Introduction
Gastric cancer, one of the most common human
malignant tumors, accounts for the first leading cause of
gastrointestinal cancer-related mortality worldwide (1), particularly in China (2). Chemotherapy has been shown to increase
overall survival in patients with early gastric cancer after
surgery over the years, yet survival in advanced and metastatic
stages has not significantly improved (3).
Diallyl disulfide (DADS) is one of the organosulfur
compounds that derive from Allium vegetables, such as
garlic. Laboratory evidence has revealed that DADS exerts multiple
antitumor effects on a variety of tumors by suppression of cell
proliferation, angiogenesis or invasion, induction of apoptosis or
differentiation (4,5), without gross toxicity based on the
experiments in vivo (6,7).
Therefore, as a putative anticancer agent to inhibit cancer cell
growth and invasion, DADS has good prospects for clinical
application of adjuvant therapy.
Many studies have shown that diverse molecular
mechanisms are implicated in the suppressive effect of DADS on
tumor growth (4,5). In particular, a few studies suggest
that DADS inhibits cancer cell invasion through reducing the
expression and activities of matrix metalloproteinases (MMPs)
(8,9). However, the underlying mechanisms are
still largely unknown. Thus, potential molecules regulated by DADS
remain to be explored to elucidate its inhibitory effect on tumor
growth, particularly metastasis.
We have previously demonstrated that DADS induces
G2/M phase cell cycle arrest in gastric cancer cell lines via
ATR/Chk1/Cdc25C/cyclin B1 signaling pathway, accompanied with an
increase in Chk1 phosphorylation (10,11)
and a decrease in Cdc25C expression through activation of p38
(12). In addition, DADS induced
cell differentiation by downregu-lating ERK signaling pathway
(13). Our recent study showed that
DADS suppresses gastric cancer growth and induces apoptosis by
upregulation of miRNA-200b and miRNA-22 (14).
To further explore potential molecular targets
affected by DADS in gastric cancer, we screened and obtained some
proteins through proteomics research approaches, which showed
significantly differential expression between DADS-treated and
untreated gastric cancer cells in the present study. The expression
changes of retinoid-related orphan nuclear receptor α (RORα), nM23,
cyclin-dependent kinase-1 (CDK1), urokinase-type plasminogen
activator receptor (uPAR) and LIM kinase-1 (LIMK1) were verified in
subsequent experiments. We further explored the effects of DADS on
the ERK/Fra-1 pathway, as well as the invasion and
epithelial-mesenchymal transition (EMT)-associated protein
expression. We proposed that DADS-induced suppression of migration
and invasion of gastric cancer cells may be in part, due to
downregulation of the uPAR and the ERK/Fra-1 pathway.
Materials and methods
Reagents and antibodies
DADS (purity 80%, the remaining 20% being diallyl
trisulfide and diallyl sulfide), purchased from Fluka Co.
(Milwaukee, WI, USA), was dissolved in Tween-80 and stored at −20°C
after a 100-fold dilution with saline. The primary antibodies for
LIMK1 (ab81046), uPAR (ab89932), Fra-1 (ab117951), E-cadherin
(ab40772), MMP-9 (ab38898), TIMP-3 (ab39184) and vimentin (ab92547)
were purchased from Abcam (Abcam, Cambridge, UK). The antibody
against uPA (17968- 1-AP) was purchased from ProteinTech
(ProteinTech Group, Chicago, IL, USA). The primary antibodies for
RORα (sc-6062), nM23-H1 (sc-343), CDK1 (sc-54), ERK1/2 (sc-135900),
p-ERK1/2 (sc-292838) and β-actin (sc-47778) and horseradish
peroxidase (HRP)-conjugated secondary antibodies were purchased
from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Cell culture and cell line
establishment
Human gastric cancer cell line MGC-803 was obtained
from the Cancer Research Institute, Xiangya Medical College, Center
South University in China. The cells were cultured in RPMI-1640
medium (Gibco, Life Technologies, Vienna, Austria), supplemented
with 10% heat-inactivated fetal bovine serum (FBS; Life
Technologies, USA), 100 μg/ml streptomycin, and 100 U/ml
penicillin G (Invitrogen, Carlsbad, CA, USA) in a humidified
atmosphere of 5% CO2 and 95% air at 37°C. MGC-803 cells
(2×106) in the logarithmic growth phase were untreated
or treated with 30 mg/l DADS. To establish the stable
uPAR-interfering cell lines, three pcDNA™6.2-GW/EmGFPmiR
uPAR-microRNA (miR)-expressing plasmids were constructed by
Invitrogen Corporation. Sequences of DNA oligomers inserted into
pcDNA™6.2-GW/EmGFPmiR were as follows: miR1 sense, 5′-TGC TGA AGT
CTA ACC CAC ACA CAA CCG TTT TGG CCA CTG ACT GAC GGT TGT GTG GGT TAG
ACT T-3′ and antisense, 5′-CCT GAA GTC TAA CCC ACA CAA CCG TCA GTC
AGT GGC CAA AAC GGT TGT GTG TGG GTT AGA CTT C-3′; miR2 sense,
5′-TGC TGA TAG GTG ACA GCC CGG CCA GAG TTT TGG CCA CTG ACT GAC TCT
GGC CGC TGT CAC CTA T-3′ and antisense, 5′-CCT GAT AGG TGA CAG CGG
CCA GAG TCA GTC AGT GGC CAA AAC TCT GGC CGG GCT GTC ACC TAT C-3′;
miR3 sense, 5′-TGC TGA ATG CAT TCG AGG TAA CGG CTG TTT TGG CCA CTG
ACT GAC AGC CGT TAT CGA ATG CAT T-3′ and antisense, 5′-CCT GAA TGC
ATT CGA TAA CGG CTG TCA GTC AGT GGC CAA AAC AGC CGT TAC CTC GAA TGC
ATT C-3′; MGC-803 cells were transfected with uPAR-miR expressing
plasmid and empty vector (control) using Lipofectamine 2000 reagent
(Invitrogen), following the instructions of the manufacturer. The
transfected cells were selected with blasticidin (Invitrogen) until
the stable transgene expression during culture maintenance. The
expression levels of uPAR in stable cell lines were evaluated by
reverse transcription-polymerase chain reaction (RT-PCR) and
western blot analysis to confirm the knockdown efficacy.
Preparation of protein samples for
two-dimensional electrophoresis (2-DE)
DADS-treated and -untreated cells were harvested and
lysed in lysis buffer containing 8 M urea, 4% CHAPS, 40 mM Tris, 1%
DDT, 0.5% immobilized pH gradient (IPG) buffer (all from Amersham
Biosciences) and protease inhibitor mixture (Roche Molecular
Biochemicals). The samples were mixed and incubated on ice for 30
min and then centrifuged at 15,000 rpm for 30 min. The supernatant
was transferred to a fresh tube and stored in −80°C until use. The
total protein concentration was measured according to the Bradford
protein assay method with bovine serum albumin as standard.
2-DE
2-DE was carried out to separate proteins from
DADS-treated and -untreated cells of three independent experiments.
Protein (200 μg) was taken from each extract then dissolved
in a 320 μl rehydration solution (8 M urea, 4% CHAPS, 65 mM
DDT, 0.5% IPG buffer and a trace of bromo-phenol blue) and placed
at room temperature for 40 min, then loaded to 18 cm IPG strips
containing a wide-range of pH gradient 3–10 (Amersham Biosciences).
Rehydration and isoelectric focusing (IEF) were performed at 20°C
for 12 h at 50 V, followed by 50 μA/IPG strip, 500 V for 1
h, 1,000 V for 1 h, 8,000 V for 4 h 30 min using an IPGphor
(Amersham Biosciences). Following IEF, the IPG strips were
equilibrated with equilibration buffer (6 M urea, 2% sodium dodecyl
sulfate, 50 mM Tris-HCl, pH 8.8, 30% glycerol and 1% DTT) for 15
min, and then they were put in the same solution, with 2.5%
iodoacetamide replacing DTT, for an additional 15 min with gentle
shaking. The equilibrated strips were transferred to the top of
12.5% SDS PAGE gels and embedded in 1% low-melting agarose
containing a trace of bromophenol blue. Electrophoresis was carried
out using Ettan DALTsix system (GE Healthcare) at 25 w/gel for 30
min, and then 17 w/gel until the bromophenol blue dye reached the
bottom of the gel. Then, the gels were fixed (40% ethanol, 10%
acetic acid) and silver stained using the PlusOne silver staining
kit (Amersham Biosciences) as described in the instructions of the
manufacturer.
Gel imaging and spot analysis
Gels were scanned using the Molecular Imager Gel Doc
XR System and analyzed using the PDQuest system (both from
Bio-Rad), according to the protocols provided by the manufacturer.
Spot intensities were quantified by calculation of spot volume
after normalization of the image using the total spot volume
normalization method. Protein spots showing at least a 2.0-fold
difference in abundance between control and treatment were
considered as differentially expressed. The selected protein spots
that were statistically analyzed by the Student’s t-test with
P<0.05 were considered to indicate a statistically significant
result.
Mass spectrometry analysis
The excised spots of interest were destained and
digested as previously described (15). Briefly, protein spots were destained
with 15 mM K3[Fe(CN)6] and 50 mM
Na2S2O3. Gel pieces were incubated
in the digestion solution with 20 μg/ml trypsin (Amresco)
for 10–12 h at 37°C. The tryptic peptide mixture was mixed with
α-cyano-4-hydroxycinnamic acid matrix solution (Sigma-Aldrich, St.
Louis, MO, USA). One microliter of the mixture was analyzed by the
Voyager System DE-STR 4307 matrix-assisted laser
desorption/ionization time of flight mass spectrometer
(MALDI-TOF-MS) (Applied Biosystems, Foster City, CA, USA). Protein
identification was performed by matching the peptide mass
fingerprints against the Swiss-Prot or the NCBInr database with the
MS-Fit search program (http://prospector.ucsf.edu/). Molecular weight (Mr)
and isoelectric point (pI) of the identified proteins were
subsequently compared with the experimental values obtained from
the 2-DE image analysis.
Reverse transcription-polymerase chain
reaction (RT-PCR)
Total RNA was extracted from the cells using TRIzol
reagent (Invitrogen). Reverse transcription was carried out using
the RT-PCR system and PCR analysis was performed using the GeneAmp
PCR kit (Promega, Madison, WI, USA). Primer sequences were as
follows: RORα F, 5′-GTC AGC AGC TTC TAC CTG GAC-3′ and R, 5′-GTG
TTG TTC TGA GAG TGA AAG GCA CG-3′; nM23 F, 5′-AGG GCA GAC CAC ATT
GCT TTT C-3′ and R, 5′-GCT GGG AGG AAG CAT TTT TAA TC-3′; β-actin
F, 5′-TCT ACA ATG AGC TGC GTG TGG-3′ and R, 5′-GGA ACC GCT CAT TGC
CAA TG-3′. For LIMK1 F, 5′-GGG GCA TCA TCA AGA GCA-3′ and R, 5′-CCA
GGC AGT TGT GGG AGT-3′; β-actin F, 5′-ACA CTG TGC CCA TCT ACG AGG
GG-3′ and R, 5′-ATG ATG GAG TTG AAG GTA GTT TCG TGG AT-3′. For uPAR
F, 5′-TCA CCT ATT CCC GAA GCC GTT AC-3′ and R, 5′-AGA GTG AGC GTT
CGT GAG TGC C-3′; β-actin F, 5′-CCC ACA CTG TGC CCA TCT AC-3′ and
R, 5′-GCC GGA CTC GTC ATA CTC CT-3′. For CDK1 F, 5′-AGT CAG TCT TCA
GGA TGT GCT-3′ and R, 5′-TGA CAT GGG ATG CTA GGC TT-3′; β-actin F,
5′-TGG CAT CCA CGA AAC TAC CT-3′ and R, 5′-TCA CCT TCA CCG TTC CAG
TT-3′. The PCR products were analyzed on a 2% agarose gel
containing ethidium bromide. The densitometric quantitation of the
products was determined using Lab-work analysis software. The ratio
of target gene to β-actin was calculated to obtain the relative
fold-changes in gene expression. A DL1000 DNA ladder and a 100 bp
DNA ladder were used as the markers (Takara, Dalian, China).
Western blot analysis
For the total protein extraction, the cells were
lysed on ice for 30 min in 0.1 ml lysis buffer [10 mmol/l Tris-HCl
(pH 7.6), 100 mmol/l NaCl, 1 mmol/l EDTA (pH 8.0), 100 μg/ml
PMSF and 1 μg/ml aprotinin]. The cell lysates were
centrifuged at 12,000 rpm for 10 min, and the protein contents were
determined by the BCA protein assay kit (Pierce, Rockford, IL,
USA). The protein extracts were resolved on 12% SDS-polyacrylamide
gels for electrophoresis and transferred onto the PVDF membranes.
The membranes were incubated with 5% skim milk in Tris-buffered
saline (TBS) containing 0.1% Tween-20 for 2 h at room temperature
and then they were incubated with the primary antibody at 4°C
overnight. After being washed with TBS-T, the membranes were then
incubated with the HRP-conjugated secondary antibody
(1:1,000–2,000). The membranes were developed by an enhanced
chemiluminescence plus kit (Amersham Biosciences, Buckinghamshire,
UK) and bands were visualized on X-ray film (Kodak). The target
protein amounts were normalized towards β-actin quantity using
densitometry, and relative fold-changes in protein levels were
calculated.
Migration assays
Cell migration was evaluated using the scratch wound
assay as previously described (16). Briefly, the cells were cultured to
90% confluence in a 6-well plate and washed with serum-free
RPMI-1640 medium. The cell monolayers were wounded with a
10-μl plastic pipette tip. After, the cells were untreated
or treated with 30 mg/l DADS, the wound areas were photographed
using an inverted microscope. The distance that the cells migrated
from the wound edge was measured and a mean value of five
independent microscope fields was calculated. The experiments were
repeated three times. Migration rates were expressed as ratios
between treated vs. control group values or between 24 vs. 0 h
group values.
Invasion assays
Invasion assays were carried out using
Transwell® (Corning, Corning, NY, USA) as previously
described (16). Matrigel (25
μg) (Becton-Dickinson, Bedford, MA, USA) dilution was added
to the upside of porous filters (pore size, 8-μm) and
allowed to gel at 37°C overnight. After the coated filters were
rehydrated with 100 μl medium, 1×105 cells in 200
μl serum-free medium supplemented with 0.2% bovine serum
albumin were seeded into the upper part of each chamber, whereas
the lower compartments were filled with 500 μl of culture
medium. The cells were treated or untreated with 30 mg/l DADS. The
invaded cells on the lower membrane surface were fixed with 4%
paraformaldehyde and stained with 0.1% crystal violet. The
experiments were repeated three times. Invasiveness was determined
by counting an average number of cells from four microscopic
fields/well, and invasion rates were expressed as ratios between
treated vs. control group values, or between 24 vs. 0 h group
values.
Statistical analyses
All the results are presented as mean ± SD or SE of
three independent experiments. The Student’s t-test and the one-way
ANOVA were used to identify statistical significance. A two-sided
P<0.05 was considered to indicate a statistically significant
result.
Results
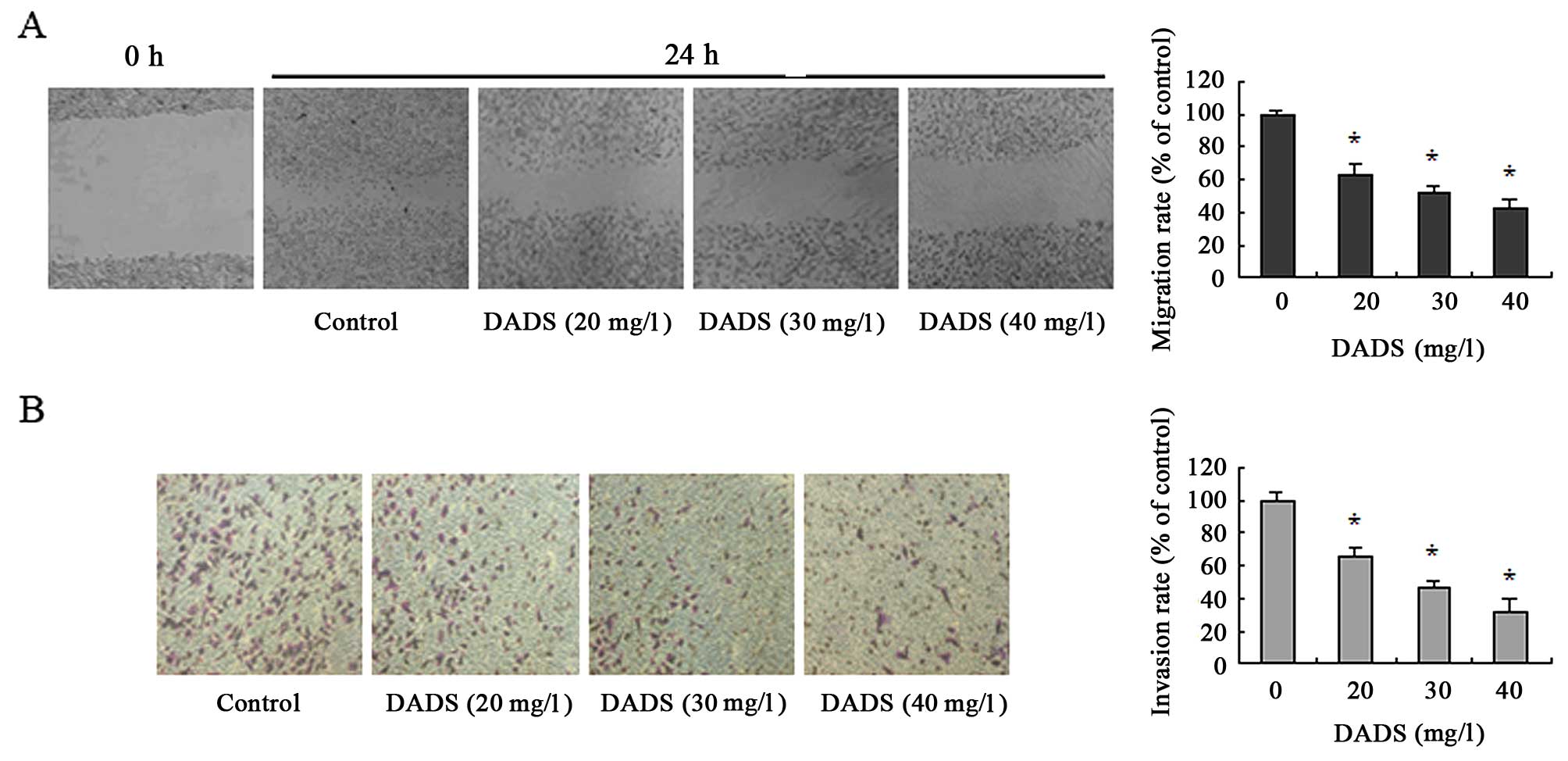
DADS inhibits MGC-803 cell migration and
invasion
We first treated cells with 20, 30 and 40 mg/l DADS
for 24 h, and examined the effects of DADS on migration and
invasion of the gastric cancer MGC-803 cell line. As shown in
Fig. 1A and B, DADS reduced
migration and invasion rates of the cells in a
concentration-dependent manner. These data indicate that treatment
with DADS significantly inhibited migration and invasion activities
of MGC-803 cells. Our previous studies have demonstrated that 30
mg/l DADS is able to cause the arrest of the G/M phase of the cell
cycle and the growth inhibition of the gastric cancer cells
(10) based on IC50
values for DADS (17). Thus,
incubation with 30 mg/l DADS was used in the subsequent
experiments.
Identification of differential expression
proteins by 2-DE and MS
To analyze the differential expression proteins
induced by DADS in gastric cancer MGC-803 cells, the cells were
treated with 30 mg/l DADS for 24 h. The proteins from DADS-treated
and untreated cells were resolved by 2-DE respectively, and the
gels were stained with silver to visualize the protein spots in the
2-DE gels. Two representative 2-DE maps from DADS-treated and
untreated group are shown in Fig. 2A
and B, respectively. By comparing the ratio of each spot in the
gels, 291 protein spots that had consistent differences (≥2-fold)
between DADS-treated group and the untreated group in triplicate
experiments were considered as differentially expressed. Among
them, 23 selected differential protein spots were excised from the
stained gels, subjected to trypsin digestion. Twenty-three
differentially expressed proteins were identified based on
MALDI-TOF-MS and database query with MS-Fit (Table I). These protein spots are marked
with arrows in Fig. 2A and B.
Compared with the untreated group, the expression levels of 9
proteins were increased, whereas the expression levels of the other
14 proteins were decreased in DADS-treated group.
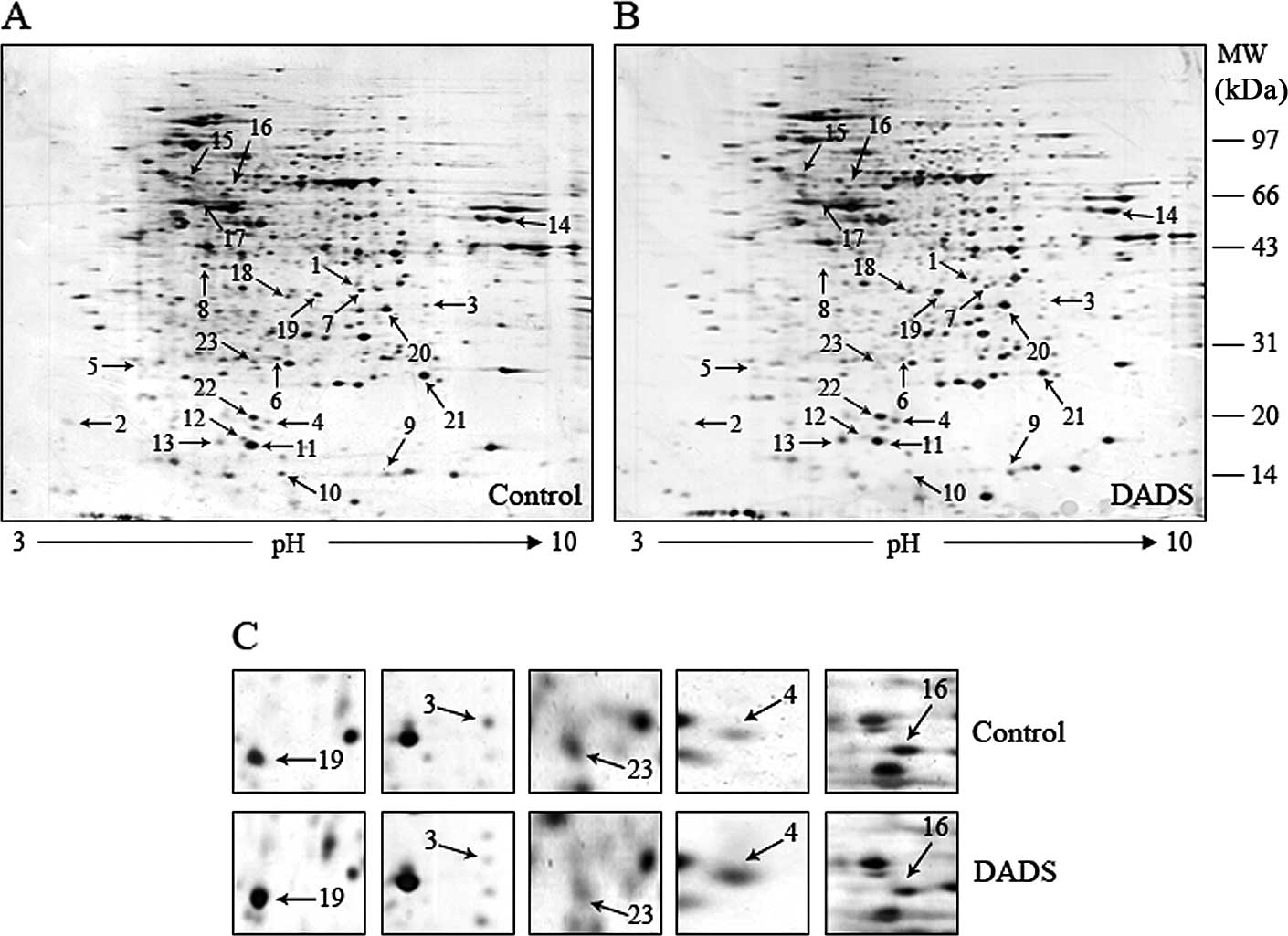 | Figure 2Two-dimensional gel analysis and
MALDI-TOF-MS identification of differential expression proteins
between untreated and DADS-treated MGC-803 cells. After the cells
were treated or untreated with 30 mg/l DADS for 24 h, proteins were
extracted and separated by 2-DE. Twenty-three protein spots showing
significantly differential expression were identified using
MALDI-TOF-MS, and are marked with arrows. Representative 2-DE maps
of (A) untreated and (B) DADS-treated MGC-803 cells are shown. (C)
Images of five differential expression protein spots, spot 3
(uPAR), spot 4 (nM23), spot 16 (LIMK1), spot 19 (RORα) and spot 23
(CDK1), are shown between untreated and DADS-treated MGC-803 cells.
MALDI-TOF-MS, matrix-assisted laser desorption/ionization time of
flight mass spectrometer; DADS, diallyl disulfide; 2-DE,
two-dimensional electrophoresis; uPAR, urokinase-type plasminogen
activator receptor; LIMK1, LIM kinase-1; RORα, retinoid-related
orphan nuclear receptor α; CDK1, cyclin-dependent kinase-1. |
 | Table IDifferential expression proteins
induced by DADS in MGC803 cells. |
Table I
Differential expression proteins
induced by DADS in MGC803 cells.
| Spot no. | Accession
no.a | Protein name | Mr (Da)/pI | Matched
peptides | Sequence coverage
(%) | Expression
alterationb |
|---|
| 1 | Q30136 | HLA-DRB1
protein | 24901.0/7.22 | 8 | 46 | Upregulated |
| 2 | O60522 | Antigen
NY-CO-45 | 85665.9/4.46 | 12 | 21 | Downregulated |
| 3 | Q03405 | Urokinase-type
plasminogen activator receptor | 31263.6/6.01 | 8 | 26 | Downregulated |
| 4 | P15531 | Metastasis
inhibition factor nm23 | 20411.6/7.06 | 10 | 51 | Upregulated |
| 5 | O00429 | Dynamin-1-like
protein | 81877.6/6.37 | 19 | 25 | Downregulated |
| 6 | O60602 | Toll-like receptor
5/Toll/interleukin-1 receptor-like protein 3 | 97692.0/6.21 | 10 | 17 | Downregulated |
| 7 | Q92782 | Zinc finger protein
neuro-d4 | 39875.8/6.28 | 7 | 28 | Downregulated |
| 8 | Q08499 | Phosphodiesterase
4D | 76466.9/4.86 | 8 | 15 | Downregulated |
| 9 | P98088 | Mucin-5AC/gastric
mucin | 91762.7/6.85 | 35 | 40 | Upregulated |
| 10 | Q86XL3 | Ankyrin repeat and
LEM domain-containing protein 2 | 87454.5/6.37 | 21 | 29 | Downregulated |
| 11 | P98170 | X-linked inhibitor
of apotosis protein | 56701.3/6.22 | 7 | 26 | Downregulated |
| 12 | P20062 |
Transcobalamin-2 | 47535.3/6.52 | 11 | 25 | Downregulated |
| 13 | P33981 | Dual specificity
protein kinase TTK | 95279.9/8.66 | 26 | 31 | Upregulated |
| 14 | P40926 | Malate
dehydrogenase | 35531.5/8.92 | 8 | 35 | Upregulated |
| 15 | P52597 | Heterogeneous
nuclear ribonucleoprotein F | 45672.1/5.38 | 6 | 23 | Downregulated |
| 16 | P53667 | LIM kinase-1 | 83045.8/6.05 | 7 | 14 | Downregulated |
| 17 | Q14934 | Nuclear factor of
activated T-cells | 95473.3/5.29 | 11 | 19 | Downregulated |
| 18 | Q99676 | Zinc finger protein
184 | 83269.5/8.50 | 12 | 26 | Upregulated |
| 19 | P35398 | Nuclear receptor
RORα | 63036.2/5.97 | 10 | 16 | Upregulated |
| 20 | P01850 | T-cell receptor β-1
chain C region | 34440.2/6.23 | 5 | 31 | Upregulated |
| 21 | P78536 | TNF-α-converting
enzyme | 92961.5/5.59 | 9 | 14 | Downregulated |
| 22 | Q9UNN8 | Endothelial protein
C receptor | 26641.6/6.70 | 5 | 33 | Upregulated |
| 23 | P06493 | Cyclin-dependent
kinase 1/CDC2 | 27502.9/6.54 | 5 | 11 | Downregulated |
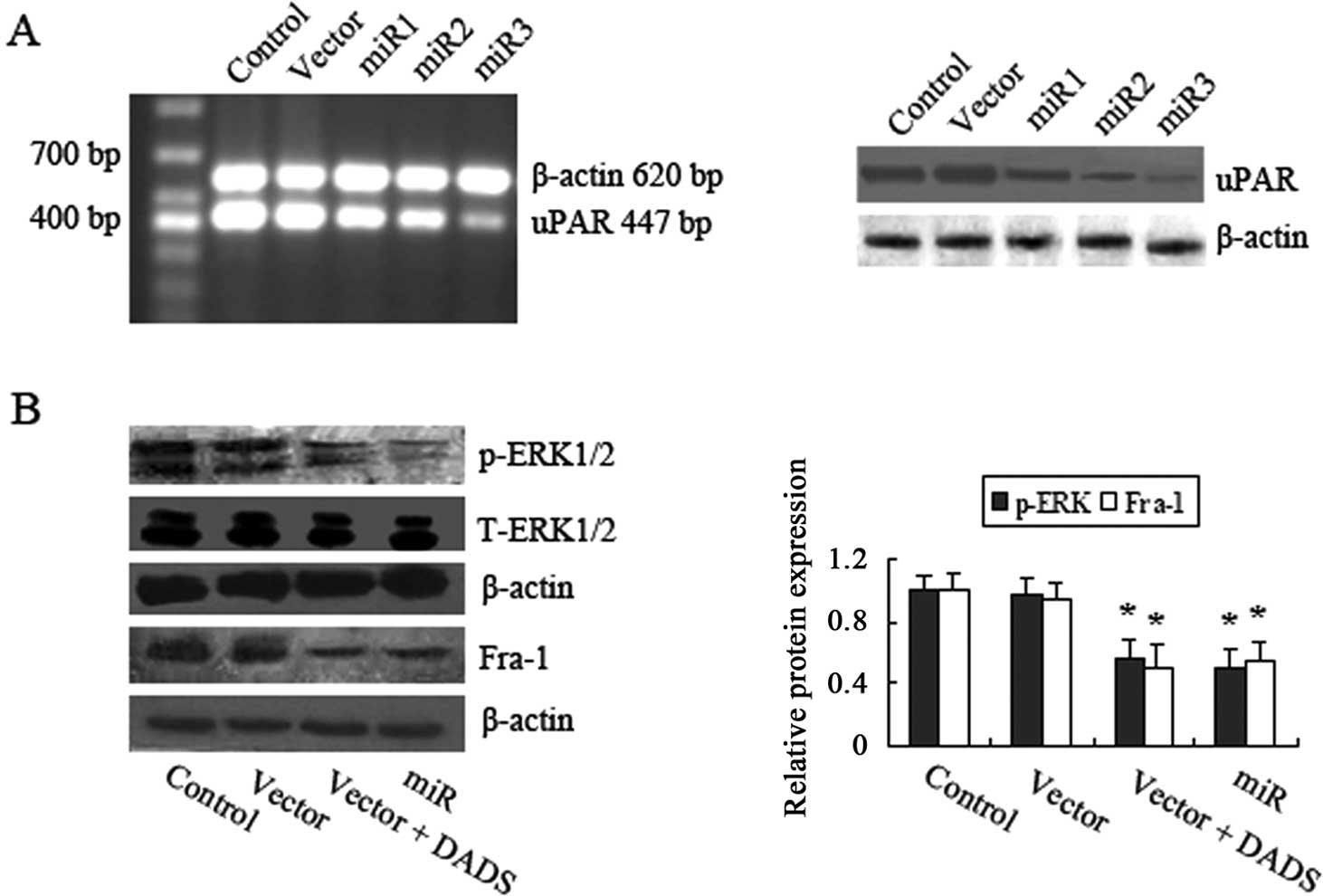
Verification of DADS-induced differential
expression genes in MGC-803 cells
Among the identified proteins, RORα, nM23, LIMK1,
uPAR and CDK1 displayed significantly differential expression in
DADS-treated cells compared with the untreated cells. A
representative comparison of the five proteins is shown in Fig. 2C. These five proteins were subjected
to further validation. RT-PCR and western blot analysis were
performed to explore the expression pattern of these five genes
after the MGC-803 cells were exposed to 30 mg/l DADS for various
times. As shown in Fig. 3A, the
mRNA levels of RORα and nM23 increased in a time-dependent manner.
Whereas, LIMK1, uPAR and CDK1 mRNA levels decreased in a
time-dependent manner. Accordingly, similar changes in the protein
levels were also observed in the MGC-803 cells upon DADS treatment
(Fig. 3B). Therefore, changes in
the gene expression induced by DADS were confirmed at the
transcriptional and translational levels, which were consistent
with the results from the comparative proteomics research.
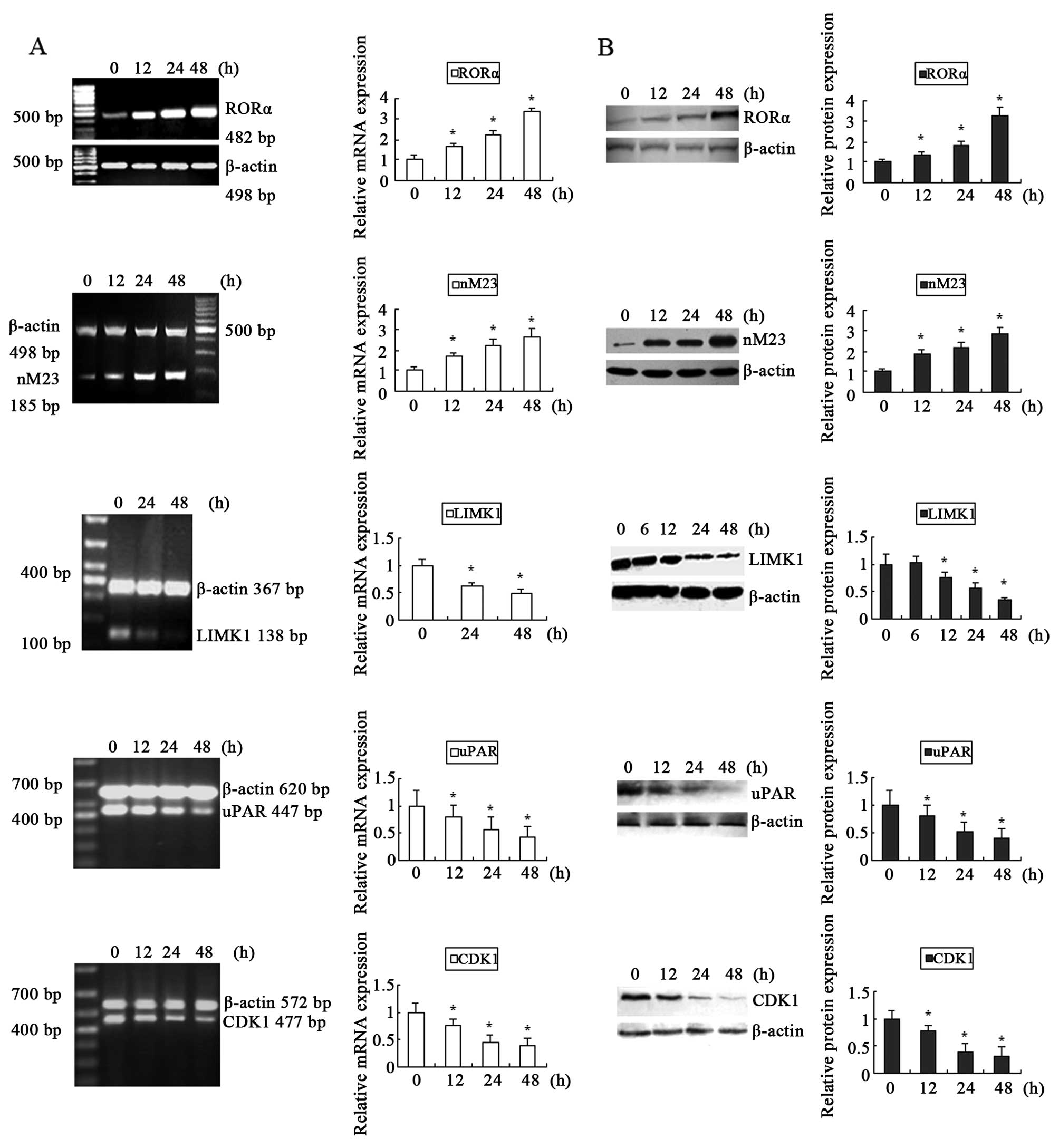 | Figure 3The genes with differential
expression induced by DADS in the MGC-803 cells. The cells were
exposed to 30 mg/l DADS for indicated times. (A) The mRNA
expression levels of RORα, nM23, LIMK1, uPAR and CDK1 were detected
by RT-PCR. β-actin was used as an internal control for
normalization. (B) Western blot analysis was used to determine the
RORα, nM23, LIMK1, uPAR and CDK1 protein levels. β-actin was used
as a loading control. The relative fold-changes in mRNA or the
protein level compared with the controls were calculated.
*P<0.05 vs. control. DADS, diallyl disulfide; RORα,
retinoid-related orphan nuclear receptor α; LIMK1, LIM kinase-1;
uPAR, urokinase-type plasminogen activator receptor; CDK1,
cyclin-dependent kinase-1. |
DADS inhibits the ERK/Fra-1 pathway
through decreasing uPAR expression in MGC-803 cells
uPAR activates diverse cell signaling pathways,
including the ERK/Fra-1 pathway. We hypothesized that the decreased
expression of uPAR induced by DADS may cause inhibition of the
ERK/Fra-1 pathway. To test this hypothesis, we first constructed
several stable uPAR-interfering MGC-803 cell lines using microRNA
(miR), and then chose miR3, which showed the most knockdown
efficacy, for the following experiments (Fig. 4A). Then, we determined the levels of
phosphorylated ERK1/2 in DADS-treated and uPAR-interfering cells.
As anticipated, knockdown of uPAR resulted in decreased levels of
p-ERK1/2 (Fig. 4B). The levels of
Fra-1 protein were also downregulated by DADS and knockdown of uPAR
(Fig. 4B). These data suggest that
downregulation of uPAR by DADS inhibited ERK1/2 activities and
Fra-1 expression, leading to blockage of ERK/Fra-1 pathway.
Downregulation of uPAR by DADS affects
invasion and EMT-associated protein expression in MGC-803
cells
Activation of the ERK/Fra-1 pathway is involved in
positively regulating cancer cell invasion and EMT-related genes,
such as uPA and MMP-9. Fig. 5A and
B showed that uPA (uroki-nase-type plasminogen activator) and
MMP-9 expression were attenuated by DADS, accompanied by
downregulation of uPAR. Moreover, similar results were observed in
the uPAR-interfering cells. Moreover, DADS upregulated TIMP-3
(tissue inhibitor of metalloproteinase-3) expression (Fig. 5C). Next, we examined the expression
levels of vimentin and E-cadherin, which were taken as markers of
EMT. Both DADS and knockdown of uPAR decreased the vimentin and
increased the E-cadherin expression (Fig. 5B and C). Moreover, knockdown of uPAR
is able to augment these effects induced by DADS (Fig. 5A–C).
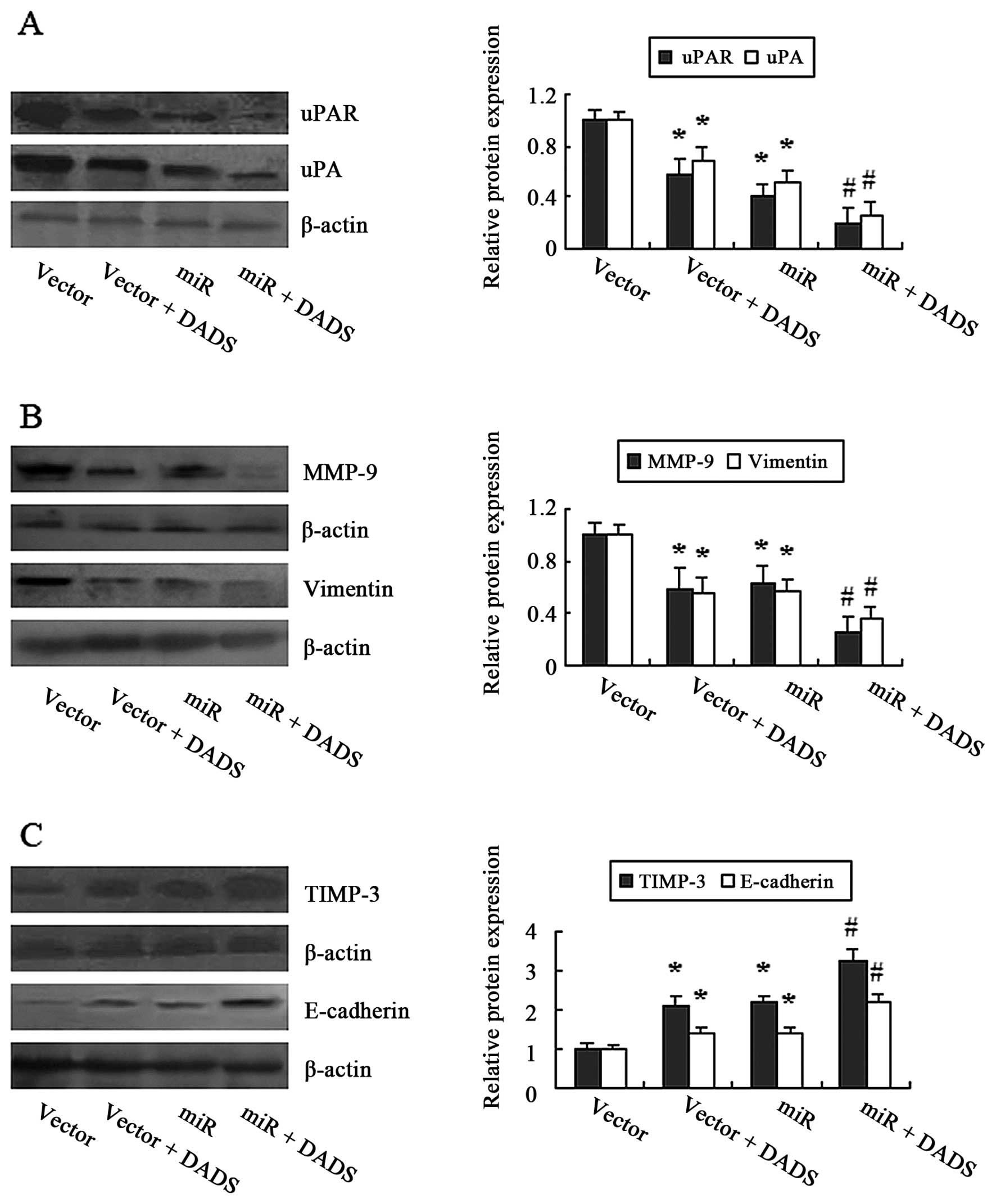 | Figure 5Downregulation of uPAR by DADS
affects invasion and EMT-associated protein expression in MGC-803
cells. The cells transfected with uPAR-miR or the empty vector were
treated or untreated with 30 mg/l DADS for 24 h. The expression of
uPAR, (A) uPA and MMP-9, (B) vimentin and TIMP-3, (C) and
E-cadherin proteins were determined by western blot analysis.
β-actin was used as a loading control. The relative fold-changes in
the protein level compared with the empty vector groups (controls)
were calculated. *P<0.05 vs. control,
#P<0.05 vs. control, the uPAR-miR or the DADS group.
uPAR, urokinase-type plasminogen activator receptor; DADS, diallyl
disulfide; EMT, epithelial-mesenchymal transition; uPA,
urokinase-type plasminogen activator; MMP-9, matrix
metalloproteinase-9; TIMP-3, tissue inhibitor of
metalloproteinase-3. |
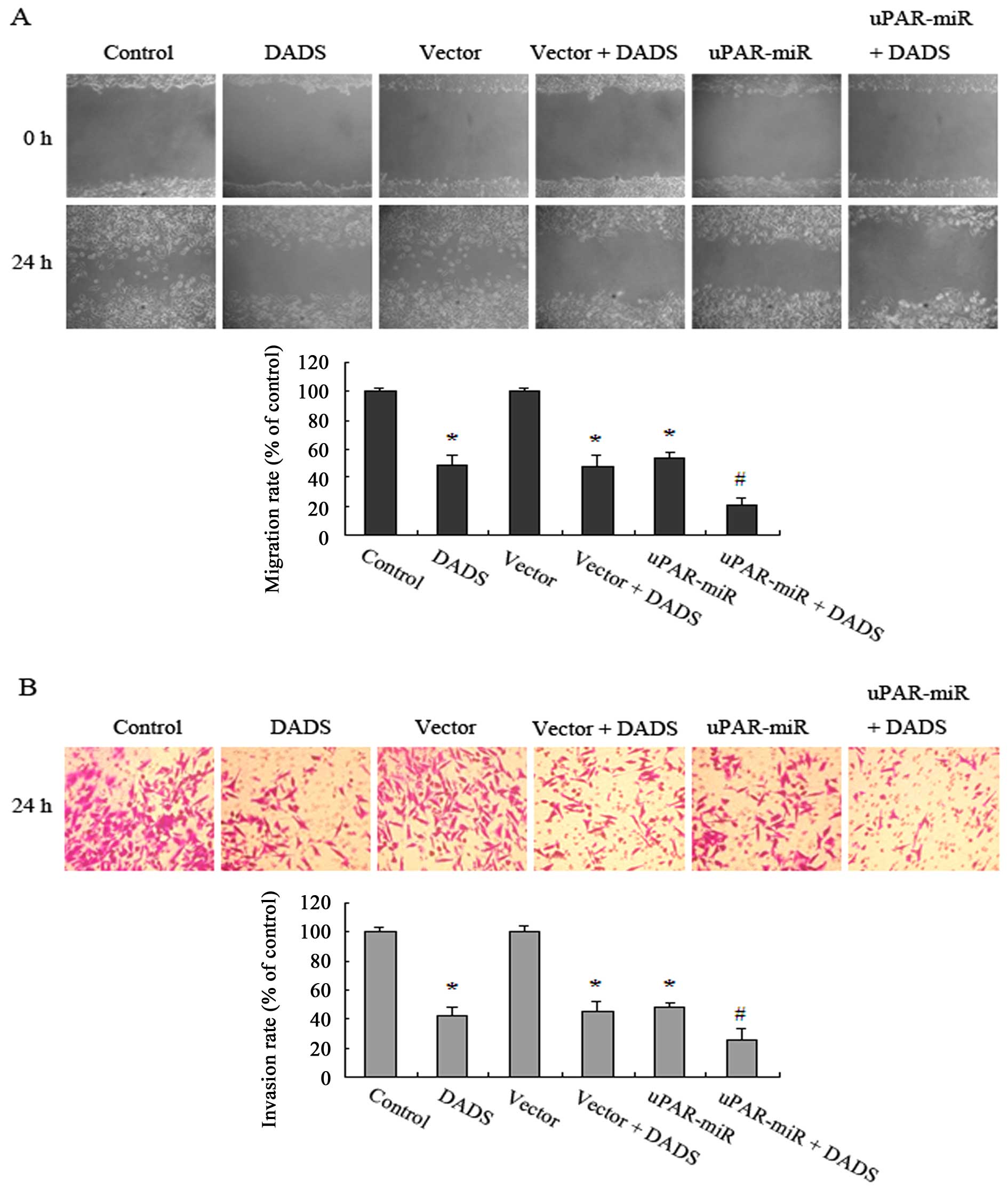
Downregulation of uPAR potentiates
inhibitory effects of DADS on MGC-803 cell migration and
invasion
To test whether DADS-induced downregulation of uPAR
is responsible for inhibition of MGC-803 cell migration and
invasion, we performed the scratch wound and invasion assays. The
results showed that the migration rates of the cells after uPAR
knockdown were obviously reduced in contrast to the vector group,
as well as that of the DADS-treated cells in contrast to the
control group (Fig. 6A). Moreover,
both DADS-treated and uPAR knockdown cells exhibited a significant
decrease in the invasion rates (Fig.
6B). In addition, stronger inhibitory effects on cell migration
and invasion were found in the uPAR-interfering cells exposed to
DADS for 24 h (Fig. 6A and B).
Discussion
In the present study, we identified differentially
expressed proteins induced by DADS in gastric cancer cells using
the comparative proteomics technique. Among them, RORα, nM23,
LIMK1, uPAR and CDK1 attracted our attention since dysregulation of
their expression and function is closely correlated with
tumorigenesis and progression, particularly invasion and
metastasis. The effects of DADS on these gene expression have not
yet been reported in the gastric cancer cells.
RORα is a member of the orphan nuclear factor family
that regulates gene expression by binding to ROR response elements.
Its decreased expression is shown in cancers, suggesting that RORα
inactivation may be one mechanism underlying tumorigenesis
(18). Activation of RORα
suppresses the expression of Wnt/β-catenin target genes, resulting
in inhibition of proliferation and migration of colon cancer cells
(19). RORα inhibits breast cancer
cell invasion by transactivation of SEMA3F, a suppressive
microenvironmental factor (20).
Thus, RORα has been proposed to act as a tumor suppressor. In
gastric cancer, methylation silence of the RORα gene may be one
reason for its inactivation (21).
Our previous studies demonstrated that DADS could upregulate p21
expression through augmenting histone acetylation (22), we speculate that this epigenetic
mechanism may be implicated in DADS-induced RORα expression in
MGC-803 cells, and upregulation of RORα by DADS may contribute to
its bioactivity against gastric cancer cell growth and
invasion.
Nm23 is characterized as a specific metastasis
suppressor ascribed to its multiple activities involving regulation
of tumor metastatic process. The decreased Nm23 expression
facilitates tumor cell motility and invasion, while its
overexpression in aggressive cancer cells suppresses their
metastatic capacity (23). Nm23
silencing promotes invado-podia formation, activation of
pro-invasive signaling pathways and upregulation of MMPs (24). Whereas, re-expression of Nm23
attenuates metastatic ability of cancer cells by altering the
expression of metastasis-related genes (25,26).
Therefore, effort has been made to develop agents to recover Nm23
function in cancer cells. For an example, lycopene reduces
experimental tumor metastasis in vivo via increasing Nm23
protein expression (27). It has
been reported that the decreased Nm23 protein level was observed in
lymph node and liver metastases in contrast to primary lesions of
gastric cancer (28). In the
present study, our data indicated that DADS upregulates Nm23
expression in gastric cancer cells. It suggests that restoration of
nm23 bioactivity may be a mechanism underlying DADS anti-invasion
effect on gastric cancer cells.
LIM kinases (LIMK1 and LIMK2) are the common
downstream effectors of Rho-ROCK, Rac-PAK and Cdc42-MRCK pathways
that regulate rearrangements of actin cytoskeleton. Functional
dysregulation of LIMKs, due to their overexpression and
overactivation, in particular LIMK1 (29,30),
promotes the formation and maturation of matrix-degrading
invadopodia, which facilitate cancer cell invasion (31). Whereas, knockdown of LIMK1
expression blocks invadopodia formation, and reduces MMP-9
expression and matrix protein degradation, decreasing the invasive
potential of cancer cells (31).
Our previous study demonstrated that the suppressive effects of
DADS on colon cancer cell motility and invasiveness are associated
with attenuating Rac1-Rock1/Pak1-LIMK1 signaling pathway (32). Thus, LIMK1 may be a candidate target
for DADS against gastric cancer cell invasion.
uPAR is a glycolipid-anchored cell surface receptor.
Its elevated expression indicates poor prognosis in many human
tumors, including gastric cancer (33). In addition to mediating the
degradation of extracellular matrix by its interaction with uPA to
activate plasminogen and MMPs, uPAR interacts with co-receptors to
activate intracellular signaling pathways (34). Furthermore, overactivation of
uPAR-dependent signaling pathways fosters EMT and invasion, and
these cellular changes are blocked by silencing uPAR (35) or uPA (36) expression. Since various signaling
pathways are involved in inducing uPAR expression, which in turn
stimulates gastric cancer cell invasiveness, uPAR has been
considered as a potential therapeutic target (37). It has been demonstrated that ERK
activation promotes the expression of the AP-1 family transcription
factor Fra-1, which is essential for cell invasion through
modulating invasion-associated gene expression, such as uPA and
MMP-9 (38). The present study
showed that DADS reduced uPAR expression in the MGC-803 cells,
suggesting that a decrease in uPAR protein may lead to suppression
of its downstream effectors of ERK/Fra-1, resulting in inhibition
of cell migration and invasion. To testify this assumption, we
examined effects of DADS on ERK/Fra-1 pathway and its target gene
expression in MGC-803 cells. Our data showed that DADS decreased
phosphorylation of ERK1/2 and Fra-1 expression, concomitant with
the decreased expression of uPA and MMP-9. Additionally, DADS
increased the expression of TIMP-3, which inhibits the catalytic
activity of MMPs to maintain extracellular matrix (ECM)
homeostasis. These changes resulting from DADS-induced
downregulation of uPAR were supported by the similar results that
were observed in uPAR knockdown cells. Notably, we found that DADS
downregulates vimentin and upregulates E-cadherin expression, and
these results were consistent with the observations in the uPAR
knockdown cells. Recent researches indicated that overactivation of
ERK/Fra-1 pathway induces EMT (39,40),
and Fra-1 is involved in regulation of the expression of vimentin
(41) and E-cadherin (42). Based on these findings, we proposed
that DADS may inhibit EMT in gastric cancer cells, which may
partially account for its inhibitory effect on MGC-803 cell
invasion. Notably, DADS is able to exert more potent effects on
uPAR knockdown cells. We speculate that DADS-induced invasion
inhibition may involve other mechanisms that remain to be revealed
except for downregualtion of uPAR, such as inhibition of LIMK1 and
upregulation of RORα and Nm23.
CDK1 plays a critical role in promoting the
transition of the G2/M phase of the cell cycle through the
formation of a complex between CDK1 and cyclin B1 or cyclin A. Its
functional dysregulation results in tumor cell unlimited
proliferation. Inhibition of CDK1 expression and activity is an
effective approach to suppress tumor growth and sensitize cancer
cells to apoptosis (43,44). We previously reported that the
decrease of CDK1 expression and its interaction with Mcl1 (an
anti-apoptotic factor) are involved in the DADS-induced G2/M phase
arrest in human leukemia HL-60 cells (45). In the present study, we revealed
that DADS reduces CDK1 expression in gastric cancer MGC-803 cells.
It suggests that the inhibitory effect of DADS on CDK1 may
contribute to the treatment of gastric cancer, since gastric tumor
tissues show higher CDK1 expression and activity than
non-neoplastic mucosa (46).
In summary, we revealed that DADS is able to
increase RORα and nM23 expression and decrease LIMK1, uPAR and CDK1
expression. DADS-induced downregulation of uPAR is able to
partially account for inhibition of the ERK/Fra-1 pathway, and
migration and invasion of gastric MGC-803 cells. These findings are
valuable for revealing potential targets for DADS, but further
studies are required to demonstrate its anti-gastric cancer
mechanisms.
Acknowledgments
The present study was supported by the National
Natural Scientific Foundation of China (nos. 81374013, 31201027,
31100935, 81102854, 31000629 and 81100375), the Key Project
Foundation of the Science and Technology program of Hunan Province
of China (no. 04SK1004), the Patency Foundation of Innovation
Platform of Hunan Provincial University of China (no. 09K074), the
Key Project of Scientific Research Foundation of Hunan Province
Education Department of China (no. 09A077), the Key Hunan
Provincial Natural Science Foundation of China (no. 07JJ3033) and
the Construct Program of the Key Discipline in Hunan Province of
China (no. 2011–76).
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yang L: Incidence and mortality of gastric
cancer in China. World J Gastroenterol. 12:17–20. 2006.PubMed/NCBI
|
|
3
|
Patel JN, Fuchs CS, Owzar K, Chen Z and
McLeod HL: Gastric cancer pharmacogenetics: Progress or old tripe?
Pharmacogenomics. 14:1053–1064. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Powolny AA and Singh SV: Multitargeted
prevention and therapy of cancer by diallyl trisulfide and related
Allium vegetable-derived organosulfur compounds. Cancer Lett.
269:305–314. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yi L and Su Q: Molecular mechanisms for
the anti-cancer effects of diallyl disulfide. Food Chem Toxicol.
57:362–370. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sundaram SG and Milner JA: Diallyl
disulfide suppresses the growth of human colon tumor cell
xenografts in athymic nude mice. J Nutr. 126:1355–1361.
1996.PubMed/NCBI
|
|
7
|
Nakagawa H, Tsuta K, Kiuchi K, Senzaki H,
Tanaka K, Hioki K and Tsubura A: Growth inhibitory effects of
diallyl disulfide on human breast cancer cell lines.
Carcinogenesis. 22:891–897. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shin DY, Kim GY, Kim JI, Yoon MK, Kwon TK,
Lee SJ, Choi YW, Kang HS, Yoo YH and Choi YH: Anti-invasive
activity of diallyl disulfide through tightening of tight junctions
and inhibition of matrix metalloproteinase activities in LNCaP
prostate cancer cells. Toxicol In Vitro. 24:1569–1576. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Park HS, Kim GY, Choi IW, Kim ND, Hwang
HJ, Choi YW and Choi YH: Inhibition of matrix metalloproteinase
activities and tightening of tight junctions by diallyl disulfide
in AGS human gastric carcinoma cells. J Food Sci. 76:T105–T111.
2011. View Article : Google Scholar
|
|
10
|
Ling H, Wen L, Ji XX, Tang YL, He J, Tan
H, Xia H, Zhou JG and Su Q: Growth inhibitory effect and
Chk1-dependent signaling involved in G2/M arrest on
human gastric cancer cells induced by diallyl disulfide. Braz J Med
Biol Res. 43:271–278. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bo S, Hui H, Li W, Hui L, Hong X, Lin D,
Dai WX, Wu YH, Ai XH, Hao J, et al: Chk1, but not Chk2, is
responsible for G2/M phase arrest induced by diallyl disulfide in
human gastric cancer BGC823 cells. Food Chem Toxicol. 68:61–70.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yuan JP, Wang GH, Ling H, Su Q, Yang YH,
Song Y, Tang RJ, Liu Y and Huang C: Diallyl disulfide-induced G2/M
arrest of human gastric cancer MGC803 cells involves activation of
p38 MAP kinase pathways. World J Gastroenterol. 10:2731–2734.
2004.PubMed/NCBI
|
|
13
|
Ling H, Zhang LY, Su Q, Song Y, Luo ZY,
Zhou XT, Zeng X, He J, Tan H and Yuan JP: Erk is involved in the
differentiation induced by diallyl disulfide in the human gastric
cancer cell line MGC803. Cell Mol Biol Lett. 11:408–423. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tang H, Kong Y, Guo J, Tang Y and Xie X,
Yang L, Su Q and Xie X: Diallyl disulfide suppresses proliferation
and induces apoptosis in human gastric cancer through Wnt-1
signaling pathway by up-regulation of miR-200b and miR-22. Cancer
Lett. 340:72–81. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gharahdaghi F, Weinberg CR, Meagher DA,
Imai BS and Mische SM: Mass spectrometric identification of
proteins from silver-stained polyacrylamide gel: A method for the
removal of silver ions to enhance sensitivity. Electrophoresis.
20:601–605. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Su B, Xiang B, Wang L, Cao L, Xiao L, Li
X, Li X, Wu M and Li G: Profiling and comparing transcription
factors activated in non-metastatic and metastatic nasopharyngeal
carcinoma cells. J Cell Biochem. 109:173–183. 2010.
|
|
17
|
Zhang LY, Ling H, Su Q, Song Y and Liang
XQ: Inhibitory effect of diallyl disulfide on human gastric cancer
cell line MGC803 in vitro. Shijie Huaren Xiaohua Zazhi.
11:1290–1293. 2003.In Chinese.
|
|
18
|
Zhu Y, McAvoy S, Kuhn R and Smith DI:
RORA, a large common fragile site gene, is involved in cellular
stress response. Oncogene. 25:2901–2908. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee JM, Kim IS, Kim H, Lee JS, Kim K, Yim
HY, Jeong J, Kim JH, Kim JY, Lee H, et al: RORalpha attenuates
Wnt/beta-catenin signaling by PKCalpha-dependent phosphorylation in
colon cancer. Mol Cell. 37:183–195. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xiong G, Wang C, Evers BM, Zhou BP and Xu
R: RORα suppresses breast tumor invasion by inducing SEMA3F
expression. Cancer Res. 72:1728–1739. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yamashita S, Tsujino Y, Moriguchi K,
Tatematsu M and Ushijima T: Chemical genomic screening for
methylation-silenced genes in gastric cancer cell lines using
5-aza-2′-deoxycytidine treatment and oligonucleotide microarray.
Cancer Sci. 97:64–71. 2006. View Article : Google Scholar
|
|
22
|
Su B, Xiang SL, Su J, Tang HL, Liao QJ,
Zhou YJ and Su Q: Diallyl disulfide increased histone acetylation
and p21WAF1 expression in human gastric cancer cells in
vivo and in vitro. Biochem Pharmacol. 1:1–10. 2012. View Article : Google Scholar
|
|
23
|
Marino N, Nakayama J, Collins JW and Steeg
PS: Insights into the biology and prevention of tumor metastasis
provided by the Nm23 metastasis suppressor gene. Cancer Metastasis
Rev. 31:593–603. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Boissan M, De Wever O, Lizarraga F, Wendum
D, Poincloux R, Chignard N, Desbois-Mouthon C, Dufour S,
Nawrocki-Raby B, Birembaut P, et al: Implication of metastasis
suppressor NM23-H1 in maintaining adherens junctions and limiting
the invasive potential of human cancer cells. Cancer Res.
70:7710–7722. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Che G, Chen J, Liu L, Wang Y, Li L, Qin Y
and Zhou Q: Transfection of nm23-H1 increased expression of
beta-catenin, E-cadherin and TIMP-1 and decreased the expression of
MMP-2, CD44v6 and VEGF and inhibited the metastatic potential of
human non-small cell lung cancer cell line L9981. Neoplasma.
53:530–537. 2006.PubMed/NCBI
|
|
26
|
Horak CE, Mendoza A, Vega-Valle E, Albaugh
M, Graff-Cherry C, McDermott WG, Hua E, Merino MJ, Steinberg SM,
Khanna C, et al: Nm23-H1 suppresses metastasis by inhibiting
expression of the lysophosphatidic acid receptor EDG2. Cancer Res.
67:11751–11759. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Huang CS, Liao JW and Hu ML: Lycopene
inhibits experimental metastasis of human hepatoma SK-Hep-1 cells
in athymic nude mice. J Nutr. 138:538–543. 2008.PubMed/NCBI
|
|
28
|
Guan-Zhen Y, Ying C, Can-Rong N, Guo-Dong
W, Jian-Xin Q and Jie-Jun W: Reduced protein expression of
metastasis-related genes (nm23, KISS1, KAI1 and p53) in lymph node
and liver metastases of gastric cancer. Int J Exp Pathol.
88:175–183. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bernard O: Lim kinases, regulators of
actin dynamics. Int J Biochem Cell Biol. 39:1071–1076. 2007.
View Article : Google Scholar
|
|
30
|
Wang W, Mouneimne G, Sidani M, Wyckoff J,
Chen X, Makris A, Goswami S, Bresnick AR and Condeelis JS: The
activity status of cofilin is directly related to invasion,
intravasation, and metastasis of mammary tumors. J Cell Biol.
173:395–404. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Scott RW, Hooper S, Crighton D, Li A,
König I, Munro J, Trivier E, Wickman G, Morin P, Croft DR, et al:
LIM kinases are required for invasive path generation by tumor and
tumor-associated stromal cells. J Cell Biol. 191:169–185. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhou Y, Su J, Shi L, Liao Q and Su Q: DADS
downregulates the Rac1-ROCK1/PAK1-LIMK1-ADF/cofilin signaling
pathway, inhibiting cell migration and invasion. Oncol Rep.
29:605–612. 2013.
|
|
33
|
Laufs S, Schumacher J and Allgayer H:
Urokinase-receptor (u-PAR): An essential player in multiple games
of cancer: A review on its role in tumor progression, invasion,
metastasis, proliferation/dormancy, clinical outcome and minimal
residual disease. Cell Cycle. 5:1760–1771. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Smith HW and Marshall CJ: Regulation of
cell signalling by uPAR. Nat Rev Mol Cell Biol. 11:23–36. 2010.
View Article : Google Scholar
|
|
35
|
Lester RD, Jo M, Montel V, Takimoto S and
Gonias SL: uPAR induces epithelial-mesenchymal transition in
hypoxic breast cancer cells. J Cell Biol. 178:425–436. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jo M, Lester RD, Montel V, Eastman B,
Takimoto S and Gonias SL: Reversibility of epithelial-mesenchymal
transition (EMT) induced in breast cancer cells by activation of
urokinase receptor-dependent cell signaling. J Biol Chem.
284:22825–22833. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ma YY and Tao HQ: Role of urokinase
plasminogen activator receptor in gastric cancer: A potential
therapeutic target. Cancer Biother Radiopharm. 27:285–290. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Belguise K, Kersual N, Galtier F and
Chalbos D: FRA-1 expression level regulates proliferation and
invasiveness of breast cancer cells. Oncogene. 24:1434–1444. 2005.
View Article : Google Scholar
|
|
39
|
Diesch J, Sanij E, Gilan O, Love C, Tran
H, Fleming NI, Ellul J, Amalia M, Haviv I, Pearson RB, et al:
Widespread FRA1-dependent control of mesenchymal
transdifferentiation programs in colorectal cancer cells. PLoS One.
9:e889502014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Huang XY, Ke AW, Shi GM, Zhang X, Zhang C,
Shi YH, Wang XY, Ding ZB, Xiao YS, Yan J, et al: αB-crystallin
complexes with 14–3–3ζ to induce epithelial-mesenchymal transition
and resistance to sorafenib in hepatocellular carcinoma.
Hepatology. 57:2235–2247. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Andreolas C, Kalogeropoulou M, Voulgari A
and Pintzas A: Fra-1 regulates vimentin during Ha-RAS-induced
epithelial mesenchymal transition in human colon carcinoma cells.
Int J Cancer. 122:1745–1756. 2008. View Article : Google Scholar
|
|
42
|
Andersen H, Mejlvang J, Mahmood S, Gromova
I, Gromov P, Lukanidin E, Kriajevska M, Mellon JK and Tulchinsky E:
Immediate and delayed effects of E-cadherin inhibition on gene
regulation and cell motility in human epidermoid carcinoma cells.
Mol Cell Biol. 25:9138–9150. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Mayes PA, Dolloff NG, Daniel CJ, Liu JJ,
Hart LS, Kuribayashi K, Allen JE, Jee DI, Dorsey JF, Liu YY, et al:
Overcoming hypoxia- induced apoptotic resistance through
combinatorial inhibition of GSK-3β and CDK1. Cancer Res.
71:5265–5275. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wu W, Ye H, Wan L, Han X, Wang G, Hu J,
Tang M, Duan X, Fan Y, He S, et al: Millepachine, a novel chalcone,
induces G2/M arrest by inhibiting CDK1 activity and
causing apoptosis via ROS-mitochondrial apoptotic pathway in human
hepatocarcinoma cells in vitro and in vivo. Carcinogenesis.
34:1636–1643. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Yi L, Ji XX, Tan H, Feng MY, Tang Y, Wen L
and Su Q: Involvement of Mcl1 in diallyl disulfide-induced G2/M
cell cycle arrest in HL-60 cells. Oncol Rep. 27:1911–1917.
2012.PubMed/NCBI
|
|
46
|
Yasui W, Ayhan A, Kitadai Y, Nishimura K,
Yokozaki H, Ito H and Tahara E: Increased expression of p34cdc2 and
its kinase activity in human gastric and colonic carcinomas. Int J
Cancer. 53:36–41. 1993. View Article : Google Scholar : PubMed/NCBI
|