Introduction
Colorectal cancer (CRC) is the second most common
malignancy of the digestive system in women and the third in men
worldwide, and is the third (women) and fourth (men) leading cause
of cancer-related mortality (1).
Conventional treatments for CRC currently include various
combinations of surgery, chemotherapy, radiotherapy and targeted
therapy. Due to early screening, reduced prevalence of risk factors
and/or improved treatment, the recurrence and the mortality rates
of CRC have largely decreased in the past decade (1). However, the prognosis of advanced or
distant-stage CRC remains poor, and the 5-year overall survival
rate is less than 13% in the US (2). Accumulating alteration of oncogenes
and tumor-suppressor genes is crucial for the pathogenesis and
progression of CRC. To improve the therapeutic efficacy for CRC
particularly metastatic CRC and its prognosis, it is therefore
urgently needed to better understand the molecular mechanisms
involved in CRC progression and metastasis, and correspondingly
identify potential CRC-associated therapeutic genes/targets.
The inhibitor of growth 4 (ING4) has been recognized
as a powerful tumor suppressor (3).
ING4 frequently exhibits alterations in human cancers such as
deletion, mutation, splicing variant and downregulation,
contributing to cancer initiation and progression as well as poor
prognosis (4). ING4 can repress the
loss of contact inhibition that is induced by myelocytomatosis
viral related oncogene, neuroblastoma derived (MYCN) and
myelocytomatosis viral oncogene homolog (MYC) (5). Growing evidence has further shown that
forced expression of ING4 can trigger tumor growth suppression via
induction of cell cycle alteration, apoptosis and toxic autophagy
in a large variety of cancers (3,6–8).
Additionally, ING4 can augment the therapeutic efficacy of
chemotherapy and intracavitary or external beam radiotherapy
(3,9–13).
Adenoviral-mediated ING4 and interleukin (IL)-24 double
tumor-suppressor gene therapy also was found to exhibit enhanced
antitumor activity (3,14,15).
Moreover, ING4 can suppress cancer metastasis via interaction with
liprin α1 and downregulation of matrix metalloproteinases (MMPs)
(7,16,17).
ING4 can also inhibit tumor angiogenesis by suppressing production
of proangiogenic factors through attenuating transcription activity
of nuclear factor κB (NF-κB) and hypoxia-inducible factor-1α
(HIF-1α) (18–20). Notably, ING4 can impair prooncogene
c-myc translation via interacting with AUF1 (21).
Clinical data have shown that ING4 is commonly
down-regulated in human CRC tissues, and is closely associated with
higher clinical stage, histological grade, microvessel density
(MVD) and lymph node metastasis (22,23).
However, the possible role and related mechanism of ING4 in the
progression of human CRC remain largely elusive. In the present
study, we assessed the expression of ING4 in low and high
metastatic human CRC cells, investigated the effect of ING4 on
growth, cell cycle distribution and invasion of high metastatic
human CRC cells following lentiviral-directed ING4 stable
expression, and delineated the potential mechanisms.
Materials and methods
Vectors, cell lines, reagents and
mice
The pAdTrack-CMV/ING4 plasmid containing the
humanized ING4 coding sequence (CDS) was previously constructed
(7). The lentiviral transfer
plasmid pLenti6.3/IRES/GFP carrying a green fluorescent protein
(GFP) marker gene and a Blasticidin S (BSD)-resistant gene, and the
lentiviral packing plasmids including pLP1, pLP2 and VSVG were
purchased from Novobio Science and Technology Inc. (Shanghai,
China). The LS174T, SW480, LoVo and SW620 human CRC cell lines, the
human colorectal mucous epithelial FHC cell line and the human
embryonic kidney 293T cell line were purchased from the Cell Bank,
Type Culture Collection of the Chinese Academy of Sciences
(Shanghai, China). RPMI-1640 medium and fetal bovine serum (FBS)
were purchased from HyClone (Logan, UT, USA). The MiniBEST
Universal RNA Extraction kit was purchased from Takara (Dalian,
Liaoning, China). The reverse transcriptase polymerase MuMLV and
the primer oligo(dT)18 were purchased from Thermo Fisher
Scientific (Shanghai, China). The FastStart Universal SYBR-Green
Master (ROX) kit was purchased from Roche (Shanghai, China).
Lipofectamine 2000 was purchased from Invitrogen (Shanghai, China).
Blasticidin S, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium
bromide (MTT) and the mammalian cell lysis kit were purchased from
Sigma (Shanghai, China). The 24-well Transwell chamber was
purchased from Corning Inc. (Corning, NY, USA). Matrigel was
purchased from BD Biosciences (Shanghai, China). The BCA protein
assay kit was purchased from Beyotime Biotechnology (Beijing,
China). The propidium iodide (PI) cell cycle detection kit was
purchased from Nanjing KeyGen Biotechnology Inc. (Nanjing, Jiangsu,
China). The human IL-6, IL-8 and vascular endothelial growth factor
(VEGF) and enzyme linked immunosorbent assay (ELISA) kits were
purchased from R&D Systems (Shanghai, China). The antibodies
specific for ING4, P21, cyclin E, CDK2, E-cadherin, N-cadherin,
vimentin, Snail (Snail1), Snail2 (Slug), ZEB1, Twist, β-actin and
CD31 were purchased from Santa Cruz Biotechnology (Shanghai, China)
and Cell Signaling Technology (Danvers, MA, USA). The Super
Enhanced chemiluminescence detection kit was purchased from
Applygen Technology Inc. (Beijing, China). The UltraSensitive™ SP
kit was purchased from Maixin (Fuzhou, Fujian, China). The primers
were synthesized from Sangon Biotechnology Inc. (Shanghai, China).
The 4-week-old female athymic BALB/c nude mice were purchased from
Shanghai Experimental Animal Center (Shanghai, China) and
maintained in the animal facility at Soochow University (Suzhou,
Jiangsu, China) according to the Animal Research Committee
guidelines of Soochow University.
Quantitative reverse transcription
(qRT)-PCR analysis
The endogenous ING4 expression in human CRC cells
was determined by quantitative real-time reverse transcription
(qRT)-PCR assay. In brief, the total cellular RNAs were extracted
from the LS174T, SW480, LoVo and SW620 human CRC cells and the
normal human colorectal mucous epithelial control FHC cells
(2×106 cells/each) using the MiniBEST Universal RNA
Extraction kit, respectively. The first-strand cDNAs were
synthesized from RNAs using reverse transcriptase MuMLV and
oligo(dT)18. The cDNAs were then subjected to SYBR-Green
I-based qPCR analysis with primers specific for human ING4:
(ING4-F1, 5′-gct cat gag gga cct aga cc-3′ and ING4-R1, 5′-ggc caa
ttt ttc ctc gga gc-3′ for amplifying 112 bp) or the housekeeping
gene β-actin (β-actin-F, 5′-ctc acc atg gat gat gat atc gc-3′ and
β-actin-R, 5′-agg aat cct tct gac cca tgc-3′ for amplifying 163 bp)
(an internal control) using the FastStart Universal SYBR-Green
Master (ROX) kit. The authenticity of the PCR products was verified
by melting curve analysis and agarose gel electrophoresis. The ING4
mRNA expression was normalized to internal control β-actin and
calculated using the 2−ΔΔCt method as previously
described (24).
Generation of a lentivirus expressing
ING4
A recombinant lentivirus expressing the humanized
ING4 tumor suppressor gene (LV-ING4) was generated as previously
described (25). Briefly, the
humanized ING4 CDS fragment was amplified by PCR using the
pAdTrack-CMV/ING4 plasmid (7) as a
template, and primers specific for humanized ING4 were: ING4-F2,
5′-gaa gct agc gcc acc atg gct gct ggg atg tat ttg-3′ and ING4-R2,
5′-ata ggc gcg ccc tat ttc ttc ttc cgt tct tg-3′ for amplifying 747
bp as primers, and then subcloned into the pLenti6.3/IRES/GFP
lentiviral transfer plasmid at the NheI and SgsI
restriction enzyme sites to construct pLenti6.3/ING4/IRES/GFP. The
pLenti6.3/ING4/IRES/GFP and the lentiviral packaging plasmids
including pLP1, pLP2 and VSVG were then cotransfected into the
human embryonic kidney packaging 293T cell line using Lipofectamine
2000. Seventy-two hours after transfection, the culture supernatant
was harvested and LV-ING4 was purified by ultracentrifugation. The
blank lentivirus without ING4 insertion (LV) used as the control
was similarly prepared as above.
Construction of the ING4-stably
transgenic CRC cell line
The biological titer (transducing U/ml, i.e. TU/ml)
of LV-ING4 and LV lentiviruses expressing the GFP marker gene was
evaluated by calculating the number of GFP-expressing 293T cells
after lentiviral infection by fluorescence microscopy following the
company protocol. The ratio of infectious lentivirus (TU) to target
cells is called the multiplicity of infection (MOI). For stable
expression of the lentiviral-mediated ING4 transgene in CRC cells,
the high metastatic human CRC LoVo cells were dispensed into
24-well plates at 1×105 cells/well and incubated with
LV-ING4 or LV at an MOI of 10 according to the infectious dose as
recommended by the manufacturer. Forty-eight hours after infection,
the GFP expression and lentivirus infection efficiency were
observed by fluorescence microscopy. To eliminate the uninfected
cells, the transfectants were then selected with Blasticidin S
(final concentration 10 µg/ml) for 1 month. Expression of
the GFP marker gene and the ING4 transgene in Blasticidin
S-resistant LV-ING4 or LV-infected LoVo (termed LoVo-ING4 or
LoVo-Mock) tumor cells were further identified by fluorescence
microscopy, RT-PCR and western blot analysis.
RT-PCR analysis
The LoVo-ING4 or LoVo-Mock human CRC and
untransfected control tumor cells (2×106 cells/each)
were collected. The lentiviral-mediated humanized ING4
transcriptional expression in the LoVo tumor cells was analyzed by
RT-PCR using primers specific for humanized ING4 (ING4-F2, 5′-gaa
gct agc gcc acc atg gct gct ggg atg tat ttg-3′ and ING4-R2, 5′-ata
ggc gcg ccc tat ttc ttc ttc cgt tct tg-3′ for amplifying 747 bp);
or the housekeeping gene β-actin (β-actin-F, 5′-ctc acc atg gat gat
gat atc gc-3′ and β-actin-R, 5′-agg aat cct tct gac cca tgc-3′ for
amplifying 163 bp). The reaction products were analyzed using
agarose gel electrophoresis.
Cell viability assay
The effect of lentiviral-mediated ING4 stable
expression on human CRC cell growth in vitro was assessed by
MTT assay. In brief, the LoVo-ING4 and LoVo-Mock tumor cells were
dispensed into 96-well plates at 1×104 cells/well/200
µl culture medium, i.e. RPMI-1640 medium supplemented with
10% FBS. At different time points of incubation (day 1–3), the
viability of the tumor cells was analyzed using the MTT kit
according to the manufacturer's protocols.
Flow cytometric analysis of cell cycle
distribution
The effect of lentiviral-mediated ING4 stable
expression on the cell cycle profile of human CRC cells in
vitro was determined by flow cytometric analysis using PI
staining. Briefly, the LoVo-ING4 and LoVo-Mock tumor cells were
harvested, washed in cold phosphate-buffered saline (PBS) and fixed
in 70% cold alcohol for 24 h. After washing, the cell pellets were
stained with PI solution (50 µg/ml PI, 50 µg/ml RNase
A and 0.1% Triton X-100) in the dark for 30 min and then analyzed
by flow cytometry.
Transwell invasion assay
The effect of lentiviral-mediated ING4 stable
expression on human CRC cell invasion in vitro was assessed
by Transwell invasion assay. In brief, 12.5 µl of Matrigel
(50 mg/l) was diluted in 87.5 µl serum-free RPMI-1640
medium. The 100 µl Matrigel diluted solution was added to a
24-well Transwell chamber, dried in a laminar hood overnight and
reconstituted in 100 µl serum-free RPMI-1640 medium for 2 h.
The LoVo-ING4 and LoVo-Mock tumor cells (2×105 cells/100
µl serum-free RPMI-1640 medium) were added to the upper
chamber of the Transwell. The lower chamber was filled with 500
µl of culture medium. After 24 h of incubation, tumor cells
on the upper surface of the filter were removed and cells invading
into the bottom side of the insert were fixed with 4%
paraformaldehyde, stained with crystal violet, photographed and
counted by investigators that were blinded to the group allocation
in 5 random ×200 high-power fields. The invasive ability of the
tumor cells was then analyzed.
Western blot analysis
The LoVo-ING4 and LoVo-Mock human CRC cells were
collected, washed with cold PBS and lysed in lysis buffer
(1×107 cells/1 ml lysis buffer) for preparation of total
cellular lysates using a mammalian cell lysis kit. The protein
concentration was determined by BCA protein assay. The total
cellular lysates (100 µg/lane) were then subjected to 12%
sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE) and western blot analysis of human ING4, P21, cyclin E,
CDK2, E-cadherin, N-cadherin, vimentin, Snail1, Snail2, ZEB1, Twist
and β-actin (an internal control). After washing, the membranes
were developed using the Super Enhanced chemiluminescence detection
kit. The bands were then visualized after their exposure to Kodak
X-ray film.
Enzyme-linked immunosorbent assay
(ELISA)
The LoVo-ING4 and LoVo-Mock human CRC cells were
dispensed into 24-well plates at 2×105 cells/well/1 ml
culture medium. The supernatants were collected at different time
points of incubation (day 1–3). The amounts of IL-6, IL-8 and VEGF
in the above supernatants were then determined by ELISA analysis
using human IL-6, IL-8 and VEGF ELISA kits, respectively.
Tumor transplantation assay in vivo
Female athymic BALB/c nude mice were subcutaneously
(s.c.) implanted with LoVo-ING4 or LoVo-Mock human CRC cells
(1×106 cells/mouse) (6 mice/group). Tumor progression
in vivo was monitored by investigators that were blinded to
the group allocation via measurement of tumor volume and weight.
Tumor volume (V) was measured with a caliper and calculated by the
formula: V = tumor size = ab2/2, where a is the larger
of the two dimensions and b is the smaller. The tumor-bearing mice
were sacrificed 4 weeks after tumor cell inoculation. The
xenografted tumors were then removed, weighed, fixed with 10%
neutral formalin and embedded in paraffin for hematoxylin &
eosin (H&E) staining and immunohistochemical analysis.
CD31 immunohistochemical analysis
The expression of tumor vessel CD31 in the LoVo-ING4
and LoVo-Mock human CRC s.c. xenografted tumors was examined by
immunohistochemical analysis using the UltraSensitive™ SP kit. Any
endothelial cell cluster immunoreactive for CD31 clearly separated
from adjacent microvessels was considered as a single countable
vessel (26). The MVD was
determined by investigators that were blinded to the group
allocation in 5 randomly selected high-power (magnification, ×200)
fields of each section under microscopy.
Statistical analysis
All data are presented as the mean ± standard
deviation (SD) and were statistically processed by the Student's
t-test for comparison of differences between two groups using SPSS
10.0 software (SPSS, Inc., Chicago, IL, USA). A value of p<0.05
was considered to indicate a statistically significant result.
Results
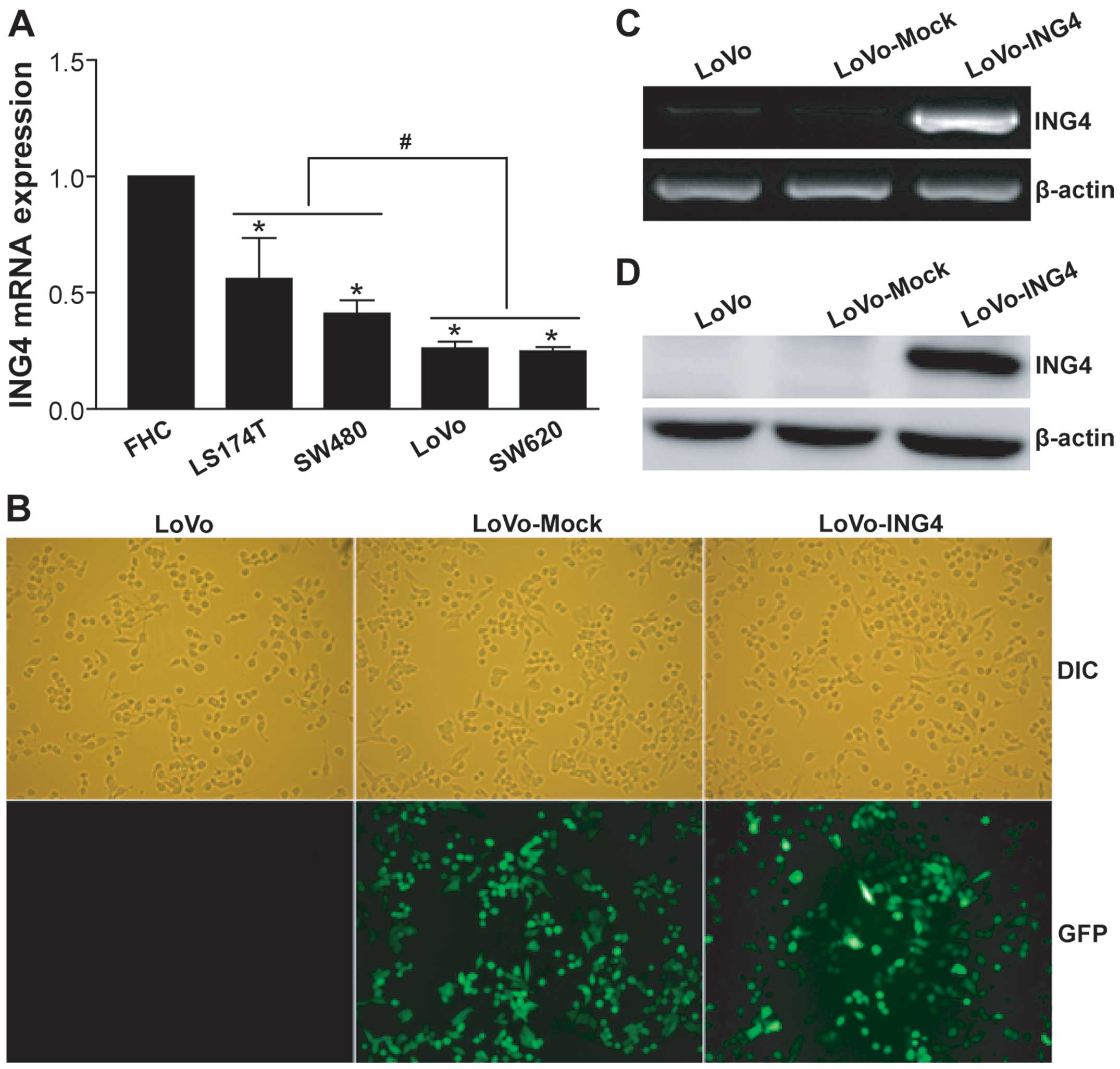
ING4 expression is downregulated in
CRC
Clinical evidence indicates that the ING4 tumor
suppressor is downregulated in human CRC (22,23).
To further evaluate ING4 expression in human CRC cells, we
quantified ING4 mRNA in a panel of human CRC cell lines including
low (LS174T and SW480) and high (LoVo and SW620) metastatic cell
lines by qRT-PCR analysis (Fig.
1A). Compared with normal human colorectal mucous epithelial
FHC cells, all of the tested low and high metastatic human CRC
cells displayed lower expression of ING4 (p<0.05). Moreover,
ING4 expression in the highly metastatic LoVo and SW620 CRC cells
was less than that in the lowly metastatic LS174T and SW480 CRC
cells (p<0.05). Our cellular model data supported previously
studied clinical data (22,23), suggesting that ING4 is reduced in
human CRC and may facilitate the progression and metastasis of
CRC.
Lentiviral-mediated ING4 stable
expression
To establish ING4 stably transgenic CRC cells, we
constructed a recombinant lentivirus LV-ING4-expressing humanized
ING4 gene and GFP marker gene. After infection of the LoVo cells
with 10 MOI LV-ING4 or LV followed by selection with Blasticidin S,
we obtained the LoVo-ING4 and LoVo-Mock transfectants. Fluorescence
microscopic analysis (Fig. 1B)
showed that >90% of GFP expression was found in the LoVo-ING4
and LoVo-Mock tumor cells, whereas GFP expression was not noted in
the uninfected LoVo control cells. To further detect the
lentiviral-mediated ING4 transgene expression, the LoVo-ING4 and
LoVo-Mock tumor and LoVo control cells were analyzed by RT-PCR
(Fig. 1C) and western blotting
(Fig. 1D). As shown in Fig. 1C and D, the lentiviral-mediated
exogenous ING4 gene was highly expressed at both the
transcriptional and translational levels in the LoVo-ING4 tumor
cells, but not in the LoVo-Mock and LoVo control cells. These
results indicated that the ING4-stably transgenic LoVo human CRC
cell line directed by the lentivirus was successfully
established.
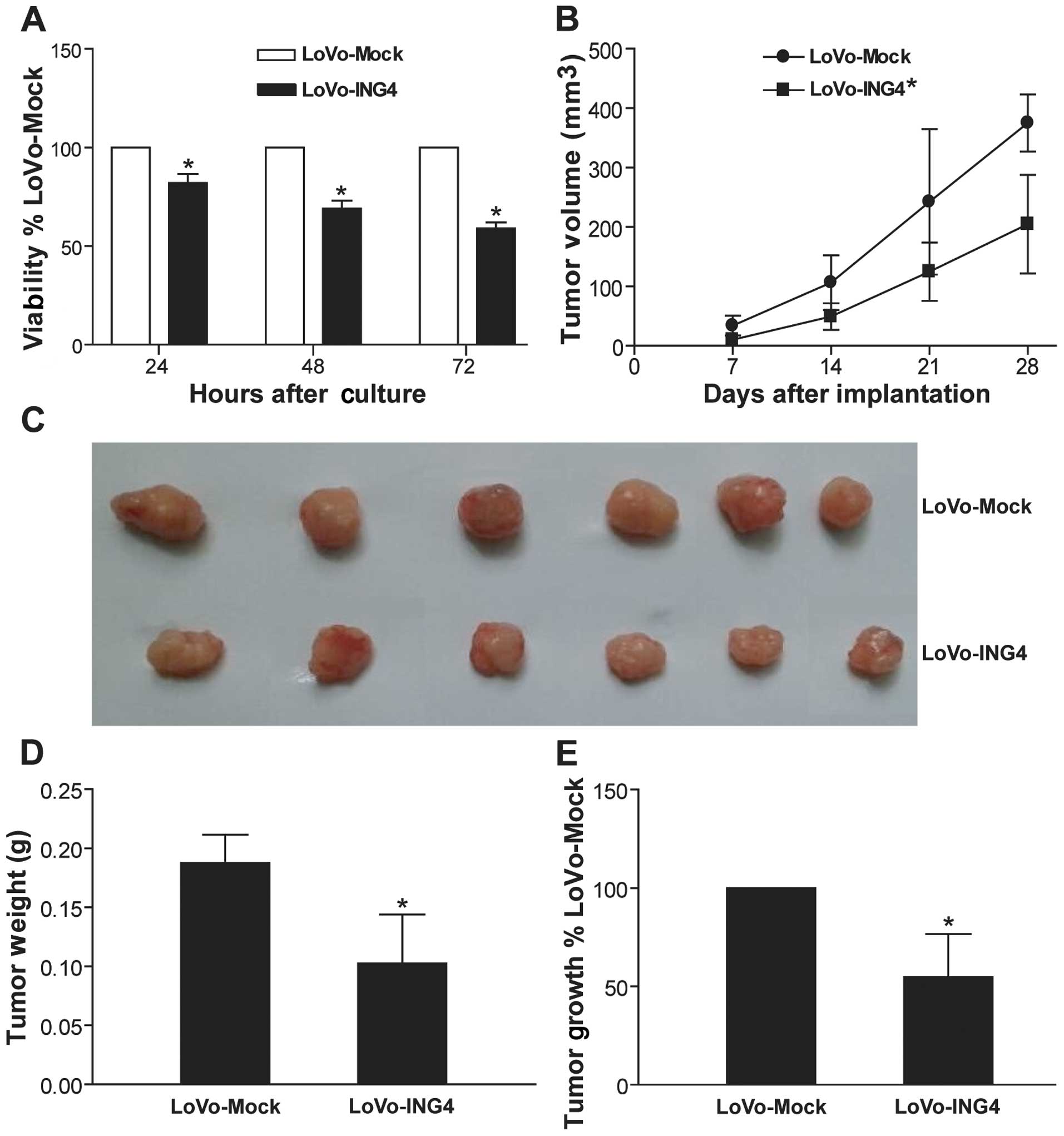
ING4 suppresses CRC growth
To examine the effect of lentiviral-mediated ING4
expression on human CRC growth in vitro, we generated
transgenic LoVo-ING4 and LoVo-Mock CRC cells and determined the
tumor cell viability by MTT assay. As shown in Fig. 2A, lentiviral-mediated ING4 gene
transfer obviously inhibited human CRC LoVo cell growth in
vitro in a time-dependent manner compared to the LoVo-Mock
group (p<0.05). To further assess whether the ING4-induced
growth-suppressive effect on CRC in vitro could be
reproduced in vivo, we monitored and compared the growth of
LoVo-ING4 and LoVo-Mock human CRC s.c. xenografted tumors in
athymic nude mice. As shown in Fig.
2B–E, the in vivo growth of the LoVo-ING4 CRC cells was
also markedly retarded (p<0.05).
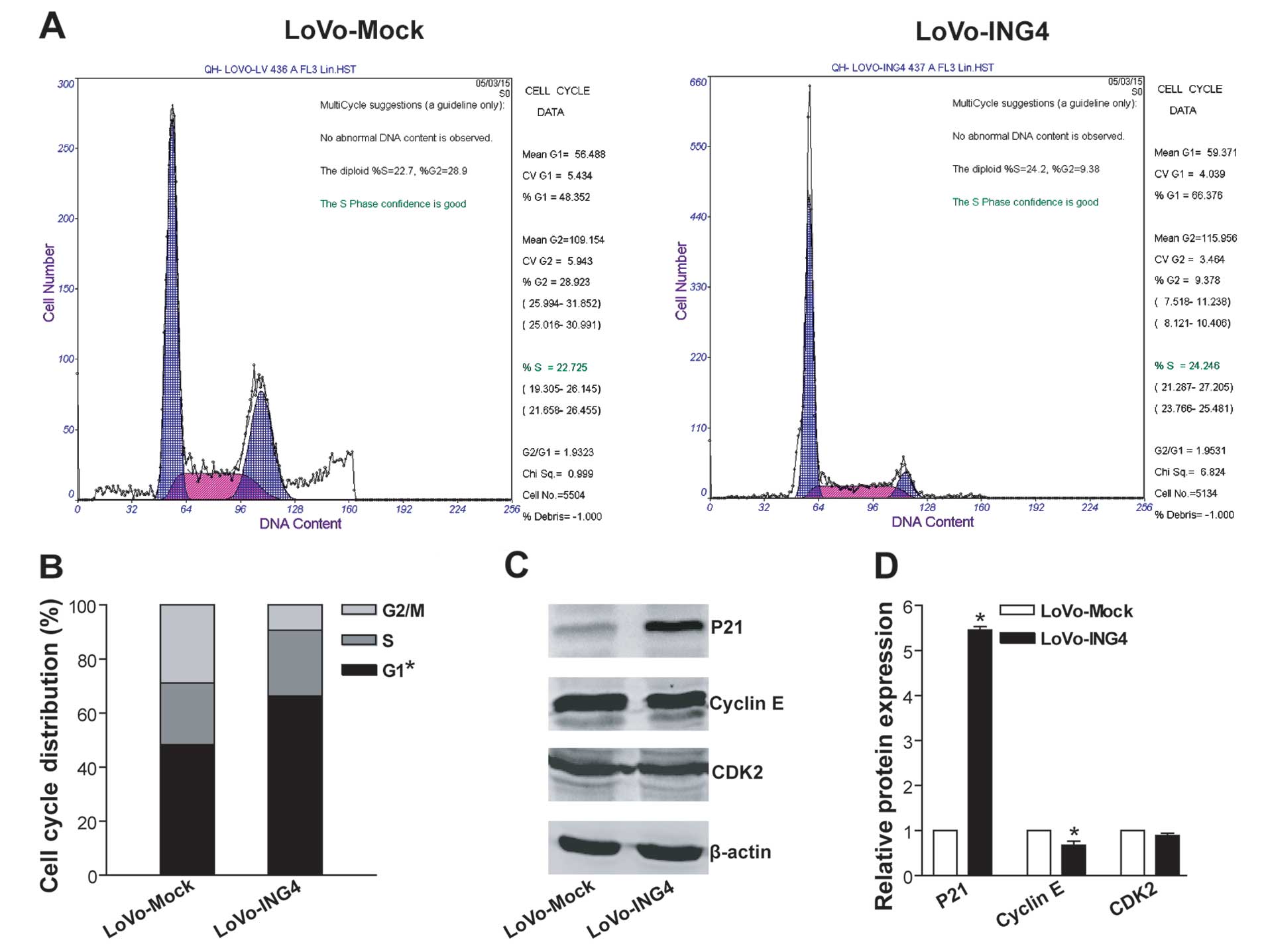
ING4 induces G1 phase arrest in CRC cells
possibly via upregulation of P21 and downregulation of cyclin
E
To explore the cellular mechanism responsible for
ING4-mediated tumor suppression in CRC cells, the cell cycle
profile of LoVo-ING4 and LoVo-Mock CRC cells was analyzed using PI
staining by flow cytometry. As shown in Fig. 3A and B, LoVo-ING4 transgenic CRC
cells exhibited a significant increase in the cell cycle G1 phase
population (G1 population in the total cell population, 67.8±5.2%)
compared with LoVo-Mock control tumor cells (48.1±3.7%)
(p<0.05), indicating that ING4 efficiently elicits LoVo CRC G1
arrest. To further elucidate the molecular mechanism underlying
ING4-induced G1 arrest, we analyzed the expression of G1-related
proteins such as Cip/Kip family cyclin-dependent kinase (CDK)
inhibitor P21, cyclin E and CDK2 by western blotting (Fig. 3C and D). As expected, ING4
substantially upregulated P21 expression as well as downregulated
cyclin E expression in the LoVo cells (p<0.05).
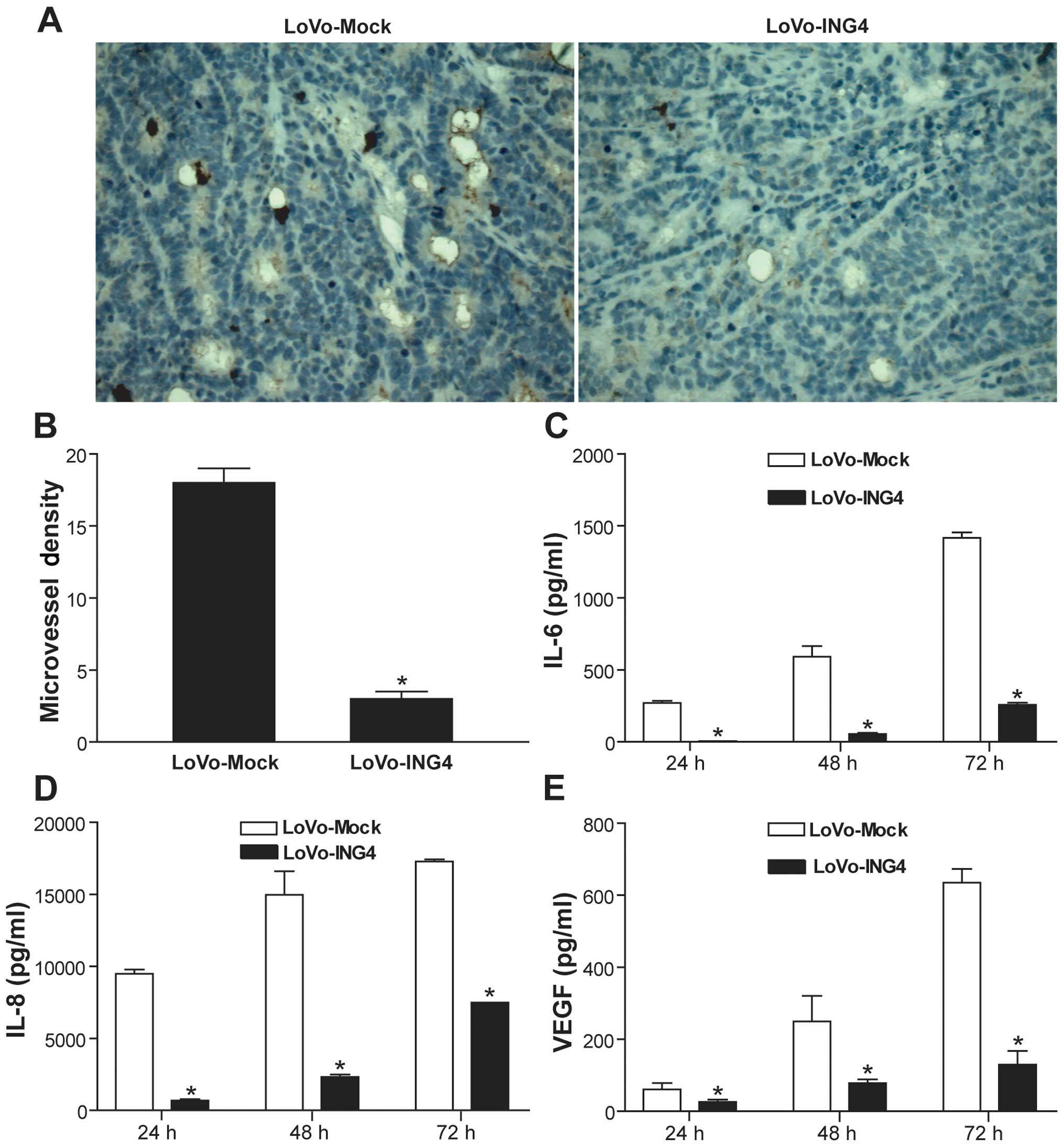
ING4 inhibits tumor angiogenesis via
downregulation of IL-6, IL-8 and VEGF
To investigate the effect of lentiviral-mediated
ING4 expression on tumor angiogenesis in vivo, the MVD in
LoVo-ING4 and LoVo-Mock human CRC s.c. xenografted tumor sections
was detected by CD31 immunohistochemical analysis. The positive
expression of CD31 was mainly presented as brownish yellow or
brownish granules in tumor vascular endothelial cells of the
xenografted tumors (Fig. 4A).
Compared with the LoVo-Mock group, the CD31 expression in the
LoVo-ING4 group was weaker or less (Fig. 4A). In addition, the MVD assessed in
the LoVo-ING4 group was significantly less than that in the
LoVo-Mock group (Fig. 4B)
(p<0.05). To delineate the mechanism involved in the
ING4-induced inhibition of in vivo tumor angiogenesis, we
examined the secretion of proangiogenic factors such as IL-6, IL-8
and VEGF from LoVo-ING4 and LoVo-Mock tumor cells by ELISA
analysis. Our data showed that lentiviral-mediated ING4 expression
markedly downregulated the levels of IL-6 (Fig. 4C), IL-8 (Fig. 4D) and VEGF (Fig. 4E) in the CRC LoVo cells
(p<0.05).
ING4 suppresses CRC invasion via reversal
of EMT
To assess the effect of ING4 on CRC cell invasion
in vitro, we compared the invasive ability of the LoVo-ING4
and LoVo-Mock tumor cells by Transwell chamber invasion assay. As
shown in Fig. 5A and B, the
lentiviral-mediated ING4 expression obviously inhibited the
invasion of the LoVo cells compared with the invasive ability of
the LoVo-Mock control cells (relative invasive ability, 43.2±3.8%
of the LoVo-Mock group) (p<0.05). To elucidate the molecular
mechanism associated with the ING4-mediated tumor invasion
suppression, the expression of epithelial marker E-cadherin,
mesenchymal markers as N-cadherin and vimentin, and
epithelial-mesenchymal transition (EMT)-inducing transcription
factors (EMT-TFs) Snail1, Snail2, ZEB1 and Twist in the LoVo-ING4
and LoVo-Mock tumor cells was analyzed by western blotting
(Fig. 5C and D). Compared with the
LoVo-Mock group, ING4 significantly increased the E-cadherin
expression as well as decreased the N-cadherin and vimentin
expression in the LoVo cells (p<0.05). Notably, ING4 markedly
suppressed the Snail1 expression in the LoVo cells (p<0.05).
These data indicated that ING4 inhibits CRC invasion and metastasis
via reversal of EMT through downregulation of Snail1 EMT-TF and a
switch from N-cadherin to E-cadherin.
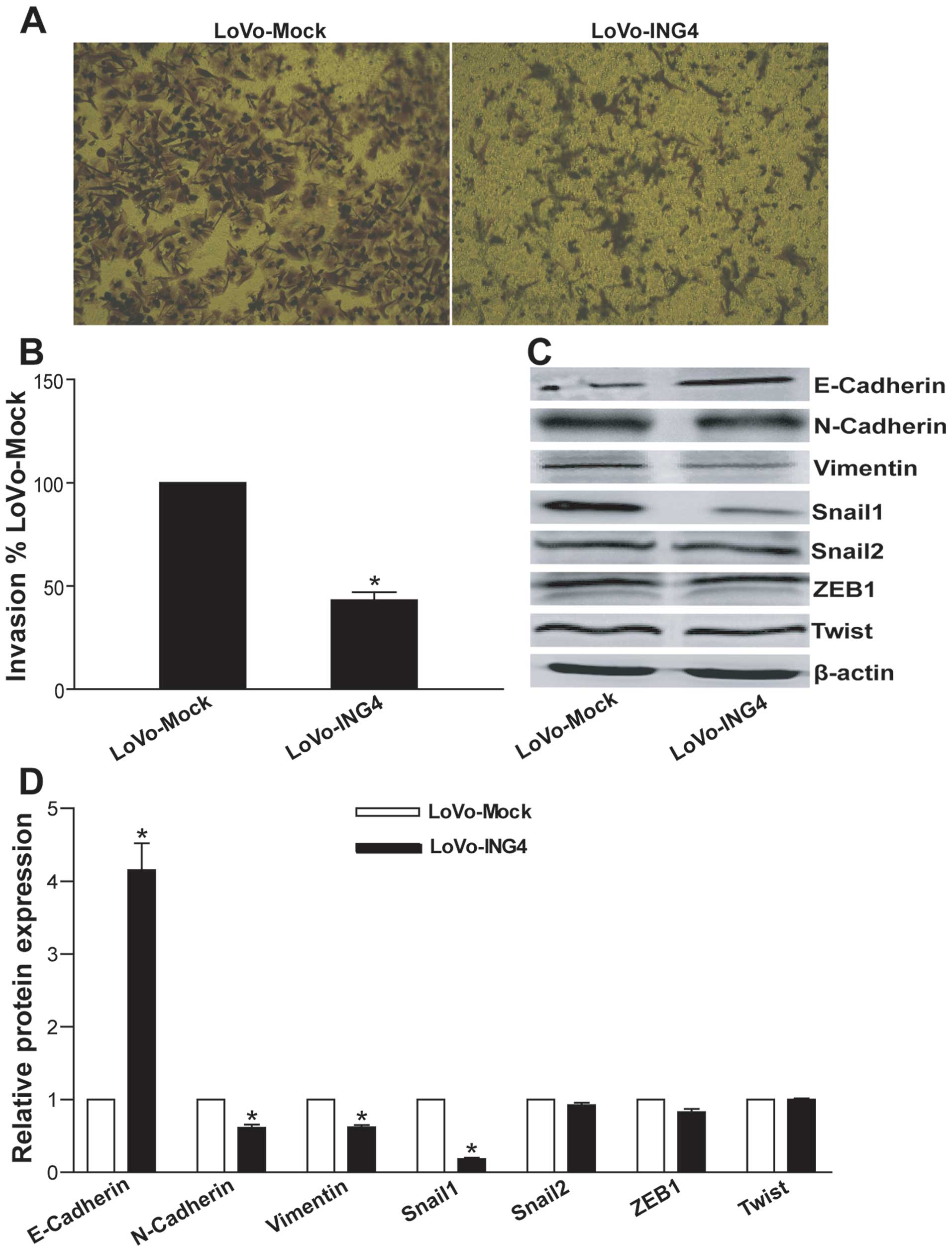 | Figure 5ING4 inhibits CRC invasion via
reversal of EMT by downregulation of Snail1. (A and B) ING4
inhibits CRC invasion in vitro. (A) Representative images of
the Transwell invasion assay are shown. (B) The relative invasive
ability of tumor cells was determined by comparison with the
LoVo-Mock group. *p<0.05 compared with LoVo-Mock
group; Student's t-test, n=3 replicates/condition, n=5
observations/replicate. (C and D) Western blot analysis of
EMT-associated proteins. The total cellular lysates derived from
the LoVo-ING4 and LoVo-Mock tumor cells were immunoblotted with a
panel of antibodies specific for E-cadherin, N-cadherin, vimentin,
Snail1, Snail2, ZEB1, Twist and β-actin (a loading control),
respectively. (C) Representative images of western blot analysis
are shown. (D) Expression of each index was normalized to the
expression level of β-actin, and the relative change was expressed
as a ratio or fold, with 1 being the value for the LoVo-Mock group.
*p<0.05 compared with the LoVo-Mock group; Student's
t-test, n=3 replicates/sample. Data shown are representative of
three independent experiments. |
Discussion
It has been found that the expression level of the
ING4 tumor suppressor in human colorectal cancer (CRC) is
negatively correlated with clinical stage, tumor angiogenesis and
metastasis (22,23). This evidence promoted us to further
examine the role of ING4 in human CRC progression using a high
metastatic LoVo CRC tumor model. In the present study, we found
that lentiviral-mediated ING4 gene transfer induced obvious tumor
growth suppression, G1 phase arrest, inhibition of invasion and
reduced MVD in the human CRC LoVo cells in vitro and/or
in vivo in athymic BALB/c nude mice.
The dyregulation of cell cycle control plays an
important role in cancer growth. Previous studies have demonstrated
that ING4 can suppress tumor growth via induction of S phase
reduction and G2/M phase arrest (6,7,10).
Inconsistent with these findings, our studies showed that
lentiviral-mediated ING4 expression efficiently induced G1 phase
arrest in the highly metastatic human CRC LoVo cells. These results
suggest that the modulatory effect of ING4 on cancer cell cycle is
cell type-dependent. The cell cycle progression is subtly tuned by
cell cycle regulatory proteins including cyclins, cyclin-dependent
kinases (CDKs) and CDK inhibitors (27). P21 as a member of the Cip/Kip family
is a critical CDK inhibitor that can inhibit the activity of the
cyclin E/CDK2 complex, resulting in G1 phase arrest (27). It has also been shown that ING4
upregulates P21 expression in tumor cells via a p53-dependent
mechanism through enhancement of p53 acetylation and transcription
activity (6). To examine the
molecular mechanism involved in the ING4-induced G1 arrest, we
analyzed the levels of P21, cyclin E and CDK2 by western blot
analysis. We found that ING4 markedly upregulated the
p53-downstream gene P21 and downregulated cyclin E in wild-type p53
LoVo cells, which may be an important mechanism responsible for
ING4-mediated LoVo CRC G1 arrest and growth inhibition.
Tumor angiogenesis as a hallmark of cancer is
indispensable for progressive tumor growth and metastasis, and is a
potential anticancer therapeutic target (28). A great deal of data has revealed
that inhibition of tumor angiogenesis and vessel normalization
represents a promising and non-toxic anticancer strategy (29,30).
ING4 has been found to suppress tumor angiogenesis via
downregulation of IL-6 and IL-8 proangiogenic factors through
attenuation of NF-κB and HIF-1α signaling (18,20).
To investigate the effect of the lentiviral-mediated ING4
expression on the angiogenesis of human CRC s.c. xenografted tumors
in vivo, the microvessel density (MVD) in LoVo xenografted
tumor tissues was determined by CD31 immunohistochemical analysis.
Our data showed that ING4 markedly downregulated tumor vessel CD31
expression and reduced MVD in the LoVo human CRC xenografted
tumors, which may be another important mechanism accountable for
ING4-induced in vivo LoVo human CRC growth inhibition in the
athymic nude mice. To delineate the mechanism underlying the
ING4-mediated inhibition of in vivo tumor angiogenesis, the
effect of ING4 on expression of proangiogenic factors IL-6, IL-8
and VEGF which are regulated by NF-κB and HIF-1α transcription
factors (18,20,31) in
LoVo CRC cells was further assessed. We found that ING4 profoundly
downregulated the expression of IL-6, IL-8 and VEGF in the LoVo
cells. These results indicated that ING4 suppresses LoVo CRC tumor
angiogenesis possibly via reduction of proangiogenic factors IL-6,
IL-8 and VEGF.
Tumor invasion and metastasis are key hallmarks of
cancer, resulting in as much as 90% of cancer-related deaths
(28,32). Distant metastasis is also the major
cause of cancer-related mortality in CRC patients.
Epithelial-mesenchymal transition (EMT) as a developmental
regulatory program has been shown to be prominently implicated in
cancer invasion and metastasis (28,33).
Downregulation of epithelial marker E-cadherin and upregulation of
mesenchymal marker N-cadherin (also referred to as cadherin switch)
is a hallmark of EMT (33).
Cancer-associated EMT in epithelial cancers induces a mesenchymal
phenotype with increased migration and invasion potential. The
activation of EMT is orchestrated by EMT-inducing transcription
factors (EMT-TFs) such as Snail (Snail1), Snail2 (Slug),
zinc-finger E-box-binding homeobox 1/2 (ZEB1/2) and Twist that
directly or indirectly repress key epithelial marker E-cadherin and
activate key mesenchymal marker N-cadherin (33). To elucidate the molecular mechanism
involved in the ING4-induced inhibition of invasion, we determined
the expression of E-cadherin, N-cadherin, vimentin and EMT-TFs
including Snail1, Snail2, ZEB1 and Twist by western blotting. We
demonstrated that ING4 obviously downregulated the expression of
Snail1 in the LoVo cells, leading to the upregulation of E-cadherin
and the reduction of N-cadherin and vimentin. Our data suggest that
ING4 suppresses CRC invasion and metastasis via reversal of EMT
through downregulation of Snail1 and a switch from N-cadherin to
E-cadherin.
In summary, our study showed that ING4 inhibits CRC
growth via induction of G1 phase arrest through regulating G1 phase
checkpoint molecules, and inhibition of tumor angiogenesis by
reducing proangiogenic factors. Importantly, the present study also
provides the first compelling evidence that ING4 is capable of
suppressing CRC invasion and metastasis via reversal of EMT through
downregulation of EMT-TF Snail1.
Acknowledgments
The present study was supported by grants from the
National Natural Science Foundation of China (NNSFC) (nos.
81372443, 81001016, 81272542, 81572992 and 81272737), and the
Science and Technology Department of Jiangsu Province (nos.
BL2014039 and BY2015039).
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics. CA Cancer
J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, Desantis C and Jemal A:
Colorectal cancer statistics, 2014. CA Cancer J Clin. 64:104–117.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cui S, Gao Y, Zhang K, Chen J, Wang R and
Chen L: The emerging role of inhibitor of growth 4 as a tumor
suppressor in multiple human cancers. Cell Physiol Biochem.
36:409–422. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Guérillon C, Bigot N and Pedeux R: The ING
tumor suppressor genes: Status in human tumors. Cancer Lett.
345:1–16. 2014. View Article : Google Scholar
|
|
5
|
Kim S, Chin K, Gray JW and Bishop JM: A
screen for genes that suppress loss of contact inhibition:
Identification of ING4 as a candidate tumor suppressor gene in
human cancer. Proc Natl Acad Sci USA. 101:16251–16256. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shiseki M, Nagashima M, Pedeux RM,
Kitahama-Shiseki M, Miura K, Okamura S, Onogi H, Higashimoto Y,
Appella E, Yokota J, et al: p29ING4 and p28ING5 bind to p53 and
p300, and enhance p53 activity. Cancer Res. 63:2373–2378.
2003.PubMed/NCBI
|
|
7
|
Xie Y, Zhang H, Sheng W, Xiang J, Ye Z and
Yang J: Adenovirus-mediated ING4 expression suppresses lung
carcinoma cell growth via induction of cell cycle alteration and
apoptosis and inhibition of tumor invasion and angiogenesis. Cancer
Lett. 271:105–116. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gong A, Ye S, Xiong E, Guo W, Zhang Y,
Peng W, Shao G, Jin J, Zhang Z, Yang J, et al: Autophagy
contributes to ING4-induced glioma cell death. Exp Cell Res.
319:1714–1723. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xie Y, Sheng W, Miao J, Xiang J and Yang
J: Enhanced antitumor activity by combining an adenovirus harboring
ING4 with cisplatin for hepatocarcinoma cells. Cancer Gene Ther.
18:176–188. 2011. View Article : Google Scholar :
|
|
10
|
Zhang X, Xu LS, Wang ZQ, Wang KS, Li N,
Cheng ZH, Huang SZ, Wei DZ and Han ZG: ING4 induces G2/M cell cycle
arrest and enhances the chemosensitivity to DNA-damage agents in
HepG2 cells. FEBS Lett. 570:7–12. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang R, Huang J, Feng B, De W and Chen L:
Identification of ING4 (inhibitor of growth 4) as a modulator of
docetaxel sensitivity in human lung adenocarcinoma. Mol Med.
18:874–886. 2012.PubMed/NCBI
|
|
12
|
Zhao Y, Su C, Zhai H, Tian Y, Sheng W,
Miao J and Yang J: Synergistic antitumor effect of
adenovirus-mediated hING4 gene therapy and 125I
radiation therapy on pancreatic cancer. Cancer Lett. 316:211–218.
2012. View Article : Google Scholar
|
|
13
|
Ling C, Xie Y, Zhao D, Zhu Y, Xiang J and
Yang J: Enhanced radiosensitivity of non-small-cell lung cancer
(NSCLC) by adenovirus-mediated ING4 gene therapy. Cancer Gene Ther.
19:697–706. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhu Y, Lv H, Xie Y, Sheng W, Xiang J and
Yang J: Enhanced tumor suppression by an ING4/IL-24 bicistronic
adenovirus-mediated gene cotransfer in human non-small cell lung
cancer cells. Cancer Gene Ther. 18:627–636. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xie Y, Lv H, Sheng W, Miao J, Xiang J and
Yang J: Synergistic tumor suppression by adenovirus-mediated
inhibitor of growth 4 and interleukin-24 gene cotransfer in
hepatocarcinoma cells. Cancer Biother Radiopharm. 26:681–695. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shen JC, Unoki M, Ythier D, Duperray A,
Varticovski L, Kumamoto K, Pedeux R and Harris CC: Inhibitor of
growth 4 suppresses cell spreading and cell migration by
interacting with a novel binding partner, liprin alpha1. Cancer
Res. 67:2552–2558. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li J, Martinka M and Li G: Role of ING4 in
human melanoma cell migration, invasion and patient survival.
Carcinogenesis. 29:1373–1379. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Garkavtsev I, Kozin SV, Chernova O, Xu L,
Winkler F, Brown E, Barnett GH and Jain RK: The candidate tumour
suppressor protein ING4 regulates brain tumour growth and
angiogenesis. Nature. 428:328–332. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ozer A, Wu LC and Bruick RK: The candidate
tumor suppressor ING4 represses activation of the hypoxia inducible
factor (HIF). Proc Natl Acad Sci USA. 102:7481–7486. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Colla S, Tagliaferri S, Morandi F, Lunghi
P, Donofrio G, Martorana D, Mancini C, Lazzaretti M, Mazzera L,
Ravanetti L, et al: The new tumor-suppressor gene inhibitor of
growth family member 4 (ING4) regulates the production of
proangiogenic molecules by myeloma cells and suppresses
hypoxia-inducible factor-1 alpha (HIF-1alpha) activity: Involvement
in myeloma-induced angiogenesis. Blood. 110:4464–4475. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lu M, Pan C, Zhang L, Ding C, Chen F, Wang
Q, Wang K and Zhang X: ING4 inhibits the translation of
proto-oncogene MYC by interacting with AUF1. FEBS Lett.
587:1597–1604. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
You Q, Wang XS, Fu SB and Jin XM:
Downregulated expression of inhibitor of growth 4 (ING4) in
advanced colorectal cancers: A non-randomized experimental study.
Pathol Oncol Res. 17:473–477. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lou C, Jiang S, Guo X and Dong XS: ING4 is
negatively correlated with microvessel density in colon cancer.
Tumour Biol. 33:2357–2364. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative CT method. Nat
Protoc. 3:1101–1108. 2008. View Article : Google Scholar
|
|
25
|
Tiscornia G, Singer O and Verma IM:
Production and purification of lentiviral vectors. Nat Protoc.
1:241–245. 2006. View Article : Google Scholar
|
|
26
|
Weidner N: Current pathologic methods for
measuring intratumoral microvessel density within breast carcinoma
and other solid tumors. Breast Cancer Res Treat. 36:169–180. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Massagué J: G1 cell cycle control and
cancer. Nature. 432:298–306. 2004. View Article : Google Scholar
|
|
28
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Welti J, Loges S, Dimmeler S and Carmeliet
P: Recent molecular discoveries in angiogenesis and antiangiogenic
therapies in cancer. J Clin Invest. 123:3190–3200. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shang B, Cao Z and Zhou Q: Progress in
tumor vascular normalization for anticancer therapy: Challenges and
perspectives. Front Med. 6:67–78. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ferrara N: VEGF and the quest for tumour
angiogenesis factors. Nat Rev Cancer. 2:795–803. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chaffer CL and Weinberg RA: A perspective
on cancer cell metastasis. Science. 331:1559–1564. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lamouille S, Xu J and Derynck R: Molecular
mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell
Biol. 15:178–196. 2014. View Article : Google Scholar : PubMed/NCBI
|