Introduction
Hepatocellular carcinoma (HCC) is one of the most
common forms of liver cancer (1)
and ranks as the third-leading cause of cancer-related mortality
(2). Although numerous therapeutic
strategies have been employed to treat this fatal disease, the
prognosis of HCC patients remains dismal with a low 5-year survival
rate of ~30% (3,4). The unsatisfactory prognosis of HCC
largely is attributed to the lack of diagnostic biomarkers and
effective therapeutic targets. Therefore, it is of great importance
to elucidate the exact mechanisms of the pathogenesis of HCC, and
subsequently find promising biomarkers and therapeutic targets of
HCC.
B-cell CLL/lymphoma-3 (BCL-3) is an atypical member
of the IκB family (5) and can bind
NF-κB homodimeric complexes of p50 or p52, which switches the
transcriptional properties of the homodimers from a repressive to
an activating state (6). It was
initially identified as a pro-oncogene in cancers of the blood,
bone marrow and lymphatic system (7–9).
Recently, emerging evidence suggests that BCL-3 also plays
promoting roles in the development and progression of various solid
tumors (10). The mRNA and protein
expression levels of BCL-3 have been reported to be overexpressed
in breast (6,11), nasopharyngeal (12), endometrial (13) and colorectal cancer (14). Nuclear BCL-3 expression has been
found to be associated with the poor prognosis of colorectal cancer
patients (14). Functionally, BCL-3
was found to regulate colony formation and cell cycle progression
by regulating ubiquitination-mediated degradation of c-Myc in
colorectal cancer (15). In
addition, together with p50, BCL-3 was found to regulate the
metastasis of renal cell carcinoma in response to antiangiogenic
therapy (16). Therefore, the
functional role of BCL-3 in human cancers seems to be cancer-type
specific. To date, the clinical significance and the exact function
of BCL-3 in HCC are poorly investigated.
In the present study, we found that the expression
level of BCL-3 in human HCC was significantly elevated, and the
increased expression level of BCL-3 was associated with unfavorable
clinical features and the poor prognosis of the HCC patients. Our
in vitro and in vivo studies demonstrated that BCL-3
promoted the tumor growth of HCC by facilitating cell viability,
proliferation and cell cycle progression. Furthermore, we found
that BCL-3 exerted its roles in HCC cells by regulating the
expression of cyclin D1. Our results indicated that BCL-3 is a
promising biomarker of HCC, and can potentially serve as a
therapeutic target of HCC.
Materials and methods
Clinical samples and data
Clinical specimens derived from 90 patients,
including 79 males and 11 females who were diagnosed with primary
HCC, were examined in the present study, after obtaining informed
consent from every patient. All patients underwent surgical
resection at the Department of Hepatobiliary Surgery, The First
Affiliated Hospital of Xi'an Jiaotong University during January
2007 to December 2009. All patients did not receive any
chemotherapy or embolization during the perioperative period. The
clinicopathological data of these 90 patients are presented in
Table I. All protocols were
approved by the Ethics Committee of Xi'an Jiaotong University
according to the Declaration of Helsinki (as revised in Tokyo
2004).
 | Table IClinicopathological correlation of
BCL-3 expression in the HCC cases. |
Table I
Clinicopathological correlation of
BCL-3 expression in the HCC cases.
| Clinicopathological
features | n | BCL-3 expression
| P-value |
|---|
| Positive (n=58) | Negative (n=32) |
|---|
| Age (years) | | | | 0.323 |
| ≤50 | 36 | 21 | 15 | |
| >50 | 54 | 37 | 17 | |
| Gender | | | | 0.692 |
| Male | 79 | 52 | 27 | |
| Female | 11 | 6 | 5 | |
| HBV infection | | | | 0.015a |
| No | 13 | 4 | 9 | |
| Yes | 77 | 54 | 23 | |
| Serum AFP level
(ng/ml) | | | | 0.102 |
| ≤20 | 27 | 14 | 13 | |
| >20 | 63 | 44 | 19 | |
| Tumor size
(cm) | | | | 0.005a |
| ≤5 | 41 | 20 | 21 | |
| >5 | 49 | 38 | 11 | |
| No. of tumor
nodules | | | | 0.183 |
| 1 | 74 | 50 | 24 | |
| ≥2 | 16 | 8 | 8 | |
| Cirrhosis | | | | 0.018a |
| Absent | 21 | 9 | 12 | |
| Present | 69 | 49 | 20 | |
| Venous
infiltration | | | | 0.293 |
| Absent | 43 | 25 | 18 | |
| Present | 47 | 33 | 15 | |
| Edmondson-Steiner
grade | | | | 0.264 |
| I+II | 68 | 46 | 22 | |
| III+IV | 22 | 12 | 10 | |
| TNM tumor
stage | | | | 0.035a |
| I+II | 67 | 39 | 28 | |
| III+IV | 23 | 19 | 4 | |
Cell lines and transfection
The human HCC cell lines, Huh7 and HepG2, were
obtained from the Institute of Biochemistry and Cell Biology,
Chinese Academy of Sciences, Shanghai, China, and were maintained
in Dulbecco's modified Eagle's medium (DMEM) containing 10% fetal
bovine serum (FBS) (both from Gibco, Grand Island, NY, USA) with
100 U/ml penicillin and 100 µg/ml streptomycin (Sigma, St.
Louis, MO, USA). All cells were incubated in a humidified incubator
with 5% CO2 at 37°C.
A specific cyclin D1 siRNA [sense, 5′-r(CCA CAG AUG
UGA AGU UCA U)d(TT)-3′ and antisense, 5′-r(AUG AAC UUC ACA UCU GUG
G)d(TT)-3′); and a non-specific duplex oligonucleotide as a
negative control (sense, 5′-r(UUC UCC GAA CGU GUC ACG U)d(TT)-3′
and antisense, 5′-r(ACG UGA CAC GUU CGG AGA A)d(TT)-3′] were
synthesized by Sangon Biotech, Shanghai, Co., Ltd. (Shanghai,
China). The siRNAs mentioned above were transfected into HCC cells
using Lipofectamine 2000 following the manufacturer's instructions
(Invitrogen, Carlsbad, CA, USA). BCL-3 shRNA plasmid (h)
(sc-29789-SH) and control shRNA plasmid (sc-108060) were purchased
from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Plasmid
transfection reagent (sc-108061; Santa Cruz Biotechnology) was used
for shRNA transfection. Retroviral vector pMMP-BCL-3 was generated
by inserting the cDNA into pMMP. Retrovirus packaging and
transduction were previously described (17).
Immunohistochemical staining
Paraffin-embedded samples were cut into 4-µm
sections, and were deparaffinized in xylene and re-hydrated through
graded ethanol. Antigen retrieval was performed in sodium citrate
buffer for 15 min at 100°C, and then these slices were quenched for
endogenous peroxidase activity in 3% hydrogen peroxide for 10 min.
They were blocked with goat plasma at 37°C for 30 min and incubated
with the BCL-3 antibody (1:100; Santa Cruz Biotechnology) at 4°C
overnight. After incubating with the biotinylated secondary
antibody (ZSGB-Bio, Beijing, China) at room temperature for 2 h,
the sections were incubated with diaminobenzidine and were
counterstained with hematoxylin. The percentage of positive tumor
cells or hepatocytes was graded according to the following
criteria: 0, <10%; 1, 10–30%; 2, 31–50%; and 3, >50%.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
TRIzol reagent (Invitrogen) was employed to collect
the total RNA from clinical specimens following the manufacturer's
protocol. RNA was reverse-transcribed into cDNA using SuperScript
II reverse transcriptase (Invitrogen) and qPCR was conducted with
the TaqMan Universal PCR Master mix (Applied Biosystems, Foster
City, CA, USA). The expression of BCL-3 relative to a housekeeping
gene, GAPDH, was measured using an ABI Prism 7700 Sequence Detector
(Applied Biosystems). The following primers were used: BCL-3
primers, 5′-GAA AAC AAC AGC CTT AGC ATG GT-3′ and 5′-CTG CGG AGT
ACA TTT GCG-3′; and GAPDH primers, 5′-CAA GCT CAT TTC CTG GTA TGA
C-3′ and 5′-CAG TGA GGG TCT CTC TCT TCC T-3′.
Protein extraction and western blot
analysis
After washing twice with cold phosphate-buffered
saline (PBS), total proteins were extracted from HCC cells using
ice-cold modified radioimmunoprecipitation assay (RIPA) buffer (50
mM Tris-HCl pH 7.4, 1% Nonidet P-40, 0.25% sodium deoxycholate, 150
mM NaCl, 1 mM Na3VO4 and 1 mM NaF). Protein
lysates (30 µg) were separated by sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and
transferred to NC membranes (Millipore Corporation, USA). The
membranes were incubated overnight with reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
following primary antibodies: BCL-3 (1:200), cyclin D1 (1:500)
(both from Santa Cruz Biotechnology) and GAPDH (1:1,000; US
Biological, Swampscott, MA, USA). Then, the blots were incubated
with peroxidase-conjugated secondary antibodies (1:2,000-1:5,000;
Bio-Rad, Hercules, CA, USA), and visualized with the ECL system
(Amersham, Piscataway, NJ, USA).
Cell viability assay
HCC cells at the log-phase of growth were seeded
into 96-well plates at 2×103 cells/well. A solution (50
µl) of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium
bromide (MTT; Roche, USA) (5 mg/ml; KeyGen, China) was added into
each well at 24, 48 and 72 h after transfection. Four hours later,
the supernatant was removed, and 200 µl of dimethyl
sulfoxide (DMSO) was added to each well. Optical density (OD) was
measured at 490 nm to represent the viability of the HCC cells.
Cell cycle assay
Forty-eight hours after transfection, the cells were
washed with cold PBS twice, and were fixed with 70% ethanol at 4°C
overnight. After extensive washing, the cells were incubated with
50 µg/ml PI and 50 µg/ml RNase A for 1 h at room
temperature. After incubation, the cells were subjected to flow
cytometric analysis using a FACSCalibur (BD Biosciences, Bedford,
MA, USA). Experiments were performed in triplicate. The results are
presented as the percentage (%) of cells in a particular phase.
In vivo experiments
The nude mouse xenograft model was established using
4- to 6-week-old female BALB/c nude mice (Centre of Laboratory
Animals, The Medical College of Xi'an Jiaotong University, Xi'an,
China). HepG2 cells (4×106) transfected with BCL-3 shRNA
or NT shRNA were mixed in 150 µl PBS, and were
subcutaneously injected into the flank of each mouse. Tumor growth
curves were generated as previously described (18). All animal protocols were approved by
the Institutional Animal Care and Use Committee of Xi'an Jiaotong
University.
Statistical analysis
All statistical analyses were performed using the
SPSS Statistical Package for Windows version 13 (SPSS, Inc.,
Chicago, IL, USA) or GraphPad Prism 5 software (GraphPad Software,
Inc., San Diego, CA, USA). The quantitative data were compared
between groups using the Student's t-test or ANOVA. Categorical
data were analyzed using the Pearson's Chi-square test. The
Kaplan-Meier method and log-rank test were used to compare the
cumulative recurrence and survival rates. The independent factors
influencing the survival and recurrence of HCC patients were
determined using the Cox proportional hazards model. A value of
P<0.05 was considered to indicate a statistically significant
result.
Results
Expression of BCL-3 is elevated in HCC
tissues
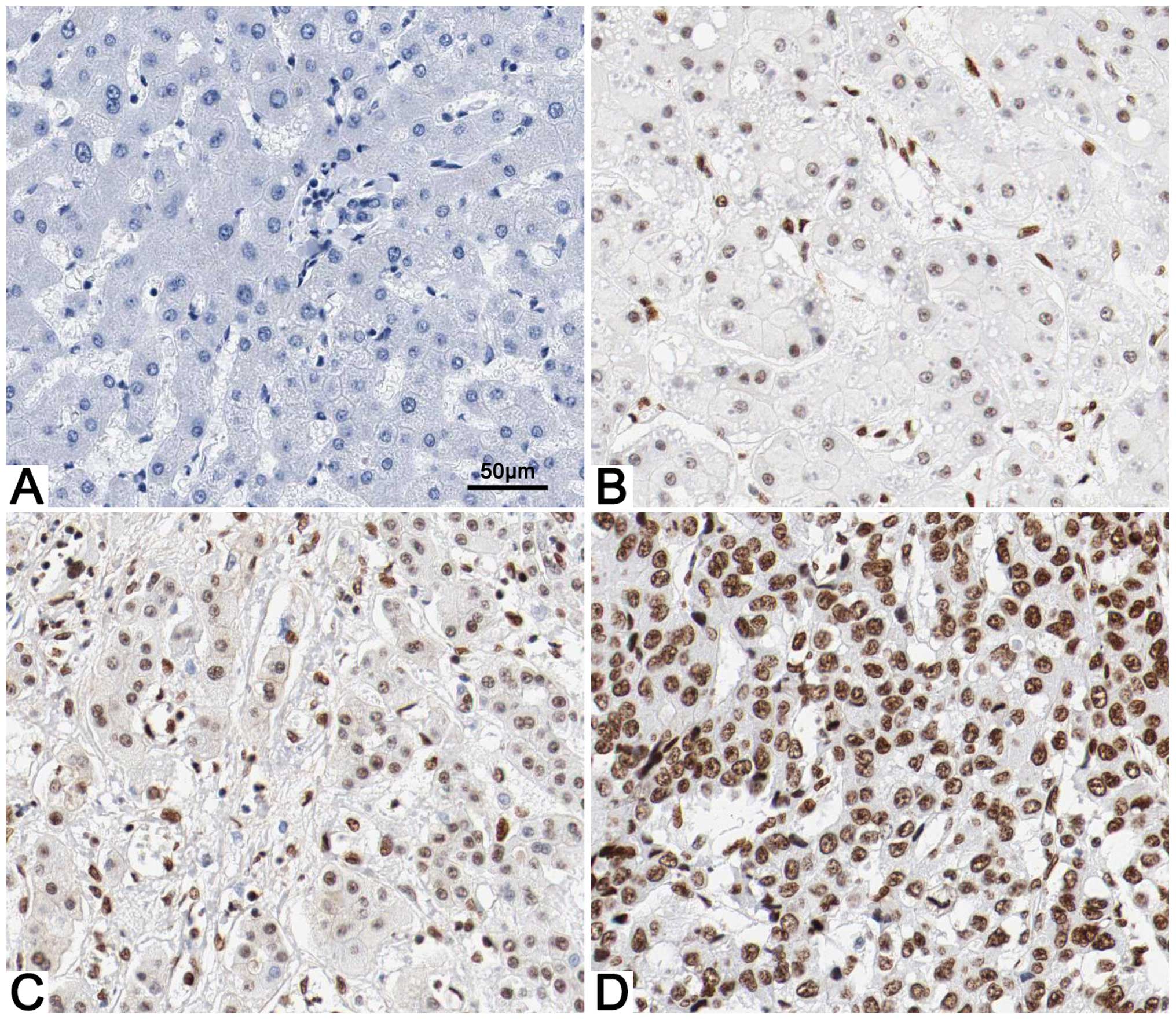
Immunohistochemical staining was first performed to
explore the difference in BCL-3 expression between the HCC and
adjacent non-tumor tissues. BCL-3 expression was considered as
either negative (scores 0–1) or positive (scores 2–3). BCL-3
expression was detected in 64.4% (58/90) of the HCC specimens,
whereas only 23.3% (21/90) of the non-cancerous tissues showed a
positive BCL-3 signal (P<0.05; Fig.
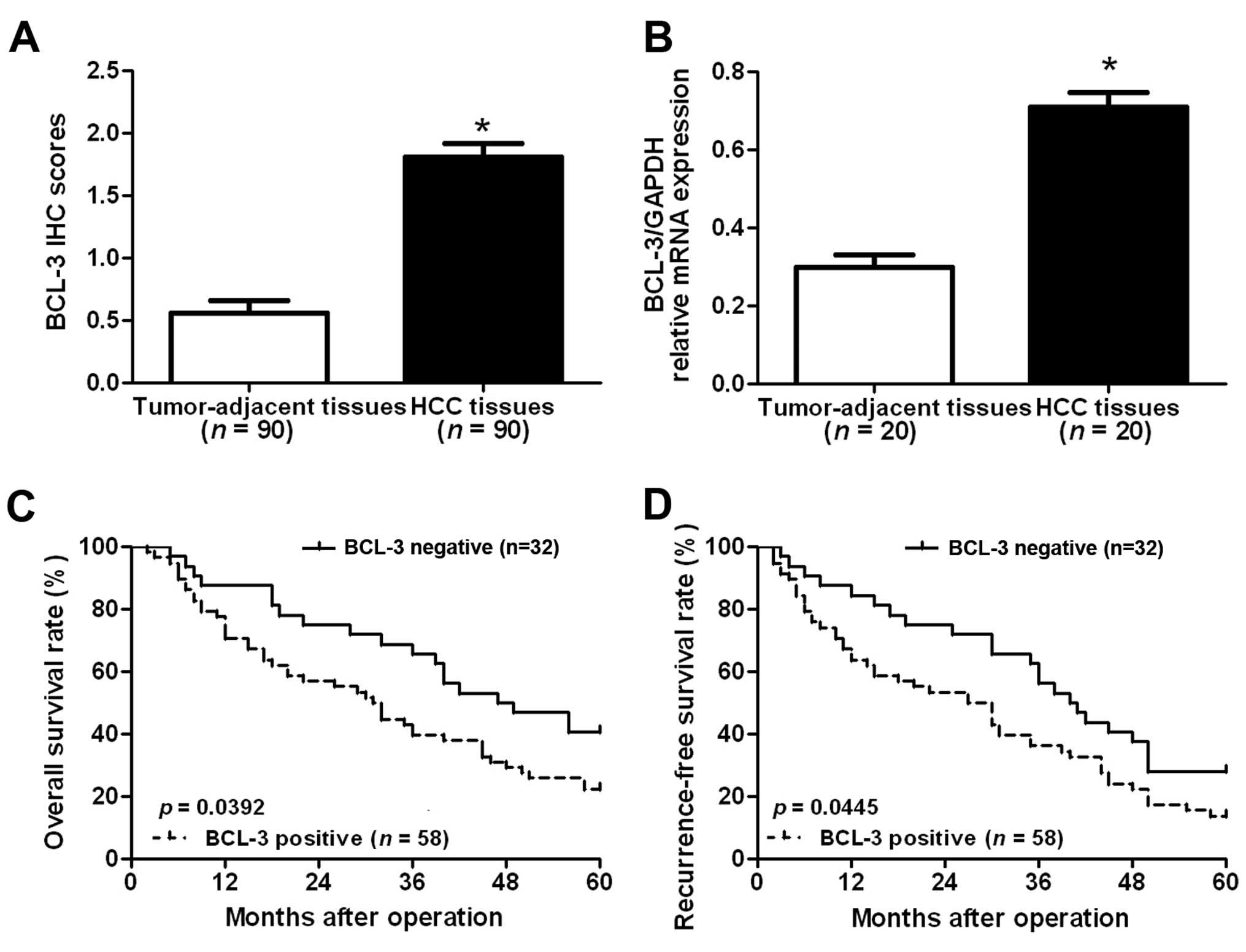
1). Furthermore, the results of IHC scores showed that the
level of BCL-3 expression in HCC tissues was significantly elevated
compared with that in the adjacent non-tumor tissues (P<0.05;
Fig. 2A). To further confirm the
results of IHC staining, we performed RT-qPCR to examine the mRNA
level of BCL-3 in the clinical tissues. The results of RT-qPCR
showed that the BCL-3 mRNA level in the HCC tissues was also
significantly elevated compared to that in the non-tumor tissues
(P<0.05; Fig. 2B). Taken
together, these results suggest that BCL-3 plays an oncogenic role
in the pathogenesis of HCC.
Positive expression of BCL-3 is
correlated with adverse clinicopathological features and poor
prognosis of HCC patients
To further determine the clinical significance and
prognostic value of BCL-3 in HCC, we first divided the patients
into two groups based on the staining of BCL-3 in HCC tissues
(positive group, n=58; negative group, n=32). Then, we compared the
clinicopathological features between patients in these two groups.
As shown in Table I, positive
expression of BCL-3 was associated with hepatitis B virus (HBV)
infection (P=0.015), tumor size (P=0.005), cirrhosis (P=0.018) and
TNM tumor stage (P=0.035). Furthermore, we compared the overall
survival (OS) and recurrence-free survival (RFS) between patients
in these two groups. Kaplan-Meier survival curves showed that
patients with positive staining of BCL-3 had significantly reduced
OS (P=0.0392; Fig. 2C) and RFS
(P=0.0445; Fig. 2D). These data
suggest that BCL-3 may be involved in the development and
progression of HCC, and can serve as a promising predictor of the
prognosis of HCC patients. Furthermore, multivariate Cox regression
analysis indicated that BCL-3 expression was an independent factor
for predicting both 5-year OS and RFS in HCC patients (P=0.016 and
0.009, respectively; Table
II).
| Table IIMultivariate Cox regression analysis
of the 5-year survival of 90 HCC patients. |
Table II
Multivariate Cox regression analysis
of the 5-year survival of 90 HCC patients.
| Variables | OS
| RFS
|
|---|
| HR | 95% CI | P-value | HR | 95% CI | P-value |
|---|
| HBV infection | 1.26 | 0.93–1.70 | 0.143 | 1.19 | 0.81–1.75 | 0.367 |
| Tumor size | 1.41 | 1.05–1.88 | 0.022a | 1.13 | 0.83–1.55 | 0.431 |
| Cirrhosis | 1.29 | 0.94–1.76 | 0.120 | 1.18 | 0.86–1.63 | 0.303 |
| TNM tumor
stage | 1.65 | 1.20–2.27 | 0.002a | 1.57 | 1.14–2.10 | 0.006a |
| BCL-3
expression | 1.58 | 1.09–2.29 | 0.016a | 1.48 | 1.10–1.98 | 0.009a |
BCL-3 promotes the growth of HCC cells
both in vitro and in vivo
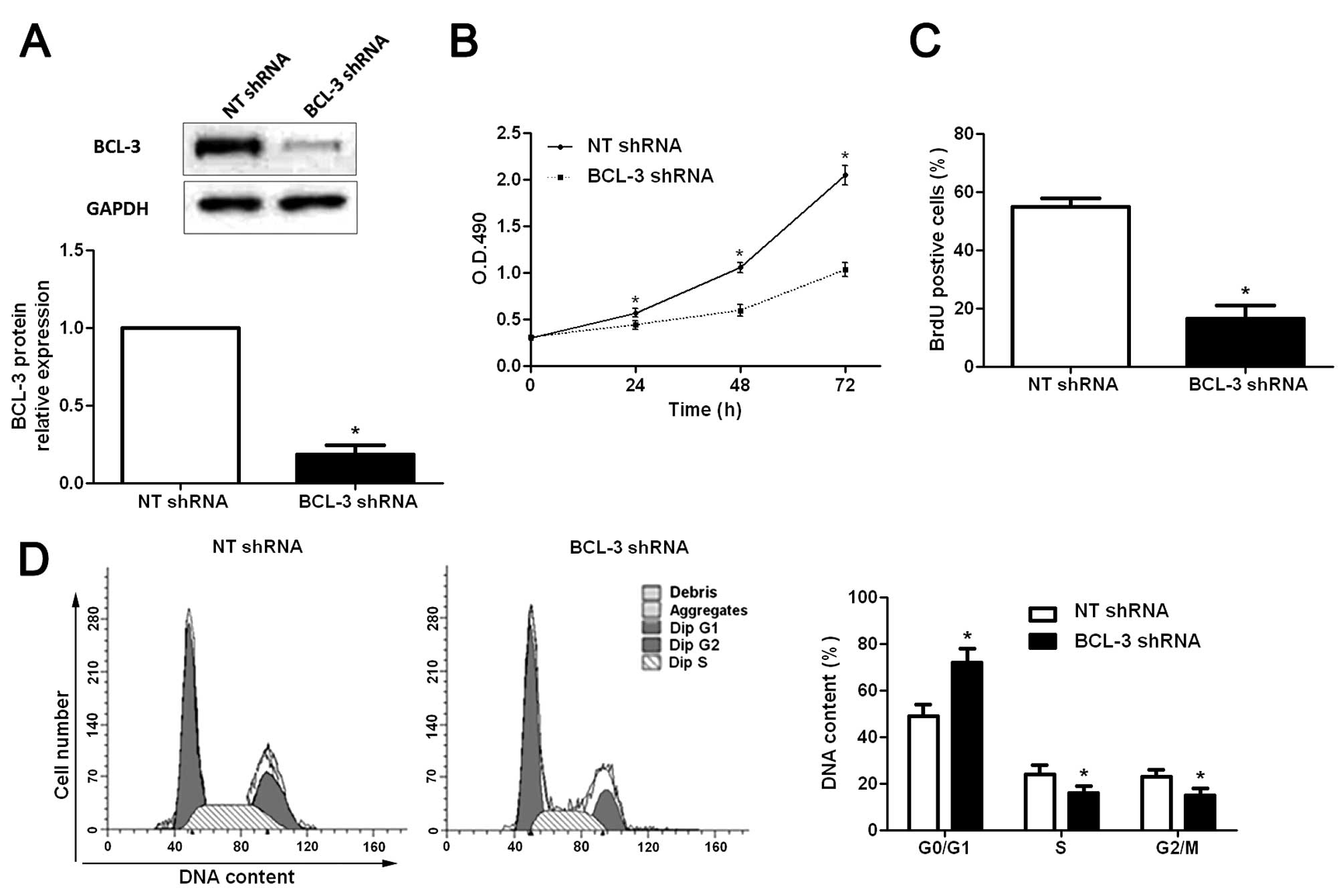
To explore the functional role of BCL-3 in HCC, a
BCL-3-specific shRNA was employed to suppress the expression of
BCL-3 in HepG2 cells. Western blotting was used to confirm the
knockdown of BCL-3 (P<0.05; Fig.
3A). Subsequently, the results of the MTT assays showed that
the viability of the HepG2 cells was significantly decreased after
BCL-3 knockdown (P<0.05; Fig.
3B). The proliferation of HepG2 cells was significantly
decreased following BCL-3 downregulation (P<0.05; Fig. 3C), as determined by BrdU
incorporation assays. In addition, cell cycle assays showed that
knockdown of BCL-3 significantly increased the percentage of cells
in the G0/G1 phase (P<0.05), and decreased the percentage of
cells in the S phase (P<0.05) and G2/M phase (P<0.05)
(Fig. 3D). In contrast, the
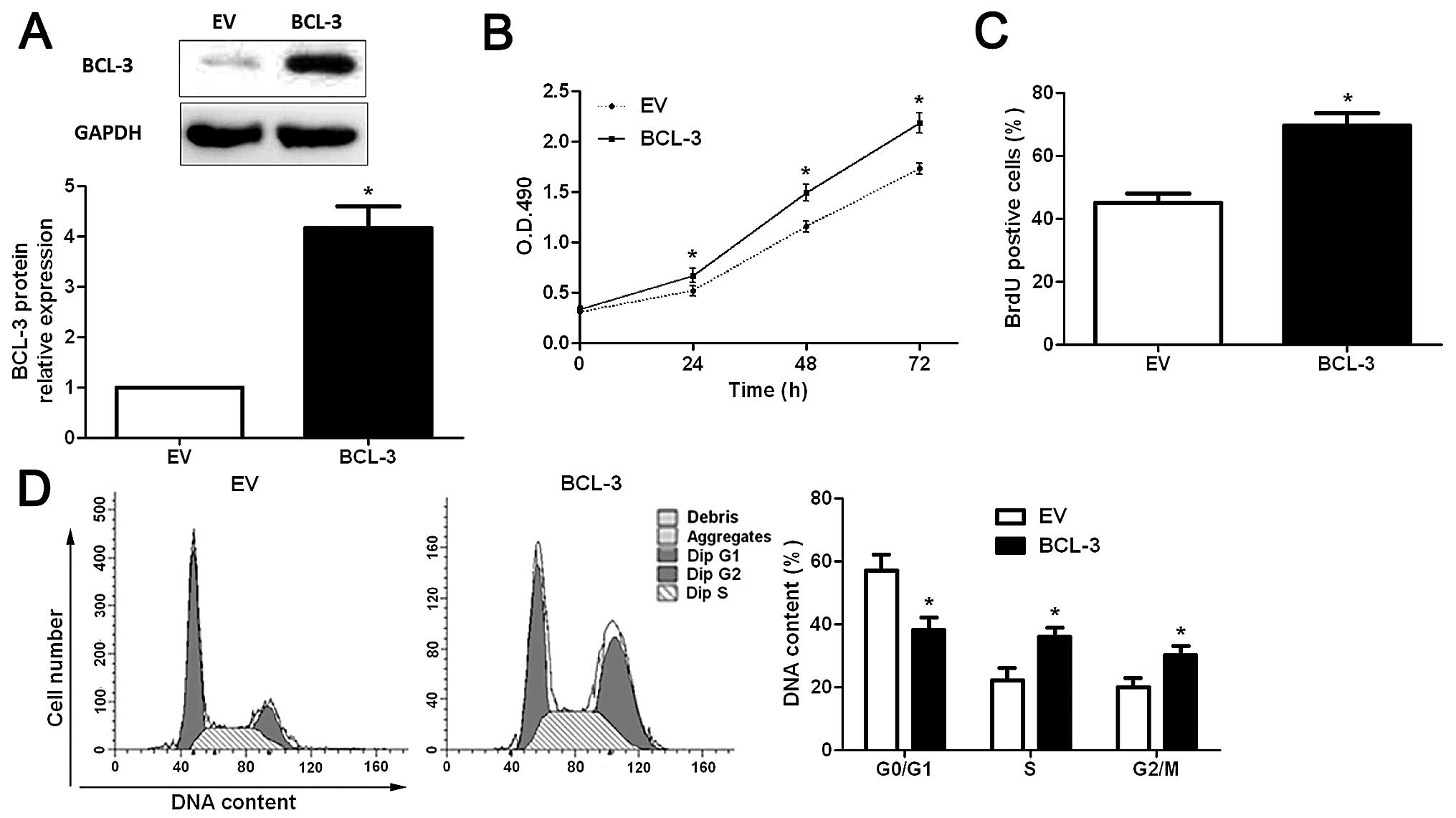
BCL-3-overexpressing plasmid obviously upregulated the expression
of BCL-3 in the Huh7 cells (P<0.05; Fig. 4A), and resulted in significantly
increased cell viability (P<0.05; Fig. 4B), proliferation (P<0.05;
Fig. 4C) and cell cycle progression
(P<0.05; Fig. 4D). These data
suggest that BCL-3 promotes the growth of HCC cells by regulating
cell viability, proliferation and cell cycle progression in
vitro.
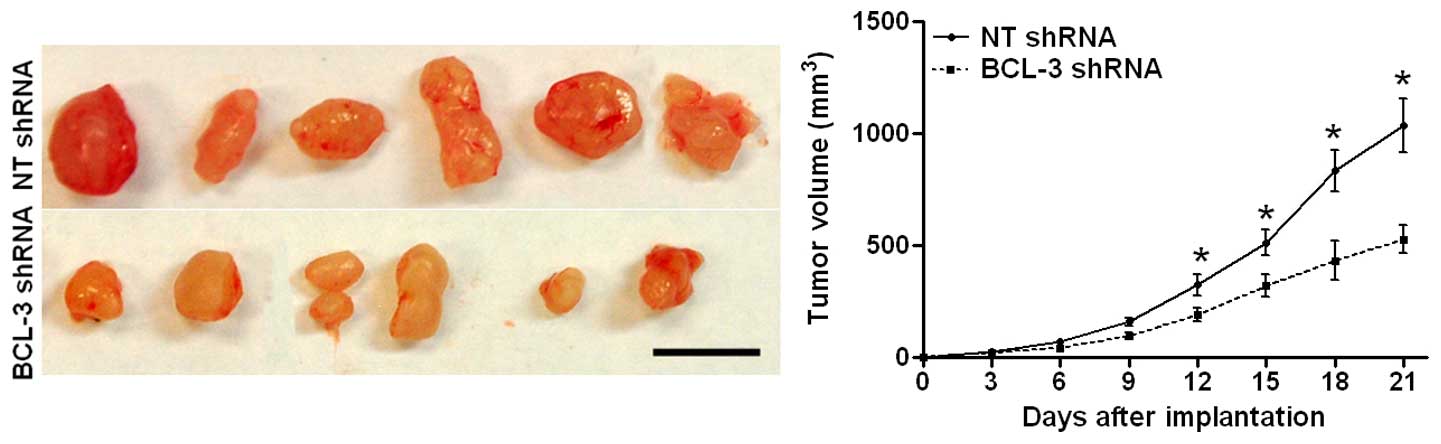
To further confirm the promoting effects of BCL-3 on
HCC cell growth in vivo, we subcutaneously injected HepG2
cells transfected with scramble shRNA or BCL-3 shRNA into nude
mice. As shown in Fig. 5A, BCL-3
knockdown significantly inhibited the growth of the HepG2 cells in
nude mice (P<0.05). Taken together, these data suggest that
BCL-3 promotes the growth of HCC cells both in vitro and
in vivo.
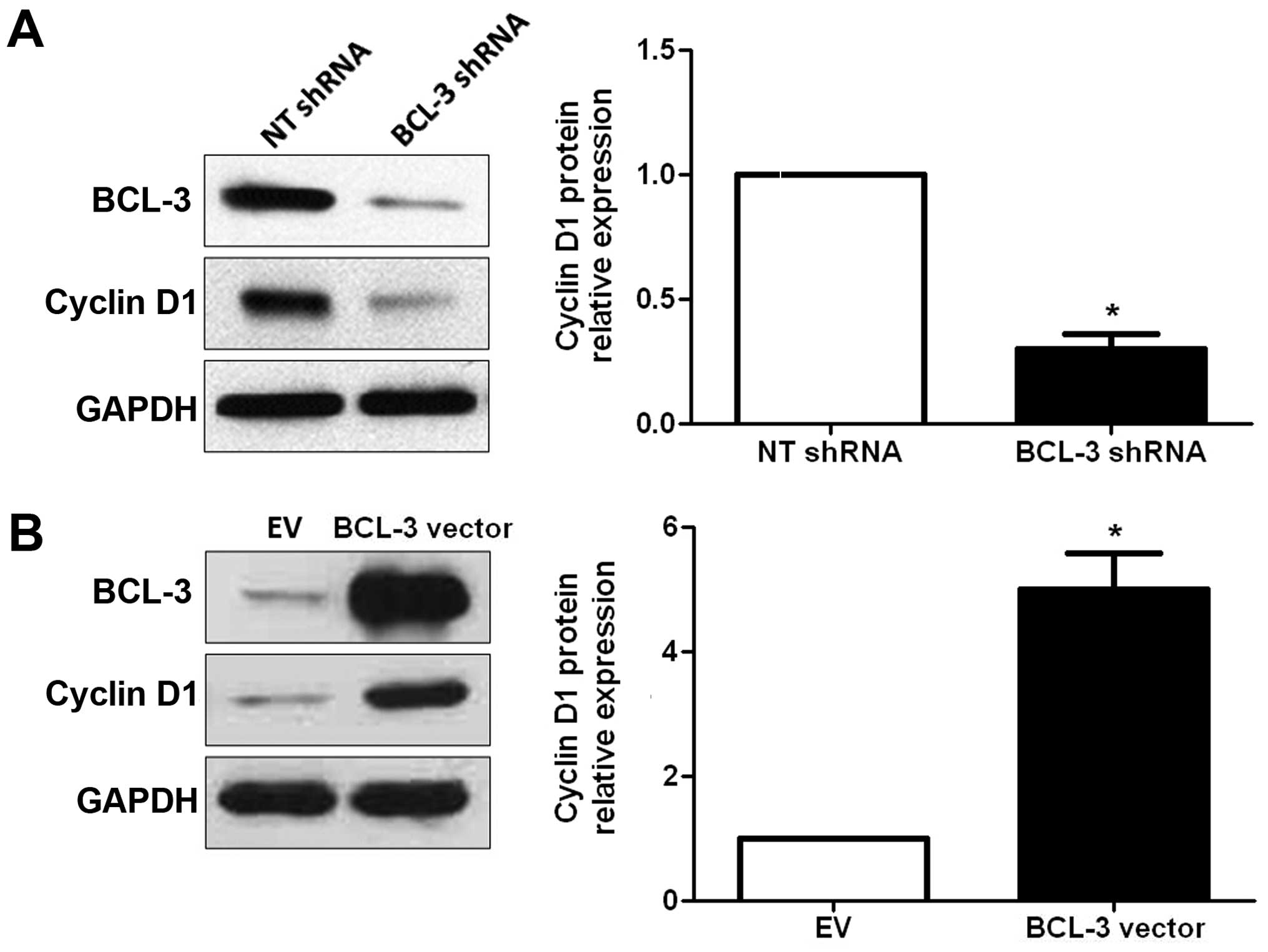
BCL-3 regulates the expression of cyclin
D1 in HCC cells
Cyclin D1 has been found to be overexpressed in HCC
(19,20), and plays an important role in
regulating cell cycle progression (21). Therefore, we ascertained whether
BCL-3 regulates the expression of cyclin D1 in HCC cells. The
results of the western blot analyses showed that the expression of
cyclin D1 was significantly reduced after downregulation of the
expression of BCL-3 (P<0.05; Fig.
6A). In contrast, overexpression of BCL-3 resulted in a
significant increase in cyclin D1 expression (P<0.05, Fig. 6B). These data indicate that BCL-3
regulates the expression of cyclin D1 in HCC cells.
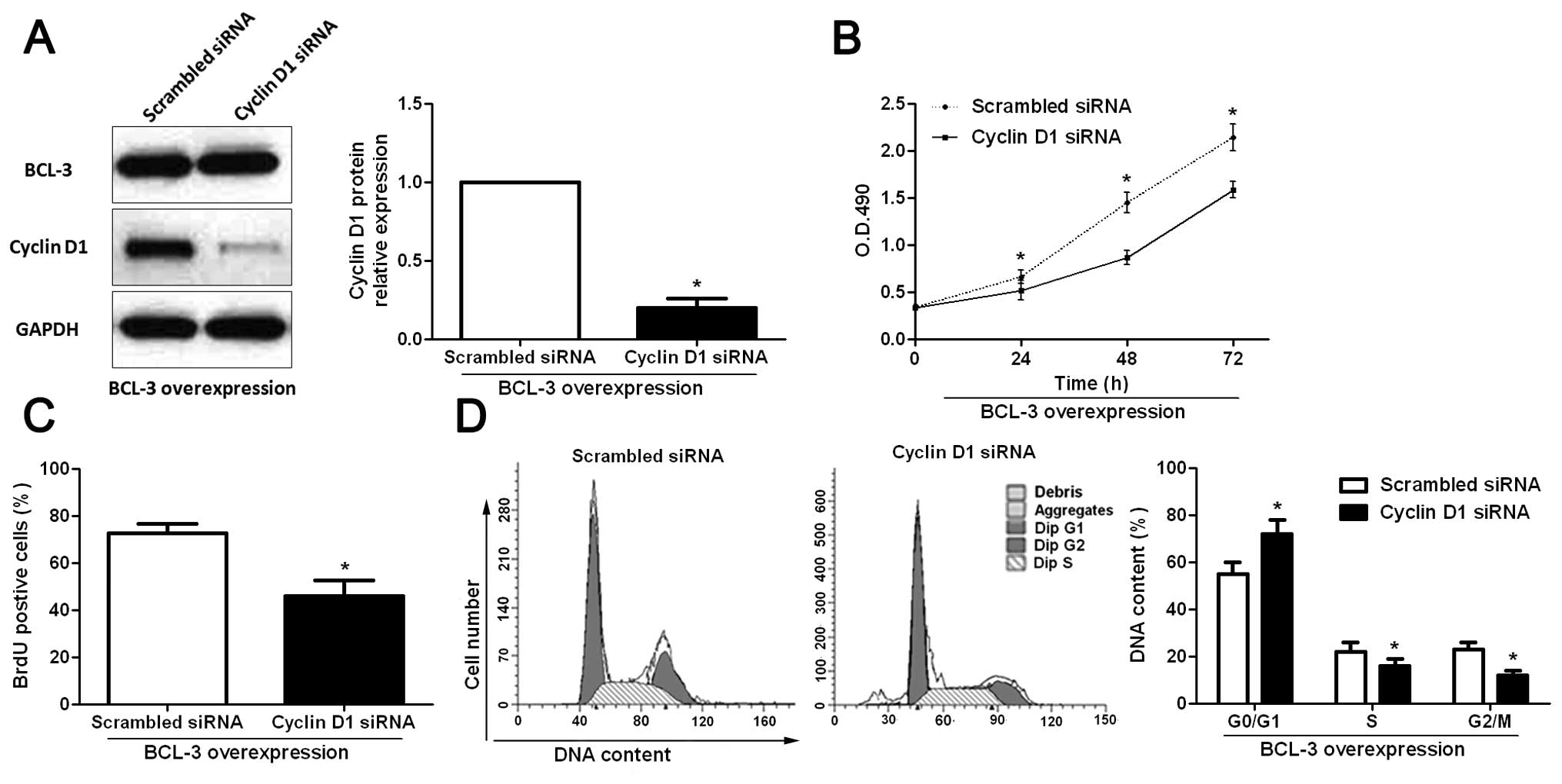
Cyclin D1 mediates the functional effects
of BCL-3 in HCC cells
To further determine whether cyclin D1 is a
functional mediator of BCL-3, we inhibited cyclin D1 expression in
the BCL-3-overexpressing Huh7 cells using cyclin D1-specific siRNA.
Cyclin D1 siRNA significantly inhibited the expression of cyclin D1
(P<0.05; Fig. 7A) in the
BCL-3-overexpressing Huh7 cells while it had no influence on the
expression of BCL-3 (Fig. 7A).
Functionally, inhibition of cyclin D1 partially abrogated the
effect of BCL-3 overexpression, resulting in a significant decrease
in cell viability (P<0.05; Fig.
7B) and proliferation (P<0.05; Fig. 7C), and cell cycle arrest (P<0.05;
Fig. 7D). These results
demonstrated that cyclin D1 is a downstream mediator of the
biological function of BCL-3 in HCC.
Discussion
Treatment of HCC, particularly for patients in an
advanced stage, is still a challenge for physicians (22). In addition, the long-term prognosis
for HCC patients is still dismal (23). Therefore, it is of great importance
to identify novel biomarkers and effective therapeutic targets,
which may promote the early diagnosis and improve the efficacy of
molecular-targeted drugs for HCC. In the present study, we examined
the expression status of BCL-3 in HCC tissues for the first time.
In addition, we found that, compared with adjacent non-tumor
tissues, HCC tissues harbored significantly higher expression of
BCL-3 at both the mRNA and protein levels. Importantly, positive
expression of BCL-3 was associated with poor clinicopathological
features and reduced survival of HCC patients. Notably, HCC
patients with HBV infection and cirrhosis showed significantly
higher levels of BCL-3 expression. Herein, previous studies of
hepatocarcinogenesis indicated that HBx protein upregulated the
expression of BCL-3 mRNA, which subsequently resulted in the
upregulation of the NF-κB2 (p52)/BCL-3 complex in the nucleus
(24). Therefore, these results
suggest that BCL-3 can potentially serve as a promising biomarker
for the early diagnosis and is a valuable predictor of the
prognosis of HCC patients.
The significant elevation of BCL-3 expression in HCC
tissues and its association with clinicopathological features and
prognosis of HCC patients, suggests a potential oncogenic role of
BCL-3 in HCC, and motivated us to explore its functional
significance in HCC. Previous studies have found that BCL-3
promotes the survival and inhibits the apoptosis of colorectal
cancer cells (25). In addition,
BCL-3 was found to be involved in the response to antiangiogenic
therapy in a mouse model of metastatic renal cell carcinoma
(16). In the present study, the
in vitro studies demonstrated that BCL-3 knockdown inhibited
cell viability, proliferation and cell cycle progression of HepG2
cells while its overexpression had promoting effects on these
biological behaviors in the Huh7 cells. In addition, in vivo
experiments showed that BCL-3 downregulation inhibited the growth
of HepG2 cells in nude mice. Therefore, our data revealed that
BCL-3 contributes to the growth of HCC by promoting cell viability,
proliferation and cell cycle progression.
Cyclin D1, a downstream mediator of the PI3K/GSK-3
signaling pathway, functions as a well-recognized regulator of cell
cycle progression and proliferation, and has been found to be
deregulated in many types of human cancers, including HCC (26,27).
In the present study, we confirmed that BCL-3 regulated the
expression of cyclin D1 in HCC cells using knockdown and
overexpression assays. Furthermore, we demonstrated that inhibition
of cyclin D1 expression in Huh7 cells overexpressing BCL-3
abrogated the functional influence of BCL-3 on Huh7 cells,
suggesting that BCL-3 not only regulated the expression of cyclin
D1, but also exerted its biological functions in HCC through cyclin
D1. Notably, a study of prostate cancer showed that BCL-3 regulated
the chemotherapeutic drug-induced apoptosis by modulating the
expression of Id-1 and Id-2 proteins (28). In addition, BCL-3 was found to
stabilize the expression CtBP1 by inhibiting proteasome-dependent
degradation, thus leading to increased resistance of breast cancer
cells to apoptosis (11).
Therefore, the functional role of BCL-3 in human cancers and the
underlying mechanisms by which BCL-3 exerts its functional
influence are largely dependent on the cancer type.
In summary, the present study confirmed for the
first time that BCL-3 is overexpressed in HCC tissues. Patients
with positive expression of BCL-3 had adverse clinicopathological
features and poorer prognosis. Functionally, BCL-3 can promote the
growth of HCC cells by promoting cell viability, proliferation and
cell cycle progression. Furthermore, BCL-3 can regulate the
expression of cyclin D1 in HCC cells, and its functional influence
on HCC cells is realized through cyclin D1. Therefore, the present
study demonstrated that BCL-3 can serve as a promising biomarker
for the early diagnosis and prognostic prediction of HCC patients,
and can potentially act as an effective therapeutic target of
HCC.
Acknowledgments
The present study was supported by a grant from the
National Natural Science Foundation of China (no. 81402039).
References
|
1
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar
|
|
2
|
Forner A, Llovet JM and Bruix J:
Hepatocellular carcinoma. Lancet. 379:1245–1255. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kim SH, Choi SB, Lee JG, Kim SU, Park MS,
Kim do Y, Choi JS and Kim KS: Prognostic factors and 10-year
survival in patients with hepatocellular carcinoma after curative
hepatectomy. J Gastrointest Surg. 15:598–607. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hanazaki K, Kajikawa S, Shimozawa N,
Mihara M, Shimada K, Hiraguri M, Koide N, Adachi W and Amano J:
Survival and recurrence after hepatic resection of 386 consecutive
patients with hepatocellular carcinoma. J Am Coll Surg.
191:381–388. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bours V, Franzoso G, Azarenko V, Park S,
Kanno T, Brown K and Siebenlist U: The oncoprotein Bcl-3 directly
transactivates through kappa B motifs via association with
DNA-binding p50B homodimers. Cell. 72:729–739. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cogswell PC, Guttridge DC, Funkhouser WK
and Baldwin AS Jr: Selective activation of NF-kappa B subunits in
human breast cancer: Potential roles for NF-kappa B2/p52 and for
Bcl-3. Oncogene. 19:1123–1131. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Au WY, Horsman DE, Ohno H, Klasa RJ and
Gascoyne RD: Bcl-3/IgH translocation (14;19)(q32;q13) in
non-Hodgkin's lymphomas. Leuk Lymphoma. 43:813–816. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Canoz O, Rassidakis GZ, Admirand JH and
Medeiros LJ: Immunohistochemical detection of BCL-3 in lymphoid
neoplasms: A survey of 353 cases. Mod Pathol. 17:911–917. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Schlette E, Rassidakis GZ, Canoz O and
Medeiros LJ: Expression of bcl-3 in chronic lymphocytic leukemia
correlates with trisomy 12 and abnormalities of chromosome 19. Am J
Clin Pathol. 123:465–471. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Maldonado V and Melendez-Zajgla J: Role of
Bcl-3 in solid tumors. Mol Cancer. 10:1522011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Choi HJ, Lee JM, Kim H, Nam HJ, Shin HJ,
Kim D, Ko E, Noh DY, Kim KI, Kim JH, et al: Bcl3-dependent
stabilization of CtBP1 is crucial for the inhibition of apoptosis
and tumor progression in breast cancer. Biochem Biophys Res Commun.
400:396–402. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Thornburg NJ, Pathmanathan R and
Raab-Traub N: Activation of nuclear factor-kappaB p50
homodimer/Bcl-3 complexes in nasopharyngeal carcinoma. Cancer Res.
63:8293–8301. 2003.PubMed/NCBI
|
|
13
|
Pallares J, Martínez-Guitarte JL, Dolcet
X, Llobet D, Rue M, Palacios J, Prat J and Matias-Guiu X:
Abnormalities in the NF-kappaB family and related proteins in
endometrial carcinoma. J Pathol. 204:569–577. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Puvvada SD, Funkhouser WK, Greene K, Deal
A, Chu H, Baldwin AS, Tepper JE and O'Neil BH: NF-κB and Bcl-3
activation are prognostic in metastatic colorectal cancer.
Oncology. 78:181–188. 2010. View Article : Google Scholar :
|
|
15
|
Liu Z, Jiang Y, Hou Y, Hu Y, Cao X, Tao Y,
Xu C, Liu S, Wang S, Wang L, et al: The IκB family member Bcl-3
stabilizes c-Myc in colorectal cancer. J Mol Cell Biol. 5:280–282.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
de Souza Braga M, da Silva Paiva KB,
Foguer K, Barbosa Chaves KC, de Sá Lima L, Scavone C and Bellini
MH: Involvement of the NF-κB/p50/Bcl-3 complex in response to
antiangiogenic therapy in a mouse model of metastatic renal cell
carcinoma. Biomed Pharmacother. 68:873–879. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tu K, Yang W, Li C, Zheng X, Lu Z, Guo C,
Yao Y and Liu Q: Fbxw7 is an independent prognostic marker and
induces apoptosis and growth arrest by regulating YAP abundance in
hepatocellular carcinoma. Mol Cancer. 13:1102014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dou C, Wang Y, Li C, Liu Z, Jia Y, Li Q,
Yang W, Yao Y, Liu Q and Tu K: MicroRNA-212 suppresses tumor growth
of human hepatocellular carcinoma by targeting FOXA1. Oncotarget.
6:13216–13228. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Deane NG, Parker MA, Aramandla R, Diehl L,
Lee WJ, Washington MK, Nanney LB, Shyr Y and Beauchamp RD:
Hepatocellular carcinoma results from chronic cyclin D1
overexpression in transgenic mice. Cancer Res. 61:5389–5395.
2001.PubMed/NCBI
|
|
20
|
Joo M, Kang YK, Kim MR, Lee HK and Jang
JJ: Cyclin D1 overexpression in hepatocellular carcinoma. Liver.
21:89–95. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Stacey DW: Cyclin D1 serves as a cell
cycle regulatory switch in actively proliferating cells. Curr Opin
Cell Biol. 15:158–163. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Severi T, van Malenstein H, Verslype C and
van Pelt JF: Tumor initiation and progression in hepatocellular
carcinoma: Risk factors, classification, and therapeutic targets.
Acta Pharmacol Sin. 31:1409–1420. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sun VC and Sarna L: Symptom management in
hepatocellular carcinoma. Clin J Oncol Nurs. 12:759–766. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Park SG, Chung C, Kang H, Kim JY and Jung
G: Up-regulation of cyclin D1 by HBx is mediated by NF-kappaB2/BCL3
complex through kappaB site of cyclin D1 promoter. J Biol Chem.
281:31770–31777. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Urban BC, Collard TJ, Eagle CJ, Southern
SL, Greenhough A, Hamdollah-Zadeh M, Ghosh A, Poulsom R, Paraskeva
C, Silver A, et al: BCL-3 expression promotes colorectal
tumorigenesis through activation of AKT signalling. Gut. Jun
1–2015.Epub ahead of print. piigutjnl-2014-308270. View Article : Google Scholar
|
|
26
|
Freiburghaus C, Janicke B,
Lindmark-Månsson H, Oredsson SM and Paulsson MA: Lactoferricin
treatment decreases the rate of cell proliferation of a human colon
cancer cell line. J Dairy Sci. 92:2477–2484. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zwijsen RM, Klompmaker R, Wientjens EB,
Kristel PM, van der Burg B and Michalides RJ: Cyclin D1 triggers
autonomous growth of breast cancer cells by governing cell cycle
exit. Mol Cell Biol. 16:2554–2560. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ahlqvist K, Saamarthy K, Syed Khaja AS,
Bjartell A and Massoumi R: Expression of Id proteins is regulated
by the Bcl-3 proto-oncogene in prostate cancer. Oncogene.
32:1601–1608. 2013. View Article : Google Scholar
|