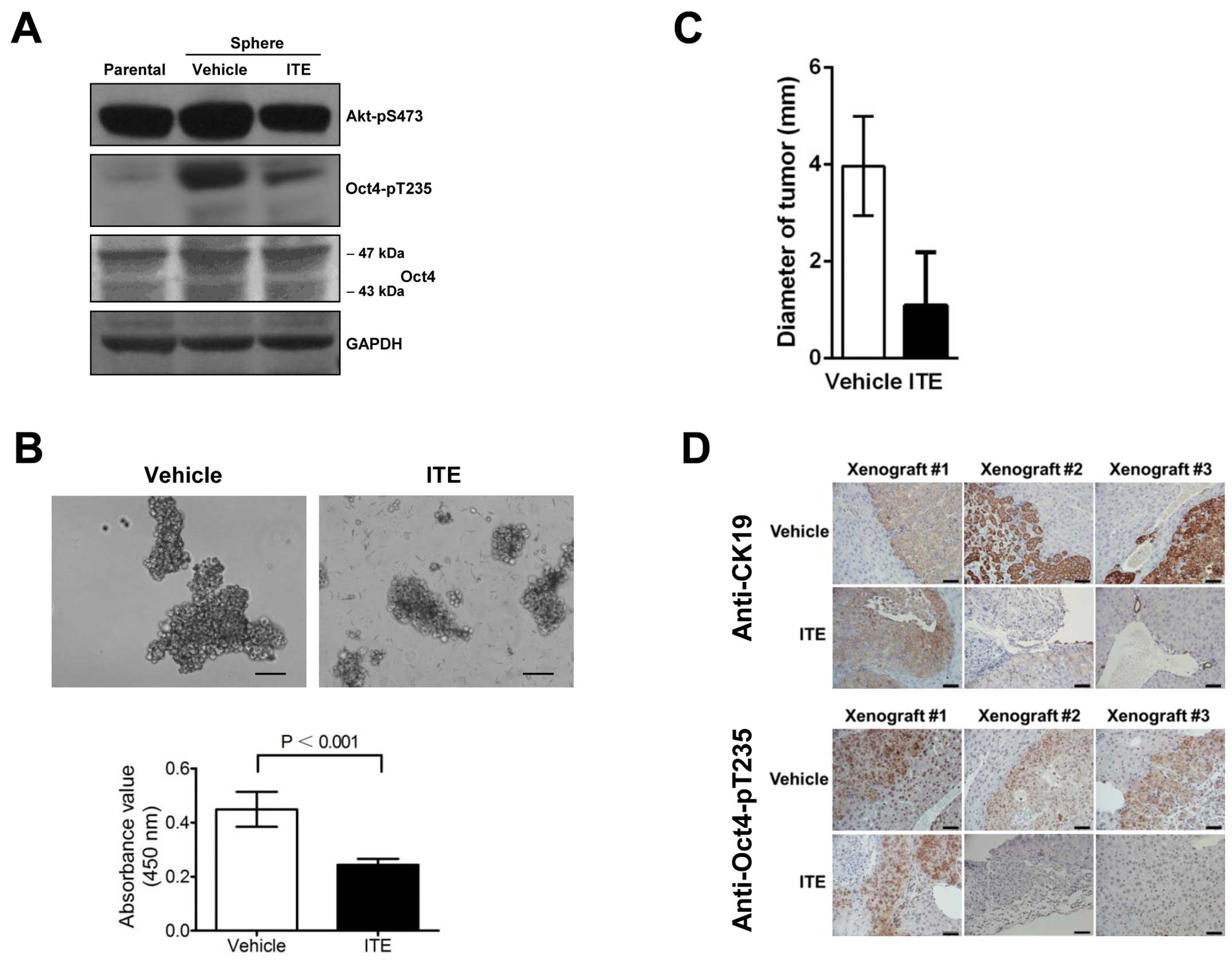
ITE decreases the Oct4-pT235 level and
inhibits the proliferation of stem-like cancer cells in
neurospheres and in a mouse xenograft model
Emerging evidence indicated an intrinsic connection
between the PI3K/Akt signaling pathway and the aryl hydrocarbon
receptor (AhR), a ligand-activated transcription factor responding
to environmental toxicants. For instance, AhR-deficient hepatoma
cells exhibited impaired activation of Akt and enhanced sensitivity
to apoptosis (34).
β-naphthoflavone (BNF), an agonist of the AhR and a putative
chemotherapeutic agent, inhibited PI3K/Akt signaling in breast
cancer cells (35). We demonstrated
that ITE, another agonist of the AhR that possesses antitumor
activities (29), significantly
reduced the Oct4-pT235 level in U87 neurospheres (Fig. 5A) and inhibited their proliferation
in vitro (Fig. 5B). To
better mimic the organ and tissue microenvironment in which the
tumors grow, we orthotopically transplanted the human
hepatocellular HCCLM3 carcinoma cells into the livers of nude mice.
Hematoxylin-eosin (H&E) staining showed that all three
vehicle-treated transplanted mice bore prominent tumors, while only
one out of three ITE-treated mice was found to contain growing
tumor, and the average tumor size of the ITE group was smaller than
that of the vehicle group (Fig.
5C). Staining for the hepatocellular carcinoma marker
cytokeratin-19 (CK19) further confirmed the observations with the
H&E staining and revealed typical and expected cytological
characteristics of the transplanted HCCLM3 cells (Fig. 5D). In the vehicle group, the levels
of Oct4-pT235 in the implanted HCCLM3 cells were significantly
higher than those in adjacent mouse liver cells, indicating that
this site-specific phosphorylation of Oct4 may be associated with
tumorigenesis and/or the maintenance of tumors. By contrast, there
were only background levels of Oct4-pT235 in two of the ITE-treated
mice (Fig. 5D), further confirming
the role of ITE in reducing the Oct4-pT235 level as seen above in
U87 neurosphere cells (Fig.
5A).
Discussion
The presence of Oct4A protein in normal or cancerous
adult tissues or cells has been widely questioned due to the lack
of convincing data. In this study, using a well-characterized
monoclonal anti-Oct4A antibody, we detected two bands (of 43 and 47
kDa) in western blots that can be significantly reduced by an shRNA
targeting POU5F1 and OCT4-pg1, indicating they are
highly associated with Oct4A protein. Detection of the upper band
(47 kDa) by anti-Oct4-pT235 further confirmed its close association
with Oct4A. Given the extremely high degree (95%) of sequence
homology between Oct4A and Oct4-pg1 proteins, it is difficult to
distinguish them at the protein level using routine approaches.
Thus, it is critical in future study to purify the endogenous 47
kDa proteins and to fully identify them by amino acid sequencing.
More importantly, irrespective of whether the 47 kDa protein
corresponds to Oct4A or its pseudogene product, additional
investigations are required to fully characterize its target genes
and functional roles in the context of human cancer cells.
To the best of our knowledge, we have shown for the
first time that a specific site in Oct4A or its homolog protein,
which corresponds to T235 in human Oct4A, in human cancer cells can
be phosphorylated by serine/threonine-protein kinase Akt. When U87
glioblastoma cells were cultured in NSC medium to induce the
formation of neurospheres that are rich in glioblastoma stem-like
cells, the level of Oct4-pT235 in sphere cells was markedly
increased compared to their monolayer parental cells. Consistent
with previous studies (36,37), we found the PI3K/Akt pathway to be
generally upregulated in U87 sphere cells. Emerging evidence
indicates that PI3K/Akt signaling is critical for the self-renewal
of stem-like cancer cells and tumor progression (38,39).
Moreover, the aberrantly activated Oct4/Tcl1/Akt signaling pathway
contributed to chemotherapeutic drug resistance in liver cancer
cells (40). The effects of Akt
were thought to be mediated by the activation of the Wnt/β-catenin
pathway and GSK3β phosphorylation in mammary stem/progenitor cells
(41), and by Akt-induced ABCG2
activation in glioblastoma stem-like cells (42).
The results of the present study suggest that Oct4
may be another important substrate for Akt in maintaining the
self-renewal of stem-like cancer cells. It is noteworthy that among
the examined human cancer cell lines, U87 exhibited the lowest Oct4
mRNA level but the highest Oct4 protein level and Oct4-pT235 level.
Since our previous study has shown that Akt-mediated
phosphorylation at the T235 site stabilizes Oct4 protein by
attenuating its proteasome-dependent degradation (25), it is possible that a high level of
Oct4-pT235 along with other PTMs may facilitate the stabilization
of Oct4 protein and increase its steady-state level in U87 cells.
In this study, we established that reducing the Oct4-pT235 level by
either Akti-1/2 or ITE is associated with attenuated proliferation
of stem-like cancer cells in vitro and in vivo.
Future studies are required to assess the potential value of
Oct4-pT235 as a diagnostic, prognostic, or predictive biomarker for
certain types of human cancer, and to fully decipher the complete
Akt-Oct4 interplay system in stem-like cancer cells which may be
useful in revealing the intrinsic connection between self-renewal
and the tumorigenesis of those cells, and in developing more
effective interventions to eradicate or inhibit them.
Acknowledgements
We thank Dr Jiasheng Song (AhR Pharmaceuticals,
Inc., Madison, WI, USA) for kindly providing the ITE. This study
was supported by the National Natural Science Foundation of China
(grant no. 31271465 and no. 81472722 to Y.-J.W., grant no. 81101662
to Q.Z., grant no. 81071767 to M.Z.).
References
|
1
|
Wang X and Dai J: Concise review: isoforms
of OCT4 contribute to the confusing diversity in stem cell biology.
Stem Cells. 28:885–893. 2010.PubMed/NCBI
|
|
2
|
Jerabek S, Merino F, Scholer HR and
Cojocaru V: OCT4: dynamic DNA binding pioneers stem cell
pluripotency. Biochim Biophys Acta. 1839.138–154. 2014.
|
|
3
|
Jung M, Peterson H, Chavez L, et al: A
data integration approach to mapping OCT4 gene regulatory networks
operative in embryonic stem cells and embryonal carcinoma cells.
PLoS One. 5:e107092010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Young RA: Control of the embryonic stem
cell state. Cell. 144:940–954. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Park IH, Zhao R, West JA, et al:
Reprogramming of human somatic cells to pluripotency with defined
factors. Nature. 451:141–146. 2008. View Article : Google Scholar
|
|
6
|
Takahashi K, Tanabe K, Ohnuki M, et al:
Induction of pluripotent stem cells from adult human fibroblasts by
defined factors. Cell. 131:861–872. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yu J, Vodyanik MA, Smuga-Otto K, et al:
Induced pluripotent stem cell lines derived from human somatic
cells. Science. 318:1917–1920. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cauffman G, Liebaers I, Van Steirteghem A
and Van de Velde H: POU5F1 isoforms show different expression
patterns in human embryonic stem cells and preimplantation embryos.
Stem Cells. 24:2685–2691. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pain D, Chirn GW, Strassel C and Kemp DM:
Multiple retropseudogenes from pluripotent cell-specific gene
expression indicates a potential signature for novel gene
identification. J Biol Chem. 280:6265–6268. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Suo G, Han J, Wang X, Zhang J, Zhao Y and
Dai J: Oct4 pseudogenes are transcribed in cancers. Biochem Biophys
Res Commun. 337:1047–1051. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang L, Guo ZY, Zhang R, et al: Pseudogene
OCT4-pg4 functions as a natural micro RNA sponge to regulate OCT4
expression by competing for miR-145 in hepatocellular carcinoma.
Carcinogenesis. 34:1773–1781. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hayashi H, Arao T, Togashi Y, et al: The
OCT4 pseudogene POU5F1B is amplified and promotes an aggressive
phenotype in gastric cancer. Oncogene. Dec 23–2013.(Epub ahead of
print). PubMed/NCBI
|
|
13
|
Panagopoulos I, Moller E, Collin A and
Mertens F: The POU5F1P1 pseudogene encodes a putative protein
similar to POU5F1 isoform 1. Oncol Rep. 20:1029–1033.
2008.PubMed/NCBI
|
|
14
|
Breyer JP, Dorset DC, Clark TA, et al: An
expressed retrogene of the master embryonic stem cell gene POU5F1
is associated with prostate cancer susceptibility. Am J Hum Genet.
94:395–404. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu J, Ma L, Xu J, et al: Spheroid
body-forming cells in the human gastric cancer cell line MKN-45
possess cancer stem cell properties. Int J Oncol. 42:453–459.
2013.
|
|
16
|
Liedtke S, Stephan M and Kogler G: Oct4
expression revisited: potential pitfalls for data misinterpretation
in stem cell research. Biol Chem. 389:845–850. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cantz T, Key G, Bleidissel M, et al:
Absence of OCT4 expression in somatic tumor cell lines. Stem Cells.
26:692–697. 2008. View Article : Google Scholar
|
|
18
|
Lengner CJ, Welstead GG and Jaenisch R:
The pluripotency regulator Oct4: a role in somatic stem cells? Cell
Cycle. 7:725–728. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mueller T, Luetzkendorf J, Nerger K,
Schmoll HJ and Mueller LP: Analysis of OCT4 expression in an
extended panel of human tumor cell lines from multiple entities and
in human mesenchymal stem cells. Cell Mol Life Sci. 66:495–503.
2009. View Article : Google Scholar
|
|
20
|
Jez M, Ambady S, Kashpur O, et al:
Expression and differentiation between OCT4A and its pseudogenes in
human ESCs and differentiated adult somatic cells. PLoS One.
9:e895462014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wei F, Scholer HR and Atchison ML:
Sumoylation of Oct4 enhances its stability, DNA binding, and
transactivation. J Biol Chem. 282:21551–21560. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xu H, Wang W, Li C, et al: WWP2 promotes
degradation of transcription factor OCT4 in human embryonic stem
cells. Cell Res. 19:561–573. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Saxe JP, Tomilin A, Scholer HR, Plath K
and Huang J: Post-translational regulation of Oct4 transcriptional
activity. PLoS One. 4:e44672009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Brumbaugh J, Hou Z, Russell JD, et al:
Phosphorylation regulates human OCT4. Proc Natl Acad Sci USA.
109:7162–7168. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lin Y, Yang Y, Li W, et al: Reciprocal
regulation of Akt and Oct4 promotes the self-renewal and survival
of embryonal carcinoma cells. Mol Cell. 48:627–640. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Campbell PA and Rudnicki MA: Oct4
interaction with Hmgb2 regulates Akt signaling and pluripotency.
Stem Cells. 31:1107–1120. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen B, Xue Z, Yang G, et al: Akt-signal
integration is involved in the differentiation of embryonal
carcinoma cells. PLoS One. 8:e648772013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Padhye SS, Guin S, Yao HP, Zhou YQ, Zhang
R and Wang MH: Sustained expression of the RON receptor tyrosine
kinase by pancreatic cancer stem cells as a potential targeting
moiety for antibody-directed chemotherapeutics. Mol Pharm.
8:2310–2319. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang K, Li Y, Jiang YZ, et al: An
endogenous aryl hydrocarbon receptor ligand inhibits proliferation
and migration of human ovarian cancer cells. Cancer Lett.
340:63–71. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Atlasi Y, Mowla SJ, Ziaee SA, Gokhale PJ
and Andrews PW: OCT4 spliced variants are differentially expressed
in human pluripotent and nonpluripotent cells. Stem Cells.
26:3068–3074. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhao GJ, Xu LX, Chu ES, et al:
Establishment of an orthotopic transplantation tumor model of
hepatocellular carcinoma in mice. World J Gastroenterol.
18:7087–7092. 2012. View Article : Google Scholar
|
|
32
|
Ledur PF, Villodre ES, Paulus R, Cruz LA,
Flores DG and Lenz G: Extracellular ATP reduces tumor sphere growth
and cancer stem cell population in glioblastoma cells. Purinergic
Signal. 8:39–48. 2012. View Article : Google Scholar :
|
|
33
|
Singh SK, Clarke ID, Terasaki M, et al:
Identification of a cancer stem cell in human brain tumors. Cancer
Res. 63:5821–5828. 2003.PubMed/NCBI
|
|
34
|
Wu R, Zhang L, Hoagland MS and Swanson HI:
Lack of the aryl hydrocarbon receptor leads to impaired activation
of AKT/protein kinase B and enhanced sensitivity to apoptosis
induced via the intrinsic pathway. J Pharmacol Exp Ther.
320:448–457. 2007. View Article : Google Scholar
|
|
35
|
Wang C, Xu CX, Bu Y, Bottum KM and
Tischkau SA: Beta-naphthoflavone (DB06732) mediates estrogen
receptor-positive breast cancer cell cycle arrest through
AhR-dependent regulation of PI3K/AKT and MAPK/ERK signaling.
Carcinogenesis. 35:703–713. 2014. View Article : Google Scholar :
|
|
36
|
Molina JR, Hayashi Y, Stephens C and
Georgescu MM: Invasive glioblastoma cells acquire stemness and
increased Akt activation. Neoplasia. 12:453–463. 2010.PubMed/NCBI
|
|
37
|
Haas-Kogan D, Shalev N, Wong M, Mills G,
Yount G and Stokoe D: Protein kinase B (PKB/Akt) activity is
elevated in glioblastoma cells due to mutation of the tumor
suppressor PTEN/MMAC. Curr Biol. 8:1195–1198. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li J and Zhou BP: Activation of β-catenin
and Akt pathways by Twist are critical for the maintenance of EMT
associated cancer stem cell-like characters. BMC Cancer. 11:492011.
View Article : Google Scholar
|
|
39
|
Dubrovska A, Kim S, Salamone RJ, et al:
The role of PTEN/Akt/PI3K signaling in the maintenance and
viability of prostate cancer stem-like cell populations. Proc Natl
Acad Sci USA. 106:268–273. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang XQ, Ongkeko WM, Chen L, et al:
Octamer 4 (Oct4) mediates chemotherapeutic drug resistance in liver
cancer cells through a potential Oct4-AKT-ATP-binding cassette G2
pathway. Hepatology. 52:528–539. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Korkaya H, Paulson A, Charafe-Jauffret E,
et al: Regulation of mammary stem/progenitor cells by
PTEN/Akt/beta-catenin signaling. PLoS Biol. 7:e10001212009.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Bleau AM, Hambardzumyan D, Ozawa T, et al:
PTEN/PI3K/Akt pathway regulates the side population phenotype and
ABCG2 activity in glioma tumor stem-like cells. Cell Stem Cell.
4:226–235. 2009. View Article : Google Scholar : PubMed/NCBI
|