Tight junctions (TJs) are the apical junctional
complex in epithelial and endothelial cells (1). TJs play a key role in maintaining
distinct tissue spaces and regulating paracellular flux of small
molecules and inflammatory cells, as well as promoting
bi-directional signaling between the intracellular and
extracellular compartments (2–4).
Furthermore, tumor differentiation is related to the reduction of
TJs, and tumor cells must overcome the frontline structures of TJs
in order to successfully metastasize (5–7).
The JAM family belongs to the immunoglobulin
subfamily involved in the formation of TJ in most cell types,
including leukocytes, platelets, endothelial and epithelial cells
(2,8–10).
JAMs have been reported to regulate epithelial TJ assembly, and
interact in both a homo- and heterotypic fashion, as well as with
integrins (3,11,12).
The JAM family comprises five members, namely JAM-A, JAM-B, JAM-C,
JAM-4, and JAM-like (13–16). There are many differences between
JAM-B and the other JAM family members. This review will highlight
that which is currently known about the structure, function, and
mechanisms of action of JAM-B and potential areas for future
study.
Members of the JAM family are type I transmembrane
glycoproteins consisting of a short N-terminal signal peptide, two
immunoglobulin-like domains, one transmembrane domain and one
cytoplasmic tail of variable length containing a C-terminal
PDZ-domain domain (4,17–20).
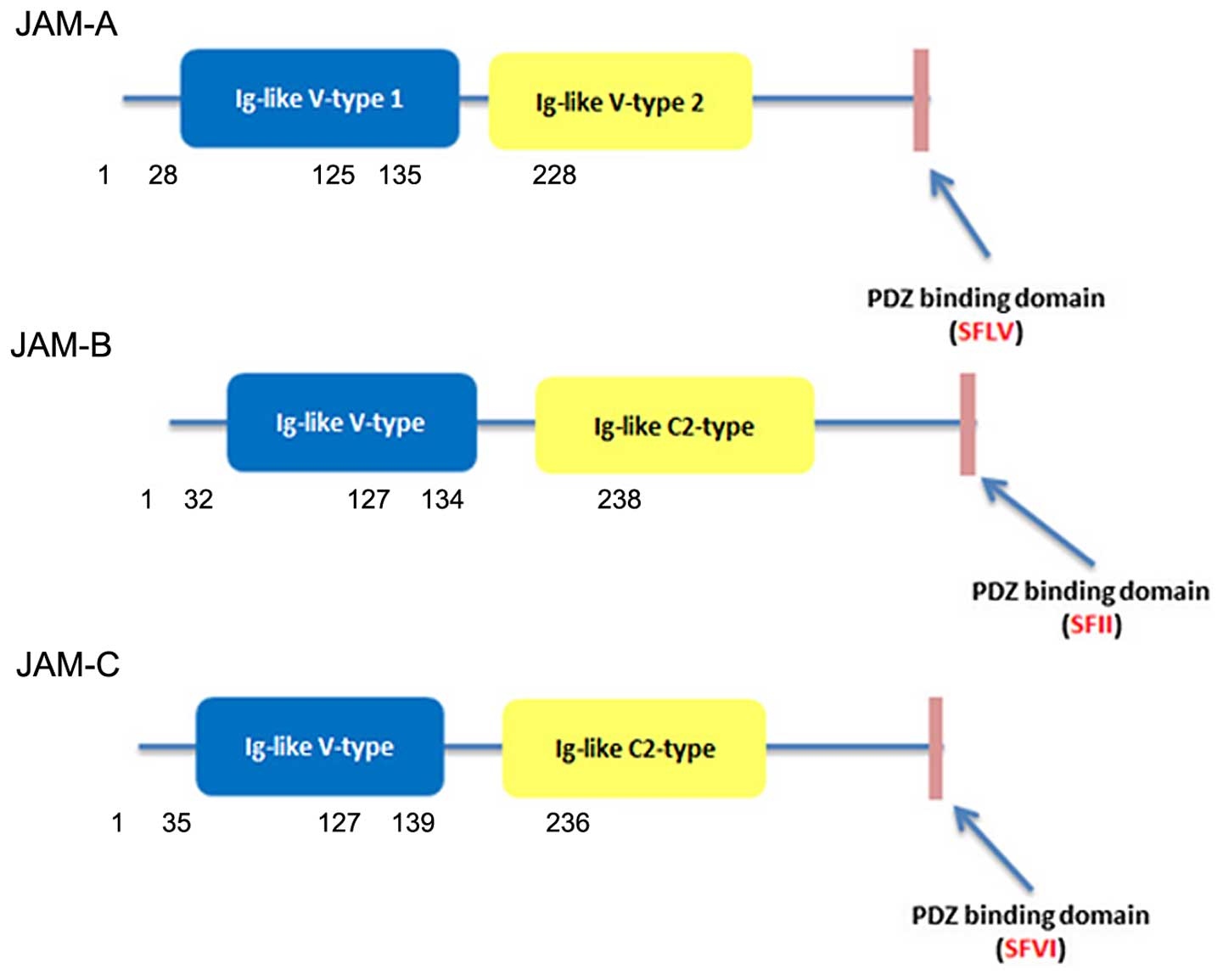
In the case of JAM-A, the sequence was described by multiple
investigators as two C2-type domains in mouse (10) and two V-type domains in human
(2). However, the Ig-like domains
of JAM-B and JAM-C are composed of V type and C2 type (18) (Fig.
1). A J-like sequence is contained in the V domain and an extra
disulfide bridge between cysteines involved incorporated in the
C2-type domain (19).
Some studies have found that JAM-A has a three amino
acid sequence (Val-Leu-Val) in the connection between the two
Ig-like domains which causes the distal Ig-like domains to bend and
bind to another three amino acid sequence (Arg-Leu-Glu) in the V
type Ig-like domains and dimerise (20,21). A
JAM-A dimer at the surface of cells may interact in a trans
orientation with another dimer of JAM2 expressed by the adjacent
cell, forming a tetramer. The successive JAM tetramer would be able
to structure a zipper-like structure surrounding the cell at the
apical cells border. It was found that JAM-B (Arg-Leu-Glu) and
JAM-C (Arg-Ile-Glu) could form the same type of dimerisation motifs
as seen in JAM-A (19).
The JAM family exhibits different expression
profiles and transcript level in various tissues, organs and tumors
(Table I). Overall, as an important
member of the TJ complex, JAMs are generally expressed in subsets
of leukocytes, as well as in epithelial and endothelial cells
(22,23). However, compared with JAM-A and
JAM-C, JAM-B displays a restricted distribution. It is expressed in
the junctions of endothelial cells of different vessels, but mostly
localizes in high endothelial venules (13,25).
Amongst different types of tumors, it is difficult
to quantitatively analyze differences in the expression of the JAM
family. Therefore, the differential expression of JAM-B in cancer
and normal tissues must be compared in a similar tumor type.
Multiple studies have shown that the expression of JAM-B shows
significant differences when comparing normal and cancerous tissues
with the data presented in Table
II.
As reported, classical JAMs, containing an
intracellular PDZ binding domain which may act as an anchor for
scaffolding adaptor proteins, can mediate cellular functions,
embryonic development (26),
junction assembly, spermatogenesis, paracellular permeability,
leukocytic transmigration, angiogenesis, tumor metastasis and cell
proliferation through binding and interacting with receptors such
as ligand and integrin (17,27–33).
The heterodimeric transmembrane receptors, the integrins, contain
α- and β-subunits, which mediate cell-to-cell adhesive
interactions.
There are several studies which report JAM-B
interacting with JAM-C and the integrin α4β1, forming multimer
interactions with integrin counter-receptors. JAM-B can interact
with the integrin α4β1, but this is dependent on the presence of
JAM-C. In 1996, Butcher and Picker described integrin-mediated
T-lymphocyte adhesion inflamed endothelium through VCAM-1 (34). However, whether the integrin could
engage JAM-B and VCAM-1 simultaneously through different binding
sites is still unclear. However, JAM-B and VCAM-1 may interact with
one another laterally and reinforce integrin through conformational
changes (35). Other reports claim
that on human circulating platelets, dendritic cells, NK cells, and
subsets of T and B cells, JAM-C plays an extended role as the
leukocyte counter receptor for JAM-B expressed on endothelial cells
(11,36–40).
In 2009, Ludwig et al found that using a model of
DNFB-induced contact hypersensitivity, JAM-B played a role in
rolling and firm adhesion of T lymphocytes by interacting with α4β1
(41). However, a recent study
proposed JAM-B to play an independent role in leukocyte
transmigration.
JAM-B has been reported involved in many kinds of
biological processes, for instance, leukocyte migration (39,42–44),
spermatogenesis (45) and
angiogenesis (46) through its
interaction with JAM-C. Previous study has shown that blocking the
interactions of JAM-B and -C can reduce monocyte numbers in the
ablumenal compartment by increasing reverse transmigration rather
than through reducing transmigration (47).
The effect of JAM-B and JAM-C in leukocyte
accumulation during inflammation has also been monitored (39,43,48).
For example, a study suggested anti-JAM-B and anti-JAM-C antibodies
played a role of addition agent in blocking leukocytes infiltration
in the model of allergic contact dermatitis, indicating that the
actions of JAM-B and -C are indispensable for development of
cutaneous inflammation (50).
Previous study has shown that blocking the interactions of JAM-B
and -C can reduce monocyte numbers in the ablumenal compartment by
increasing reverse transmigration rather than through reducing
transmigration. Overall, the results indictate that the blocking of
JAM-B/-C interactions decreased the number of monocytes in inflamed
tissues owing to considerably increased level of
reverse-transmigrated monocytes in the peripheral blood.
The cellular localization of JAMs is related to
specific signals of the extracellular environment. Phosphorylation
of the cytoplasmic tail of JAMs plays a key role in the mechanism
of regulating the localization of JAMs. The classical JAM proteins
all contain C-terminal PDZ-binding domains which can be modified by
kinases and may facilitate the interaction with PDZ-containing
proteins, such as TJ-associated cytoplasmic proteins AF6, PAR3,
ZO1, CASK, MAGI and MUPP1 (15,50–54).
In 2003, Ebnet et al showed that JAM-B and JAM-C could
directly associate with PAR-3 through its first PDZ domain, and
could also associate with protein ZO-1 in a PDZ domain-dependent
manner (55). Recently, antibodies
against murine JAM-B inhibited microvessel endothelial cell
proliferation were demonstrated through in vitro endothelial
network formation assays and ex vivo aortic rings. This
research also showed that JAM-B can negatively regulate
pro-angiogenic pathways and inhibit VEGF-induced ERK1/2 activation
through integrin activation in MAPK pathway (56).
Following the investigation of JAM-B protein
involvement in the control of cellular motility and formation of
cellular projections in cancer cells, a number of researchers have
focused on the roles of JAM-B and regulation of this molecule in
cancer cells and tissues. Clinical studies have mentioned different
expression profiles and roles of JAM-B in various types of cancers.
The inconsistency of the results in different cancers has led to an
uncertainty regarding the role of JAM-B in carcinogenesis.
Tumor metastasis is the movement or spreading of
tumor cells from the primary site to a distant one. Circulating
tumor cells, separated from primary tumor, can interact with
vascular endothelial cells during hematogenous metastatic
dissemination. Metastatic and invasive potential of a tumor cell
depends on the expression of various proteins at each step of the
metastatic cascade. Numerous studies showed JAM-B played an
important role in the metastasis of tumor cells (57–59).
Gastric cancer, one of the most common malignancies,
is the second leading cause of cancer-related death globally. Many
patients who had gastric cancer previously could still suffer from
tumor recurrence and metastasis following curative gastrectomy.
Deregulation of the expression and function of TJ proteins leads to
the initiation and progression of cancer by activation of
cytoskeleton mechanism. Hajjari et al examined the changes
in expression levels of genes encoding tight junction-associated
proteins of JAM-B and JAM-C in gastric adenocarcinoma in comparison
with their corresponding marginal normal gastric tissues from the
same patients. They found that JAM-B was upregulated significantly
in tumor samples compared with adjacent normal tissues and was
higher in high grade tumors than in the low grade and intermediate
grade tumors. Moreover, they showed JAM-B and JAM-C activated the
expression of actin filament-associated protein (AFAP) gene as a
downstream factor of JAM-B and JAM-C and this resulted in a
significantly higher expression of AFAP (57). Recently, whole genome and
transcriptome sequencing in primary and peritoneal metastatic
gastric carcinoma indicated JAM-B was only expressed in primary
tumors (58). Overall, deregulation
of JAM-B and JAM-C expression may potentially be involved in
progression of gastric adenocarcinoma tumors. However, JAM-A was
downregulated in gastric cancer tissues compared to adjacent
non-tumor tissues from the same patients, and low JAM-A expression
contributed to poor clinical outcome and increased cell invasion
and migration in gastric cancer (59).
Brain invasion is a biological hallmark of glioma
which contributes to its aggressiveness and prognosis. Deregulated
expression of JAMs in glioma cells has been implicated in this
process. Recently, a study showed that JAM-B was aberrantly
expressed in glioma as a high affinity JAM-C ligand. The
interaction of JAM-B and JAM-C could activate c-Src proto-oncogene,
which is known as a central upstream molecule in the pathways
regulating cell invasion and migration. Also it was shown that
JAM-B/C blocking antibodies impaired in vivo glioma invasion
and proliferation in mice (63).
Magnetic nanoparticle mediated JAM-2 silencing also inhibited the
growth and migration of glioma in vitro and in vivo
(64). A similar study from the
same laboratory also found that the silencing of JAM-2 by
proton-sponge coated quantum dots inhibited the migration of glioma
in vitro through NOTCH pathway blockage (65). In short, targeting of the JAMs
family may provide a potential lead for the treatment of human
gliomas.
Melanoma is the most dangerous type of skin cancer.
Many factors have been proposed to influence the metastasis of
melanoma in patients. Arcangeli et al indicted JAM-B
expressed by endothelial cells was involved in melanoma cell
metastasis via its interaction with JAM-C on tumor cells.
Furthermore, using a JAM-B-deficient mouse model, it was found that
the differential migration disappeared between JAM-C expressing
cells and JAM-C silenced cells, revealing that JAM-B acted as a
counter-receptor for JAM-C mediated reservation of melanoma cells
in the lungs (66).
Esophageal squamous cell carcinoma is an aggressive
and malignant tumor for which there is limited treatment option.
The reason for this is that most cases are diagnosed late, and
patients are already in the advanced stages of the disease. Lymph
node metastasis often arises from this type of cancer, due to the
rich lymphatic drainage system in the esophagus. Recently, it was
demonstrated that the methylation of JAM-B was increased in the
region downstream of the gene and its expression was decreased
(67). Furthermore, a similar
result showed that the expression JAM-B was downregulated and JAM-B
was identified as a key player in the signal transduction networks
in ESCC (68).
In females, breast cancer is the leading cancer with
high incidence and frequently diagnosed malignancy, with lower
survival rates in patients. JAM-B is a necessary component of
apical junctional components. Recently, Coradini et al
reported that mutant p53 tumors of breast cancer were characterized
by a dramatic under-expression of several genes coding for apical
junctional components (69).
Further, a report from Bhan et al stated that JAM-B was
associated with breast cancer progression (70). In 2010, our laboratory showed JAM-B
had a significantly lower level in breast tumors from patients who
developed metastasis than those who were disease-free. We concluded
that JAM-B decreased migration and metastasis of breast cancer
cells. In contrast, JAM-A expression was upregulated in breast
cancer tissues compared to the normal specimen and was correlated
to reduced survival of breast tumor patients, thus JAM-A
overexpression could be a possible mechanism promoting breast
cancer cell migration (71).
Lung cancer is a malignant lung tumor characterized
by uncontrolled cell proliferation in tissues of the lung. JAM-2
was negatively expressed in normal mouse lung, but positively
expressed in spleen and lymph nodes surrounding the lung (72). An interesting discovery showed that
overexpression of JAM-B in a trisomy-21 mouse model of Down's
syndrome was responsible for inhibiting VEGF-induced angiogenesis
and, thus, restraining tumor effects in a lung carcinoma mouse
model (73). Conversely, the
expression level of JAM-A in lung tumor tissues was higher than
corresponding normal lung tissues and served as a negative
predictor of survival in lung cancer patients. Thus, JAM-A
contributes to lung cell growth through cell cycle regulation
(74).
Oral squamous cell carcinoma (OSCC)
characteristically metastasizes from the primary organ to regional
lymph nodes in the early stages of the cancer. It was demonstrated
that JAM-B correlated with the metastatic potential, as it was
upregulated in human oral cancer LNM Tca8113 cells which
metastasized to lymph nodes at an higher rate than its parental
cell line Tca8113 (75). Therefore
it was suggested JAM-B acts as a potential factor for the
metastasis of OSCC.
Recent studies focusing on the interaction of JAMs
have determined multiple homophilic and heterophilic
cis/trans-interactions. Further, regulation of JAM
functions arises by establishing intracellular/extracellular
multiprotein complexes. The family of JAMs shows stability in
expression levels under various conditions, indicating a vital role
under pathological and homeostatic conditions (42,77).
JAMs also play an important role in decreasing epithelial
paracellular permeability. An increasing number of studies has
concentrated on the role of JAMs in tumors. It is clear that JAMs
regulate cell growth, invasion and migration. Perturbed expression
of JAMs has been observed in various malignancy types. The abnormal
expression of certain JAMs is related to disease progression and
poor prognosis. Different JAMs may play contrasting roles in the
same cancer, while the same JAM may undertake converse roles in
different tumor types.
JAM-B interact with various receptors and cytokines
to modulate downstream signal transduction. Most studies concerning
JAM-B showed it does not assume a single role in regulating tumors,
but always with JAM-C-dependence. JAM-B/-C is an important complex
in cell-cell signaling. Its molecular mechanism of action is in
need of further investigation. Many researchers have shown that
JAM-B is an important regulator in various diseases. According to
the above cited studies, the expression level of JAM-B is different
in various tumors compared with normal tissues from the same
patients. Thus, JAM-B may have different potentiality in the
regulation of cancer metastasis. Further, insight into the detailed
mechanisms involved in JAM-B-mediated regulation would be useful in
the understanding its function and role in tumorigenesis and tumor
metastasis. Further investigations will highlight its clinical
significance and application for predicting prognosis. In the
future JAM-B may well provide preoperative diagnosis and a new
avenue for targeted therapy in certain malignancies.
This study was supported by Beijing Municipal
Science and Technology Commission (no. Z151100001615039). HSC was a
recipient of Cardiff university China Medical Scholarship. The
authors wish to than Cancer Research Wales and the Welsh Network of
Life Science.
|
1
|
Runkle EA and Mu D: Tight junction
proteins: From barrier to tumorigenesis. Cancer Lett. 337:41–48.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Martìn-Padura I, Lostaglio S, Schneemann
M, Williams L, Romano M, Fruscella P, Panzeri C, Stoppacciaro A,
Ruco L, Villa A, et al: Junctional adhesion molecule, a novel
member of the immunoglobulin superfamily that distributes at
intercellular junctions and modulates monocyte transmigration. J
Cell Biol. 142:117–127. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Liu Y, Nusrat A, Schnell FJ, Reaves TA,
Walsh S, Pochet M and Parkos CA: Human junction adhesion molecule
regulates tight junction resealing in epithelia. J Cell Sci.
113:2363–2374. 2000.PubMed/NCBI
|
|
4
|
Mandell KJ and Parkos CA: The JAM family
of proteins. Adv Drug Deliv Rev. 57:857–867. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Martin TA, Watkins G, Mansel RE and Jiang
WG: Loss of tight junction plaque molecules in breast cancer
tissues is associated with a poor prognosis in patients with breast
cancer. Eur J Cancer. 40:2717–2725. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Martin TA, Mansel RE and Jiang WG:
Antagonistic effect of NK4 on HGF/SF induced changes in the
transendothelial resistance (TER) and paracellular permeability of
human vascular endothelial cells. J Cell Physiol. 192:268–275.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hoevel T, Macek R, Mundigl O, Swisshelm K
and Kubbies M: Expression and targeting of the tight junction
protein CLDN1 in CLDN1-negative human breast tumor cells. J Cell
Physiol. 191:60–68. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Itoh M, Sasaki H, Furuse M, Ozaki H, Kita
T and Tsukita S: Junctional adhesion molecule (JAM) binds to PAR-3:
A possible mechanism for the recruitment of PAR-3 to tight
junctions. J Cell Biol. 154:491–497. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tajima M, Hirabayashi S, Yao I, Shirasawa
M, Osuga J, Ishibashi S, Fujita T and Hata Y: Roles of
immunoglobulin-like loops of junctional cell adhesion molecule 4;
involvement in the subcellular localization and the cell adhesion.
Genes Cells. 8:759–768. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liang TW, DeMarco RA, Mrsny RJ, Gurney A,
Gray A, Hooley J, Aaron HL, Huang A, Klassen T, Tumas DB, et al:
Characterization of huJAM: Evidence for involvement in cell-cell
contact and tight junction regulation. Am J Physiol Cell Physiol.
279:C1733–C1743. 2000.PubMed/NCBI
|
|
11
|
Liang TW, Chiu HH, Gurney A, Sidle A,
Tumas DB, Schow P, Foster J, Klassen T, Dennis K, DeMarco RA, et
al: Vascular endothelial-junctional adhesion molecule (VE-JAM)/JAM
2 interacts with T, NK, and dendritic cells through JAM 3. J
Immunol. 168:1618–1626. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Martin TA and Jiang WG: Loss of tight
junction barrier function and its role in cancer metastasis.
Biochim Biophys Acta. 1788:872–891. 2009. View Article : Google Scholar
|
|
13
|
Bazzoni G: The JAM family of junctional
adhesion molecules. Curr Opin Cell Biol. 15:525–530. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cunningham SA, Arrate MP, Rodriguez JM,
Bjercke RJ, Vanderslice P, Morris AP and Brock TA: A novel protein
with homology to the junctional adhesion molecule. Characterization
of leukocyte interactions. J Biol Chem. 275:34750–34756. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hirabayashi S, Tajima M, Yao I, Nishimura
W, Mori H and Hata Y: JAM4, a junctional cell adhesion molecule
interacting with a tight junction protein, MAGI-1. Mol Cell Biol.
23:4267–4282. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shin K, Fogg VC and Margolis B: Tight
junctions and cell polarity. Annu Rev Cell Dev Biol. 22:207–235.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ebnet K, Suzuki A, Ohno S and Vestweber D:
Junctional adhesion molecules (JAMs): More molecules with dual
functions? J Cell Sci. 117:19–29. 2004. View Article : Google Scholar
|
|
18
|
Aurrand-Lions M, Johnson-Leger C, Wong C,
Du Pasquier L and Imhof BA: Heterogeneity of endothelial junctions
is reflected by differential expression and specific subcellular
localization of the three JAM family members. Blood. 98:3699–3707.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Garrido-Urbani S, Bradfield PF and Imhof
BA: Tight junction dynamics: The role of junctional adhesion
molecules (JAMs). Cell Tissue Res. 355:701–715. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Prota AE, Campbell JA, Schelling P,
Forrest JC, Watson MJ, Peters TR, Aurrand-Lions M, Imhof BA,
Dermody TS and Stehle T: Crystal structure of human junctional
adhesion molecule 1: Implications for reovirus binding. Proc Natl
Acad Sci USA. 100:5366–5371. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kostrewa D, Brockhaus M, D'Arcy A, Dale
GE, Nelboeck P, Schmid G, Mueller F, Bazzoni G, Dejana E, Bartfai
T, et al: X-ray structure of junctional adhesion molecule:
Structural basis for homophilic adhesion via a novel dimerization
motif. EMBO J. 20:4391–4398. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Naik UP, Ehrlich YH and Kornecki E:
Mechanisms of platelet activation by a stimulatory antibody:
Cross-linking of a novel platelet receptor for monoclonal antibody
F11 with the Fc gamma RII receptor. Biochem J. 310:155–162. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Malergue F, Galland F, Martin F, Mansuelle
P, Aurrand-Lions M and Naquet P: A novel immunoglobulin superfamily
junctional molecule expressed by antigen presenting cells,
endothelial cells and platelets. Mol Immunol. 35:1111–1119. 1998.
View Article : Google Scholar
|
|
24
|
Williams LA, Martin-Padura I, Dejana E,
Hogg N and Simmons DL: Identification and characterisation of human
junctional adhesion molecule (JAM). Mol Immunol. 36:1175–1188.
1999. View Article : Google Scholar
|
|
25
|
Palmeri D, van Zante A, Huang CC,
Hemmerich S and Rosen SD: Vascular endothelial junction-associated
molecule, a novel member of the immunoglobulin superfamily, is
localized to intercellular boundaries of endothelial cells. J Biol
Chem. 275:19139–19145. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kobayashi I, Kobayashi-Sun J, Kim AD,
Pouget C, Fujita N, Suda T and Traver D: JAM1a-JAM2a interactions
regulate haematopoietic stem cell fate through Notch signalling.
Nature. 512:319–323. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bazzoni G and Dejana E: Endothelial
cell-to-cell junctions: Molecular organization and role in vascular
homeostasis. Physiol Rev. 84:869–901. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wegmann F, Ebnet K, Du Pasquier L,
Vestweber D and Butz S: Endothelial adhesion molecule ESAM binds
directly to the multidomain adaptor MAGI-1 and recruits it to cell
contacts. Exp Cell Res. 300:121–133. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Naik MU and Naik UP: Junctional adhesion
molecule-A-induced endothelial cell migration on vitronectin is
integrin alpha v beta 3 specific. J Cell Sci. 119:490–499. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mirza M, Raschperger E, Philipson L,
Pettersson RF and Sollerbrant K: The cell surface protein
coxsackie- and adenovirus receptor (CAR) directly associates with
the Ligand-of-Numb Protein-X2 (LNX2). Exp Cell Res. 309:110–120.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sobocka MB, Sobocki T, Babinska A, Hartwig
JH, Li M, Ehrlich YH and Kornecki E: Signaling pathways of the F11
receptor (F11R; a.k.a. JAM-1, JAM-A) in human platelets: F11R
dimerization, phosphorylation and complex formation with the
integrin GPIIIa. J Recept Signal Transduct Res. 24:85–105. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Reymond N, Garrido-urbani S, Borg JP,
Dubreuil P and Lopez M: PICK-1: A scaffold protein that interacts
with Nectins and JAMs at cell junctions. FEBS Lett. 579:2243–2249.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kansaku A, Hirabayashi S, Mori H, Fujiwara
N, Kawata A, Ikeda M, Rokukawa C, Kurihara H and Hata Y:
Ligand-of-Numb protein X is an endocytic scaffold for junctional
adhesion molecule 4. Oncogene. 25:5071–5084. 2006.PubMed/NCBI
|
|
34
|
Butcher EC and Picker LJ: Lymphocyte
homing and homeostasis. Science. 272:60–66. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Santoso S, Sachs UJH, Kroll H, Linder M,
Ruf A, Preissner KT and Chavakis T: The junctional adhesion
molecule 3 (JAM-3) on human platelets is a counterreceptor for the
leukocyte integrin Mac-1. J Exp Med. 196:679–691. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Moog-Lutz C, Cavé-Riant F, Guibal FC,
Breau MA, Di Gioia Y, Couraud PO, Cayre YE, Bourdoulous S and Lutz
PG: JAML, a novel protein with characteristics of a junctional
adhesion molecule, is induced during differentiation of myeloid
leukemia cells. Blood. 102:3371–3378. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Cunningham SA, Rodriguez JM, Arrate MP,
Tran TM and Brock TA: JAM2 interacts with alpha4beta1. Facilitation
by JAM3. J Biol Chem. 277:27589–27592. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Arrate MP, Rodriguez JM, Tran TM, Brock TA
and Cunningham SA: Cloning of human junctional adhesion molecule 3
(JAM3) and its identification as the JAM2 counter-receptor. J Biol
Chem. 276:45826–45832. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Johnson-Léger CA, Aurrand-Lions M,
Beltraminelli N, Fasel N and Imhof BA: Junctional adhesion
molecule-2 (JAM-2) promotes lymphocyte transendothelial migration.
Blood. 100:2479–2486. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Langer HF, Daub K, Braun G, Schönberger T,
May AE, Schaller M, Stein GM, Stellos K, Bueltmann A, Siegel-Axel
D, et al: Platelets recruit human dendritic cells via Mac-1/JAM-C
interaction and modulate dendritic cell function in vitro.
Arterioscler Thromb Vasc Biol. 27:1463–1470. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ludwig RJ, Hardt K, Hatting M, Bistrian R,
Diehl S, Radeke HH, Podda M, Schön MP, Kaufmann R, Henschler R, et
al: Junctional adhesion molecule (JAM)-B supports lymphocyte
rolling and adhesion through interaction with alpha4beta1 integrin.
Immunol. 128:196–205. 1990. View Article : Google Scholar
|
|
42
|
Keiper T, Al-Fakhri N, Chavakis E,
Athanasopoulos AN, Isermann B, Herzog S, Saffrich R, Hersemeyer K,
Bohle RM, Haendeler J, et al: The role of junctional adhesion
molecule-C (JAM-C) in oxidized LDL-mediated leukocyte recruitment.
FASEB J. 19:2078–2080. 2005.PubMed/NCBI
|
|
43
|
Aurrand-Lions M, Lamagna C, Dangerfield
JP, Wang S, Herrera P, Nourshargh S and Imhof BA: Junctional
adhesion molecule-C regulates the early influx of leukocytes into
tissues during inflammation. J Immunol. 174:6406–6415. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lamagna C, Meda P, Mandicourt G, Brown J,
Gilbert RJ, Jones EY, Kiefer F, Ruga P, Imhof BA and Aurrand-Lions
M: Dual interaction of JAM-C with JAM-B and alpha(M)beta2 integrin:
Function in junctional complexes and leukocyte adhesion. Mol Biol
Cell. 16:4992–5003. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gliki G, Ebnet K, Aurrand-Lions M, Imhof
BA and Adams RH: Spermatid differentiation requires the assembly of
a cell polarity complex downstream of junctional adhesion
molecule-C. Nature. 431:320–324. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Lamagna C, Hodivala-Dilke KM, Imhof BA and
Aurrand-Lions M: Antibody against junctional adhesion molecule-C
inhibits angiogenesis and tumor growth. Cancer Res. 65:5703–5710.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Bradfield P, Scheiermann C, Nourshargh S,
Ody C, Luscinskas F, Rainger G, et al: JAM-C, a turnstile for
monocyte transendothelial migration in inflammation. Swiss Med
Wkly. 137:12s2007.
|
|
48
|
Vonlaufen A, Aurrand-Lions M, Pastor CM,
Lamagna C, Hadengue A, Imhof BA and Frossard JL: The role of
junctional adhesion molecule C (JAM-C) in acute pancreatitis. J
Pathol. 209:540–548. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Ludwig RJ, Zollner TM, Santoso S, Hardt K,
Gille J, Baatz H, Johann PS, Pfeffer J, Radeke HH, Schön MP, et al:
Junctional adhesion molecules (JAM)-B and -C contribute to
leukocyte extravasation to the skin and mediate cutaneous
inflammation. J Invest Dermatol. 125:969–976. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Hamazaki Y, Itoh M, Sasaki H, Furuse M and
Tsukita S: Multi-PDZ domain protein 1 (MUPP1) is concentrated at
tight junctions through its possible interaction with claudin-1 and
junctional adhesion molecule. J Biol Chem. 277:455–461. 2002.
View Article : Google Scholar
|
|
51
|
Martinez-Estrada OM, Villa A, Breviario F,
Orsenigo F, Dejana E and Bazzoni G: Association of junctional
adhesion molecule with calcium/calmodulin-dependent serine protein
kinase (CASK/LIN-2) in human epithelial caco-2 cells. J Biol Chem.
276:9291–9296. 2001. View Article : Google Scholar
|
|
52
|
Ebnet K, Schulz Cu, Meyer Zu, Brickwedde
MK, Pendl GG and Vestweber D: Junctional adhesion molecule
interacts with the PDZ domain-containing proteins AF-6 and ZO-1. J
Biol Chem. 275:27979–27988. 2000.PubMed/NCBI
|
|
53
|
Bazzoni G, Martinez-Estrada OM, Orsenigo
F, Cordenonsi M, Citi S and Dejana E: Interaction of junctional
adhesion molecule with the tight junction components ZO-1,
cingulin, and occludin. J Biol Chem. 275:20520–20526. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Ebnet K, Suzuki A, Horikoshi Y, Hirose T,
Meyer Zu, Brickwedde MK, Ohno S and Vestweber D: The cell polarity
protein ASIP/PAR-3 directly associates with junctional adhesion
molecule (JAM). EMBO J. 20:3738–3748. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Ebnet K, Aurrand-Lions M, Kuhn A, Kiefer
F, Butz S, Zander K, Meyer zu Brickwedde MK, Suzuki A, Imhof BA and
Vestweber D: The junctional adhesion molecule (JAM) family members
JAM-2 and JAM-3 associate with the cell polarity protein PAR-3: A
possible role for JAMs in endothelial cell polarity. J Cell Sci.
116:3879–3891. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Meguenani M, Miljkovic-Licina M, Fagiani
E, Ropraz P, Hammel P, Aurrand-Lions M, Adams RH, Christofori G,
Imhof BA and Garrido-Urbani S: Junctional adhesion molecule B
interferes with angiogenic VEGF/VEGFR2 signaling. FASEB J.
29:3411–3425. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Hajjari M, Behmanesh M, Sadeghizadeh M and
Zeinoddini M: Junctional adhesion molecules 2 and 3 may potentially
be involved in progression of gastric adenocarcinoma tumors. Med
Oncol. 30:3802013. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Zhang J, Huang JY, Chen YN, Yuan F, Zhang
H, Yan FH, Wang MJ, Wang G, Su M, Lu G, et al: Whole genome and
transcriptome sequencing of matched primary and peritoneal
metastatic gastric carcinoma. Sci Rep. 5:137502015. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Huang JY, Xu YY, Sun Z, Wang ZN, Zhu Z,
Song YX, Luo Y, Zhang X and Xu HM: Low junctional adhesion molecule
A expression correlates with poor prognosis in gastric cancer. J
Surg Res. 192:494–502. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Kok-Sin T, Mokhtar NM, Ali Hassan NZ,
Sagap I, Mohamed Rose I, Harun R and Jamal R: Identification of
diagnostic markers in colorectal cancer via integrative epigenomics
and genomics data. Oncol Rep. 34:22–32. 2015.PubMed/NCBI
|
|
61
|
Oster B, Thorsen K, Lamy P, Wojdacz TK,
Hansen LL, Birkenkamp-Demtröder K, Sørensen KD, Laurberg S, Orntoft
TF and Andersen CL: Identification and validation of highly
frequent CpG island hypermethylation in colorectal adenomas and
carcinomas. Int J Cancer. 129:2855–2866. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Bujko M, Kober P, Mikula M, Ligaj M,
Ostrowski J and Siedlecki JA: Expression changes of cell-cell
adhesion-related genes in colorectal tumors. Oncol Lett.
9:2463–2470. 2015.PubMed/NCBI
|
|
63
|
Tenan M, Aurrand-Lions M, Widmer V,
Alimenti A, Burkhardt K, Lazeyras F, Belkouch MC, Hammel P, Walker
PR, Duchosal MA, et al: Cooperative expression of junctional
adhesion molecule-C and -B supports growth and invasion of glioma.
Glia. 58:524–537. 2010.
|
|
64
|
Qi LF, Liu J, Zhu HY, Li ZQ, Lu K, Li T
and Shi D: Inhibition of glioma proliferation and migration by
magnetic nanoparticle mediated JAM-2 silencing. J Mater Chem B
Mater Biol Med. 2:7168–7175. 2014. View Article : Google Scholar
|
|
65
|
Qi LF, Shao WJ and Shi DL: JAM-2 siRNA
intracellular delivery and real-time imaging by proton-sponge
coated quantum dots. J Mater Chem B Mater Biol Med. 1:654–660.
2013. View Article : Google Scholar
|
|
66
|
Arcangeli ML, Frontera V, Bardin F,
Thomassin J, Chetaille B, Adams S, Adams RH and Aurrand-Lions M:
The junctional adhesion molecule-B regulates JAM-C-dependent
melanoma cell metastasis. FEBS Lett. 586:4046–4051. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Li X, Wu Z, Mei Q, Li X, Guo M, Fu X and
Han W: Long non-coding RNA HOTAIR, a driver of malignancy, predicts
negative prognosis and exhibits oncogenic activity in oesophageal
squamous cell carcinoma. Br J Cancer. 109:2266–2278. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Xu CQ, Zhu ST, Wang M, Guo SL, Sun XJ,
Cheng R, Xing J, Wang WH, Shao LL and Zhang ST: Pathway analysis of
differentially expressed genes in human esophageal squamous cell
carcinoma. Eur Rev Med Pharmacol Sci. 19:1652–1661. 2015.PubMed/NCBI
|
|
69
|
Coradini D, Fornili M, Ambrogi F, Boracchi
P and Biganzoli E: TP53 mutation, epithelial-mesenchymal
transition, and stemlike features in breast cancer subtypes. J
Biomed Biotechnol. 2012:2540852012. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Bhan A, Hussain I, Ansari KI, Kasiri S,
Bashyal A and Mandal SS: Antisense transcript long noncoding RNA
(lncRNA) HOTAIR is transcriptionally induced by estradiol. J Mol
Biol. 425:3707–3722. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
McSherry EA, McGee SF, Jirstrom K, Doyle
EM, Brennan DJ, Landberg G, Dervan PA, Hopkins AM and Gallagher WM:
JAM-A expression positively correlates with poor prognosis in
breast cancer patients. Int J Cancer. 125:1343–1351. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Singh B, Tschernig T, van Griensven M,
Fieguth A and Pabst R: Expression of vascular adhesion protein-1 in
normal and inflamed mice lungs and normal human lungs. Virchows
Arch. 442:491–495. 2003.PubMed/NCBI
|
|
73
|
Reynolds LE, Watson AR, Baker M, Jones TA,
D'Amico G, Robinson SD, Joffre C, Garrido-Urbani S,
Rodriguez-Manzaneque JC, Martino-Echarri E, et al: Tumour
angiogenesis is reduced in the Tc1 mouse model of Down's syndrome.
Nature. 465:813–817. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Zhang M, Luo W, Huang B, Liu Z, Sun L,
Zhang Q, Qiu X, Xu K and Wang E: Overexpression of JAM-A in
non-small cell lung cancer correlates with tumor progression. PLoS
One. 8:e791732013. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Zhuang Z, Jian P, Longjiang L, Bo H and
Wenlin X: Oral cancer cells with different potential of lymphatic
metastasis displayed distinct biologic behaviors and gene
expression profiles. J Oral Pathol Med. 39:168–175. 2010.
View Article : Google Scholar
|
|
76
|
Zhang Y, Zhang H, Huang Y, Sun R, Liu R
and Wei J: Human leukocyte antigen (HLA)-C polymorphisms are
associated with a decreased risk of rheumatoid arthritis. Mol Biol
Rep. 41:4103–4108. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Harita Y, Miyauchi N, Karasawa T, Suzuki
K, Han GD, Koike H, Igarashi T, Shimizu F and Kawachi H: Altered
expression of junctional adhesion molecule 4 in injured podocytes.
Am J Physiol Renal Physiol. 290:F335–F344. 2006. View Article : Google Scholar
|