Introduction
The human skin is the most exposed organ of the
human body and protects against diverse environmental injuries.
Meanwhile, human skin is also at risk of continuous and repetitive
environmental damage. Solar ultraviolet (UVA, UVB and UVC)
radiation is a sort of high-energy electromagnetic radiation that
is thought to have xenotoxic effects on all living organisms.
Particularly UVB (280–320 nm), which is the main component of solar
UV, is incapable of penetrating deeply into the skin and it only
affects the epidermis, the superficial layer of the skin, which is
composed predominantly of keratinocytes (1). UVB irradiation of the skin can result
in a variety of injuries and leads to various skin diseases
(2). It is reported that UVB
irradiation has been shown to induce a transient increase in the
intracellular level of reactive oxygen species (ROS) in human skin,
particularly in keratinocytes (HaCaT) (3–9). These
reactive oxygen species (ROS), including hydroxyl radicals,
superoxide radicals, peroxyl radical and their active precursors
namely singlet oxygen, hydrogen peroxide and ozone (10), attack the cell membrane and cellular
organelles resulting in lipid peroxidation and organelle damage.
UVB irradiation of HaCaT cells results in a
post-irradiation-dependent increase in ROS and subsequent cell
membrane fluidity decrease and mitochondrial membrane
depolarization (1). Plasma membrane
fluidity affects a number of cellular functions, such as
carrier-mediated transport, the properties of certain
membrane-bound enzymes and cell growth. Membrane fluidity decrease
has deleterious effects on cells and triggers apoptosis (11). Mitochondria are both the major
producers and targets of ROS in mammalian cells (12). UVB-induced ROS can attack
mitochondria and result in mitochondrial structure change and the
collapse of mitochondrial membrane potential (ΔΨm) of mitochondria
and the subsequent outflow of apoptosis-inducing factors and
apoptosis. UVB irradiation-induced ROS play an important role in
the occurrence of apoptosis (10)
and solar radiation is a crucial factor that deeply affects this
specialized epithelium (13) and
skin diseases.
Thus, reducing intracellular ROS levels may
represent an effective strategy for preventing UVB-induced HaCaT
apoptosis and skin damage. In recent years, naturally occurring
compounds, flavonoids, which are abundant in fruits, vegetables,
green tea, and red wine and possess a variety of biological
activities including antioxidant, have gained considerable
attention as protective agents against UVB-induced skin damage via
scavenging ROS. Quercetin is one of the flavonoids derived from
numerous fruits and vegetables with high antioxidant action and
provides protection against UV-induced damage to plants. Studies
have reported that quercetin has diverse pharmacological
activities, and the protective effects of quercetin have been
attributed to the inhibition of key signaling enzymes involved in
the regulation of cell proliferation, angiogenesis and apoptosis
(14). In addition, quercetin has
strong antioxidant activity and can scavenge free radicals and
inhibit lipid peroxidation and metal ion chelation (15–22).
Due to the antioxidant properties of quercetin, we hypothesized
that quercetin may provide protection against ROS attack on the
cell membrane and mitochondria and block lipid peroxidation and
release of apoptosis-inducing factors from mitochondria via ROS
clearing activity.
In the present study, we investigated the protective
effect of quercetin against ROS-induced cell membrane and
mitochondrial damage in UVB-irradiated HaCaT cells. The results
showed that quercetin effectively cleared UVB irradiation-induced
ROS in the HaCaT cells and blocked ROS-induced lipid peroxidation,
mitochondrial membrane depolarization and subsequent outflow of
apoptosis-inducing factors and apoptosis.
Materials and methods
Materials
Quercetin was purchased from Sigma Aldrich (St.
Louis, MO, USA). 4′, 6-Diamidino-2-phenylindole dihydrochloride
(DAPI) was purchased from Sangon Biotech (Shanghai, China).
DCFH-DA, Dio, JC-1 were purchased from Beyotime Biotechnology
(Shanghai, China). All other chemicals and reagents of the highest
quality were commercially available and used as received.
Antibodies against cytochrome c, Bcl-2, Bax, caspase-3 and
PARP were purchased from Cell Signaling Technology Inc. (Beverly,
MA, USA).
Cell culture and UVB irradiation
Human skin keratinocytes, HaCaT, were cultured in
Dulbecco's modified Eagle's medium (DMEM; Gibco BRL, Grand Island,
NY, USA) supplemented with 10% heat-inactivated fetal bovine serum
(FBS; Biological Industries Ltd., Israel) at 37°C in 5%
CO2. The cells were incubated with or without quercetin
for 8 h prior to UVB irradiation. Then, the cells were washed with
phosphate buffered saline (PBS; pH 7.4) and irradiated using a
microprocessor-controlled UV Crosslinker (XL-1000; Spectrolinker™,
Westbury, NY, USA). The irradiation intensity was monitored by a
UVB radiometer (Beijing Normal University, China). Immediately
after UVB irradiation, the cells were returned to the incubator and
incubated with drug-free media for the indicated times.
Assessment of cell viability
The cell viability was determined using MTT assay.
The cells were seeded in 96-well plates at a concentration of 5,000
cells/ml. The cells underwent pretreatments with quercetin mixed
with dimethyl sulfoxide (DMSO) at various concentrations. Before
irradiation, the cells were incubated for 18 h and then, after
irradiation, the cells were incubated for the indicated times.
Following this, 20 μl MTT (2 mg/ml) was added, and the cells were
incubated for another 4 h. Finally, 200 μl DMSO was added to
dissolve the crystals, and the plates were read immediately on a
plate reader at a test wavelength of 490 nm.
Measurement of intracellular ROS
levels
Cells grown on coverslips in 6-well plates were
exposed to the indicated treatments. Quercetin was applied, and the
incubation time after irradiation was 24 h. Cells were incubated
with DCFH-DA according to the instructions and immediately examined
by confocal laser scanning microscopy (Olympus FV1000-IX81, Tokyo,
Japan) using an argon laser with a 488-nm excitation band. The
laser intensity, pinhole diameter and photomultiplier settings were
kept constant for every experiment. For quantitative analysis, the
ROS levels were examined using a Partec PAS III flow cytometer
(Partec, Munster, Germany).
Fluorescence recovery after
photobleaching (FRAP)
FRAP analysis was performed as previously described
(23–25). Briefly, after incubation with
quercetin for 8 h, the cells were irradiated with UVB (15
mJ/cm2). Twelve hours after irradiation, the cells were
incubated with the Dio probe according to the protocol, and
photobleaching was immediately conducted by confocal laser scanning
microscopy (FV1000-IX81; Olympus, Tokyo, Japan) (with excitation
band 488 nm, absorption band 530 nm, bleaching band 405 nm, power
91%, time 1 sec).
R(recovery)=(F1–F0)/(F2–F0)x100%D(diffusioncoefficients)=βω2/4t1/2;
where F2 is the fluorescence intensity after the
recovery, F1 is the the fluorescence intensity before the
photobleaching, F0 is the fluorescence intensity immediately after
the photobleaching, β is the conversion factor, ω is the radius of
the bleached area and t1/2 is the half time
(t1/2) of recovery into the bleached region.
Measurement of intracellular
malondialdehyde (MDA) level
Cells were seeded in 6-well plates at a
concentration of 1×107 cells/ml. After the cells had
undergone the indicated treatments, quercetin was applied, and the
incubation time after irradiation was 24 h. The MDA levels were
estimated using the commercially available colorimetric MDA-586
assay kit from Beyotime Biotechnology. The absorbance was
determined using a microplate reader (ELX808; Bio-Tek Instruments,
Inc., Winooski, VT, USA) at a wavelength of 586 nm. The protein
content was measured using a BCA assay kit from Beyotime
Biotechnology.
Detection of mitochondrial membrane
depolarization
Cells were seeded into 6-well plates until reaching
80% confluency. Quercetin was applied, and the incubation time
after irradiation was 24 h. Cells were stained with JC-1 according
to the manufacturer's protocol and were immediately examined by
confocal laser scanning microscopy (FV1000-IX81; Olympus). The
laser intensity, pinhole diameter and photomultiplier settings were
kept constant for every experiment. For quantitative analysis, the
ΔΨm was examined using a Partec PAS III flow cytometer
(Partec).
Immunocytochemical staining
Cells were seeded in a 12-well plate with a cover
glass and incubated at 37°C with 5% CO2 overnight.
Quercetin was added to the medium and incubated for 8 h. Then the
cells were washed and irradiated with the indicated irradiation
intensity. Twenty-four hours after irradiation, the cells were
fixed, permeabilized and the immunofluorescence staining with
anti-Tom antibody was performed.
Immunoblotting
Immunoblot analysis was performed as previously
described (26). Briefly, the cell
lysates were resolved on 12% SDS-PAGE and analyzed by
immunoblotting using cytochrome c, Bcl-2, Bax, caspase-3 and
PARP antibodies, followed by enhanced chemiluminescence (ECL)
detection (Thermo Scientific).
Statistical methodology
The data are presented as mean ± SE. Significant
differences between two groups were analyzed with the Student's
t-test. One-way analysis of variance (ANOVA) was applied to analyze
differences in data of biochemical parameters among the
experimental groups, followed by Dunnett's test for pair-wise
multiple comparisons. P<0.05 was considered to indicate a
statistically significant difference.
Results and Discussion
Quercetin increases the viability of
UVB-treated HaCaT cells
Quercetin is known to possess antioxidant properties
due to the o-hydroxy structure in the B ring, the 2–3 double bond
in conjugation with the 4-oxo function in the C-ring and the 3- and
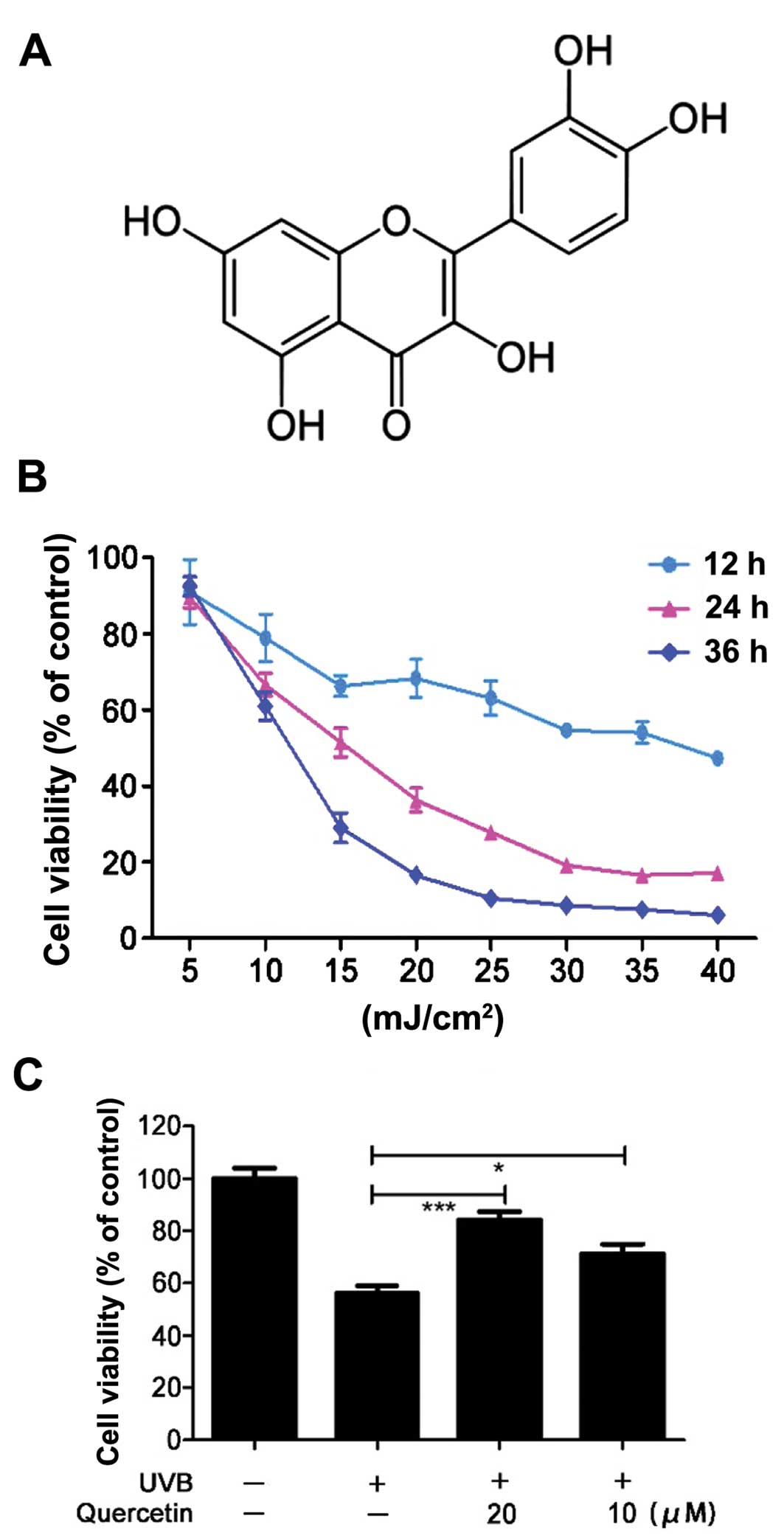
5-OH groups with the 4-oxo function in the A and C rings (Fig. 1A) (27). It is reported that quercetin reduces
cell death in cutaneous tissue-associated cell types (28). Thus, we investigated the protective
effect of quercetin against UVB irradiation-induced damage in HaCaT
cells. The cell viability (% of the control) in the UVB-irradiated
HaCaT cells was evaluated by MTT assay. UVB irradiation caused cell
death in a time and dose-dependent manner (Fig. 1B). Pretreatment with quercetin
significantly increased cell viability 24 h post-irradiation at the
dose of 15 mJ/cm2 (Fig.
1C). This demonstrated that quercetin could rescue HaCaT cells
from UVB irradiation-induced cell injury. As UVB radiation is known
to have deleterious effects on cells via ROS (29,30),
we hypothesized that one of the possible mechanisms for the
protective effect of quercetin could be attributed to ROS
scavenging capacity.
Quercetin reduces UVB
irradiation-induced intracellular ROS levels
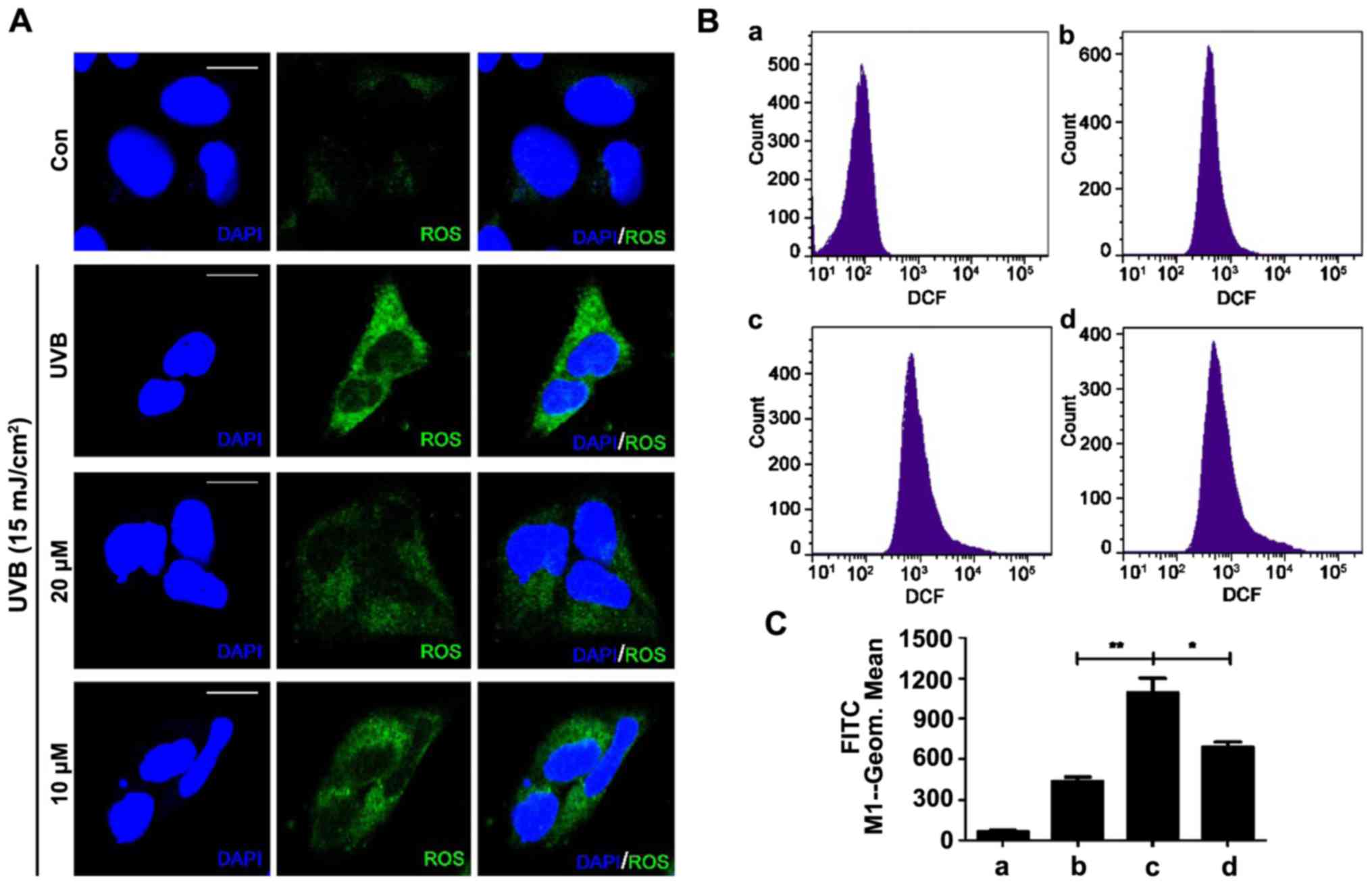
To investigate whether quercetin reduces UVB-induced
ROS generation, a fluorescence assay using DCFH-DA was performed.
DCFH-DA is hydrolyzed by intracellular esterase and is converted to
nonfluorescent DCFH, which is oxidized to highly fluorescent DCF in
the presence of ROS (10,31). The fluorescence intensity is a
measurement of intracellular ROS activity. The qualitative
assessment of the cellular level of ROS was conducted by confocal
laser scanning microscopy. UVB irradiation of 15 mJ/cm2
significantly increased green fluorescence intensity (Fig. 2A). When the cells were pretreated
with quercetin, the DCF fluorescence intensity was clearly reduced
and the group treated with 20 µM quercetin had a weaker
fluorescence than the 10 µM quercetin-treated group. Fig. 2B shows the quantitative analysis of
the cellular ROS levels and the bar graph in Fig. 2C displays the fluorescence intensity
of the flow cytometric results. The results demonstrated that UVB
irradiation caused intracellular ROS generation and quercetin
effectively reduced the intracellular ROS generated by UVB
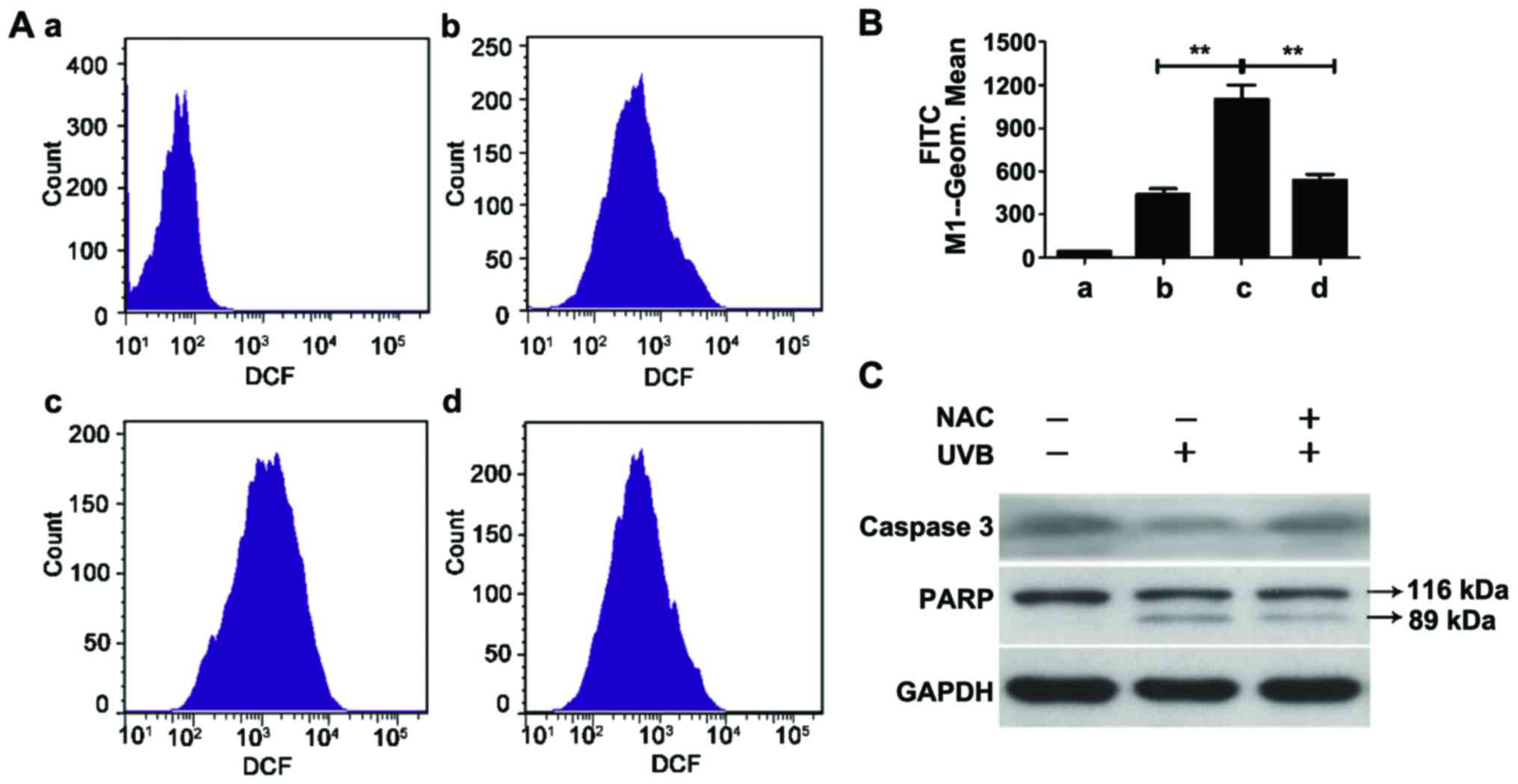
irradiation. Furthermore, N-acetyl-cysteine (NAC), an effective ROS
clearing agent, effectively scavenged UVB-induced ROS in the HaCaT
cells (Fig. 3A and B).
Quercetin prevents cells from
ROS-induced membrane fluidity decrease and lipid peroxidation
UVB irradiation-induced ROS may attack the cell
membrane and result in lipid peroxidation and cell membrane
fluidity decrease. As quercetin effectively reduced UVB
irradiation-induced ROS generation, we hypothesized that quercetin
prevents cells from ROS-induced membrane fluidity decrease and
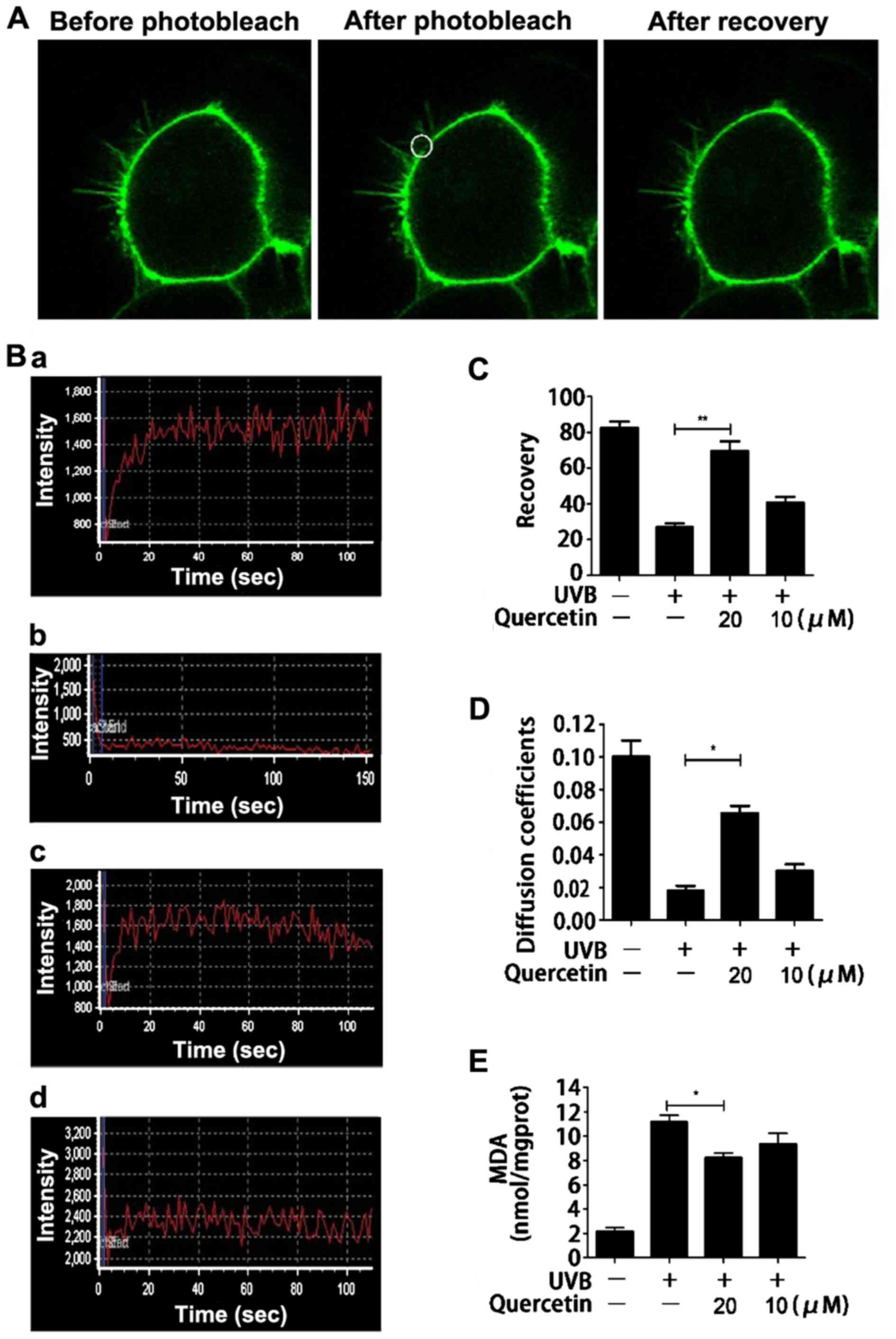
lipid peroxidation. FRAP is a technique that monitors the diffusion
of fluorescent molecules into a region in which fluorophores have
been irreversibly photobleached by a high intensity laser pulse. A
fluorescent probe (Dio) was used to stain the cell membrane, and
the membrane fluidity was detected via lateral diffusion by
monitoring the recovery of fluorescence into a region following a
single photobleaching event. The fluorescence intensity before the
photobleaching, the fluorescence intensity immediately after
photobleaching and the fluorescence intensity after the recovery,
respectively are shown in Fig. 4A.
The fluorescence intensity curves represent the membrane fluidity
of the cells (Fig. 4B). UVB
irradiation significantly reduced cell membrane fluidity, and
pretreatment with quercetin at the concentration of 20 μM markedly
rescued cell membrane fluidity. The concentration of 20 µM resulted
in a more effective protective effect than the concentration of 10
µM. Fig. 4C and D show the recovery
(R) and effective diffusion coefficient (D) of the cell membrane
which suggested that quercetin had an obvious protective effect
against ROS-induced cell membrane fluidity decrease.
Then, we further detected the lipid peroxidation of
the cell membrane, which is the most common consequence of
oxidative stress. MDA is a common indicator with which to determine
the degree of lipid peroxidation. It is the final product of lipid
peroxidation and is toxic to cells and the cell membrane (10,32).
Fig. 4E shows that UVB exposure
induced an apparent increase in MDA and pretreatment with quercetin
clearly inhibited the increase. The highest protective effect of
quercetin was obtained at the concentration of 20 μM. The results
suggest that quercetin effectively prevents cells from ROS-induced
membrane fluidity decrease and lipid peroxidation.
Quercetin protects cells from
mitochondrial damage
UVB irradiation induces a transient increase in the
intracellular level of ROS, and the increased levels of ROS can
attack the cell membrane and organelles, such as the mitochondria.
Quercetin effectively prevented cells from ROS-induced cell
membrane damage. Then, we investigated whether quercetin had a
protective effect on preventing ROS-induced mitochondrial
damage.
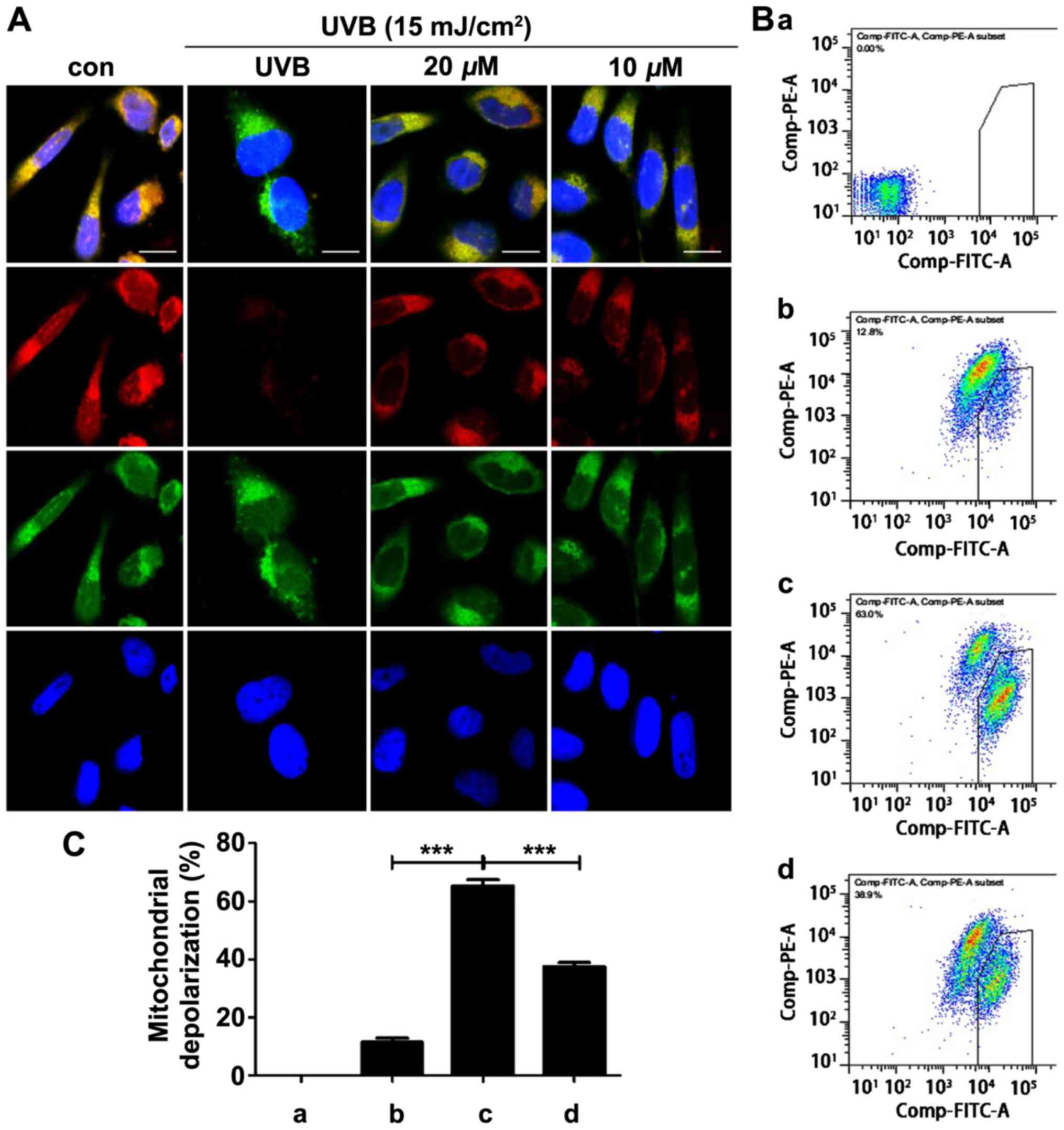
The integrity of ΔΨm was assessed using JC-1
fluorescence staining and visualized via fluorescence microscopy.
The red fluorescence decreased and the green fluorescence
correspondingly increased after the UVB irradiation, indicating the
disruption of ΔΨm and mitochondrial dysfunction (Fig. 5A). Pretreatment with quercetin
markedly rescued mitochondrial membrane depolarization. Fig. 5B shows the quantitative analysis of
ΔΨm and the bar graph in Fig. 5C
displays the values of the flow cytometric results.
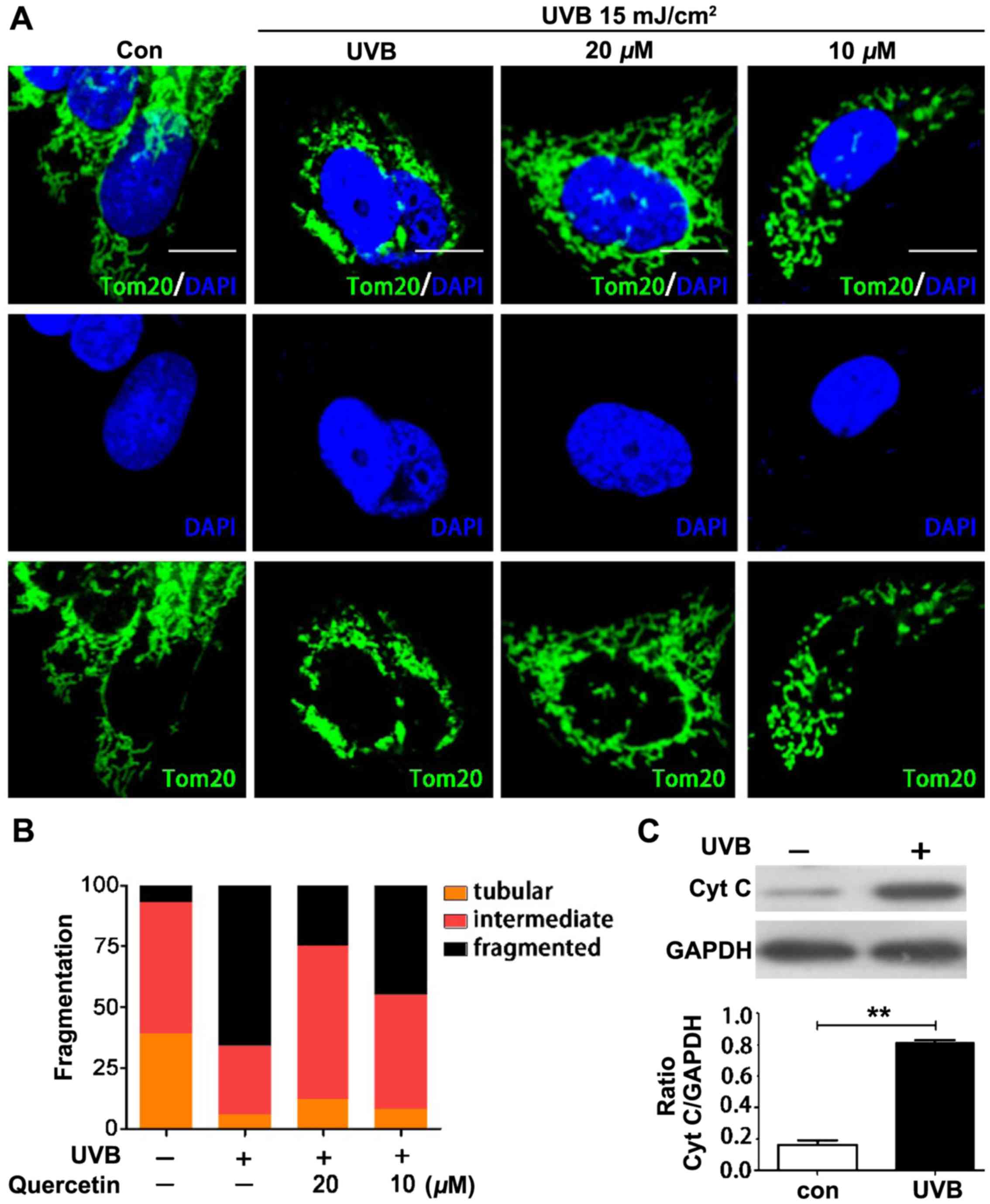
We further assessed the potential effect of ROS on
mitochondrial morphology and the effect of quercetin on
mitochondrial morphologic change. Tom20 was used to identify
mitochondrial morphology in each cell. Cells were classified into
three groups. Cells classified as having tubular mitochondria
contained almost entirely mitochondria with a length/width (axis)
ratio >10. Cells classified as having fragmented mitochondria
contained mitochondria with an axis ratio <3, and cells
classified as having intermediate mitochondria contained both
tubular and fragmented mitochondria (10,33).
Fig. 6A represents the morphology
of the mitochondria in the differently treated cells. UVB
irradiation interrupted tubular mitochondria and increased the
numbers of fragmented mitochondria. Pretreatment with quercetin
blocked ROS-induced mitochondrial fragmentation. Fig. 6B shows the numbers of cells with
different mitochondrial morphology. The results suggest that
quercetin protected cells from ROS-induced mitochondrial
morphological change. Then, we further investigated whether the
mitochondrial fragmentation accompanied the outflow of
apoptosis-inducing factors, such as cytochrome c. Fig. 6C shows that the release of
cytochrome c occurred in the mitochondrial fragmented
cells.
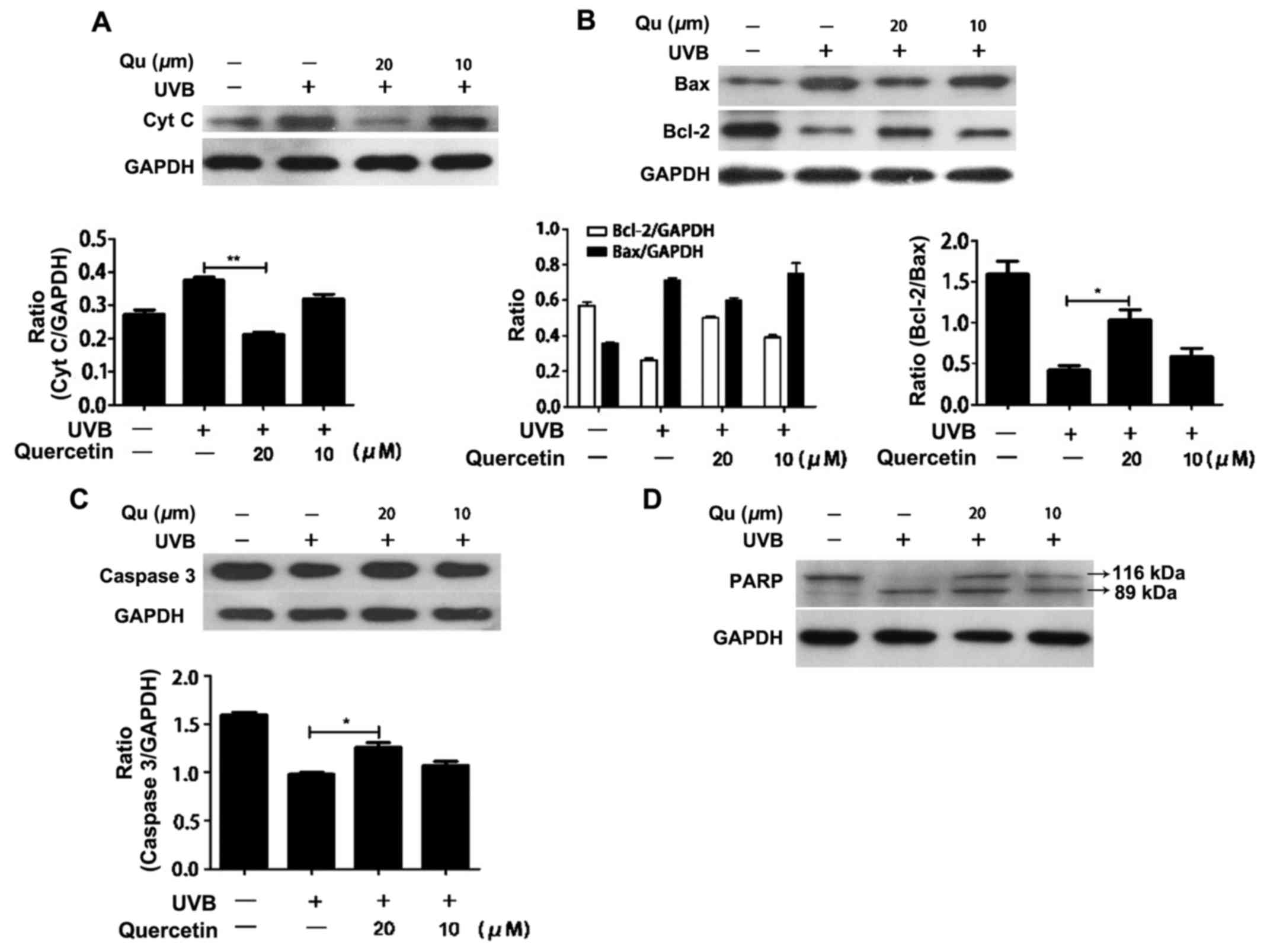
Quercetin blocks the release of
cytochrome c and inhibits apoptosis in the UVB-irradiated
cells
UVB irradiation increases intracellular ROS levels
and the increased ROS results in mitochondrial membrane
depolarization, mitochondrial morphological change and the outflow
of cytochrome c from the mitochondria. The release of
cytochrome c into the cytosol can subsequently activates the
caspase cascade that results in apoptosis. Thus, we examined the
effect of quercetin on cytochrome c release. Quercetin
markedly inhibited the outflow of cytochrome c from the
mitochondria (Fig. 7A). Next, we
examined the effect of quercetin on the expression of Bcl-2 family
proteins, which play an important role in the regulation of
mitochondrial permeability, in the UVB-irradiated HaCaT cells.
Western blot analysis revealed that pretreatment with quercetin
increased the level of Bcl-2 and decreased the level of Bax in the
UVB-irradiated HaCaT cells. Therefore, the ratio of Bax to Bcl-2
was increased (Fig. 7B). The
caspase family is believed to play a central role in mediating
various apoptotic responses. To confirm the induction of the
apoptotic pathway, we examined the activation of caspase-3 and
PARP. Pretreatment with quercetin induced a significant increase in
caspase-3 after UVB irradiation (Fig.
7C and D). PARP, which is a substrate of caspase-3, was cleaved
in the quercetin-pretreated cells in response to UVB irradiation.
These findings suggest that quercetin protects cells against UVB
irradiation-induced cell death via the mitochondrial-mediated
apoptotic pathway.
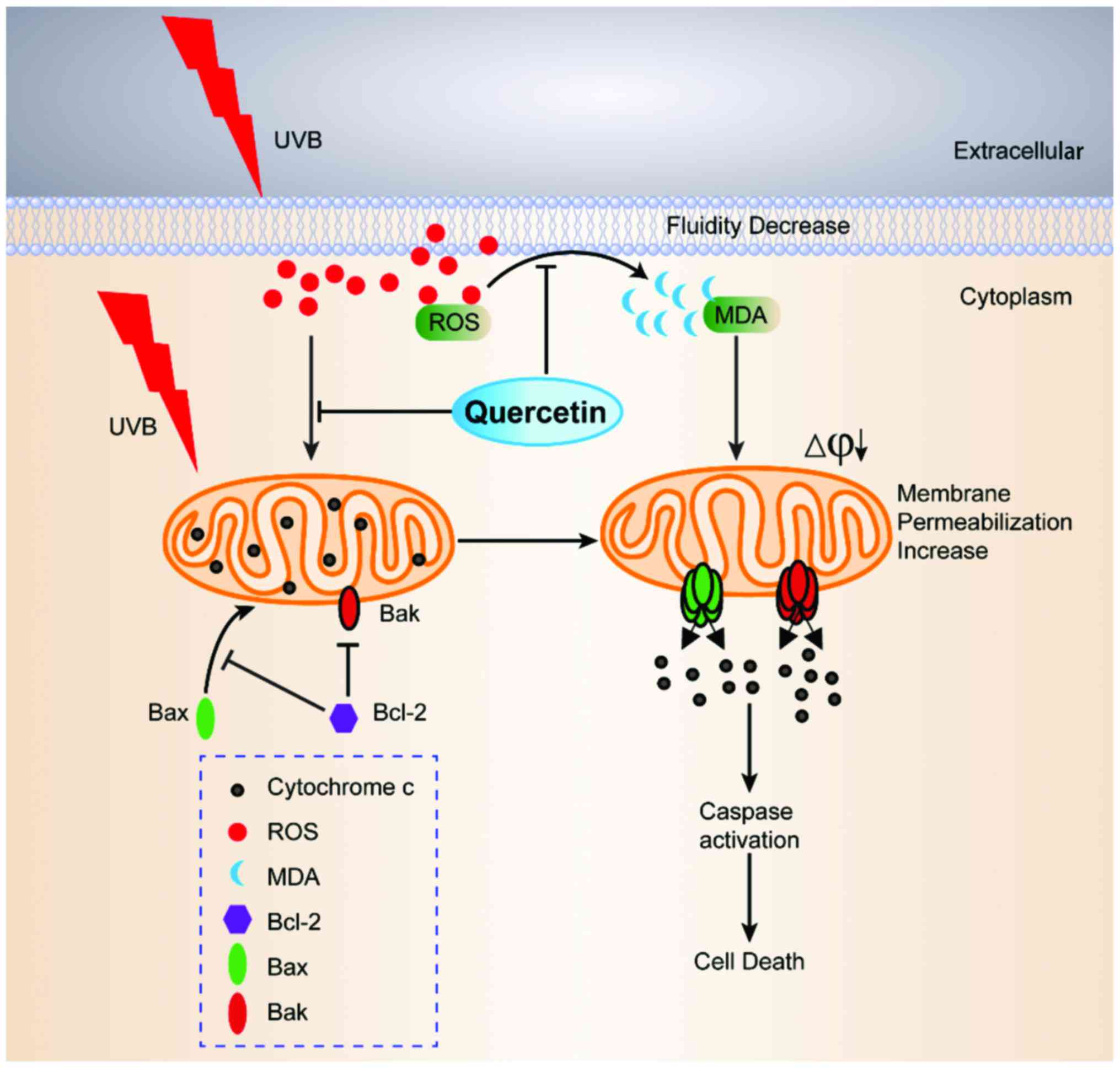
In conclusion, the present study demonstrated that
quercetin can effectively scavenge intracellular ROS, and
preincubation with quercetin blocked ROS attack on the cell
membrane and mitochondria after UVB exposure. The decrease in cell
membrane fluidity was inhibited by quercetin. The integrity of ΔΨm
was rescued and the mitochondrial morphology was protected.
Moreover, the release of cytochrome c was prevented and an
elevation in the Bcl-2/Bax ratio was observed. These data indicate
that quercetin prevented the cell membrane and mitochondria from
ROS-induced damage and blocked the mitochondrial-dependent
apoptotic pathway. The ROS clearing ability of quercetin provides
protection against UVB irradiation-induced apoptosis of HaCaT cells
(Fig. 8). In addition, ROS clearing
agent NAC effectively inhibited UVB-induced apoptosis (Fig. 3C) and further proved that the
protective effect of quercetin on UVB-induced cell death was
associated with ROS clearing ability. These findings suggest that
quercetin exerts beneficial effects against UVB irradiation-induced
skin photo-damage.
Acknowledgements
The present study was supported by a grant to XJ.S.
from the ‘Shenzhen Hong Kong Innovation Circle’ Research Program
(ZYB200907100147A) and the Novel Innovation Project of Graduate
School at Shenzhen, Tsinghua University (JZ2016000).
References
|
1
|
Paz ML, González Maglio DH, Weill FS,
Bustamante J and Leoni J: Mitochondrial dysfunction and cellular
stress progression after ultraviolet B irradiation in human
keratinocytes. Photodermatol Photoimmunol Photomed. 24:115–122.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Katiyar SK, Matsui MS, Elmets CA and
Mukhtar H: Polyphenolic antioxidant (−)-epigallocatechin-3-gallate
from green tea reduces UVB-induced inflammatory responses and
infiltration of leukocytes in human skin. Photochem Photobiol.
69:148–153. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hanson KM and Clegg RM: Observation and
quantification of ultraviolet-induced reactive oxygen species in ex
vivo human skin. Photochem Photobiol. 76:57–63. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Beak SM, Lee YS and Kim JA: NADPH oxidase
and cyclooxygenase mediate the ultraviolet B-induced generation of
reactive oxygen species and activation of nuclear factor-kappaB in
HaCaT human keratinocytes. Biochimie. 86:425–429. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sander CS, Chang H, Hamm F, Elsner P and
Thiele JJ: Role of oxidative stress and the antioxidant network in
cutaneous carcinogenesis. Int J Dermatol. 43:326–335. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bickers DR and Athar M: Oxidative stress
in the pathogenesis of skin disease. J Invest Dermatol.
126:2565–2575. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Black HS: Potential involvement of free
radical reactions in ultraviolet light-mediated cutaneous damage.
Photochem Photobiol. 46:213–221. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wlaschek M, Tantcheva-Poór I, Naderi L, Ma
W, Schneider LA, Razi-Wolf Z, Schüller J and Scharffetter-Kochanek
K: Solar UV irradiation and dermal photoaging. J Photochem
Photobiol B. 63:41–51. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Heck DE, Vetrano AM, Mariano TM and Laskin
JD: UVB light stimulates production of reactive oxygen species:
Unexpected role for catalase. J Biol Chem. 278:22432–22436. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Abdul-Muneer PM, Chandra N and Haorah J:
Interactions of oxidative stress and neurovascular inflammation in
the pathogenesis of traumatic brain injury. Mol Neurobiol.
51:966–979. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tavolari S, Munarini A, Storci G, Laufer
S, Chieco P and Guarnieri T: The decrease of cell membrane fluidity
by the non-steroidal anti-inflammatory drug Licofelone inhibits
epidermal growth factor receptor signalling and triggers apoptosis
in HCA-7 colon cancer cells. Cancer Lett. 321:187–194. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Orrenius S, Gogvadze V and Zhivotovsky B:
Mitochondrial oxidative stress: Implications for cell death. Annu
Rev Pharmacol Toxicol. 47:143–183. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Segre JA: Epidermal barrier formation and
recovery in skin disorders. J Clin Invest. 116:1150–1158. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Morrow DMP, Fitzsimmons PEE, Chopra M and
McGlynn H: Dietary supplementation with the anti-tumour promoter
quercetin: Its effects on matrix metalloproteinase gene regulation.
Mutat Res 480–481. 269–276. 2001. View Article : Google Scholar
|
|
15
|
Galati G, Moridani MY, Chan TS and O'Brien
PJ: Peroxidative metabolism of apigenin and naringenin versus
luteolin and quercetin: Glutathione oxidation and conjugation. Free
Radic Biol Med. 30:370–382. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pannala A Sekher, Chan TS, O'Brien PJ and
Rice-Evans CA: Flavonoid B-ring chemistry and antioxidant activity:
Fast reaction kinetics. Biochem Biophys Res Commun. 282:1161–1168.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sawa T, Nakao M, Akaike T, Ono K and Maeda
H: Alkylperoxyl radical-scavenging activity of various flavonoids
and other phenolic compounds: Implications for the
anti-tumor-promoter effect of vegetables. J Agric Food Chem.
47:397–402. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ioku K, Tsushida T, Takei Y, Nakatani N
and Terao J: Antioxidative activity of quercetin and quercetin
monoglucosides in solution and phospholipid bilayers. Biochim
Biophys Acta. 1234:99–104. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bonina F, Lanza M, Montenegro L, Puglisi
C, Tomaino A, Trombetta D, Castelli F and Saija A: Flavonoids as
potential protective agents against photo-oxidative skin damage.
Int J Pharm. 145:87–94. 1996. View Article : Google Scholar
|
|
20
|
Morel I, Lescoat G, Cogrel P, Sergent O,
Pasdeloup N, Brissot P, Cillard P and Cillard J: Antioxidant and
iron-chelating activities of the flavonoids catechin, quercetin and
diosmetin on iron-loaded rat hepatocyte cultures. Biochem
Pharmacol. 45:13–19. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Casagrande R, Georgetti SR, Verri WA Jr,
Dorta DJ, dos Santos AC and Fonseca MJV: Protective effect of
topical formulations containing quercetin against UVB-induced
oxidative stress in hairless mice. J Photochem Photobiol B.
84:21–27. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Casagrande R, Georgetti SR, Verri WA Jr,
Jabor JR, Santos AC and Fonseca MJV: Evaluation of functional
stability of quercetin as a raw material and in different topical
formulations by its antilipoperoxidative activity. AAPS
PharmSciTech. 7:E102006. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Goodwin JS and Kenworthy AK:
Photobleaching approaches to investigate diffusional mobility and
trafficking of Ras in living cells. Methods. 37:154–164. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pucadyil TJ and Chattopadhyay A: Effect of
cholesterol on lateral diffusion of fluorescent lipid probes in
native hippocampal membranes. Chem Phys Lipids. 143:11–21. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Axelrod D, Koppel DE, Schlessinger J,
Elson E and Webb WW: Mobility measurement by analysis of
fluorescence photobleaching recovery kinetics. Biophys J.
16:1055–1069. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Høyer-Hansen M, Bastholm L, Szyniarowski
P, Campanella M, Szabadkai G, Farkas T, Bianchi K, Fehrenbacher N,
Elling F, Rizzuto R, et al: Control of macroautophagy by calcium,
calmodulin-dependent kinase kinase-beta, and Bcl-2. Mol Cell.
25:193–205. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Rice-Evans CA, Miller NJ and Paganga G:
Structure-antioxidant activity relationships of flavonoids and
phenolic acids. Free Radic Biol Med. 20:933–956. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Skaper SD, Fabris M, Ferrari V, Carbonare
M Dalle and Leon A: Quercetin protects cutaneous tissue-associated
cell types including sensory neurons from oxidative stress induced
by glutathione depletion: Cooperative effects of ascorbic acid.
Free Radic Biol Med. 22:669–678. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Svobodova A, Walterova D and Vostalova J:
Ultraviolet light induced alteration to the skin. Biomed Pap Med
Fac Univ Palacky Olomouc Czech Repub. 150:25–38. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shindo Y and Hashimoto T: Ultraviolet
B-induced cell death in four cutaneous cell lines exhibiting
different enzymatic antioxidant defences: Involvement of apoptosis.
J Dermatol Sci. 17:140–150. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
LeBel CP, Ischiropoulos H and Bondy SC:
Evaluation of the probe 2′,7′-dichlorofluorescin as an indicator of
reactive oxygen species formation and oxidative stress. Chem Res
Toxicol. 5:227–231. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Millán-Plano S, García JJ,
Martínez-Ballarín E, Reiter RJ, Ortega-Gutiérrez S, Lázaro RM and
Escanero JF: Melatonin and pinoline prevent aluminium-induced lipid
peroxidation in rat synaptosomes. J Trace Elem Med Biol. 17:39–44.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Nakamura K, Nemani VM, Azarbal F,
Skibinski G, Levy JM, Egami K, Munishkina L, Zhang J, Gardner B,
Wakabayashi J, et al: Direct membrane association drives
mitochondrial fission by the Parkinson disease-associated protein
alpha-synuclein. J Biol Chem. 286:20710–20726. 2011. View Article : Google Scholar : PubMed/NCBI
|