Introduction
Lung cancer is one of the most common causes of
death from cancer worldwide. Non-small cell lung cancer (NSCLC),
which accounts for ~80% of lung cancers, is associated with a poor
5-year survival in developing countries (less than 9%) and in the
USA (not more than 15%) owing to its aggressiveness (1,2). Thus,
it is necessary to explore novel therapeutic approaches for
improving the quality of life of patients with lung cancer
including NSCLC.
Accumulating evidence has emerged that a small
population of cells possesses tumor-initiating activity in lung
cancer and these cells are termed ‘cancer stem-like cells (CSLCs)’
(3,4), which possess a variety of unique
biologic properties including self-renewal, expression of specific
biological markers and stem cell genes, and the use of common
signaling pathways and the stem cell niche (5–7). CSLCs
differ from normal stem cells in their tumorigenic capacities for
cancer initiation, recurrence, metastasis, and therapy resistance.
Although the existence of CSLCs in human lung cancer has been
previously studied, the regulation of self-renewal and stemness
maintenance in lung cancer stem-like cells (LCSLCs) in the
initiation of lung cancer remains unclear (8–10).
Gonadotropin-releasing hormone (GnRH) is synthesized
in a small subset of neurons in the septal-preoptic-hypothalamic
region (11,12). These neurons secrete the
neurohormone into the hypophysial portal circulation, through which
it reaches the anterior pituitary to stimulate the
synthesis/release of the two gonadotropins that, in turn, regulate
gonadal gender steroid production (13). In addition, GnRH and its receptor
were also found to be expressed in several types of cancer tissues,
either related (prostate, breast, ovarian and endometrial cancers)
or unrelated (melanoma, glioblastoma, lung and pancreatic cancers)
to the reproductive system, indicating that the expression of GnRH
may be linked to tumor progression. Recent studies indicate that
GnRH agonists have strong antitumor activity, such as
anti-proliferation, anti-metastasis and anti-angiogenesis (14–16).
These opposite biological effects of GnRH-R activation seem to be
linked to specific intracellular signaling cascades that are
coupled to this receptor in the different tissues (17,18).
Therefore, GnRH has been considered a promising candidate for novel
molecular-targeted strategies for the treatment of tumors.
In the present study, we found that GnRH is
associated with the regulation of self-renewal and stemness
maintenance in LCSLCs. Firstly, an in vitro sphere culture
system was established to isolate and enrich A549 LCSLCs. We then
determined the expression levels of tumorigenesis-related genes by
microarray analysis in the LCSLCs. Compared with the adherent
monolayer A549 cells, we found that the GnRH expression level was
much higher in the LCSLCs, suggesting that GnRH may play a role in
the regulation of self-renewal and stemness maintenance in LCSLCs.
Further examinations indicated that GnRH expression was essential
for the self-renewal and stemness maintenance by knockdown or
overexpression of GnRH expression in the A549 LCSLCs. Based on our
findings, GnRH and its receptor may represent an effective
molecular target for the development of innovative tumor-targeted
anticancer drugs directly or specifically for LCSLCs.
Materials and methods
Ethics statement
The present study was carried out in strict
accordance with the recommendations in the Guide for the Care and
Use of Laboratory Animals of Huazhong University of Science and
Technology. The animal procedures were approved by the Committee on
the Ethics of Animal Experiments of Huazhong University of Science
and Technology.
Cell culture and transfection
Human NSCLC cell line A549 was purchased from the
American Type Culture Collection (ATCC; Manassas, VA, USA), and
cultured in Dulbecco’s modified Eagle’s medium (DMEM) supplemented
with 10% fetal bovine serum (FBS) (both from Gibco, Grand Island,
NY, USA), 100 U/ml penicillin and streptomycin (both from
Invitrogen, Carlsbad, CA, USA) in a humidified 37°C incubator with
a 5% CO2 atmosphere. The cells stably overexpressing
GnRH were transfected with GnRH recombinant vector pCDNA3.1(+)-GnRH
by using Lipofectamine 2000™ and selected with 500 µg/ml
G418 (both from Invitrogen). After 72 h, the cells were analyzed by
quantitative real-time PCR (qRT-PCR) or western blot analysis.
Isolation of the A549 lung sphere
cells
The isolation of A549 sphere cells was modified
using a previously reported protocol (3,19). To
obtain sphere cultures, cells were plated at a density of
2×104 cells/well in 24-well ultra-low plates (Corning
Inc., Corning, NY, USA), supplemented with commercial hormone mix
B27 (Gibco), 20 ng/ml EGF (Sigma-Aldrich, St. Louis, MO, USA), 10
ng/ml bFGF (Upstate, Lake Placid, NY, USA), 0.4% bovine serum
albumin (BSA; Gibco), 4 µg/ml insulin (Sigma-Aldrich), 100
U/ml penicillin and streptomycin at 37°C. After being cultured for
7 days, the lung cancer spheres were collected, dissociated into a
single-cell suspension and resuspended in fresh medium for serial
subcultivation every 7 days. Adherent monolayer A549 cells were
used as the controls.
Colony-formation assay
For colony formation, the adherent monolayer A549
and the A549 sphere cells were dissociated into a single-cell
suspension. The cells were plated into 100-mm dishes at a density
of 2×103 cells/well in DMEM with 10% FBS. The plates
were further incubated for 2 weeks at 37°C with 5% CO2
until colonies were visible. The colonies were stained with 0.01%
crystal violet and counted using inverted microscopy. The dimension
of each colony was measured with Image J software (NIH, Bethesda,
MD, USA).
Microarray and data analysis
Human genome microarray analysis was conducted by
CapitalBio Corporation. Genes were determined to be differentially
expressed when logarithmic gene expression ratios were >5-fold
different and P-values <0.05. For data validation, the mRNA
levels and the expression of the genes of interest were confirmed
by qRT-PCR and western blot analysis.
qRT-PCR and western blotting
Total RNA was extracted from the tumor cells using
TRIzol reagent (Invitrogen) according to the manufacturer’s
protocol. qRT-PCR was performed using the SYBR PrimeScript RT-PCR
kit (Takara, Japan) on a Rotor-Gene 6000 real-time genetic analyzer
(Corbett Life Science, USA) according to the manufacturer’s
instructions. The GnRH forward primer was 5′atgaagccaattca
aaaact3′, and the reverse primer was 5′ttaaatcttcttctgccca3′. The
PCR protocol included a denaturation program (95°C for 2 min),
followed by 40 cycles of amplification and quantification program
(95°C for 5 sec, 55–57°C for 30 sec) and a melting curve program
(55–95°C, with a 0.5°C increment each cycle). Each sample was
replicated 3 times.
To analyze the protein expression levels, the cells
were washed by PBS 3 times and then lysed [1% Triton X-100 in PBS
buffer with protease inhibitor (Roche, Indianapolis, IN, USA)] for
30 min on ice. After incubation, the cell lysates were centrifuged
at 16,000 × g for 10 min. The supernatant was mixed with 5X SDS
sample buffer and boiled for 10 min, and electrophoresed at 80 V
through a 10% polyacrylamide-SDS denaturing gel. Separated proteins
were electrotransferred to nitrocellulose, and the expression of
GnRH, CD44, CD133, aldehyde dehydrogenase 1 (ALDH1), Sox2, JNK1,
c-Jun or GAPDH protein was detected with the desired antibodies
including mouse anti-GnRH monoclonal antibody (1:1,000; Santa Cruz
Biotechnology, Santa Cruz, CA, USA), mouse anti-CD44 monoclonal
antibody (1:1,000; Proteintech, Inc., Chicago, IL, USA), mouse
anti-CD133 monoclonal antibody (1:200; Miltenyi Biotec, Germany),
mouse anti-ALDH1 monoclonal antibody (1:1,000; Cell Signaling
Technology, Danvers, MA, USA), rabbit anti-Sox2 polyclonal antibody
(1:1,000) and rabbit anti-JNK1 polyclonal antibody (1:1,000) (both
from Abcam, Cambridge, UK), rabbit anti-phospho-c-Jun (Ser73)
polyclonal antibody (1:2,000; Cell Signaling Technology) and mouse
anti-GAPDH monoclonal antibody (1:3,000; GenScript Corp.,
Piscataway, NJ, USA), respectively. The signals were detected using
a SuperSignal West Femto Maximum Sensitivity Substrate (Thermo
Scientific, Waltham, MA, USA).
Immunofuorescence microscopy
The cells were seeded on a sterile coverslip and
washed with PBS 48 h later, and then fixed in 4% paraformaldehyde
(PFA) in PBS at room temperature for 30 min. The fixed cells were
washed with PBST (1X PBS containing 0.5% Triton X-100) and
incubated with 2% BSA in PBST (blocking buffer), and then incubated
with anti-GnRH antibody (1:200) in blocking buffer at 4°C
overnight. After 4 washes with PBST at room temperature for 15 min
each, the cells were incubated with FITC-conjugated goat anti-mouse
IgG (1:300; Jackson ImmunoResearch, West Grove, PA, USA). The cells
were then washed 4 times, followed by staining with propidium
iodide (PI; Sigma-Aldrich) to visualize the nuclear DNA, before
being mounted in mounting medium (Vector Laboratories, Burlingham,
CA, USA), and sealed under a coverslip with nail polish. The
prepared samples were observed and photographed under an Olympus
FluoView FV1000 confocal microscope.
Xenograft tumor growth assay
The desired numbers of tumor cells were injected
subcutaneously into the back of 4-week-old BALB/C-nude mice (n=5
each, Center of Experimental Animals, Huazhong University of
Science and Technology, China). The mice were reared for 2 months,
and the tumor growth and tumorigenic time were examined visually.
At the end of the experiment, the mice were sacrificed and tumor
xenografts were removed and measured. The tumor volume (TV) was
calculated in accordance with the formula: TV (mm3) =
d2 × D/2, where d and D represent the shortest and the
longest diameter, respectively.
Statistical analysis
All experiments were carried out at least 3 times
with triplicate samples. Data are presented as the mean ± SEM.
Statistical analysis was conducted using SPSS13.0 software. A
statistically significant difference was assessed by one-way ANOVA
followed by multiple mean comparisons by the Student’s t-test. A
value of P<0.05 was considered to be statistically
significant.
Results
LCSLCs derived from the A549 cell line
display self-renewal capacity
In our study, we established a stable sphere culture
system for isolating and enriching A549 sphere cells, which are
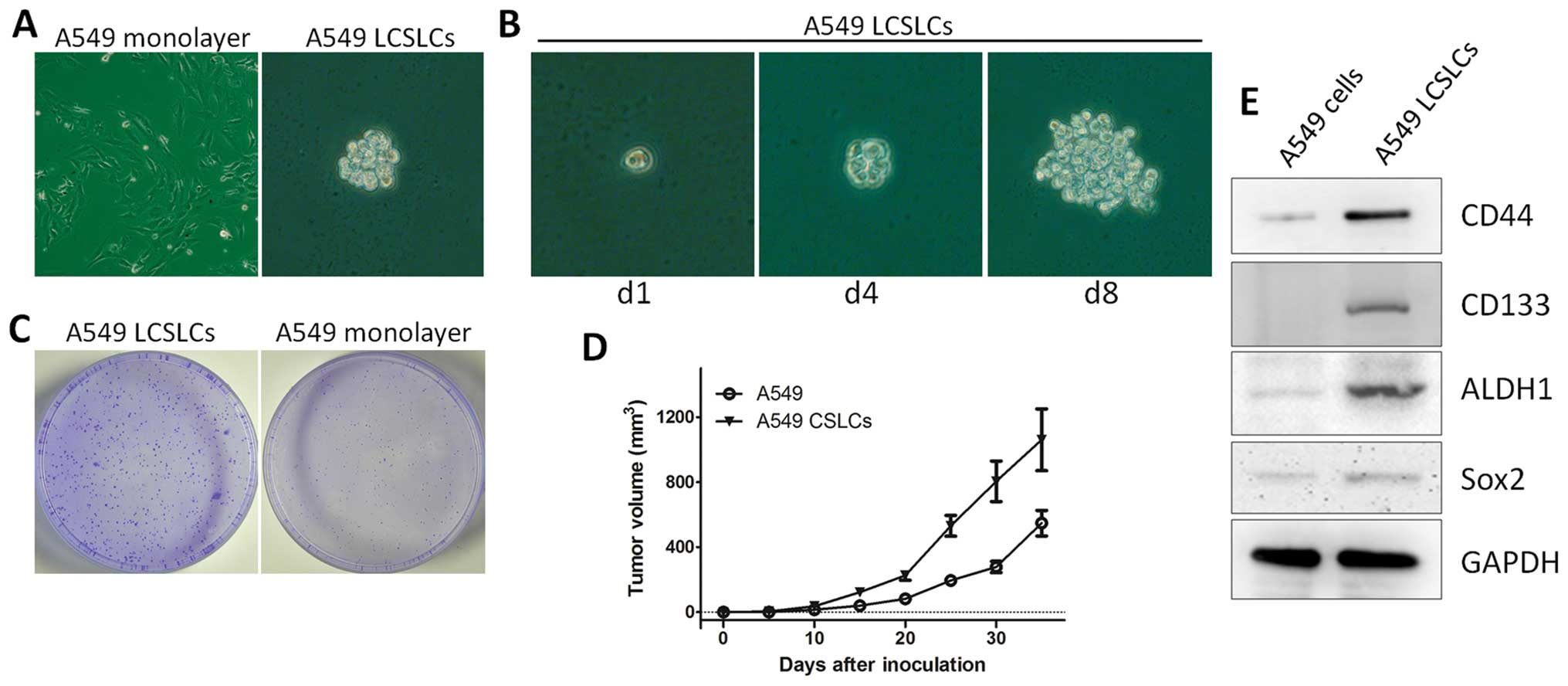
regarded as LCSLCs, to elucidate their biological behaviors. As
shown in Fig. 1, A549 cells that
were cultured in the SFM with EGF and bFGF generated non-adherent,
multicellular sphere LCSLCs (Fig.
1A). The result revealed that sphere LCSLCs exist in human lung
cancer A549 cells. Moreover, it was found that single cells of the
CSLCs from the lung cancer A549 cell line were able to form new
spheres (Fig. 1B), suggesting that
the A549 sphere has self-renewal capacity. We then assessed the
self-renewal capacity of A549 spheres by colony formation assay.
Compared with the A549 monolayer cells, the LCSLCs exhibited
enhanced colony formation (Fig.
1C).
The self-renewal capacity of LCSLCs in vivo
was revealed by examining the tumorigenic potential of A549
monolayer cells and A549 LCSLCs. We found that tumors derived from
the A549 monolayer cells yielded much smaller sized tumors than
those from the A549 LCSLCs (Fig.
1D). In addition, the xenograft tumor model in nude mice was
also used to investigate the tumorigenicity of the A549 monolayer
cells and A549 LCSLCs in BALB/C immunodeficient mice. The results
indicate that 1×104 A549 LCSLCs were enough to form
tumors in vivo, whereas 1×106 A549 monolayer
cells were required to induce stable tumor formation (Table I).
 | Table IXenotransplantation of human lung
cancer A549 cells and A549 LCSLCs into BALB/C immunodeficient
mice. |
Table I
Xenotransplantation of human lung
cancer A549 cells and A549 LCSLCs into BALB/C immunodeficient
mice.
| Cell type | Inoculum size | Tumor
incidence |
|---|
| A549 cells |
1×104 | 0/5 |
|
1×105 | 0/5 |
|
1×106 | 2/5 |
| A549 LCSLCs |
1×104 | 2/5 |
|
1×105 | 5/5 |
|
1×106 | 5/5 |
LCSLCs derived from the A549 cell line
overexpress stem cell markers
We next investigated the expression of cancer stem
cell (CSC) biomarkers or stemness-related transcription factors,
such as CD44, CD133, Sox2, ALDH1 and GAPDH in the A549 LCSLCs using
western blotting. The results demonstrated that, compared with the
A549 monolayer cells, CD44, CD133, Sox2 and ALDH1 protein
expression levels were apparently increased in the A549 LCSLCs
(Fig. 1E), suggesting that the
LCSLCs possess characteristics of CSCs.
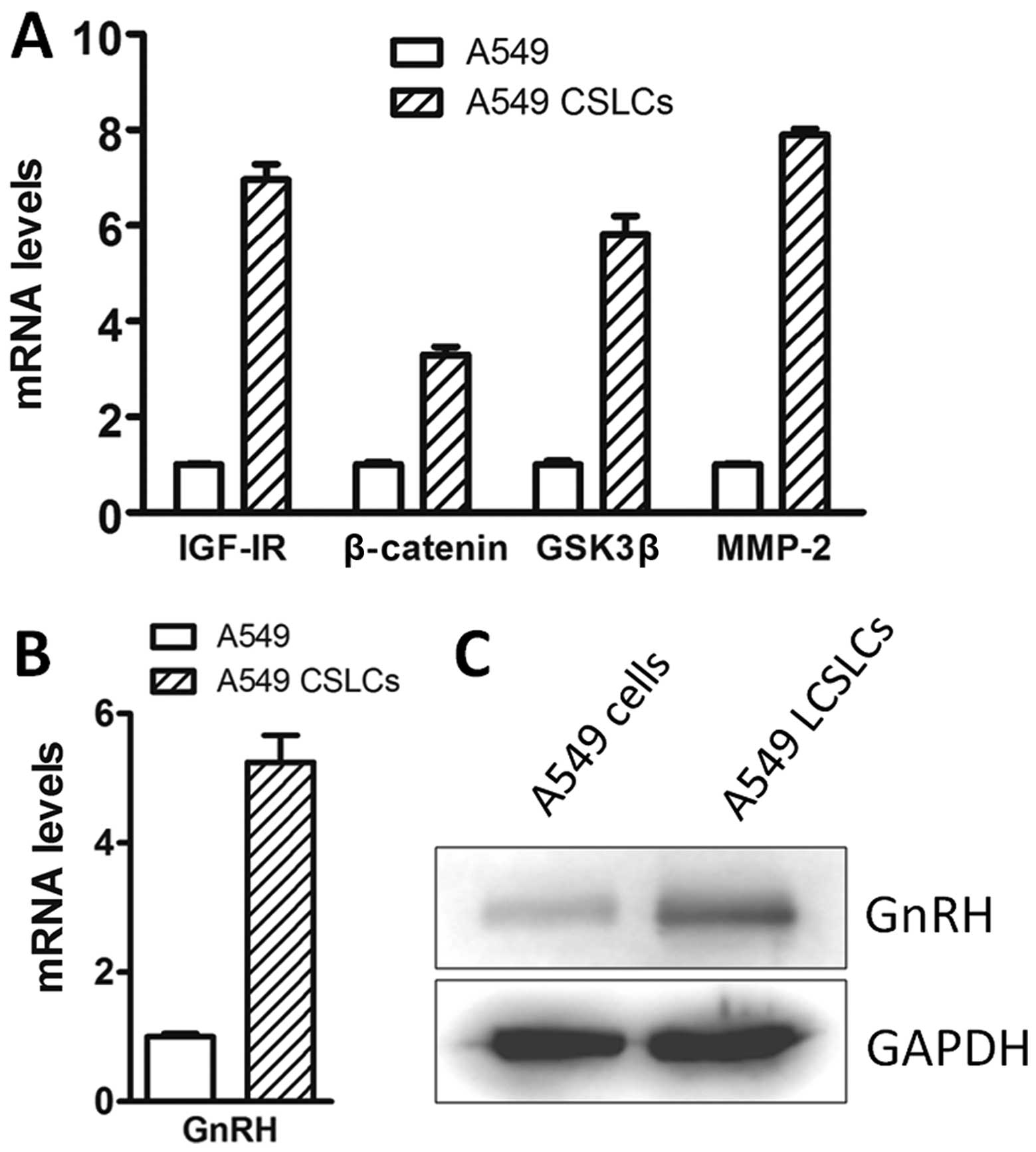
Upregulation of GnRH expression in the
A549 LCSLCs
After establishing the stable sphere culture system
for enriching and isolating LCSLCs from A549 cells, we performed
microarray analysis to further investigate the characteristics of
the LCSLCs. The categories of the microarray data (P<0.001) were
summarized on the basis of a gene ontology database (data not
shown). Genes that were significantly altered in the LCSLCs were
functionally categorized into signaling pathways in the KEGG
database. Top 10 signaling pathways were correlated with the
differentially expressed genes in the LCSLCs, such as IGF,
phosphatidylinositol-3-kinase (PI3K)/AKT/GSK3β and Wnt/β-catenin
pathways (Fig. 2A), which is
consistent with previous reports (10,19–21).
Intriguingly, the genes in the GnRH signaling pathway were
significantly altered in the LCSLCs, and qRT-PCR and western
blotting also confirmed that the mRNA and protein expression levels
of GnRH expression were significantly upregulated in the LCSLCs
(Fig. 2B and C), suggesting that
GnRH may play a role in self-renewal and stemness maintenance of
LCSLCs.
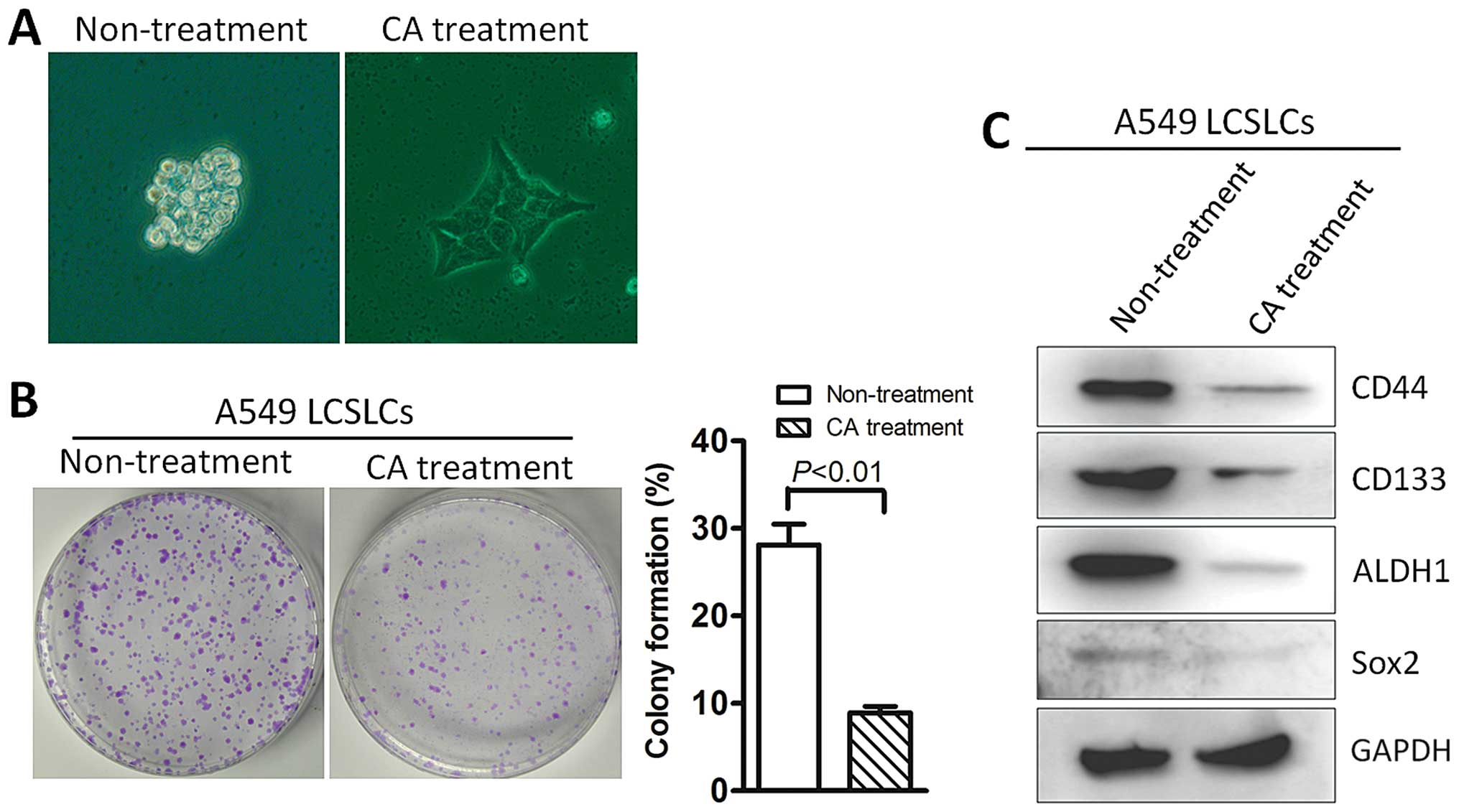
Suppression of GnRH capacity negatively
regulates the self-renewal and cell stemness of the lung cancer
cells
Since higher expression of GnRH was found in the
LCSLCs, we next characterized the stemness-associated properties of
the A549 cells and examined the biological functions of GnRH on
these properties. We then blocked the effects of GnRH by the
treatment of cetrorelix acetate, which is a man-made antagonist of
GnRH (Selleck, Houston, TX, USA). The cetrorelix acetate was
dissolved in sphere cell culture medium, and the concentration was
5 nM, which was modified according to a previous protocol (22). The sphere-forming assay was used to
identify stem cells retrospectively based on the utility of the
assay in evaluating self-renewal and differentiation at the
single-cell level in vitro. In our LCSLC culture system,
spheroid formation was observed on day 6, indicating that the A549
cells possessed a significant portion of LCSLCs. However, the
cetrorelix acetate treatment in the A549 cells significantly
inhibited spheroid formation (Fig.
3A). We also found that the treatment of cetrorelix acetate
inhibited the cell proliferation of the A549 LCSLCs as shown by the
colony-formation assay (Fig. 3B).
Western blotting demonstrated that the expression levels of CD44,
CD133, Sox2 and ALDH1 were significantly decreased in the
cetrorelix acetate-treated A549 cells in the sphere culture system,
compared with the non-treated A549 cells, suggesting that GnRH
expression was associated with the self-renewal capacity of the
LCSLCs in the A549 cells (Fig.
3C).
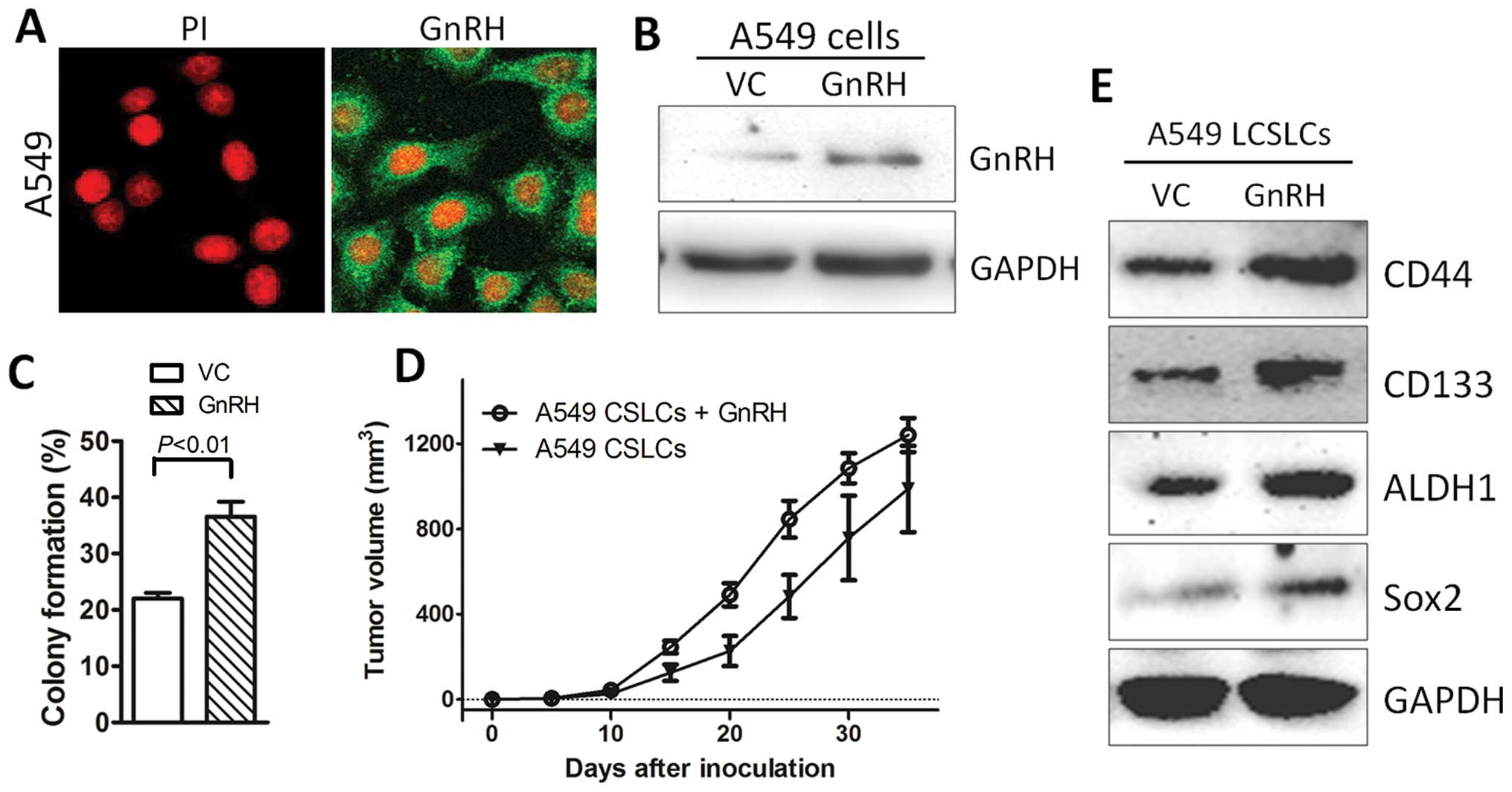
Excessive expression of GnRH induces
stemness properties in the A549 cells
We then analyzed the consequences of overexpression
of GnRH protein by stably transfecting pCDNA3.1(+)-GnRH vector into
the A549 cells. After examination using confocal microscopy, we
found that most of the A549 cells expressed GnRH protein in the
stable transfectants. In addition, at overexpression levels, GnRH
was primarily localized in the cytoplasm in the A549 cells
(Fig. 4A). Similarly, western
blotting further confirmed the overexpression of GnRH in the A549
cells, indicating the stable expression of GnRH protein in this
transfectant (Fig. 4B).
Furthermore, overexpression of GnRH increased the colony formation
in the LCSLCs (Fig. 4C), which was
significantly decreased by cetrorelix acetate treatment (Fig. 3B), indicating a higher frequency of
LCSLCs derived from the GnRH over-expressing spheres.
The role of GnRH in the self-renewal of LCSLCs in
vivo was also revealed by examining the tumorigenic potentials
of the GnRH overexpressing LCSLCs. We found that tumors derived
from the LCSLCs with GnRH overexpression had larger sizes than
those of the non-treated cells (Fig.
4D). Moreover, western blotting demonstrated higher expression
of stemness markers or stemness-related transcription factors in
the GnRH overexpressing LCSLCs (Fig.
4E), indicating that GnRH is a positive regulator of stemness
maintenance in LCSLCs.
Promotion of GnRH function for
self-renewal and stemness maintenance in the LCSLCs through the JNK
pathway
To further elucidate the molecular mechanism of GnRH
function in the self-renewal and stemness maintenance in LCSLCs, we
further investigated the regulation of the signaling pathways,
which may be related to stemness maintenance by GnRH expression in
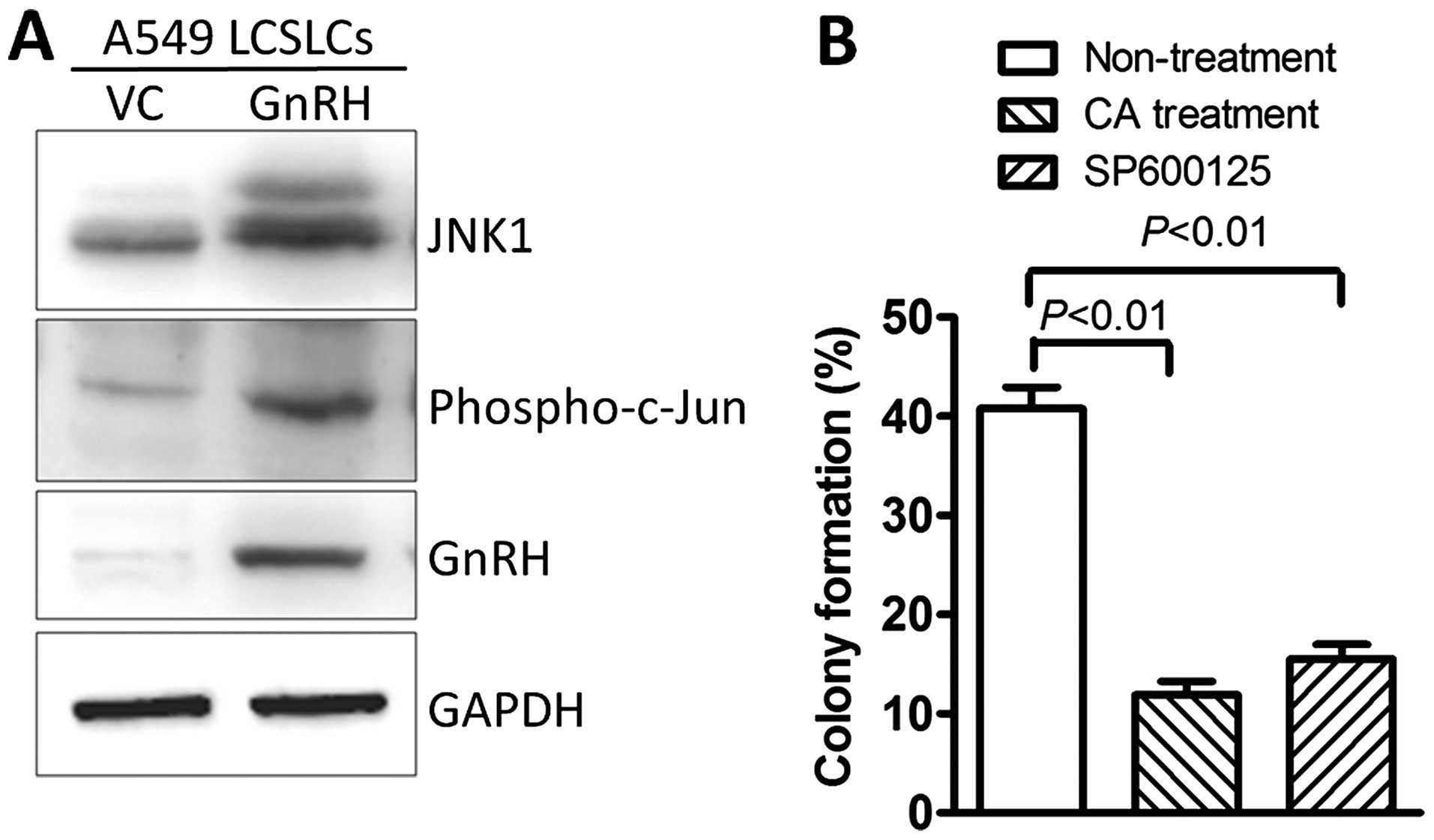
LCSLCs. After determining the expression of several genes, we found
that GnRH overexpression regulated the JNK pathway in LCSLCs
(Fig. 5A), which is consistent with
the previous reports in other tumor tissue (23,24).
Furthermore, we found that SP600125, a reversible JNK inhibitor,
can significantly inhibit the colony formation capacity of GnRH
overexpressing LCSLCs (Fig. 5B),
suggesting that GnRH may be associated with the capacity of
self-renewal and stemness maintenance through upregulation of the
JNK signaling pathway in LCSLCs.
Discussion
CSLCs are a subpopulation of cancer cells, which
possess unique survival capabilities and distinct stem cell
properties (25), play a critical
role in cancer initiation, progression, recurrence, and cancer
therapeutic failure (3,26,27).
The pathways that regulate self-renewal and cell fate in the CSC
system are beginning to be elucidated. To date, little is known
regarding the self-renewal and stemness maintenance, as well as the
regulatory mechanisms of CSCs. In the present study, we found that
GnRH was required for the self-renewal and stemness maintenance of
lung cancer cells. The self-renewal capacity can be regulated by
GnRH overexpression or inhibition in the LCSLCs. Moreover, we found
that this inhibition of GnRH function for self-renewal and stemness
maintenance was through the JNK signaling pathway.
In the last few years, it has been established that
GnRH is expressed in a variety of human tumor tissues, such as
ovary, endometrium, placenta, breast, prostate, and lung tumors
(28,29). For example, in both the endometrium
and the placenta, activation of locally expressed GnRH increases
the activity of the urokinase type plasminogen activator and matrix
metalloproteinase (MMP) system, suggesting the GnRH may reduce the
migratory and the invasive behaviors of cancer cells by regulating
MMP-2 expression/activity (30,31).
In addition, GnRH angonists have been shown to exert strong
anti-proliferative effects on human prostate, breast and ovarian
cancer cells (32–34). Moreover, GnRH may exert antitumor
activity not only by reducing cancer cell proliferation and
metastasis, but also by affecting the process of angiogenesis. It
has been found that GnRH analogs significantly decrease the
synthesis and secretion of VEGF in melanoma cells (35). Based on these observations, tumor
GnRH and its receptor are now considered a very promising candidate
for novel molecular-targeted strategies for the treatment of
tumors. However, limited data exist on the significance of GnRH
expression in CSCs, and the association between GnRH and stemness
maintenance in CSCs.
To investigate the functions of GnRH in CSCs, we
firstly isolated the sphere subpopulation from human NCLSC A549
cells using a sphere-forming enrichment culture method with
selective serum-free medium. The results demonstrated that the
sphere cells could proliferate and form a new sphere, confirming
that the sphere cell subpopulation possessed self-renewal capacity
(Fig. 1). We then discovered that
the expression levels of CSC biological markers, such as CD44,
CD133 and ALDH1, and stemness-related transcription factors Sox2 in
the LCSLCs, were much higher than that in the monolayer A549 cells.
As known, CD44, CD133, ALDH1, and Sox2 are important for the
sorting and identification of various types of CSCs, especially in
lung cancer.
Although the functions of GnRH in various tumors
have been well studied in recent years, the data available on the
presence of GnRH in lung cancer are still limited (36). After microarray assay analysis, we
found many alterations in gene expression levels in the A549
monolayer cells and LCSLCs. The higher expression level of GnRH in
LCSLCs encouraged us to investigate the functions of GnRH in the
regulation of stemness maintenance in LCSLCs (Fig. 2). Notably, both overexpression and
inactivation of GnRH demonstrated that it is essential for the
self-renewal and stemness maintenance in vivo and in
vitro (Figs. 3 and 4). Regardless of the mechanisms, our
results suggest that GnRH capacity regulates the stemness
maintenance of LCSCs.
GnRH has been known to regulate different
intracellular signaling cascades, such as MAPK (p38/MAPK, ERK1/2,
and JNK), PI3K, and phosphotyrosine phosphatase (PTP) (23,37–39).
In ovarian and endometrial cancer cells, the GnRH agonist
triptorelin has been reported to increase JNK activity, leading to
c-Jun binding to DNA and, ultimately, to an anti-proliferative
effect (17,40). Moreover, JNK was shown to mediate
the GnRH-stimulated invasive behavior of ovarian cancer cells by
increasing the expression of MMP-2 and MMP-9 (24). Therefore, we hypothesized that the
GnRH function of self-renewal and stemness maintenance in LCSLCs
was through the JNK signaling pathway. Song et al provided
evidence that the JNK/c-Jun signaling pathway mediates
EPHA2-dependent tumor cell proliferation, motility, and cancer stem
cell-like properties in NSCLCs (41). A recent study also demonstrated that
JNK signaling is crucial for the maintenance of self-renewal and
tumorigenicity of glioma stem-like cells and drug/IR resistance,
and can be considered a promising target for eliminating stem-like
cancer cells in glioma (42).
Furthermore, in ovarian CSCs, it was found that JNK plays an
essential role in the maintenance of the self-renewal and
tumor-initiating capacity (43). As
expected, our findings demonstrated similar results, that GnRH
overexpression can induce the activation of the JNK signaling
pathway in LCSLCs (Fig. 5),
suggesting that the GnRH-mediated activation of the JNK pathway may
be essential for self-renewal and stemness maintenance in
LCSLCs.
Although our studies suggest that JNK signaling
mediates GnRH-dependent stemness maintenance in LCSLCs, it cannot
be excluded that other GnRH-regulated signaling pathways may also
play a role in this process. It still remains largely unknown
whether the precise role of GnRH in the self-renewal and stemness
maintenance in LCSLCs is through regulation of the JNK pathway or
through other signaling pathways. Therefore, the precise mechanisms
and pathways, by which GnRH participates in the upregulation of the
JNK pathway for self-renewal and stemness maintenance, require
further investigation.
Acknowledgments
The present study was supported by grant WX13B02
from the Foundation of Health and the Family Planning Commission of
Wuhan Municipality, China.
References
|
1
|
Esposito L, Conti D, Ailavajhala R, Khalil
N and Giordano A: Lung cancer: are we up to the challenge? Curr
Genomics. 11:513–518. 2010. View Article : Google Scholar
|
|
2
|
Jemal A, Siegel R, Ward E, Murray T, Xu J,
Smigal C and Thun MJ: Cancer statistics, 2006. CA Cancer J Clin.
56:106–130. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Eramo A, Lotti F, Sette G, Pilozzi E,
Biffoni M, Di Virgilio A, Conticello C, Ruco L, Peschle C and De
Maria R: Identification and expansion of the tumorigenic lung
cancer stem cell population. Cell Death Differ. 15:504–514. 2008.
View Article : Google Scholar
|
|
4
|
Hermann PC, Bhaskar S, Cioffi M and
Heeschen C: Cancer stem cells in solid tumors. Semin Cancer Biol.
20:77–84. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Clevers H: The cancer stem cell: premises,
promises and challenges. Nat Med. 17:313–319. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sneddon JB and Werb Z: Location, location,
location: the cancer stem cell niche. Cell Stem Cell. 1:607–611.
2007. View Article : Google Scholar
|
|
7
|
Zhang WC, Shyh-Chang N, Yang H, Rai A,
Umashankar S, Ma S, Soh BS, Sun LL, Tai BC, Nga ME, et al: Glycine
decarboxylase activity drives non-small cell lung cancer
tumor-initiating cells and tumorigenesis. Cell. 148:259–272. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen YC, Hsu HS, Chen YW, Tsai TH, How CK,
Wang CY, Hung SC, Chang YL, Tsai ML, Lee YY, et al: Oct-4
expression maintained cancer stem-like properties in lung
cancer-derived CD133-positive cells. PLoS One. 3:e26372008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chiou SH, Wang ML, Chou YT, Chen CJ, Hong
CF, Hsieh WJ, Chang HT, Chen YS, Lin TW, Hsu HS, et al:
Coexpression of Oct4 and Nanog enhances malignancy in lung
adenocarcinoma by inducing cancer stem cell-like properties and
epithelial-mesenchymal transdifferentiation. Cancer Res.
70:10433–10444. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xu C, Xie D, Yu SC, Yang XJ, He LR, Yang
J, Ping YF, Wang B, Yang L, Xu SL, et al: β-catenin/POU5F1/SOX2
transcription factor complex mediates IGF-I receptor signaling and
predicts poor prognosis in lung adenocarcinoma. Cancer Res.
73:3181–3189. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Anthony EL, King JC and Stopa EG:
Immunocytochemical localization of LHRH in the median eminence,
infundibular stalk, and neurohypophysis. Evidence for multiple
sites of releasing hormone secretion in humans and other mammals.
Cell Tissue Res. 236:5–14. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Stopa EG, Koh ET, Svendsen CN, Rogers WT,
Schwaber JS and King JC: Computer-assisted mapping of
immunoreactive mammalian gonadotropin-releasing hormone in adult
human basal forebrain and amygdala. Endocrinology. 128:3199–3207.
1991. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Conn PM and Crowley WF Jr:
Gonadotropin-releasing hormone and its analogs. Annu Rev Med.
45:391–405. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Montagnani Marelli M, Moretti RM,
Januszkiewicz-Caulier J, Motta M and Limonta P:
Gonadotropin-releasing hormone (GnRH) receptors in tumors: a new
rationale for the therapeutical application of GnRH analogs in
cancer patients? Curr Cancer Drug Targets. 6:257–269. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cheung LW and Wong AS:
Gonadotropin-releasing hormone: GnRH receptor signaling in
extrapituitary tissues. FEBS J. 275:5479–5495. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
So WK, Cheng JC, Poon SL and Leung PC:
Gonadotropin-releasing hormone and ovarian cancer: a functional and
mechanistic overview. FEBS J. 275:5496–5511. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gründker C, Schlotawa L, Viereck V and
Emons G: Protein kinase C-independent stimulation of activator
protein-1 and c-Jun N-terminal kinase activity in human endometrial
cancer cells by the LHRH agonist triptorelin. Eur J Endocrinol.
145:651–658. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Limonta P, Moretti RM, Marelli MM, Dondi
D, Parenti M and Motta M: The luteinizing hormone-releasing hormone
receptor in human prostate cancer cells: messenger ribonucleic acid
expression, molecular size, and signal transduction pathway.
Endocrinology. 140:5250–5256. 1999.PubMed/NCBI
|
|
19
|
Kim IG, Kim SY, Choi SI, Lee JH, Kim KC
and Cho EW: Fibulin-3-mediated inhibition of
epithelial-to-mesenchymal transition and self-renewal of ALDH lung
cancer stem cells through IGF1R signaling. Oncogene. 1:1–10.
2013.
|
|
20
|
Mo XM, Li HH, Liu M and Li YT:
Downregulation of GSK3β by miR-544a to maintain self-renewal
ability of lung cancer stem cells. Oncol Lett. 8:1731–1734.
2014.PubMed/NCBI
|
|
21
|
Liu F, Cao X, Liu Z, Guo H, Ren K, Quan M,
Zhou Y, Xiang H and Cao J: Casticin suppresses self-renewal and
invasion of lung cancer stem-like cells from A549 cells through
down-regulation of pAkt. Acta Biochim Biophys Sin (Shanghai).
46:15–21. 2014. View Article : Google Scholar
|
|
22
|
Metallinou C, Köster F, Diedrich K,
Nikolettos N and Asimakopoulos B: The effects of cetrorelix and
triptorelin on the viability and steroidogenesis of cultured human
granulosa luteinized cells. In Vivo. 26:835–839. 2012.PubMed/NCBI
|
|
23
|
Kraus S, Levy G, Hanoch T, Naor Z and
Seger R: Gonadotropin-releasing hormone induces apoptosis of
prostate cancer cells: role of c-Jun NH2-terminal kinase, protein
kinase B, and extracellular signal-regulated kinase pathways.
Cancer Res. 64:5736–5744. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cheung LW, Leung PC and Wong AS:
Gonadotropin-releasing hormone promotes ovarian cancer cell
invasiveness through c-Jun NH2-terminal kinase-mediated activation
of matrix metalloproteinase (MMP)-2 and MMP-9. Cancer Res.
66:10902–10910. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Huntly BJ and Gilliland DG: Cancer
biology: summing up cancer stem cells. Nature. 435:1169–1170. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Clarke MF, Dick JE, Dirks PB, Eaves CJ,
Jamieson CH, Jones DL, Visvader J, Weissman IL and Wahl GM: Cancer
stem cells - perspectives on current status and future directions:
AACR workshop on cancer stem cells. Cancer Res. 66:9339–9344. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sueblinvong V and Weiss DJ: Stem cells and
cell therapy approaches in lung biology and diseases. Transl Res.
156:188–205. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Limonta P, Moretti RM, Montagnani Marelli
M and Motta M: The biology of gonadotropin hormone-releasing
hormone: role in the control of tumor growth and progression in
humans. Front Neuroendocrinol. 24:279–295. 2003. View Article : Google Scholar
|
|
29
|
Harrison GS, Wierman ME, Nett TM and Glode
LM: Gonadotropin-releasing hormone and its receptor in normal and
malignant cells. Endocr Relat Cancer. 11:725–748. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chou CS, Tai CJ, MacCalman CD and Leung
PC: Dose-dependent effects of gonadotropin releasing hormone on
matrix metalloproteinase (MMP)-2, and MMP-9 and tissue specifc
inhibitor of metalloproteinases-1 messenger ribonucleic acid levels
in human decidual stromal cells in vitro. J Clin Endocrinol Metab.
88:680–688. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Moretti RM, Montagnani Marelli M, Mai S
and Limonta P: Gonadotropin-releasing hormone agonists suppress
melanoma cell motility and invasiveness through the inhibition of
α3 integrin and MMP-2 expression and activity. Int J Oncol.
33:405–413. 2008.PubMed/NCBI
|
|
32
|
Dondi D, Limonta P, Moretti RM, Marelli
MM, Garattini E and Motta M: Antiproliferative effects of
luteinizing hormone-releasing hormone (LHRH) agonists on human
androgen-independent prostate cancer cell line DU 145: evidence for
an autocrine-inhibitory LHRH loop. Cancer Res. 54:4091–4095.
1994.PubMed/NCBI
|
|
33
|
Kéri G, Balogh A, Szöke B, Teplán I and
Csuka O: Gonadotropin-releasing hormone analogues inhibit cell
proliferation and activate signal transduction pathways in
MDA-MB-231 human breast cancer cell line. Tumour Biol. 12:61–67.
1991. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gründker C, Günthert AR, Westphalen S and
Emons G: Biology of the gonadotropin-releasing hormone system in
gynecological cancers. Eur J Endocrinol. 146:1–14. 2002. View Article : Google Scholar
|
|
35
|
Moretti RM, Mai S, Montagnani Marelli M,
Bani MR, Ghilardi C, Giavazzi R, Taylor DM, Martini PG and Limonta
P: Dual targeting of tumor and endothelial cells by
gonadotropin-releasing hormone agonists to reduce melanoma
angiogenesis. Endocrinology. 151:4643–4653. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Taratula O, Garbuzenko OB, Chen AM and
Minko T: Innovative strategy for treatment of lung cancer: targeted
nanotechnology-based inhalation co-delivery of anticancer drugs and
siRNA. J Drug Target. 19:900–914. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Maudsley S, Davidson L, Pawson AJ, Chan R,
López de Maturana R and Millar RP: Gonadotropin-releasing hormone
(GnRH) antagonists promote proapoptotic signaling in peripheral
reproductive tumor cells by activating a Galphai-coupling state of
the type I GnRH receptor. Cancer Res. 64:7533–7544. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Malik SN, Brattain M, Ghosh PM, Troyer DA,
Prihoda T, Bedolla R and Kreisberg JI: Immunohistochemical
demonstration of phospho-Akt in high Gleason grade prostate cancer.
Clin Cancer Res. 8:1168–1171. 2002.PubMed/NCBI
|
|
39
|
Kraus S, Naor Z and Seger R:
Gonadotropin-releasing hormone in apoptosis of prostate cancer
cells. Cancer Lett. 234:109–123. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Günthert AR, Gründker C, Hollmann K and
Emons G: Luteinizing hormone-releasing hormone induces JunD-DNA
binding and extends cell cycle in human ovarian cancer cells.
Biochem Biophys Res Commun. 294:11–15. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Song W, Ma Y, Wang J, Brantley-Sieders D
and Chen J: JNK signaling mediates EPHA2-dependent tumor cell
proliferation, motility, and cancer stem cell-like properties in
non-small cell lung cancer. Cancer Res. 74:2444–2454. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yoon CH, Kim MJ, Kim RK, Lim EJ, Choi KS,
An S, Hwang SG, Kang SG, Suh Y, Park MJ, et al: c-Jun N-terminal
kinase has a pivotal role in the maintenance of self-renewal and
tumorigenicity in glioma stem-like cells. Oncogene. 31:4655–4666.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Seino M, Okada M, Shibuya K, Seino S,
Suzuki S, Ohta T, Kurachi H and Kitanaka C: Requirement of JNK
signaling for self-renewal and tumor-initiating capacity of ovarian
cancer stem cells. Anticancer Res. 34:4723–4731. 2014.PubMed/NCBI
|