1. Introduction
Cancer is a fatal disease and has created an urgent
challenge for humankind. Malignant tumors are one of the leading
causes of mortality in the majority of the world's countries, being
slightly behind cardiovascular diseases among seniors and almost at
the same level with infectious diseases and injuries among young
patients of an active working age (1).
The main sites contributing to the oncologic
mortality rate are the lungs, breast, colon and prostate (1,2). Usage
of new generation targeted chemotherapeutic agents has helped to
prolong the lives of patients with solid tumors in economically
developed countries (3). However,
this success has not been extended to the treatment of glial brain
tumors.
Glioblastoma multiforme (GBM) or grade IV
astrocytoma according to the World Health Organization
classification is the most widespread, highly invasive primary
glial brain tumor that accounts for more than 50% of all primary
brain tumors and ~20% of all intracranial neoplasms. The frequency
of diagnosis is 4.65–5.26 cases/100,000 people, while only in the
US more than 17,000 new cases are diagnosed each year. The
treatment of GBM patients usually involves tumor removal,
radiotherapy and chemotherapy. The prognosis is not promising. If
patients with glioblastoma follow all procedures of the modern
complex treatment, the median survival is 16.2 months for patients
20–44 years of age, 7.9 months for patients 45–69 years of age, and
3.2 months for patients over 70 years of age (5,6).
One of the main reasons for the therapeutic
resistance of GBM is attributed to cancer stem cells (CSCs). There
are no pharmaceutical means for effective elimination of these
cells. In vitro and in vivo experiments have shown
that CSCs could be affected by targeted treatment in order to block
adhesion, proliferation and migration as well as some other key
functions. The main goal of this therapy is to reach membranous
targets of intracellular signaling pathways of CSCs that have not
been affected by neoplastic transformation. Stimulating such a
difficult target requires creating radically innovative
biotechnological approaches relevant to the complexity of the
object. The key methods of dealing with this issue include
application of cell and post-genomic technologies. The main focus
of this review is to organize the scientific and technical
information on the creation and methodology of using cell and
post-genomic technologies in the complex treatment of primary brain
tumors.
Materials used were scientific and technical
studies, patents and current literature on the topic available in
the Web of Science, Scopus and PubMed databases.
2. Existing methods of glial brain tumor
treatment
The current standard of treating malignant tumors
includes a unified complex of procedures, such as tumor removal,
systemic chemotherapy and radiotherapy (7). Surgical treatment must be extensive
and radical. If the tumor is located deep in the brain matter or
close to vital centers of the brain stem, it requires the use of
high-precision neurosurgical equipment or modern radiosurgical
systems (8,9).
Traditional radiotherapy includes 25–30 fractions of
X-ray treatment for 5–6 weeks. The frequency of treatment is 5
times weekly with a dose of 1.8–2.0 Gy using three-field
arrangement. The life expectancy is correlated with a total boost
dose of the amount up to 60 Gy (10). Further increase in the dose is
limited by the development of radiation necrosis (11,12).
There is an opinion that total brain irradiation with 50 Gy helps
to prevent relapse of glioblastoma although a systematic
irradiation with 50 Gy for 3–5 years leads to radiation damage of
the CNS and inevitable mental disability. In case of a favorable
result, a patient may be seriously disabled with a severe
post-radiation intellectual-mnestic and neurological disorder
(13–15).
Temozolomide is usually chosen for glioblastome
multiforme treatment (16). In
addition, PVC drug combination [procarbazine, lomustine (CCNU),
vincristine] is effective, while alternative methods include
cyclophosphamide (CAP; doxorubicin, cisplatin) and a combination of
lomustine (CCNU), doxorubicin and teniposide. Treatment is used
after dehydration and together with corticosteroids and symptomatic
medication (17–19). Bevacizumab is currently the most
widely used targeted medication. The effectiveness of chemotherapy
ranges from 20 to 60% (20).
Partially, the first stage of standard brain tumor
treatment is effective. As a rule, this stage is followed by a
'clean slate' in the disease progression. Relapse may take the form
of neurological deficit and continuation of tumor growth that
usually is located in the tumor bed and occurs in 95% of cases in
7–20 months after the surgery (4,6). GBM
relapse requires a second surgery and X-ray treatment or extended
chemotherapy if radiotherapy is not possible. Targeted therapeutic
methods used for GBM have not been proven to have a significant
influence on survival rates for this type of patient.
Thus, traditional treatment of CNS tumors is based
on the classic principles of tumor treatment and is concentrated on
removing cancerous cells. The majority of modern methods and
techniques have almost zero effect on the CSCs that are believed to
be the main cause of glioblastoma therapeutic resistance.
3. Concept of cancer stem cells
The term 'CSC' is an artificial product of
experimental medicine in a way. The distinguishing feature of these
cells is their ability to trigger tumor creation when implanted
into experimental animal bodies even in microscopic quantities.
Originally these cells were identified in leucosis, breast cancer
and gliobastoma (21).
Characteristics of CSCs can be found in CD133+ cells of
glioblastoma, ESA+/CD44+CD24−/low
of breast, CD44+ of prostate cancer, and
ESA+/CD44+CD24− of pancreatic
cancer, and side population of cells without Hoechst pigment in the
case of ovarian cancer (22–24).
The question of the origin of CSCs still has no
definite answer. They are highly probable to appear after
disruption of tissue mechanisms that control clonogenic cell
proliferation that causes the possibility to reverse transitions
from normal to altered or modified cell phenotype. Furthermore,
carcinogens influence oncogene activation, suppress
tumor-suppressor genes, and continues the damage of epigenetic
control of gene expression. As a result, there appears to be a
clonal version of the cells having maximum autonomy, individuality
and complete independence from their medium. These cells develop
mechanisms that prevent depletion of telomeres and become
immortalized giving them a strategic advantage in their fight for
survival. This concept was mainly described in the studies of
Duesberg et al who believed carcinogenesis to be a type of
species formation (25).
The idea regarding cancer as being a stage of
species evolution instead of just a disease was expressed by Julian
Huxley and some evolutionary biologists in 1956. The theory of
species formation that is a logical offspring of the mutation
theory partially explains the mechanisms of the appearance of CSCs.
Yet, it lacks argumentation for the fact that this type of cell has
both the ability of infinite self-renewal and production of
different cell types depending on the medium in which they have
been placed.
According to a second version, CSCs appear to be the
result of the neoplastic transformation of normal stem cells.
Neurogenesis in an adult brain of mammals was described for the
first time by Altman in the 1960's (26). Thirty years later, neural stem cells
(NSCs) were extracted from an adult brain of a mammal and can be
considered as the most significant breakthrough of 20th century
neurobiology (27). Originally NSCs
were discovered in the subependymal zone in the lateral ventricles
of adult mice and the rat brain and were later extracted from the
hippocampus of adult primates and humans. NSCs are able to
constantly regenerate, migrate and produce cells of different types
(28). The typical germinal zone of
an adult brain also contains neural progenitor cells that are
direct descendants of NSCs and have the highest proliferative
activity of germinal zone cells (29,30).
Neural stem and progenitor cells are the most
probable source of GBM CSCs. This is indicated by their similarity
on the basis of the main immunocytochemical marker of the cell
surface (21) and identical nature
of 63.5% of the NSC and CSC proteome in human glioblastoma
(31). We cannot deny the
experimental data. The transfer of Myc and Ras oncogenes into
nervous system cells causes the creation of invasive tumors only
after they have entered NSCs (32).
The NSC karyotype is very unstable, and in the case of long
cultivation these cells accumulate chromosomal anomalies that
become replicated in the following passages. There have been
registered cases of trisomy of chromosomes 7, 12, 17, 19 and X,
different forms of aneuploidy, an elevated level of telomerase
expression, and cases of spontaneous neoplastic transformation of
NSCs after a series of in vitro passages (33–35).
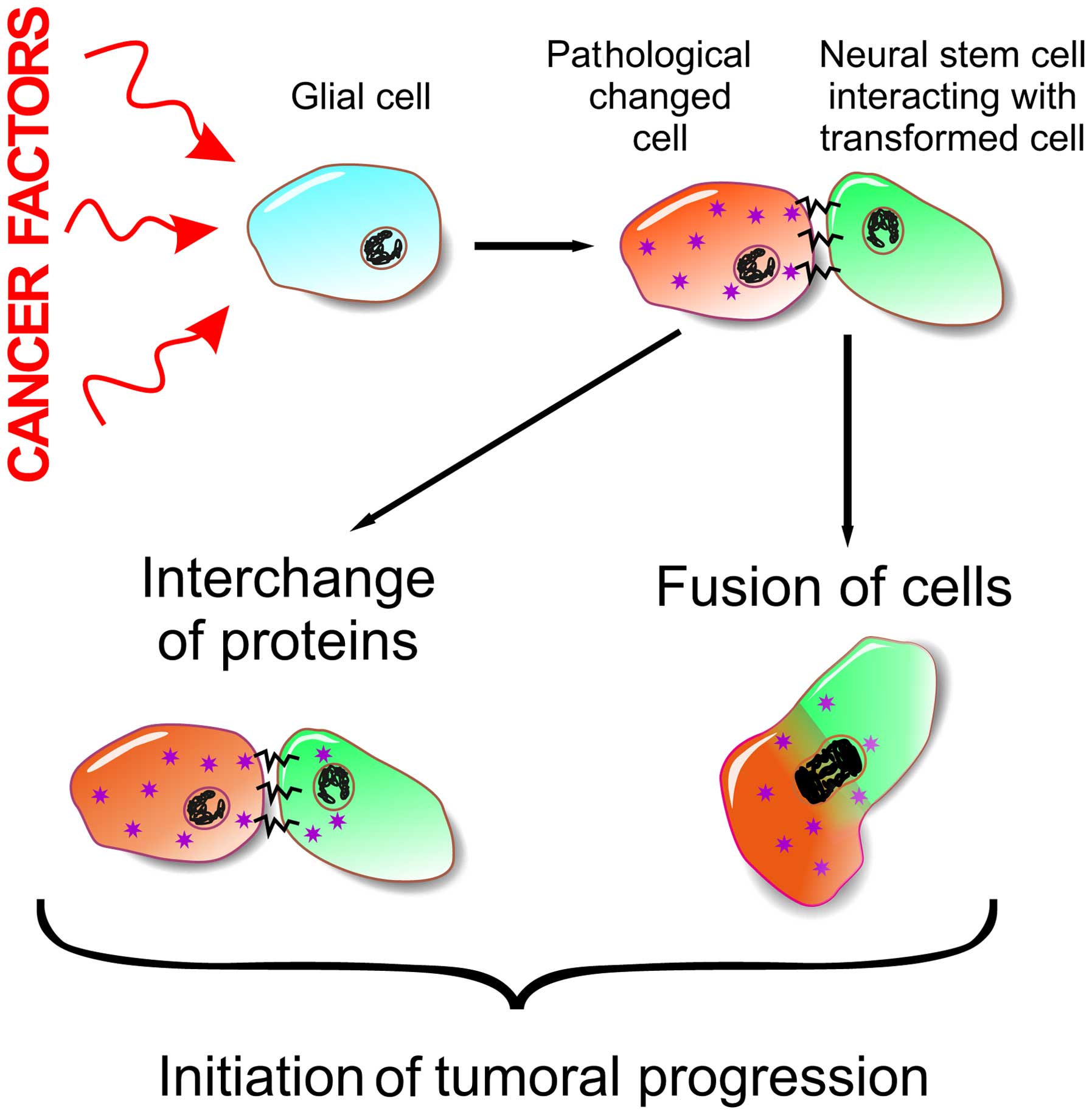
It is obvious that constant proliferation during a
lifetime of an individual results in the accumulation of mutations
that become a starting point for tumor development. Another
possible mechanism of CSC development can be reprogramming of NSCs
by oncogene expression products as a result of their interaction
with pathologically altered cells, elements of the cell matrix
(Fig. 1) or during cell fusion
(36,37).
CSCs have a system-building role. They organize
processes of invasive growth, secure formation of timorous blood,
lymph and nervous networks, interact with fibroblasts and vascular
endothelium cells creating a barrier that protects cancer cells
from chemical medication. CSCs are able to restore damaged DNA
using CD133+ cells to produce a special ferment called
O6-methylguanine-DNA-methyltransferase or MGMT factor.
This mechanism along with activation of ATM, Rad17, Chk2 and Chk1
proteins allows a cell with a damaged genome to pass a basic stage
of the cell cycle and proliferate actively, resulting in tumor
relapse after its removal. The ability of the rapid restoration of
the neoplastic population totally depends on the amount of CSCs
(38–40).
The amount of CSCs in a tumor is a very important
and dynamic factor. According to the literature, the number of CSCs
in GBM tissues varies between 0.1 and 11% and can reach up to 50%
of the cell population. This partially explains its aggressiveness
(41,42). This parameter is one of the main
characteristics of the tumor condition. Research by Shen et
al showed that CD133+ phenotype cells (main CSC
marker) constitute 87.2% of the glioma C6 cell population that can
be extracted from glioma spheres freely moving in vitro. The
CD133 marker can reveal either the CSC potential of a cell or its
epithelial nature, or both. Taking into consideration the location
of NSCs, noting this marker in C6 cells is not unexpected. At the
same time the number of GFAP-positive cells does not exceed 0.95%.
After the cells are attached to the surface, the amount of
CD133+ cells is reduced to 30.2%, while the number of
GFAP-positive cells reaches 50.1% (43).
In an in-house study, 96% of C6 glioma cells
extracted from freely circulating glioma spheres became stained by
antibodies against nestin. After adhesion to the substrate the
number of nestin-positive cells was reduced to 52.5%. Nestin is one
of the most important stem cell markers (44). Adhesion to the substrate is clearly
an important mechanism of regulating a number of CSCs, otherwise
CD133 and nestin expression is mostly characteristic of migrating
NSCs that suggests the hypothesis of transforming NSCs into CSCs in
the conditions of atypical influence of stromal or cellular
microenvironment.
CSCs are a main target in GBM treatment. The
majority of immunocytochemical markers of these cells such as
CD133, nestin, L1CAM (CD171), CD15, CD44, CD81, TPT1 and A2B5
(45–47) have been discovered and the main
signaling pathways that they use for proliferation such as Notch,
Hedgehog-Gli, RTK-Akt, BMPs/TGFβ, Wnt-β-catenin and STAT3 have been
identified (21,48). However, there is no medication for
the effective elimination of these cells. The possibilities of
affecting certain target characteristic of these cells are being
studied. For example, sunitinib affects the MAPK pathway. Rapamycin
inhibits the OPI3K/Akt pathway, cyclopamine blocks the SHH pathway,
while there are still no effective and safe medication for
suppressing Wnt signal transduction cascade of CSCs (49). Effective drugs blocking the adhesion
mechanisms of CSCs have not yet been developed.
Thus, CSCs are a crucial element in glioblastoma
development, a key moderator of invasive growth processes and a
universal form of tumor cell survival. Various arguments suggest
that CSCs of primary brain tumors originate from NSCs of the human
brain, although these suggestions support alternative ideas. A
large number of CSCs indicates a very bleak prognosis. Stem
features of cells (cell rounding, disappearance of some surface
markers, disruption of intercellular contacts) appear to be a
fundamental survival mechanism for a cancer cell that requires
development of completely new methods for managing this
process.
4. Theoretical prerequisites for using cell
and post-genome technologies for treating glioblastoma
Stem cells have been successfully used in oncology
for more than 50 years. Their usage in the complex treatment of
malignant tumors in the 1960 became a breakthrough in science.
Today they are widely used in post-chemotherapy for reconstructing
the hematopoietic system, treating immunodeficiency, creating
anticancer vaccines, cytotoxic lymphocytes and dendritic cells.
Stem cell transplants have become one of the central practices in
the treatment of serious diseases and CNS disorders. The
neurotrophic and neoplastic effects of stem cells on pathologically
modified nerve tissue have been proven, while further plans for
their application are connected with brain tumor treatment. Key
mechanisms of the anticancer effect of stem cells include their
targeted migration to the tumor nodule and intercellular
interaction in the neoplasia area.
The phenomenon of stem cell-targeted migration to
the area of trauma, ischemic or neoplastic damage of the brain have
been closely studied by neurobiologists. Our research group
published a number of experimental and theoretical studies based on
the subject. Numerous scientists treat this phenomenon only as a
repair mechanism. Researchers have identified 80 cytokines and
>20 types of receptors controlling this process. The key role
here is attributed to the interaction of stromal cell-derived
factor (SDF-1α) with the CXCR4 receptor of the stem cell membrane.
The importance of stem cell factor (SCF), hepatocyte growth factor
(HGF), vascular endothelial growth factor (VEGF), high-mobility
group protein B1 (HMGB1), monocyte chemotactic protein 1 (MCP1) and
other ligands released by damaged tissues has been proven (50). It is traditionally believed that the
main sources of cytokines are damaged neurons, astrocytes,
microglial cells and retrograde elements of the intercellular
matrix released into blood vessels.
However, experimental data show that the original
source of cytokines can be tumor cells themselves (51,52).
Having autonomy, they can synthesize their own cytokines. The
scientific literature has a detailed description of neuroepithelial
tumors able to produce tenastin, fibronectin, laminin and collagen
that change the direction of stem cell migration (53). One of the key parameters in
glioblastoma biology is hypoxia that causes excessive production of
hypoxia-induced factor (HIF) molecules. They are the main inducing
substances for producing SDF-1α, SCF, HGF and other cytokines that
stimulate stem cell migration (54).
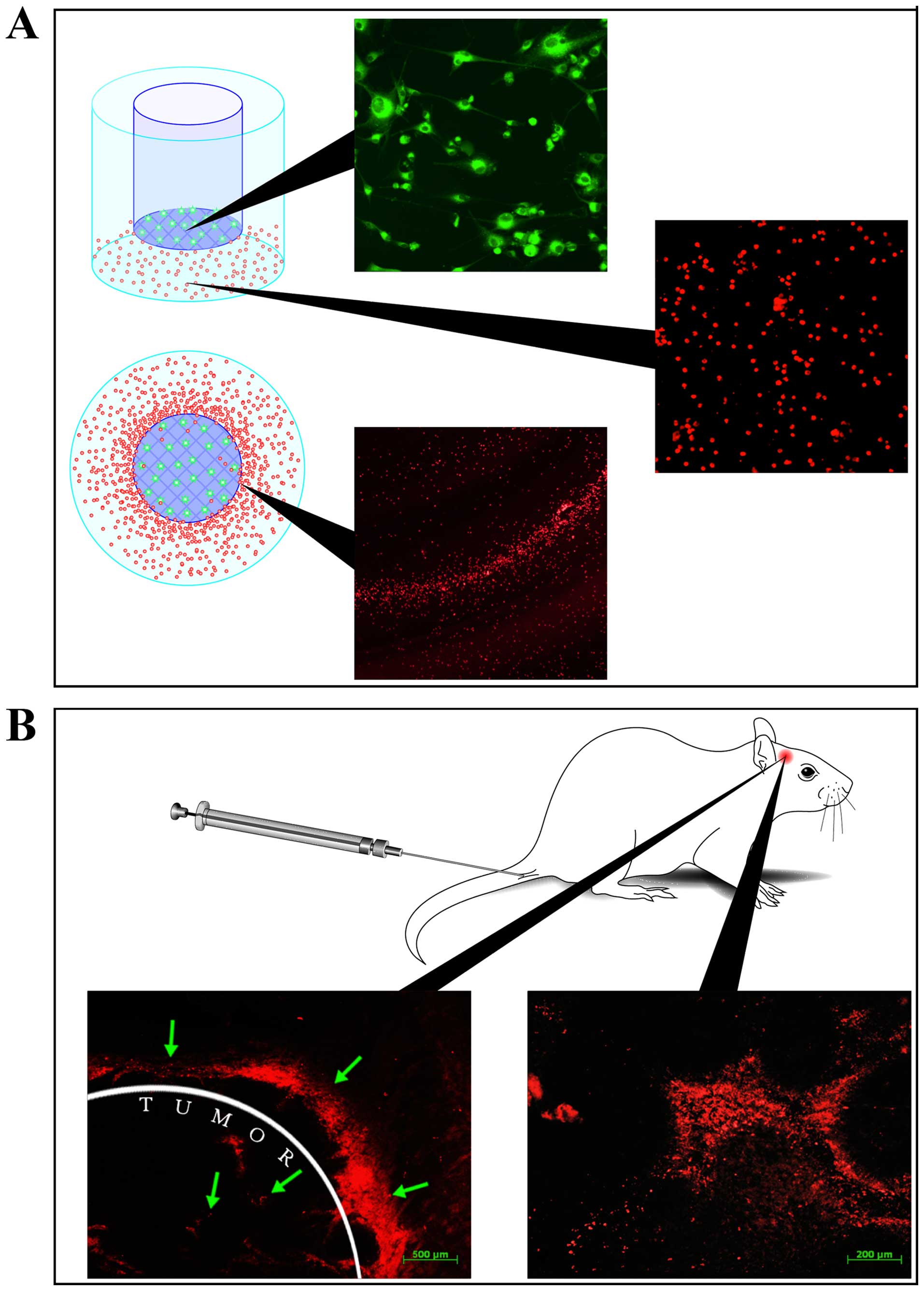
Cytokines cause stem cells to migrate towards cancer
cells. In a study by Moore et al, it was suggested that the
absolute majority of CD34+ stem cells was found in the
neoplastic tissue after being injected in an animal with a brain
tumor (55), and this fact was
completely proven by the research (Fig.
2). The above mentioned finding allows considering the
phenomenon of targeted migration of stem cells to neoplastic focus
to be the most important mechanism of tissue homeostasis regulation
(56).
Aboody et al reported a molecular adhesion
effect (57) which represents the
unique ability of stem cells to follow a cancer cell into the brain
parenchyma on its cytokine trail, reach the neoplastic cell and
attach itself to it by straddling it like a jockey. This mechanism
opens vast possibilities for the targeted cell therapy of glial and
metastatic brain tumors.
The majority of glioblastoma cells infiltrating the
brain parenchyma during its invasive growth are in interphase.
These cells are insusceptible to traditional cytostatic agents,
while the hypoxic metabolism neutralizes the effects of radiation.
Molecular adhesion allows the direct delivery of ferments
metabolizing inactive substances to cytotoxic matter,
nanoencapsulated medication used for photothermal ablation,
therapeutic genes and oncolytic viruses (58–60).
However, the phenomenon of targeted migration cannot
be viewed only from the standpoint of addressed delivery. Stem
cells will undoubtedly find a cancer cell once being introduced
into a blood vessel, implanted vertically or incorporated into
biopolymer matrices filling the tissue deficiency remaining after
tumor removal. In the latter case, being conjugated with
immunoliposomes (61), they use the
shortest way to transport therapeutic agents to the remaining
cancer cells infiltrating the brain parenchyma, reaching the most
hypoxic zones and creating the maximum concentration of the
therapeutic agents to minimize side-effects of the treatment and
reinforce the cytotoxic effect.
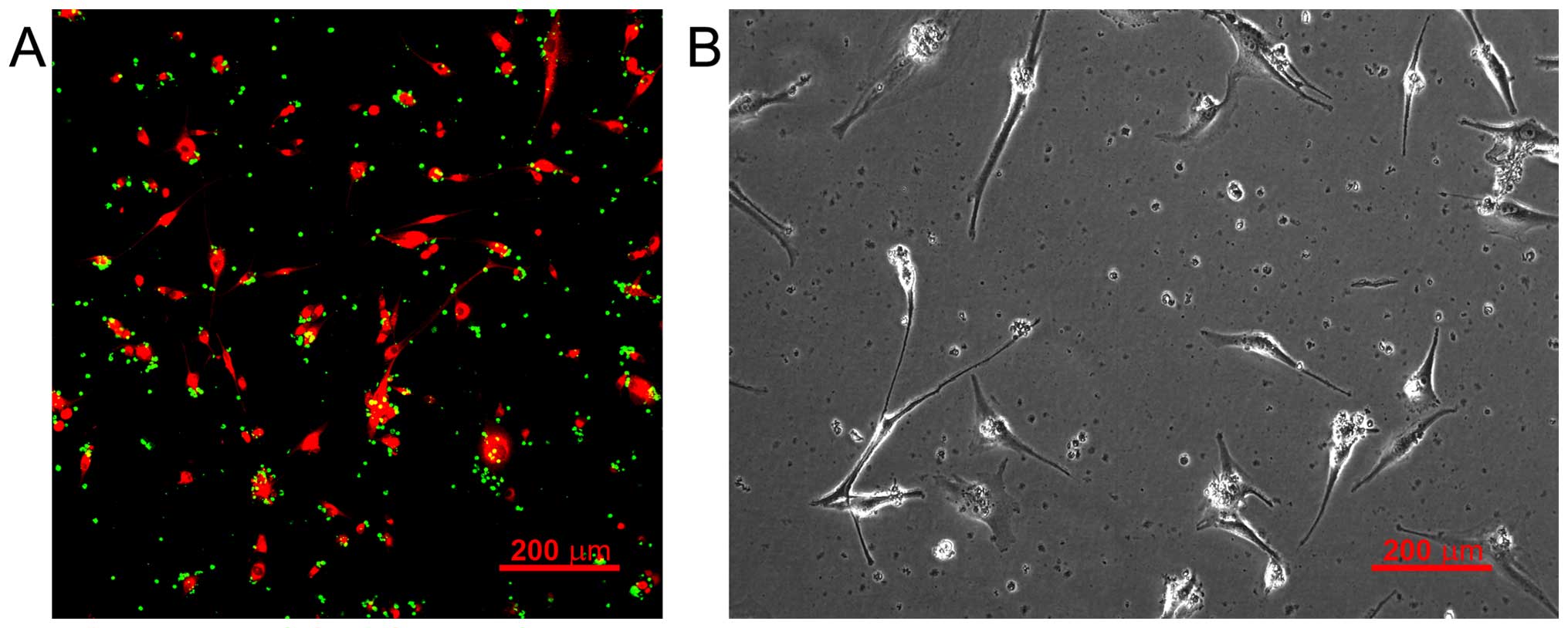
Upon reaching the neoplasia zone, stem cells become
attached to cancer cells and interact with them (Fig. 3). Mechanisms of intercellular
communication include exchange of specific regulatory proteins,
activation of cell surface receptors, bystander effect, stimulation
of inflammatory response, arrest of G1 phase cancer cell cycle and
reprogramming of tumor cells via cytoplasmic transcription factors
during the fusion of stem and cancer cells (62).
Accumulation of healthy stem cells in the neoplasm
zone inhibits tumor development which has been experimentally
proven many times. If the ratio of stem to cancer cells is 2:1, the
proliferation speed of the latter drops significantly, while when
there are a large number of stem cells, cancer cell proliferation
stops (63–65). In the case of combined cultivation
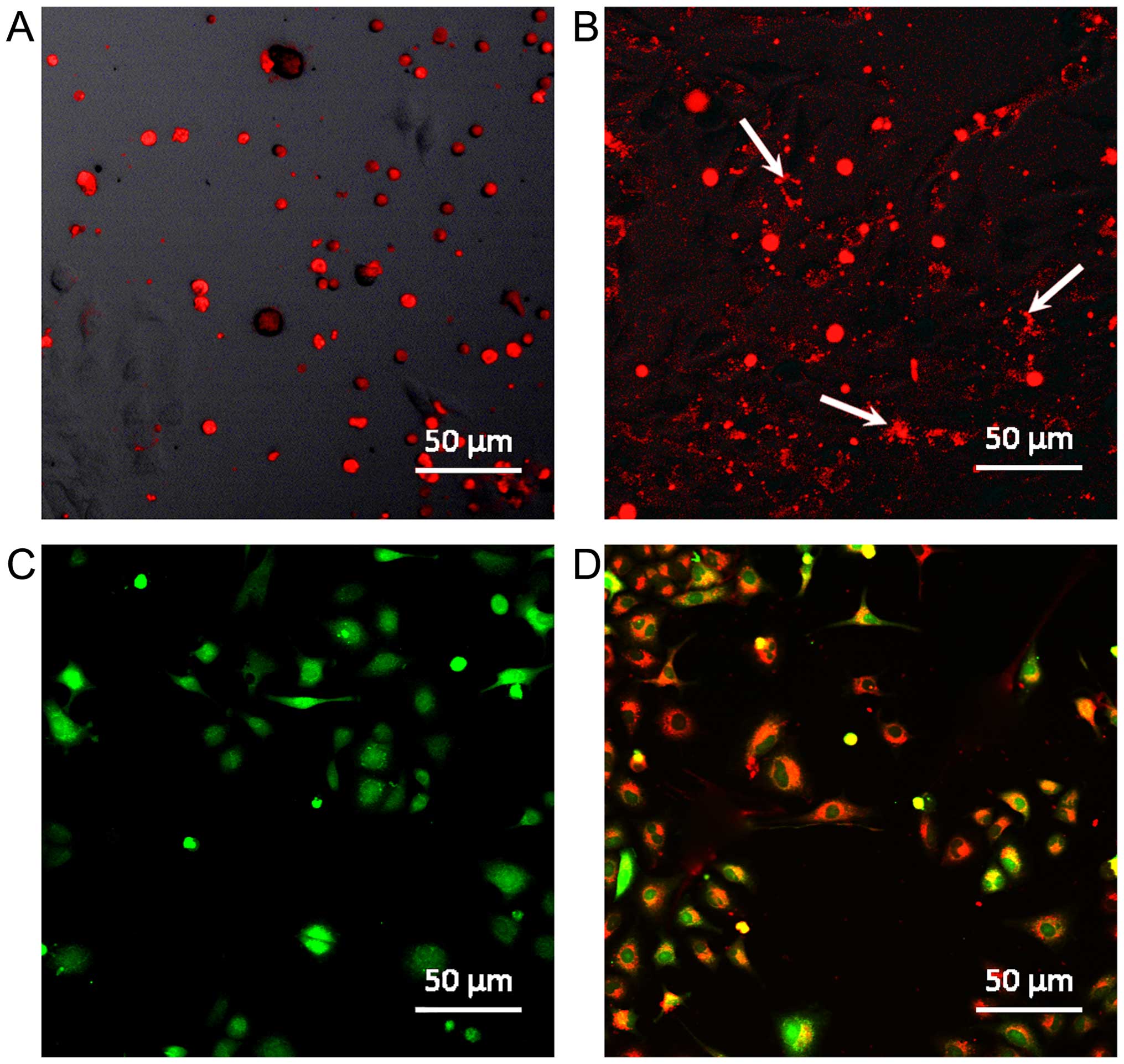
of hematopoietic stem cells (HSCs) CD34+CD45+
with C6 glioma there is an active exchange of molecules that is
proven by the fact of cancer cell accumulation of pigments
connected with HSC cytoplasmic proteins. This phenomenon is also
observed in the cultivation of lung cancer cells and breast cancer
cells with HSCs (Fig. 4A–D). The
mechanisms of this phenomenon probably include creation of
structural and functional syncytium and cell fusion effect between
interacting cells (66).
Cell fusion is a fundamental biological process.
This mechanism enables the exchange of both regulation and
defective proteins (67,68). Serious disruptions of DNA
methylation significantly destabilize a genome, while microRNA
exchange resulting from mixing cytoplasm suppresses onco-suppressor
gene expression (for instance, miR-34/deactivates c-Met
gene, miR-146a/gene deactivates Notch gene, miR-7/gene
EGFR, miR-128/gene Bmi1) and stimulates oncogene
activity (69–71). This mechanism needs to be studied.
Yet, it is already clear that the stem cell proteome profile
modification related to regulatory protein production allows us to
create a cell biological drug for inhibiting the activity of
glioblastoma cells located deep in the brain parenchyma and some
metastatic tumors.
At important inducer of cancer cell elimination is
tumor necrosis factor (TNF). This family includes 19 proteins that
have a function of ligand for 29 receptors. Receptors of the
superfamily interact with adaptor proteins including TRADD, TRAF,
RIP that define a cell response to cytokine activation, for
instance, stimulation of apoptosis or inflammation. Currently,
there are methods of addressed delivery of TRAIL, S-TRAIL proteins
right into the cancer cells via using stem cell potential (72). There are methods of HSC modification
by incorporating therapeutic genes into them to induce TNF
production (73,74). The important feature is the ability
of TNF to stimulate the production of granulocyte
colony-stimulating factor that mobilizes stem cells from their
repository in the bone marrow, thus stimulating their migration to
the neoplasia zone (75).
The 'Bystander' effect is a special case of
inductive interaction between cells. The main principle of this
effect involves triggering key functions in one of the interacting
cells. This phenomenon is frequently observed in radiology when the
death of a cell having received a lethal radiation dose is followed
by suspended death of the tumor cells closest to it. Radiation
stimulates production of IL-1, IL-2, IL-6, IL-8, TNFα and TGFβ by
cells. The mechanism opens the possibilities of creating cellular
systems with induced apoptosis that can transfer destructive
influence to the cancer cell (76–78).
Thus, the advances in cellular biology and
post-genome technologies even now allow the creation of totally new
methods of glioblastoma treatment. However, CSCs have very strong
repair mechanisms. It is highly probable that selective treatment
slows down tumor development, but later CSCs will react creating
new resistant cell clones that will provoke a relapse. This
situation requires the development of specific methods for managing
the key functions of CSCs.
5. Cell and post-genome technologies in the
complex treatment of glioblastoma multiforme
A disadvantage of the existing strategies for the
treatment of malignant tumors is having an ideal goal of destroying
all neoplastic cells. This is unrealistic due to the ideal nature,
and it is particularly true in the case of non-encapsulated tumors
of the nervous system. Glioblastoma is an autonomous dynamically
changing structure with the ability to adapt in response to growing
intensity of treatment procedures. One reaction for adapting to
drugs and radiation is an increase in the number of CSCs.
Neurosurgery is able to solve a number of problems
at the same time. It reduces intracranial hypertension, eliminates
a large part of cancer cells, including their clones, and this
activates the proliferation processes of the remaining pools and
makes them vulnerable to medication. In addition, surgery also
breaks the integrity of the hematoencephalic barrier that increases
the spectrum of applicable medication (80).
Further chemotherapy mostly is based on a standard
procedure that has been previously described. A key breakthrough is
cytokine therapy designed to disintegrate the interaction between
tumor cells and the local microenvironment; excessive expression of
epidermal growth factor receptor (EGFR) is characteristic of 40% of
GBM. In order to influence this target, drugs such as gefinitibum
(Iressa®), erlotinib (Tarceva®) and lapatinib
(Tyverb®) have been suggested. Platelet-derived growth
factor (PDGF) is one of the major ligands in GBM biology. To
suppress PDGF, tandutinib (MLN518) and sunitinib
(Sutent®) are suggested. These drugs directly suppress
tyrosine kinase signaling and block proliferation, migration and
anti-apoptosis mechanisms in cancer cells (81–83).
Hypoxia is a key parameter of GBM biology, and this
offers certain clinical potential for the use of such drugs as
inhibitors of the signaling pathway PI3K/AKT/MTOR that include
temsorolimus (Torisel®) and everolimus
(Afinitor®). These drugs reduce HIF and VEGF levels in
cancer tissues and helps to suppress the development of blood
vessels and reduces the ability of CSCs to adapt in a hypoxic media
(84–86).
One widespread approach is the use of immunotherapy,
and it is not unreasonable. Since a person normally has up to
500,000 (105) cancer cells, a patient with pre-cancer
has ~1 billion (<109) of them, while a patient with a
malignant tumor has more than a billion cells. The healthy body has
these cells under control via a natural regulatory system. Surgery
helps to reduce the amount of cancer cells up to 109,
while radiation, radio and chemotherapy allow the reduction of
cancer cells from 109 to 107.
Sanogenetic abilities of our immune system may be
activated only if the number of cancer cells does not exceed
105. A gap between sanogenetic abilities of the immune
system and therapeutic possibilities of classic antitumor treatment
is ~2 orders (from 105 to 107 cancer cells).
Researchers have high hopes for immune therapy in order to
eliminate those remaining 102 cancer cells, and in some
cases it may prolong the life of neuro-oncological patients.
However, using even the most modern methods of
cytoreductive therapy without targeting CSCs will not have the
expected outcome. If we treat the 'stem' nature of cancer cells as
a reason of their survival, than there is a reasonable assumption
that a tumor will increase the number of CSCs in order to fight the
treatment. Radiation induces the selection of cell clones that are
more resistant to hypoxia. Low partial pressure of oxygen prevents
the creation of free radicals that neutralize the effect of
radiotherapy. With partial pressure of oxygen being between 0.01
and 5%, cancer cells experience cell cycle arrest of the G0/1/Gs
phase preserving their invasive potential and making them resistant
to cytostatic chemotherapy. In their turn, a large number of CSCs
initiates invasive growth mechanisms very rapidly after
normalization of external conditions (87–89).
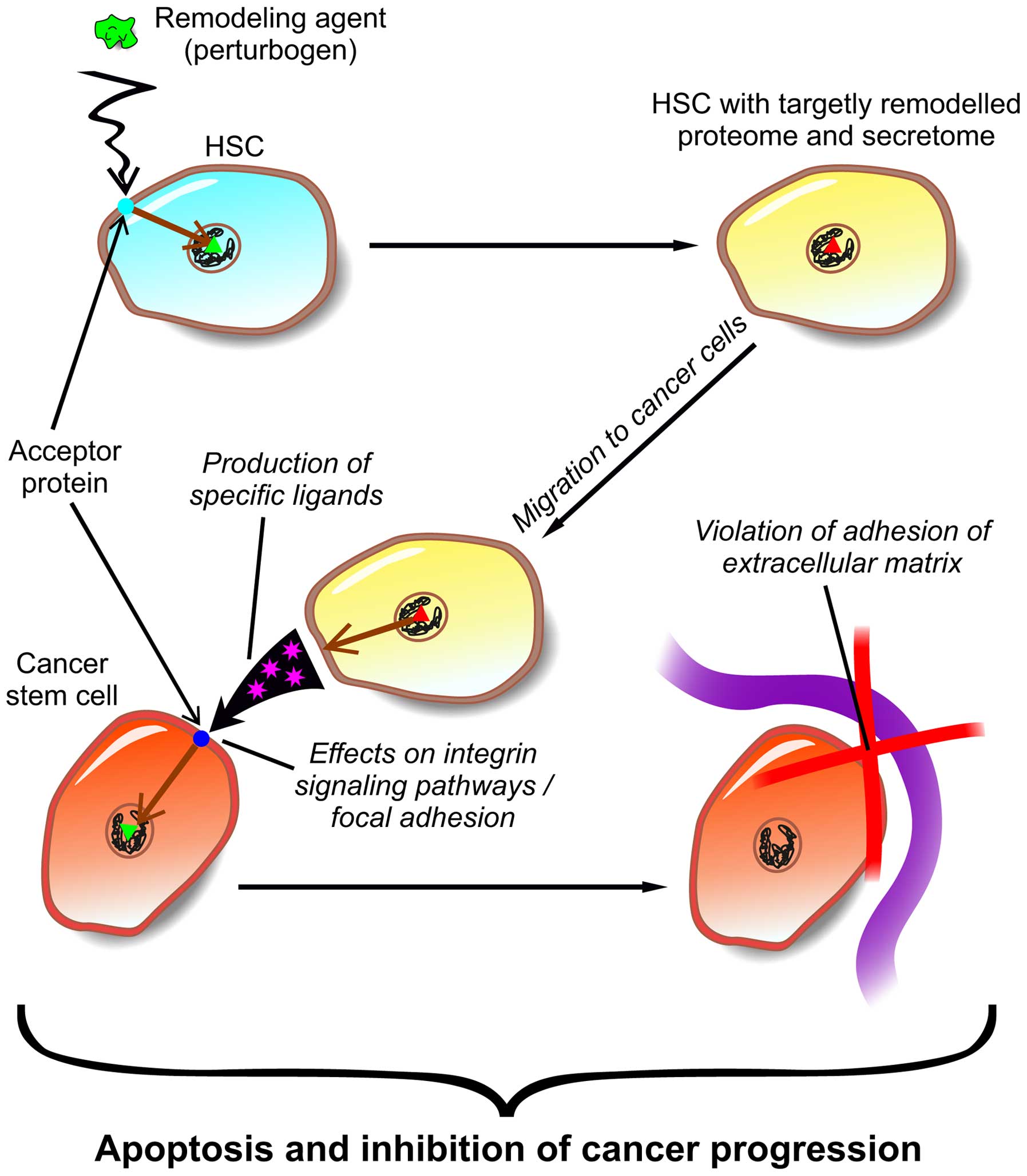
In this respect, the use of cell and post-genome
technologies may include specific mechanisms of interaction between
CSCs and the cell matrix. The result of NSC and CSC comparative
proteome mapping of glioblastoma U87 cells concluded that a pathway
of focal adhesion in CSCs in glioblastoma was not affected during
neoplastic transformation (31) and
can be used for treatment targeted at disintegration of the
CSC-extracellular matrix bond (Fig.
5).
A signaling pathway of focal adhesion starts with
transferring a signal to the cell by growth factors (EGF, FIGF,
HGF, IGF1, PDGFA, PDGFB, PGF, PDGFC, PDGFD, VEGFA, VEGFB and VEGFC)
and proteins of the extracellular matrix (lamenines, fibronectin
FN1, chondroadherin CHAD, cartilage oligomeric matrix protein COMP,
integrin-binding sialoprotein IBSP, tenascins TNC, TNN, TNR, TNXB,
reelin released by phosphoprotein 1, thrombospondins THBS1, THBS2,
THBS3 and THBS4, vitronectine and von Willebrand factor). The
external side of the plasmatic membrane is a site for transferring
signals via integrins, caveolins and tyrosine kinase receptors
(EGFR, ERBB2, FLT1, FLT4, IGF1R, KDR, MET, PDGFRA and PDGFRB). This
set of protein targets helps to influence the CSC adhesion
processes in a direct way. Correspondingly, a whole set of protein
targets is obtained allowing to interfere with the adhesion
processes of CSCs (31,90,91).
Modification of the transcriptomic profile of
autologous stem cells in terms of IL-23 and a series of ligands
activating acceptor proteins of the cell surface in signaling
pathway of focal adhesion allows targeted influence on the
interaction of glioblastoma CSCs and the intercellular matrix that
limits the proliferation abilities of this type of cells and
induces apoptosis (93). Cell-based products can be injected into
the blood vessels of patients or incorporated into biopolymer
collagen matrix implanted into the cavity of the removed tumor. In
the latter case, there is a possibility for the use of a 'molecular
trace' and reach tumor cells having migrated to the brain
parenchyma by the shortest path. In addition, biopolymer matrix
significantly changes the regeneration process by creating the
least favorable medium for CSC reduplication.
Thus, the success of cell and post-genome
technologies already allows us to start using biological drugs for
the treatment of CSCs of glioblastoma. The question concerning the
application of these technologies in the complex structure of GBM
cannot be answered even in terms of theory. A large amount of CSCs
before treatment is an unfavorable factor that requires using
disintegration methods for systems including CSC-extracellular
matrix and CSC-local microenvironment as a first stage of therapy
which is mostly used for inoperable tumors. The obvious choice of
method here could be the complex use of proteomics-based biological
drugs together with cytokine therapy.
Surgery is an obligatory stage of GBM treatment. The
extracted tissue is subjected to immunocytochemical and
morphological analysis, and mapping of the patient genetic and
epigenetic defects. Chemotherapy effectiveness can be significantly
increased by prescribing medication taking into consideration the
individual tumor characteristics of the patients.
CSC extraction is a strategically important stage of
treatment. Comparative proteome mapping, profiling and
bioinformation on CSCs and somatic stem cells of a patient are
recommended for all cases while choosing proteome-personalized cell
therapy. This allows the identification of acceptor proteins in the
CSC surface that are connected with intracellular pathways of
signal transduction unaffected by neoplastic transformation and
suitable for managing specific functions of CSCs.
6. Conclusion
The theory of CSCs is one of the latest advances
identified by experimental oncology. As a crossing point of all
modern views on carcinogenesis, the CSC theory explains the
processes of dynamic modification and adaptation of tumor tissue in
the case of using conventional schemes of anticancer therapy. It
perfectly explains the reasons for medicinal and radiation
resistance of GBM, disease relapse and generally low effectiveness
of modern GBM treatment. This is why disregard for the role and
significance of targeted dynamic regulation of cancer cells and the
key functions of CSCs in the modern system of GBM treatment is a
big oversight. Complex GBM treatment should be complemented by
biomedical proteome-based cell drugs intended for managing the key
functions of CSCs. These methods should be used along with modern
targeted pharmaceuticals that allow management of the functions of
CSCs and improvement of the survival rates of neuro-oncologic
patients.
Acknowledgments
The present study was funded by a grant from the
Russian Scientific Foundation (project 14-15-00084).
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Smith RA, Manassaram-Baptiste D, Brooks D,
Doroshenk M, Fedewa S, Saslow D, Brawley OW and Wender R: Cancer
screening in the United States, 2015: A review of current American
Cancer Society guidelines and current issues in cancer screening.
CA Cancer J Clin. 65:30–54. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Louis DN, Perry A, Burger P, Ellison DW,
Reifenberger G, von D eimling A, Aldape K, Brat D, Collins VP,
Eberhart C, et al: International Society Of Neuropathology -
Haarlem: International Society Of Neuropathology - Haarlem
consensus guidelines for nervous system tumor classification and
grading. Brain Phatol. 24:429–435. 2014. View Article : Google Scholar
|
|
5
|
Brodbelt A, Greenberg D, Winters T,
Williams M, Vernon S and Collins VP; (UK) National Cancer
Information Network Brain Tumour Group: Glioblastoma in England:
2007–2011. Eur J Cancer. 51:533–542. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Omuro A and DeAngelis LM: Glioblastoma and
other malignant gliomas: A clinical review. JAMA. 310:1842–1850.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Konovalov AN, Potapov AA, Loshakov VA,
Oliushin VE, Kornienko VN, Iartsev VV, Pronin IN, Korshunov AG,
Golanov AV, Kobiakov GL, et al: Standards, guidelines, and options
in the treatment of glial tumors of the brain in adults. Zh Vopr
Neirokhir Im NN Burdenko. 2:3–11. 2006.In Russian.
|
|
8
|
Xhumari A, Rroji A, Enesi E, Bushati T,
Sallabanda Diaz K and Petrela M: Glioblastoma after AVM
radiosurgery. Case report and review of the literature. Acta
Neurochir. 157:889–895. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Greto D, Livi L, Bonomo P, Masi L, Detti
B, Meattini I, Mangoni M, Doro R, Favuzza V, Cipressi S, et al:
Cyberknife stereotactic radiosurgery for the re-irradiation of
brain lesions: A single-centre experience. Radiol Med. 119:721–726.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tsao MN, Mehta MP, Whelan TJ, Morris DE,
Hayman JA, Flickinger JC, Mills M, Rogers CL and Souhami L: The
American Society for Therapeutic Radiology and Oncology (ASTRO)
evidence-based review of the role of radiosurgery for malignant
glioma. Int J Radiat Oncol Biol Phys. 63:47–55. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Freund D, Zhang R, Sanders M and Newhauser
W: Predictive risk of radiation induced cerebral necrosis in
pediatric brain cancer patients after VMAT versus proton therapy.
Cancers. 7:617–630. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kamiya-Matsuoka C and Gilbert MR: Treating
recurrent glioblastoma: An update. CNS Oncol. 4:91–104. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dincoglan F, Beyzadeoglu M, Sager O,
Demiral S, Gamsiz H, Uysal B, Ebruli C, Akin M, Oysul K, Sirin S,
et al: Management of patients with recurrent glioblastoma using
hypofractionated stereotactic radiotherapy. Tumori. 101:179–184.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kickingereder P, Hamisch C, Suchorska B,
Galldiks N, Visser-Vandewalle V, Goldbrunner R, Kocher M, Treuer H,
Voges J and Ruge MI: Low-dose rate stereotactic iodine-125
brachytherapy for the treatment of inoperable primary and recurrent
glioblastoma: Single-center experience with 201 cases. J
Neurooncol. 120:615–623. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang B, Tanaka K, Ji B, Ono M, Fang Y,
Ninomiya Y, Maruyama K, Izumi-Nakajima N, Begum N, Higuchi M, et
al: Total body 100-mGy X-irradiation does not induce Alzheimer's
disease-like pathogenesis or memory impairment in mice. J Radiat
Res. 55:84–96. 2014. View Article : Google Scholar :
|
|
16
|
Stupp R, Mason WP, van den Bent MJ, Weller
M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn
U, et al European Organisation for Research and Treatment of Cancer
Brain Tumor and Radiotherapy Groups: National Cancer Institute of
Canada Clinical Trials Group: Radiotherapy plus concomitant and
adjuvant temozolomide for glioblastoma. N Engl J Med. 352:987–996.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Taal W, Bromberg JE and van den Bent MJ:
Chemotherapy in glioma. CNS Oncol. 4:179–192. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zorzan M, Giordan E, Redaelli M, Caretta A
and Mucignat-Caretta C: Molecular targets in glioblastoma. Future
Oncol. 11:1407–1420. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Deutsch MB, Panageas KS, Lassman AB and
Deangelis LM: Steroid management in newly diagnosed glioblastoma. J
Neurooncol. 113:111–116. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ghiaseddin A and Peters KB: Use of
bevacizumab in recurrent glioblastoma. CNS Oncol. 4:157–169. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bryukhovetskyi IS, Bryukhovetskyi AS,
Kumeiko VV, Mischenko PV and Khotimchenko YS: Stem cells in
carcinogenesis of glioblastoma multiforme. Cell Transplant Tissue
Eng. 8:13–19. 2013.
|
|
22
|
Chen D: Tumor formation and drug
resistance properties of human glioblastoma side population cells.
Mol Med Rep. 11:4309–4314. 2015.PubMed/NCBI
|
|
23
|
Liu Y, Zhang X, Liu J, Hou G, Zhang S and
Zhang J: Everolimus in combination with letrozole inhibit human
breast cancer MCF-7/Aro stem cells via PI3K/mTOR pathway: An
experimental study. Tumour Biol. 35:1275–1286. 2014. View Article : Google Scholar
|
|
24
|
Li C, Lee CJ and Simeone DM:
Identification of human pancreatic cancer stem cells. Methods Mol
Biol. 568:161–173. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Duesberg P, Mandrioli D, McCormack A and
Nicholson JM: Is carcinogenesis a form of speciation? Cell Cycle.
10:2100–2114. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Altman J: Are new neurons formed in the
brains of adult mammals? Science. 135:1127–1128. 1962. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lepousez G, Nissant A and Lledo PM: Adult
neurogenesis and the future of the rejuvenating brain circuits.
Neuron. 86:387–401. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Doetsch F, Caillé I, Lim DA,
García-Verdugo JM and Alvarez-Buylla A: Subventricular zone
astrocytes are neural stem cells in the adult mammalian brain.
Cell. 97:703–716. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Maldonado-Soto AR, Oakley DH, Wichterle H,
Stein J, Doetsch FK and Henderson CE: Stem cells in the nervous
system. Am J Phys Med Rehabil. 93(Suppl 3): S132–S144. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang S and Cui W: Sox2, a key factor in
the regulation of pluripotency and neural differentiation. World J
Stem Cells. 6:305–311. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bryukhovetskiy A, Shevchenko V, Kovalev S,
Chekhonin V, Baklaushev V, Bryukhovetskiy I and Zhukova M: To the
novel paradigm of proteome-based cell therapy of tumors: Through
comparative proteome mapping of tumor stem cells and tissuespecific
stem cells of humans. Cell Transplant. 23(Suppl 1): S151–S170.
2014. View Article : Google Scholar
|
|
32
|
Vescovi AL, Galli R and Reynolds BA: Brain
tumour stem cells. Nat Rev Cancer. 6:425–436. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Amendola D, Nardella M, Guglielmi L,
Cerquetti L, Carico E, Alesi V, Porru M, Leonetti C, Bearzi C,
Rizzi R, et al: Human placenta-derived neurospheres are susceptible
to transformation after extensive in vitro expansion. Stem Cell Res
Ther. 5(55)2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wu W, He Q, Li X, Zhang X, Lu A, Ge R,
Zhen H, Chang AE, Li Q and Shen L: Long-term cultured human neural
stem cells undergo spontaneous transformation to tumor-initiating
cells. Int J Biol Sci. 7:892–901. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ilkanizadeh S, Lau J, Huang M, Foster DJ,
Wong R, Frantz A, Wang S, Weiss WA and Persson AI: Glial
progenitors as targets for transformation in glioma. Adv Cancer
Res. 121:1–65. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Schichor C, Albrecht V, Korte B, Buchner
A, Riesenberg R, Mysliwietz J, Paron I, Motaln H, Turnšek TL,
Jürchott K, et al: Mesenchymal stem cells and glioma cells form a
structural as well as a functional syncytium in vitro. Exp Neurol.
234:208–219. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wurmser AE and Gage FH: Stem cells: Cell
fusion causes confusion. Nature. 416:485–487. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Friedmann-Morvinski D: Glioblastoma
heterogeneity and cancer cell plasticity. Crit Rev Oncog.
19:327–336. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rahman M, Reyner K, Deleyrolle L, Millette
S, Azari H, Day BW, Stringer BW, Boyd AW, Johns TG, Blot V, et al:
Neurosphere and adherent culture conditions are equivalent for
malignant glioma stem cell lines. Anat Cell Biol. 48:25–35. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Fine HA: New strategies in glioblastoma:
Exploiting the new biology. Clin Cancer Res. 21:1984–1988. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Pavon LF, Marti LC, Sibov TT, Malheiros
SM, Brandt RA, Cavalheiro S and Gamarra LF: In vitro analysis of
neurospheres derived from glioblastoma primary culture: A novel
methodology paradigm. Front Neurol. 4(214)2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Golebiewska A, Bougnaud S, Stieber D,
Brons NH, Vallar L, Hertel F, Klink B, Schröck E, Bjerkvig R and
Niclou SP: Side population in human glioblastoma is non-tumorigenic
and characterizes brain endothelial cells. Brain. 136:1462–1475.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Shen G, Shen F, Shi Z, Liu W, Hu W, Zheng
X, Wen L and Yang X: Identification of cancer stem-like cells in
the C6 glioma cell line and the limitation of current
identification methods. In Vitro Cell Dev Biol Anim. 44:280–289.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Neradil J and Veselska R: Nestin as a
marker of cancer stem cells. Cancer Sci. 106:803–811. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Piccirillo SG, Binda E, Fiocco R, Vescovi
AL and Shah K: Brain cancer stem cells. J Mol Med Berl.
87:1087–1095. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chen R, Nishimura MC, Bumbaca SM,
Kharbanda S, Forrest WF, Kasman IM, Greve JM, Soriano RH, Gilmour
LL, Rivers CS, et al: A hierarchy of self-renewing tumor-initiating
cell types in glioblastoma. Cancer Cell. 17:362–375. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Barrett LE, Granot Z, Coker C, Iavarone A,
Hambardzumyan D, Holland EC, Nam HS and Benezra R: Self-renewal
does not predict tumor growth potential in mouse models of
high-grade glioma. Cancer Cell. 21:11–24. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Huang Z, Cheng L, Guryanova OA, Wu Q and
Bao S: Cancer stem cells in glioblastoma - molecular signaling and
therapeutic targeting. Protein Cell. 1:638–655. 2010. View Article : Google Scholar
|
|
49
|
Bryukhovetskii IS, Bryukhovetskii AS and
Khotimchenko YS: New biomolecular approaches to the treatment of
glioblastoma multiforme. Bull Exp Biol Med. 158:794–799. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Bryukhovetskiy IS, Bryukhovetskiy AS,
Mischenko PV and Khotimchenko YS: The role of systemic migration
and homing mechanisms of stem cells in the development of malignant
tumors of the central nervous system and the development of new
cancer therapies. Russian Biotherapeutic J. 4:3–12. 2013.In
Russian.
|
|
51
|
Bryukhovetskiy IS, Mischenko PV, Tolok EV,
Zaitcev SV, Khotimchenko YS and Bryukhovetskiy AS: Directional
migration of adult hematopoeitic progenitors to C6 glioma in vitro.
Oncol Lett. 9:1839–1844. 2015.PubMed/NCBI
|
|
52
|
Bryukhovetskiy IS, Bryukhovetskiy AS,
Mischenko PV, et al: Migration of human hematopoietic stem cells to
cells of glioblastoma line U87 in vitro. Russ Biotherapeutic J.
4:31–36. 2014.
|
|
53
|
Kang R, Zhang Q, Zeh HJ III, Lotze MT and
Tang D: HMGB1 in cancer: Good, bad, or both? Clin Cancer Res.
19:4046–4057. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Bordji K, Grandval A, Cuhna-Alves L,
Lechapt-Zalcman E and Bernaudin M: Hypoxia-inducible factor-2α
(HIF-2α), but not HIF-1α, is essential for hypoxic induction of
class III β-tubulin expression in human glioblastoma cells. FEBS J.
281:5220–5236. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Moore XL, Lu J, Sun L, Zhu CJ, Tan P and
Wong MC: Endothelial progenitor cells' 'homing' specificity to
brain tumors. Gene Ther. 11:811–818. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Rolando C, Parolisi R, Boda E, Schwab ME,
Rossi F and Buffo A: Distinct roles of Nogo-a and Nogo receptor 1
in the homeostatic regulation of adult neural stem cell function
and neuroblast migration. J Neurosci. 32:17788–17799. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Aboody KS, Brown A, Rainov NG, Bower KA,
Liu S, Yang W, Small JE, Herrlinger U, Ourednik V, Black PM, et al:
Neural stem cells display extensive tropism for pathology in adult
brain: Evidence from intracranial gliomas. Proc Natl Acad Sci USA.
97:12846–12851. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Aboody KS, Najbauer J, Metz MZ, D'Apuzzo
M, Gutova M, Annala AJ, Synold TW, Couture LA, Blanchard S, Moats
RA, et al: Neural stem cell-mediated enzyme/prodrug therapy for
glioma: Preclinical studies. Sci Transl Med.
5:184ra592013.PubMed/NCBI
|
|
59
|
Schnarr K, Mooney R, Weng Y, Zhao D,
Garcia E, Armstrong B, Annala AJ, Kim SU, Aboody KS and Berlin JM:
Gold nanoparticle-loaded neural stem cells for photothermal
ablation of cancer. Adv Healthc Mater. 2:976–982. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Ahmed AU, Thaci B, Alexiades NG, Han Y,
Qian S, Liu F, Balyasnikova IV, Ulasov IY, Aboody KS and Lesniak
MS: Neural stem cell-based cell carriers enhance therapeutic
efficacy of an oncolytic adenovirus in an orthotopic mouse model of
human glioblastoma. Mol Ther. 19:1714–1726. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Chekhonin VI, Bryukhovetskiy AS, Semenova
AB, Ukhova OV and Pavlov KA: The antitumor agent based
immunoliposomalnoy biological construction, its production method
and vector delivery to the central nervous system with the
malignancy. RF Patent 2336901. Filed August 10, 2007; issued
October 27, 2008.
|
|
62
|
Intercellular interactions. 2nd edition.
revised and supplemented. Meditsina Publishers; Moscow: pp.
p2882003
|
|
63
|
Yang AP, Tang LT and Chen JM: Inhibitory
effect of bone marrow mesenchymal stem cells on lymphoma cell
proliferation. Zhongguo Shi Yan Xue Ye Xue Za Zhi. 22:1610–1615.
2014.In Chinese. PubMed/NCBI
|
|
64
|
Wang M, Cai J, Huang F, Zhu M, Zhang Q,
Yang T, Zhang X, Qian H and Xu W: Pre-treatment of human umbilical
cordderived mesenchymal stem cells with interleukin-6 abolishes
their growth-promoting effect on gastric cancer cells. Int J Mol
Med. 35:367–375. 2015.
|
|
65
|
Ahn JO, Chae JS, Coh YR, Jung WS, Lee HW,
Shin IS, Kang SK and Youn HY: Human adipose tissue-derived
mesenchymal stem cells inhibit T-cell lymphoma growth in vitro and
in vivo. Anticancer Res. 34:4839–4847. 2014.PubMed/NCBI
|
|
66
|
Bryukhovetskiy IS, Mischenko PV, Tolok EV,
et al: Interaction of hematopoietic stem cells and tumor cells in
vitro. Pa Med J. 58:31–37. 2014.In Russian.
|
|
67
|
He X, Li B, Shao Y, Zhao N, Hsu Y, Zhang Z
and Zhu L: Cell fusion between gastric epithelial cells and
mesenchymal stem cells results in epithelial-to-mesenchymal
transition and malignant transformation. BMC Cancer. 15(24)2015.
View Article : Google Scholar
|
|
68
|
Sumi S and Yanai G: Fusion of mesenchymal
stem cells and islet cells for cell therapy. Methods Mol Biol.
1313:107–113. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Aguilar PS, Baylies MK, Fleissner A,
Helming L, Inoue N, Podbilewicz B, Wang H and Wong M: Genetic basis
of cell-cell fusion mechanisms. Trends Genet. 29:427–437. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Willkomm L and Bloch W: State of the art
in cell-cell fusion. Methods Mol Biol. 1313:1–19. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Fatima A, Hescheler J and Šaric T:
Chromosome tracking in fused cells by single nucleotide
polymorphisms. Methods Mol Biol. 1313:95–106. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Bagci-Onder T, Du W, Figueiredo JL,
Martinez-Quintanilla J and Shah K: Targeting breast to brain
metastatic tumours with death receptor ligand expressing
therapeutic stem cells. Brain. 138:1710–1721. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Ezzelarab M, Ezzelarab C, Wilhite T, Kumar
G, Hara H, Ayares D and Cooper DK: Genetically-modified pig
mesenchymal stromal cells: Xenoantigenicity and effect on human
T-cell xenoresponses. Xenotransplantation. 18:183–195. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
He HL, Liu L, Chen QH, Cai SX, Han JB, Hu
SL, Chun P, Yang Y, Guo FM, Huang YZ, et al: MSCs modified with
ACE2 restore endothelial function following LPS challenge by
inhibiting the activation of RAS. J Cell Physiol. 230:691–701.
2015. View Article : Google Scholar
|
|
75
|
Sainathan SK, Hanna EM, Gong Q, Bishnupuri
KS, Luo Q, Colonna M, White FV, Croze E, Houchen C, Anant S, et al:
Granulocyte macrophage colony-stimulating factor ameliorates
DSS-induced experimental colitis. Inflamm Bowel Dis. 14:88–99.
2008. View Article : Google Scholar
|
|
76
|
Najafi M, Fardid R, Hadadi G and Fardid M:
The mechanisms of radiation-induced bystander effect. J Biomed Phys
Eng. 4:163–172. 2014.
|
|
77
|
Marín A, Martín M, Liñán O, Alvarenga F,
López M, Fernández L, Büchser D and Cerezo L: Bystander effects and
radiotherapy. Rep Pract Oncol Radiother. 20:12–21. 2015. View Article : Google Scholar
|
|
78
|
Xiao L, Liu W, Li J, Xie Y, He M, Fu J,
Jin W and Shao C: Irradiated U937 cells trigger inflammatory
bystander responses in human umbilical vein endothelial cells
through the p38 pathway. Radiat Res. 182:111–121. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Barani IJ and Larson DA: Radiation therapy
of glioblastoma. Cancer Treat Res. 163:49–73. 2015. View Article : Google Scholar
|
|
80
|
Dolecek TA, Propp JM, Stroup NE and
Kruchko C: CBTRUS statistical report: Primary brain and central
nervous system tumors diagnosed in United States in 2005–2009.
Neuro Oncol. 14(Suppl 5): v1–v49. 2012. View Article : Google Scholar :
|
|
81
|
Friedmann-Morvinski D: Glioblastoma
heterogeneity and cancer cell plasticity. Crit Rev Oncog.
19:327–336. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Labussière M, Boisselier B, Mokhtari K, Di
Stefano AL, Rahimian A, Rossetto M, Ciccarino P, Saulnier O,
Paterra R, Marie Y, et al: Combined analysis of TERT, EGFR, and IDH
status defines distinct prognostic glioblastoma classes. Neurology.
83:1200–1206. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Lee JK, Joo KM, Lee J, Yoon Y and Nam DH:
Targeting the epithelial to mesenchymal transition in glioblastoma:
The emerging role of MET signaling. Onco Targets Ther. 7:1933–1944.
2014.PubMed/NCBI
|
|
84
|
Lassen U, Sorensen M, Gaziel TB,
Hasselbalch B and Poulsen HS: Phase II study of bevacizumab and
temsirolimus combination therapy for recurrent glioblastoma
multiforme. Anticancer Res. 33:1657–1660. 2013.PubMed/NCBI
|
|
85
|
Chinnaiyan P, Won M, Wen PY, Rojiani AM,
Wendland M, Dipetrillo TA, Corn BW and Mehta MP: RTOG 0913: A phase
1 study of daily everolimus (RAD001) in combination with radiation
therapy and temozolomide in patients with newly diagnosed
glioblastoma. Int J Radiat Oncol Biol Phys. 86:880–884. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Venkatesh HS, Chaumeil MM, Ward CS,
Haas-Kogan DA, James CD and Ronen SM: Reduced phosphocholine and
hyperpolarized lactate provide magnetic resonance biomarkers of
PI3K/Akt/mTOR inhibition in glioblastoma. Neuro Oncol. 14:315–325.
2012. View Article : Google Scholar :
|
|
87
|
Gardner LB, Li Q, Park MS, Flanagan WM,
Semenza GL and Dang CV: Hypoxia inhibits G1/S transition
through regulation of p27 expression. J Biol Chem. 276:7919–7926.
2001. View Article : Google Scholar
|
|
88
|
Goda N, Ryan HE, Khadivi B, McNulty W and
Rickert RC: Hypoxia-inducible factor 1alpha is essential for cell
cycle arrest during hypoxia. Mol Cell Biol. 23:359–369. 2003.
View Article : Google Scholar :
|
|
89
|
Mjelle R, Hegre SA, Aas PA, Slupphaug G,
Drabløs F, Saetrom P and Krokan HE: Cell cycle regulation of human
DN A repair and chromatin remodeling genes. DNA Repair. 30:53–67.
2015. View Article : Google Scholar
|
|
90
|
Justus CR and Yang LV: GPR4 decreases
B16F10 melanoma cell spreading and regulates focal adhesion
dynamics through the G13/Rho signaling pathway. Exp Cell
Res. 334:100–113. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Xiang L, Chi T, Tang Q, Yang X, Ou M, Chen
X, Yu X, Chen J, Ho RJ, Shao J, et al: A pentacyclic triterpene
natural product, ursolic acid and its prodrug US597 inhibit targets
within cell adhesion pathway and prevent cancer metastasis.
Oncotarget. 6:9295–9312. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Bryukhovetskiy AS, Bryukhovetskiy IS,
Shevchenko VE and Davidov MI: Antitumor individual proteome-based
effective targeted cell preparation method for its preparation and
using of this drug for the treatment of cancer and other
malignancies. RF Patent 2335972. Filed December 24, 2012; issued
October 20, 2014.
|