Introduction
Angiogenesis, the formation of new blood vessels
from pre-existing neighboring vessels, is essential for
pathological conditions including cancer, ocular disorders,
arthritis and obesity as well as physiological processes such as
wound-healing, menstruation and ovulation (1,2). The
process of angiogenesis includes endothelial cell proliferation,
migration, adhesion, invasion, tube formation and recruitment of
pericytes, and is tightly regulated by the complex interplay of
angiogenic and anti-angiogenic factors within tissue
microenvironment (3–5). Matrix metalloproteinases (MMPs) play
important roles in tissue remodeling by degrading extracellular
matrix (ECM) components and cell surface molecules, leading to
angiogenic responses associated with cancer growth and progression
(6–8). MMP-mediated cleavage of vascular
endothelial (VE)-cadherin in cell surfaces may promote vascular
permeability, proliferation, invasion and capillary-like structure
formation by dissociating cadherin-catenin complex and disrupting
cell-cell adhesion (9–14). The activities of these MMPs are
regulated by endogenous inhibitors, tissue inhibitors of
metalloproteinases (TIMPs) (15).
In addition to MMP-inhibitory activity, many investigations
demonstrate that TIMPs regulate cell fates such as proliferation,
migration, apoptosis and differentiation through MMP-independent
mechanism (16–21). Molecular mechanisms underlying
regulation of expression and activities of MMPs and TIMPs may be
attractive therapeutic targets and strategy for intervention in
angiogenesis-related disorders including cancer. However, MMP
inhibitors in multiple clinical trials show severe musculoskeletal
pain and inflammation as well as limited efficacy, suggesting that
the identification of the specific MMP target is absolutely
required for realizing the clinical potential of MMP inhibitors
(22,23).
Ligularia fischeri (LF) (Ledebour)
Turczaninow var. spiciformis Nakai (Compositae), an edible
herb distributed in Eastern Asia including Korea, China and Japan,
has been used in traditional medicine to treat rheumatoid
arthritis, erysipelas, scarlet fever and jaundice. Previous
investigations demonstrate that the extracts and biologically
active components of LF exert anti-inflammatory, anti-oxidative,
anti-hepatotoxic and anti-obesity activities (24–29).
In addition, several studies show the anticancer activity of LF
against various cancer cell lines including acute promyelocytic
leukemia, oral cancer, breast cancer, lung cancer and ovarian
cancer cells (30–33). However, the biological effects of LF
on angiogenesis associated with cancer growth and progression have
not yet been explored. In the present study, we evaluated the
regulatory effects and molecular mechanisms of LF on cell
proliferation, invasion and capillary-like structure formation in
human umbilical vein endothelial cells (HUVECs).
Materials and methods
Cell culture conditions
Primary cultures of HUVECs were purchased from Lonza
(Walkersville, MD, USA) and used between passages 3 and 6 for all
experiments. Cells were cultured in EGM-2® BulletKit
containing endothelial basal medium-2 (EBM-2) and the following
growth supplements (EGM-2® SingleQuots Kit: human
epidermal growth factor, vascular endothelial growth factor,
R3-insulin-like growth factor-1, human fibroblast growth factor,
ascorbic acid, hydrocortisone, heparin, fetal bovine serum and
gentamicin/amphotericin B) (designated as complete media),
according to the manufacturer’s instructions (Lonza).
Reagents
The following pharmacological agents and antibodies
were purchased from commercial sources: anti-phospho-extra-cellular
signal-regulated kinase (ERK) (T202/Y204), anti-phospho-Akt (S473),
anti-phospho-p70S6K (T421/S424),
anti-phospho-p38MAPK (T180/Y182), anti-MMP-2,
anti-MMP-9, anti-phospho-pRb (S780) and anti-phospho-pRb
(S807/S811) (all from Cell Signaling Technology, Beverly, MA, USA);
anti-p27Kip1 (BD Biosciences, Bedford, MA, USA);
anti-ERK, anti-Akt, anti-p70S6K,
anti-p38MAPK, anti-cyclin-dependent kinase (Cdk)4,
anti-Cdk2, anti-cyclin D, anti-cyclin E, anti-actin antibodies and
mouse and rabbit IgG-horseradish peroxidase conjugates (Santa Cruz
Biotechnology, Santa Cruz, CA, USA).
Preparation of LF extract
Five hundred grams of LF were extracted with 2
liters of ethanol and stirring for 5 h. The extract of LF was
obtained as previously reported (32).
Cell viability and proliferation
assay
Subconfluent HUVECs, plated on 6-well plates
(1×105 cells/well; BD Biosciences), were serum-starved
for 14 h in EBM-2 media to synchronize cells in
G1/G0 phase of cell cycle and incubated for
24 h in EGM-2 BulletKit media in the presence or absence of LF (1
and 10 µg/ml). Following culture for 24 h, cell viability
was determined by a Muse™ Cell Analyzer using Cell Count and
Viability Assay kit (Merck Millipore, Billerica, MA, USA), and the
cell proliferation was quantified as previously described (34). The results from triplicate
determinations (mean ± standard deviation) are presented as the
fold-increase of the untreated controls or the percentage of viable
cells of the total cell count.
Cell cycle analysis
Quiescent HUVECs were incubated for 24 h in EGM-2
BulletKit media in the presence or absence of LF (10 µg/ml).
Cells were harvested with trypsin-EDTA, rinsed with
phosphate-buffered saline (PBS, pH 7.4) and then fixed with
ice-cold 70% ethanol for 3 h. After washing with PBS, cells were
stained with Muse™ cell cycle reagent. The profile of cells in the
G1/G0, S and G2/M phases of the
cell cycle was analyzed with a Muse™ Cell Analyzer (Merck
Millipore) (35).
Western blot analysis
Quiescent HUVECs in 100-mm dishes (1×106
cells/dish; BD Biosciences) were incubated for 15 min or 24 h in
EGM-2 Bulletkit media in the presence or absence of LF (10
µg/ml). Cells were rinsed twice with ice-cold PBS and lysed
by incubation in 50 mM Tris-HCl (pH 7.4), 150 mM NaCl, 10%
glycerol, 1% Triton X-100, 1 mM EDTA, 100 µg/ml
4-(2-aminoethyl)benzenesulfonyl fluoride, 10 µg/ml
aprotinin, 1 µg/ml pepstatin A, 0.5 µg/ml leupeptin,
80 mM β-glycerophosphate, 25 mM sodium fluoride and 1 mM sodium
orthovanadate for 30 min at 4°C. Cell lysates were clarified at
12,500 × g for 20 min at 4°C and the supernatants were subjected to
western blot analysis as described previously (36–38).
All western blot analyses are representative of at least three
independent experiments. Bands of interest were integrated and
quantified by the use of National Institutes of Health (NIH) ImageJ
version 1.34s software.
Invasion assay
The upper side of the Transwell insert (Costar,
6.5-mm diameter insert, 8-µm pore size) (Corning Inc.,
Corning, NY, USA) was coated with 50 µl of 1 mg/ml
Matrigel® basement membrane matrix (10.4 mg/ml; BD
Biosciences) diluted in EBM-2. Aliquots (100 µl) of HUVECs
(5×104 cells/ml) resuspended in EBM-2 were added to the
upper compartment of the Matrigel-coated Transwell and 600
µl of EBM-2 was added to the lower compartment. After serum
starvation with EBM-2 for 2 h, cells were incubated for 18 h in
EGM-2 BulletKit media in the presence or absence of LF (1 and 10
µg/ml). The inserts were fixed with methanol and using a
cotton-tipped swab the non-invasive cells were removed from the top
of the membrane. After staining with 0.04% Giemsa solution
(Sigma-Aldrich, St. Louis, MO, USA), the number of invasive cells
was determined from six different fields using ×200 objective
magnification.
Tube formation assay
Each well of pre-chilled 24-well plates was coated
with 200 µl Matrigel (BD Biosciences). Following serum
starvation with EBM-2 for 2 h, HUVECs (3×104 cells/ml)
were added to Matrigel®-coated plates and incubated for
6 h in EGM-2 BulletKit media in the presence or absence of LF (1
and 10 µg/ml). Tube formation was observed with an Olympus
CKX41 inverted microscope (CAchN 10/0.25php objective) and ToupTek
Toupview software (version ×86, 3.5.563; Hangzhou ToupTek Photonics
Co., Zhejiang, China) (39).
Immunofluorescence microscopy
Quiescent HUVECs on gelatin-coated coverslips in
12-well plates were incubated for 30 min in EGM-2 BulletKit media
in the presence or absence of LF (10 µg/ml), fixed with 3.7%
paraformaldehyde for 5 min, washed with PBS, permeabilized with
0.1% Triton X-100 for 10 min, washed with PBS and blocked with PBS
containing 5% BSA for 1 h. Primary antibodies diluted 1:100 in 5%
BSA-PBS were incubated for 2 h at room temperature, washed with
PBS, and followed by Alexa Fluor 488-conjugated goat anti-mouse IgG
(Life Technologies, Grand Island, NY, USA). Images were obtained
with Carl Zeiss microscope (Axio Imager. M2) and AxioVision Rel.
4.8 software (Zeiss Co., Gottingen, Germany).
Statistical analysis
Statistical analysis was performed using the
Student’s t-test and was based on at least three different
experiments. The results were considered to be statistically
significant at P<0.05.
Results
LF suppresses endothelial cell
proliferation through regulating the expression of cell
cycle-related proteins
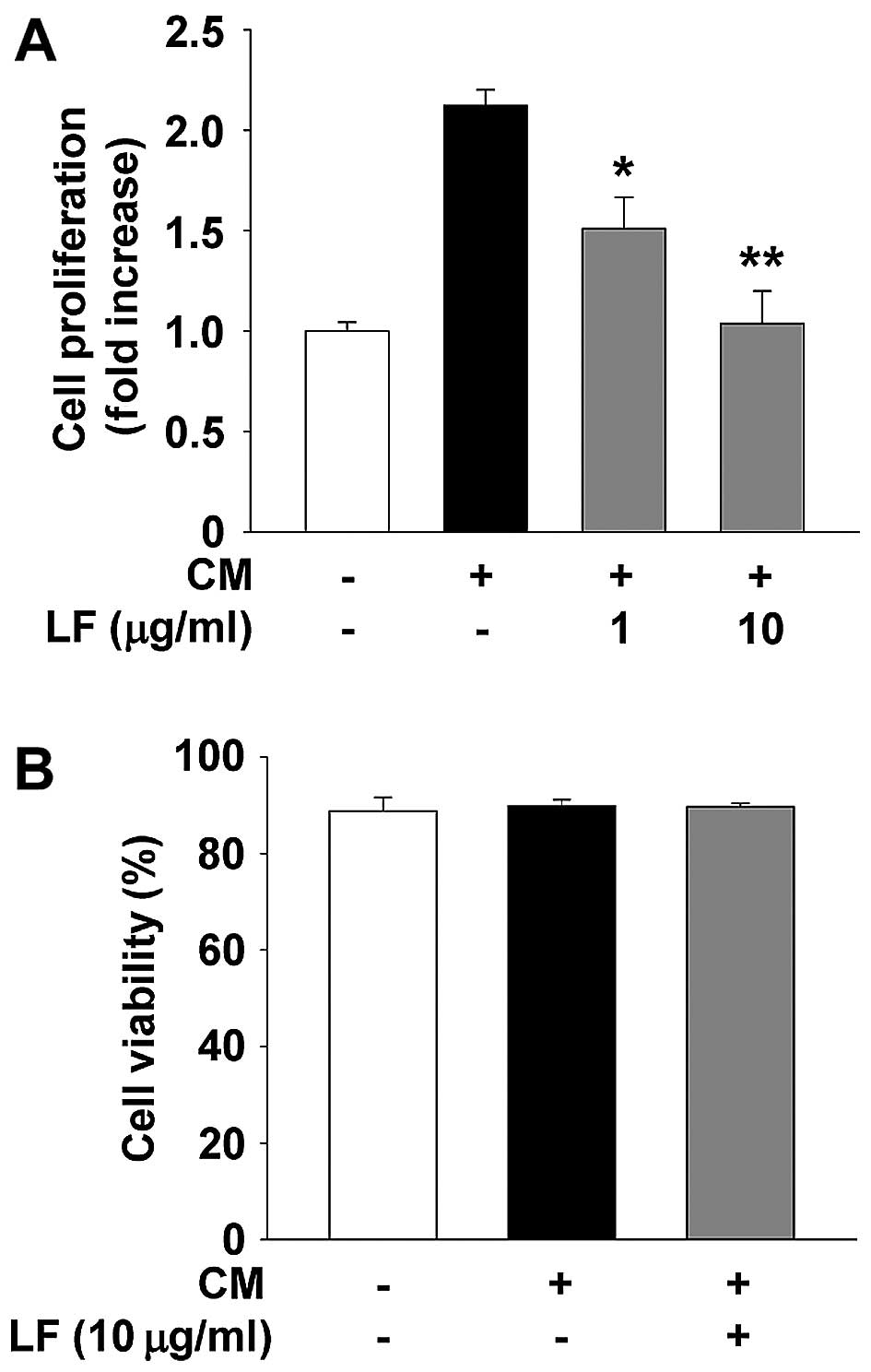
We first examined the ability of LF to modulate cell
proliferation of HUVECs. LF treatment suppressed endothelial cell
proliferation in a dose-dependent manner (Fig. 1A) and did not alter cell viability
(Fig. 1B), indicating that LF
inhibition of endothelial cell proliferation is not mediated by
induction of apoptosis or cytotoxicity. This finding is similar to
the patterns of LF in other cell types as previously reported
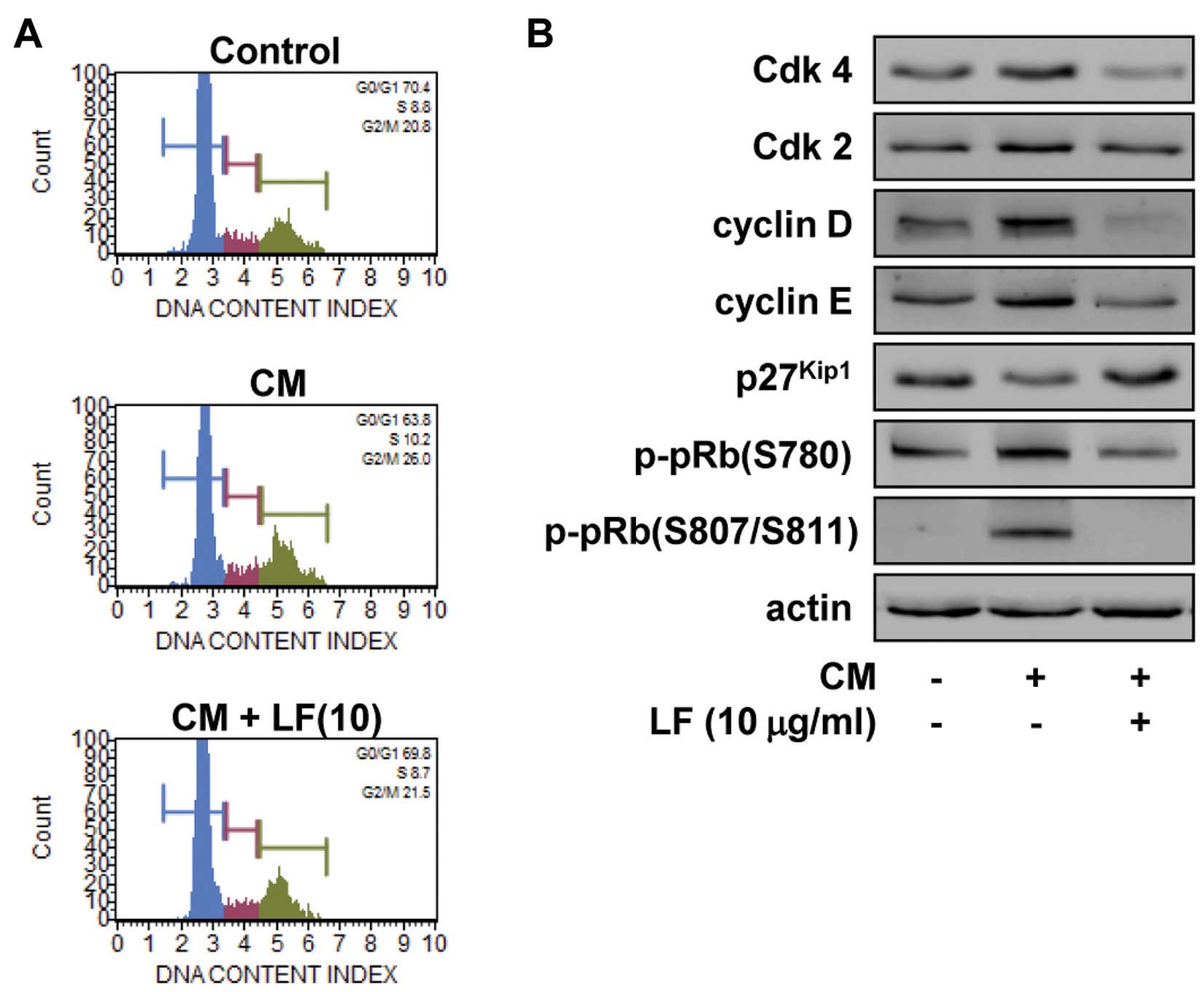
(32,33). We next examined the effect of LF on
the cell cycle by DNA content analysis (Fig. 2A). LF treatment prevented the
increase in S phase (10.2 vs. 8.7%) and G2/M phase (26.0
vs. 21.5%) and the decrease in G1 phase (63.8 vs. 69.8%)
associated with mitogenic stimulation, similar to those of
untreated controls. These observations suggest that LF inhibits the
transition from G1 to S phase, leading to G1
arrest, which is well correlated with inhibition of cell
proliferation (Fig. 1A). We have
previously reported that the ethanolic extract and ethyl caffeate,
a natural phenolic compound isolated from LF, had
growth-suppressive activity in different types of cancer cells
including non-small cell lung and ovarian cancer, this inhibitory
effect was found to be mediated by downregulation of Cdks and
cyclins (32,33). Based on these findings, we analyzed
the changes of cell cycle-related proteins such as Cdks, cyclins
and Cdk inhibitor p27Kip1 in LF-treated HUVECs. As shown
in Fig. 2B, LF treatment markedly
reduced the expression of Cdks and cyclins, enhanced the levels of
p27Kip1, leading to inhibition of pRb phosphorylation in
response to mitogenic stimulation. These findings clearly show the
regulatory effects of LF on cell cycle progression and
proliferation in HUVECs.
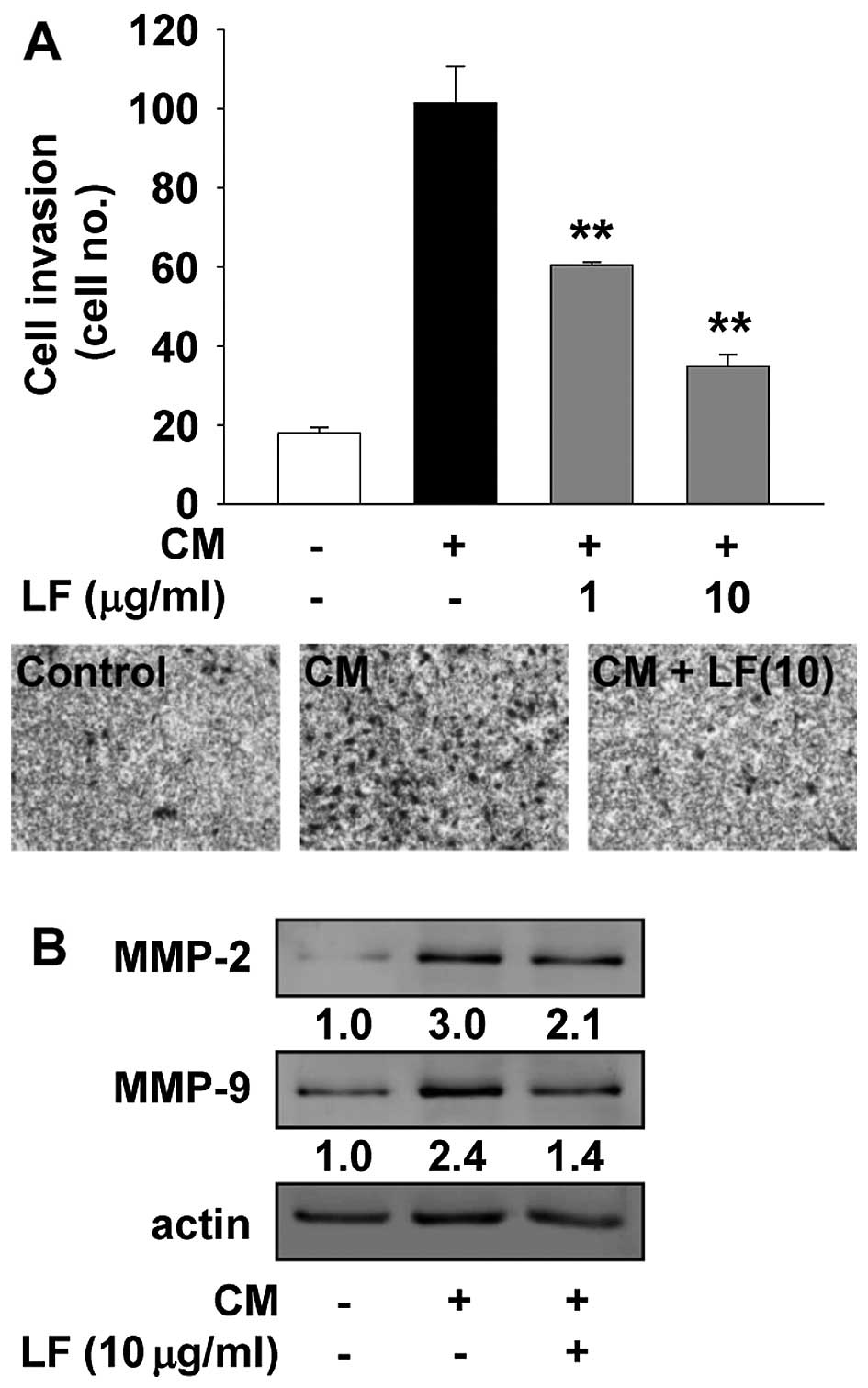
LF inhibits endothelial cell invasion and
tube formation
The release of a variety of biologically active
molecules from ECM and cell surface components by MMP-mediated
proteolytic degradation is associated with the regulation of
cellular behavior such as cell adhesion, migration and invasion
(6–8). Thus, we next examined the changes of
cell invasion and MMP expression in LF-treated HUVECs. As shown in
Fig. 3, LF treatment markedly
inhibited the invasion and MMP expression in response to mitogenic
stimuli, suggesting that the anti-invasive activity of LF may be
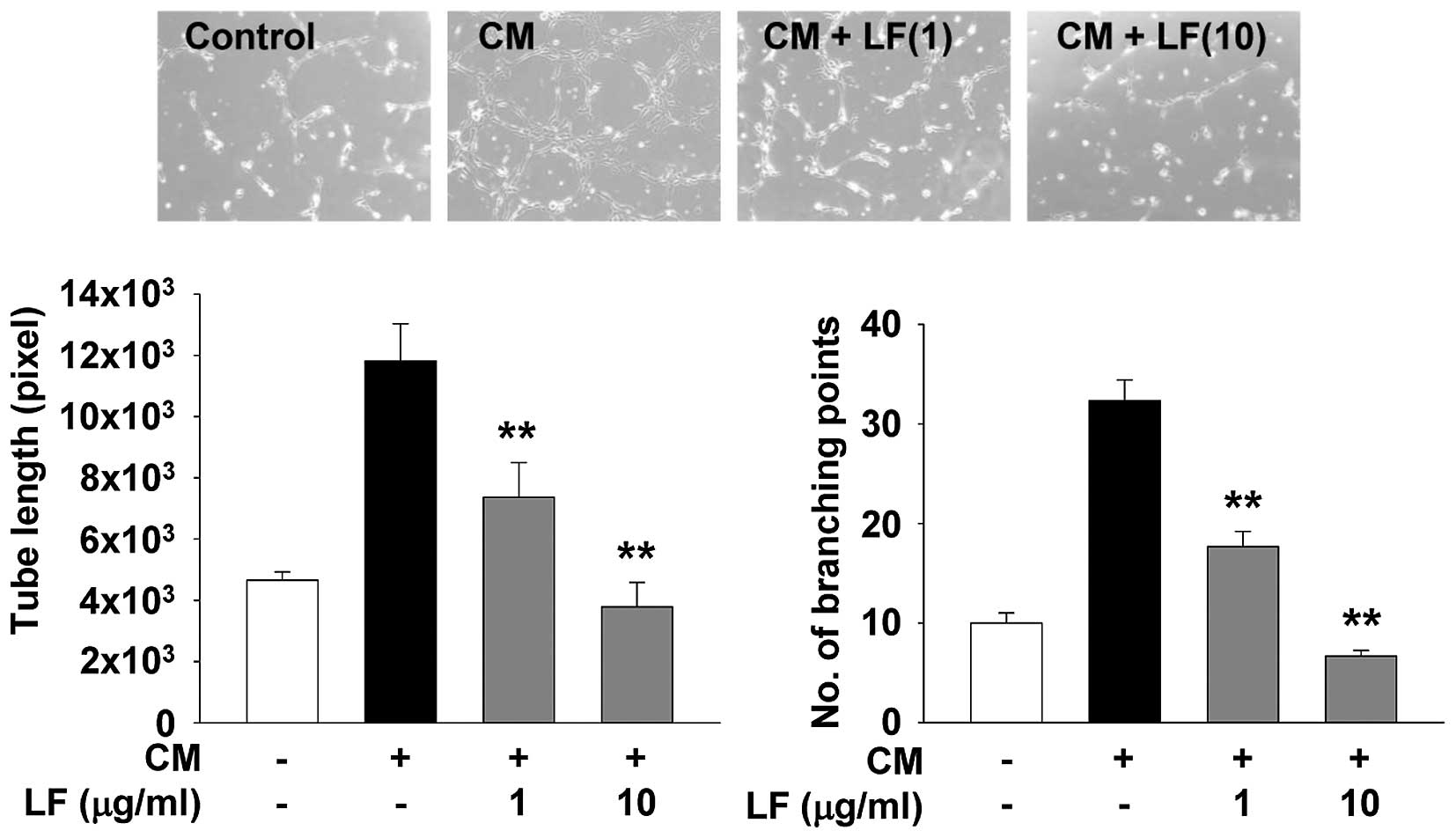
mediated through downregulation of MMP expression. In addition, LF
treatment completely suppressed mitogen-induced capillary-like
structure formation to the levels observed in untreated controls
(Fig. 4). Collectively, these
findings clearly show the pharmacological roles of LF in regulating
endothelial cell proliferation, invasion and tube formation.
Anti-angiogenic effects of LF are
mediated through the down-regulation of mitogenic signaling
pathways and VE-cadherin expression
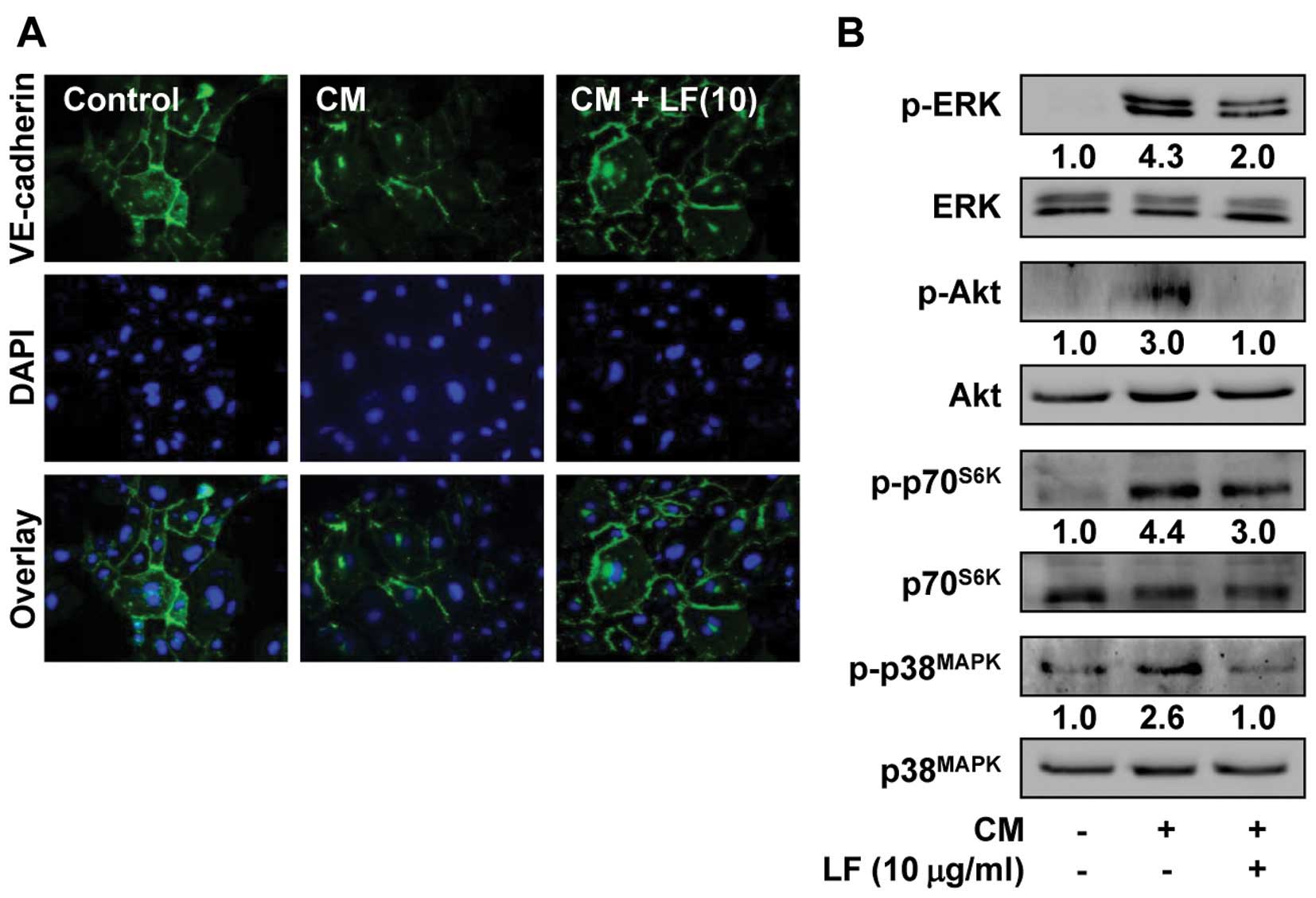
Distribution of VE-cadherin at cell-cell contacts
has been reported to enhance the stability of adherens junctions,
resulting in maintenance of endothelial barrier function (9,10).
Angiogenic factors such as VEGF-A disrupt the loss of VE-cadherin
from the endothelial cell surfaces and induces endothelial cell
responses such as permeability, proliferation, invasion and tube
formation (13,14). Change of VE-cadherin function can be
assessed by the levels of VE-cadherin detectable at cell-cell
contacts. As shown in Fig. 5A, LF
treatment prevented the mitogen-induced loss of VE-cadherin from
cell-cell contacts, similar to untreated controls. To further
investigate the molecular mechanisms by which LF modulates
mitogen-induced endothelial cell responses, we examined the changes
in activation of mitogenic signaling pathways including ERK, Akt,
p70S6K and p38MAPK (40). As shown in Fig. 5B, LF treatment markedly inhibited
the mitogen-induced phosphorylation/activation of ERK, Akt,
p70S6K and p38MAPK in HUVECs. Collectively,
these observations suggest that anti-angiogenic activities of LF
may be mediated at least in part through the inactivation of
mitogenic signaling pathways and redistribution of VE-cadherin at
cell-cell contacts.
Discussion
Overexpressed receptor tyrosine kinases (RTKs) and
dysregulation of RTK downstream signaling pathways are closely
associated with pathological conditions including cancer (40). Selective inhibition or normalization
of RTK-mediated signaling pathways has widely been appreciated as a
rational therapeutic strategy. However, many drugs that target RTKs
and the downstream signaling networks frequently lead to drug
resistance and adverse effects in clinical trials or use.
Therefore, natural products which act simultaneously on multiple
molecular targets can sometimes be of therapeutic benefit in
treating diseases. LF has been used for improving liver function,
as well as in inflammatory and infectious disorders. These
applications of LF may be mediated through anti-inflammatory,
anti-oxidative and anti-hepatotoxic effects (25–28).
We have previously reported that the ethanolic extract of LF
inhibits proliferation and migration of non-small cell lung cancer
cells through the inactivation of signaling pathways such as ERK,
Akt and p70S6K, and the downregulation of epidermal
growth factor receptor, integrin β1 and integrin-linked kinase
(ILK) (32). In addition, ethyl
caffeate, a natural phenolic compound isolated from the ethanolic
extract of LF, exerts anti-proliferative, anti-migratory and
anti-invasive activities in ovarian cancer cells. The mechanism of
these effects involves suppression of signaling pathways including
ERK, Akt, p70S6K and p38MAPK, and
downregulation of human epidermal growth factor receptor 2,
fibroblast growth factor receptor-1, vascular endothelial growth
factor receptor-2, integrin α3β1, ILK and N-cadherin (33).
In the present study, we demonstrate for the first
time that LF inhibits mitogen-induced endothelial cell
proliferation, invasion and tube formation. These anti-angiogenic
activities of LF were found to be mediated through the inactivation
of mitogenic signaling pathways, redistribution of VE-cadherin at
cell-cell contacts and downregulation of MMP expression. Based on
the regulatory effects of LF on MMP expression, we examined the
ability of LF to alter the levels of TIMP-2, an endogenous
inhibitor of MMPs which has been known to regulate cell
proliferation and differentiation through MMP-dependent and/or
MMP-independent mechanism (15,16,21).
LF treatment showed little or no change of TIMP-2 expression in
mitogen-treated HUVECs (data not shown). These findings indicate
that anti-angiogenic effects of LF may be mediated through the
regulation of MMP-2 and MMP-9, but not that of TIMP-2. However, it
cannot exclude the possibility that LF may modulate the expression
and activity of other MMP and TIMP family members.
In conclusion, the present study demonstrates the
pharmacological roles and mechanisms of LF in the regulation of
angiogenesis, and warrants further evaluation and development of LF
for the prevention and treatment of pathological states associated
with angiogenesis.
Acknowledgments
This study was supported by the High Value-added
Food Technology Development Program (112060-3) through the Ministry
of Agriculture, Food and Rural Affairs, and by the Basic Science
Research Program (2010-0021913, 2014R1A1A2058015) through the
National Research Foundation of Korea, Ministry of Education.
References
|
1
|
Cristofanilli M, Charnsangavej C and
Hortobagyi GN: Angiogenesis modulation in cancer research: novel
clinical approaches. Nat Rev Drug Discov. 1:415–426. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Folkman J: Angiogenesis: an organizing
principle for drug discovery? Nat Rev Drug Discov. 6:273–286. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Carmeliet P and Jain RK: Principles and
mechanisms of vessel normalization for cancer and other angiogenic
diseases. Nat Rev Drug Discov. 10:417–427. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cook KM and Figg WD: Angiogenesis
inhibitors: current strategies and future prospects. CA Cancer J
Clin. 60:222–243. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nyberg P, Xie L and Kalluri R: Endogenous
inhibitors of angiogenesis. Cancer Res. 65:3967–3979. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Stetler-Stevenson WG: Matrix
metalloproteinases in angiogenesis: a moving target for therapeutic
intervention. J Clin Invest. 103:1237–1241. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kessenbrock K, Plaks V and Werb Z: Matrix
metalloproteinases: regulators of the tumor microenvironment. Cell.
141:52–67. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bourboulia D and Stetler-Stevenson WG:
Matrix metalloproteinases (MMPs) and tissue inhibitors of
metalloproteinases (TIMPs): positive and negative regulators in
tumor cell adhesion. Semin Cancer Biol. 20:161–168. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dejana E, Orsenigo F and Lampugnani MG:
The role of adherens junctions and VE-cadherin in the control of
vascular permeability. J Cell Sci. 121:2115–2122. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kim SH, Cho YR, Kim HJ, Oh JS, Ahn EK, Ko
HJ, Hwang BJ, Lee SJ, Cho Y, Kim YK, et al: Antagonism of
VEGF-A-induced increase in vascular per meability by an integrin
α3β1-Shp-1-cAMP/PKA pathway. Blood. 120:4892–4902. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Herren B, Levkau B, Raines EW and Ross R:
Cleavage of beta-catenin and plakoglobin and shedding of
VE-cadherin during endothelial apoptosis: evidence for a role for
caspases and metalloproteinases. Mol Biol Cell. 9:1589–1601. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Grazia Lampugnani M, Zanetti A, Corada M,
Takahashi T, Balconi G, Breviario F, Orsenigo F, Cattelino A,
Kemler R, Daniel TO, et al: Contact inhibition of VEGF-induced
proliferation requires vascular endothelial cadherin, beta-catenin,
and the phosphatase DEP-1/CD148. J Cell Biol. 161:793–804. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
George SJ and Dwivedi A: MMPs, cadherins,
and cell proliferation. Trends Cardiovasc Med. 14:100–105. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Spring K, Chabot C, Langlois S, Lapointe
L, Trinh NT, Caron C, Hebda JK, Gavard J, Elchebly M and Royal I:
Tyrosine phosphorylation of DEP-1/CD148 as a mechanism controlling
Src kinase activation, endothelial cell permeability, invasion, and
capillary formation. Blood. 120:2745–2756. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Brew K and Nagase H: The tissue inhibitors
of metalloproteinases (TIMPs): an ancient family with structural
and functional diversity. Biochim Biophys Acta. 1803:55–71. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Seo D-W, Li H, Guedez L, Wingfield PT,
Diaz T, Salloum R, Wei BY and Stetler-Stevenson WG: TIMP-2 mediated
inhibition of angiogenesis: an MMP-independent mechanism. Cell.
114:171–180. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Qi JH, Ebrahem Q, Moore N, Murphy G,
Claesson-Welsh L, Bond M, Baker A and Anand-Apte B: A novel
function for tissue inhibitor of metalloproteinases-3 (TIMP3):
inhibition of angiogenesis by blockage of VEGF binding to VEGF
receptor-2. Nat Med. 9:407–415. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jung KK, Liu XW, Chirco R, Fridman R and
Kim HR: Identification of CD63 as a tissue inhibitor of
metalloproteinase-1 interacting cell surface protein. EMBO J.
25:3934–3942. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Seo DW, Li H, Qu CK, Oh J, Kim YS, Diaz T,
Wei B, Han JW and Stetler-Stevenson WG: Shp-1 mediates the
antiproliferative activity of tissue inhibitor of
metalloproteinase-2 in human microvascular endothelial cells. J
Biol Chem. 281:3711–3721. 2006. View Article : Google Scholar :
|
|
20
|
Seo DW, Kim SH, Eom SH, Yoon HJ, Cho YR,
Kim PH, Kim YK, Han JW, Diaz T, Wei BY, et al: TIMP-2 disrupts
FGF-2-induced downstream signaling pathways. Microvasc Res.
76:145–151. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Stetler-Stevenson WG: Tissue inhibitors of
metalloproteinases in cell signaling: metalloproteinase-independent
biological activities. Sci Signal. 1:re62008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Coussens LM, Fingleton B and Matrisian LM:
Matrix metalloproteinase inhibitors and cancer: trials and
tribulations. Science. 295:2387–2392. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Vandenbroucke RE and Libert C: Is there
new hope for therapeutic matrix metalloproteinase inhibition? Nat
Rev Drug Discov. 13:904–927. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hwang BY, Lee JH, Koo TH, Kim HS, Hong YS,
Ro JS, Lee KS and Lee JJ: Furanoligularenone, an eremophilane from
Ligularia fischeri, inhibits the LPS-induced production of nitric
oxide and prostaglandin E2 in macrophage RAW2647 cells. Planta Med.
68:101–105. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lee KH and Choi EM: Analgesic and
anti-inflammatory effects of Ligularia fischeri leaves in
experimental animals. J Ethnopharmacol. 120:103–107. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Choi EM: Ligularia fischeri leaf extract
prevents the oxidative stress in DBA/1J mice with type II
collagen-induced arthritis. J Appl Toxicol. 27:176–182. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shang YF, Kim SM, Song DG, Pan CH, Lee WJ
and Um BH: Isolation and identification of antioxidant compounds
from Ligularia fischeri. J Food Sci. 75:C530–C535. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Choi J, Park JK, Lee KT, Park KK, Kim WB,
Lee JH, Jung HJ and Park HJ: In vivo antihepatotoxic effects of
Ligularia fischeri var. spiciformis and the identification of the
active component, 3,4-dicaffeoylquinic acid. J Med Food. 8:348–352.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cha KH, Song DG, Kim SM and Pan CH:
Inhibition of gastrointestinal lipolysis by green tea, coffee, and
gomchui (Ligularia fischeri) tea polyphenols during simulated
digestion. J Agric Food Chem. 60:7152–7157. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jeong SH, Koo SJ, Choi JH, Park JH, Ha J,
Park HJ and Lee KT: Intermedeol isolated from the leaves of
Ligularia fischeri var. spiciformis induces the differentiation of
human acute promyeocytic leukemia HL-60 cells. Planta Med.
68:881–885. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xie WD, Li X, Weng CW, Liu SS and Row KH:
Fischerisin A and B, cytotoxic sesquiterpenoid-geranylhydroquinones
from Ligularia fischeri. Chem Pharm Bull (Tokyo). 59:511–514. 2011.
View Article : Google Scholar
|
|
32
|
Cho YR, Kim JK, Kim J, Oh J and Seo DW:
Ligularia fischeri regulates lung cancer cell proliferation and
migration through down-regulation of epidermal growth factor
receptor and integrin β1 expression. Genes Genom. 35:741–746. 2013.
View Article : Google Scholar
|
|
33
|
Lee HN, Kim JK, Kim JH, Lee SJ, Ahn EK, Oh
JS and Seo DW: A mechanistic study on the anti-cancer activity of
ethyl caffeate in human ovarian cancer SKOV-3 cells. Chem Biol
Interact. 219:151–158. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kim HJ, Cho YR, Kim SH and Seo DW:
TIMP-2-derived 18-mer peptide inhibits endothelial cell
proliferation and migration through cAMP/PKA-dependent mechanism.
Cancer Lett. 343:210–216. 2014. View Article : Google Scholar
|
|
35
|
Kim HJ, Ko HY, Choi SW and Seo DW:
Anti-angiogenic effects of Siegesbeckia glabrescens are mediated by
suppression of the Akt and p70S6K-dependent signaling
pathways. Oncol Rep. 33:699–704. 2015.
|
|
36
|
Cho YR, Choi SW and Seo DW: The in vitro
antitumor activity of Siegesbeckia glabrescens against ovarian
cancer through suppression of receptor tyrosine kinase expression
and the signaling pathways. Oncol Rep. 30:221–226. 2013.PubMed/NCBI
|
|
37
|
Lee HN, Joo JH, Oh JS, Choi SW and Seo DW:
Regulatory effects of Siegesbeckia glabrescens on non-small cell
lung cancer cell proliferation and invasion. Am J Chin Med.
42:453–463. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yoon HJ, Cho YR, Joo JH and Seo DW:
Knockdown of integrin α3β1 expression induces proliferation and
migration of non-small cell lung cancer cells. Oncol Rep.
29:662–668. 2013.
|
|
39
|
Cho YR, Kim JH, Kim JK, Ahn EK, Ko HJ, In
JK, Lee SJ, Bae GU, Kim YK, Oh JS, et al: Broussonetia kazinoki
modulates the expression of VEGFR-2 and MMP-2 through the
inhibition of ERK, Akt and p70S6K-dependent signaling
pathways: its implication in endothelial cell proliferation,
migration and tubular formation. Oncol Rep. 32:1531–1536.
2014.PubMed/NCBI
|
|
40
|
Lemmon MA and Schlessinger J: Cell
signaling by receptor tyrosine kinases. Cell. 141:1117–1134. 2010.
View Article : Google Scholar : PubMed/NCBI
|