Introduction
It is generally agreed that MDR is one of the most
important causes of failure in cancer chemotherapy. MDR is a
phenomenon in which cancer cells are resistant to the cytotoxic
effects of various structurally and mechanistically unrelated
chemotherapeutic agents. The result of MDR is often caused by
overexpression of cell membrane-bound ATP-binding cassette (ABC)
transporters (1–4). Forty-nine ABC transporters have been
identified in the human genome and are divided into seven
subfamilies (termed A-G) on the basis of sequence similarities
(5), among which, ABCB1
(P-glycoprotein, ABCB1/MDR1), ABCCs (multidrug
resistance-associated proteins), and ABCG2 (breast cancer
resistance protein/mitoxantrone resistance-associated
transporter/ABCP) play major roles in producing MDR in tumor cells.
Central to the mechanism of resistance to most chemotherapeutic
regimens is the overexpression of mdr1 gene, which encodes a
170-kDa transmembrane glycoprotein named as P-glycoprotein (P-gp).
Novel strategies targeting ABCB1, including the downregulation of
ABCB1 expression and/or function, may effectively circumvent drug
resistance and eliminate MDR cells to achieve better
chemotherapeutic effect (4). In
this regard, we have previously demonstrated that apatinib
(6,7), axitinib (8), crizotinib (9) and vandetanib (10) inhibit various ABC transporters and
reverse MDR in leukemia and solid tumors.
Pristimerin is a quininemethide triterpenoid
compound which has been found in various species belonging to
Celastraceae and Hippocrateaceae families and has
long been used as anti-inflammatory, antioxidant, antimalarial, and
insecticidal agents (11,12). It has been reported that
pristimerin, as a new proteasome inhibitor, has promising clinical
potential as both a therapeutic and chemopreventive agent for
cancer (13). Indeed, pristimerin
induces apoptotic cell death in certain human cancer cells,
including breast and lung cancer (14) and human acute myeloid leukemia
(15). Our previous data showed
that triterpenoid pristimerin induced HepG2 cells apoptosis through
ROS-mediated mitochondrial dysfunction (16) which revealed that pristimerin might
be a promising compound offering better anticancer treatment
options. In this study, we further investigated the effect of this
compound overcoming ABCB1-mediated chemotherapeutic drug resistance
in vitro and related molecular mechanisms.
Materials and methods
Chemicals and reagents
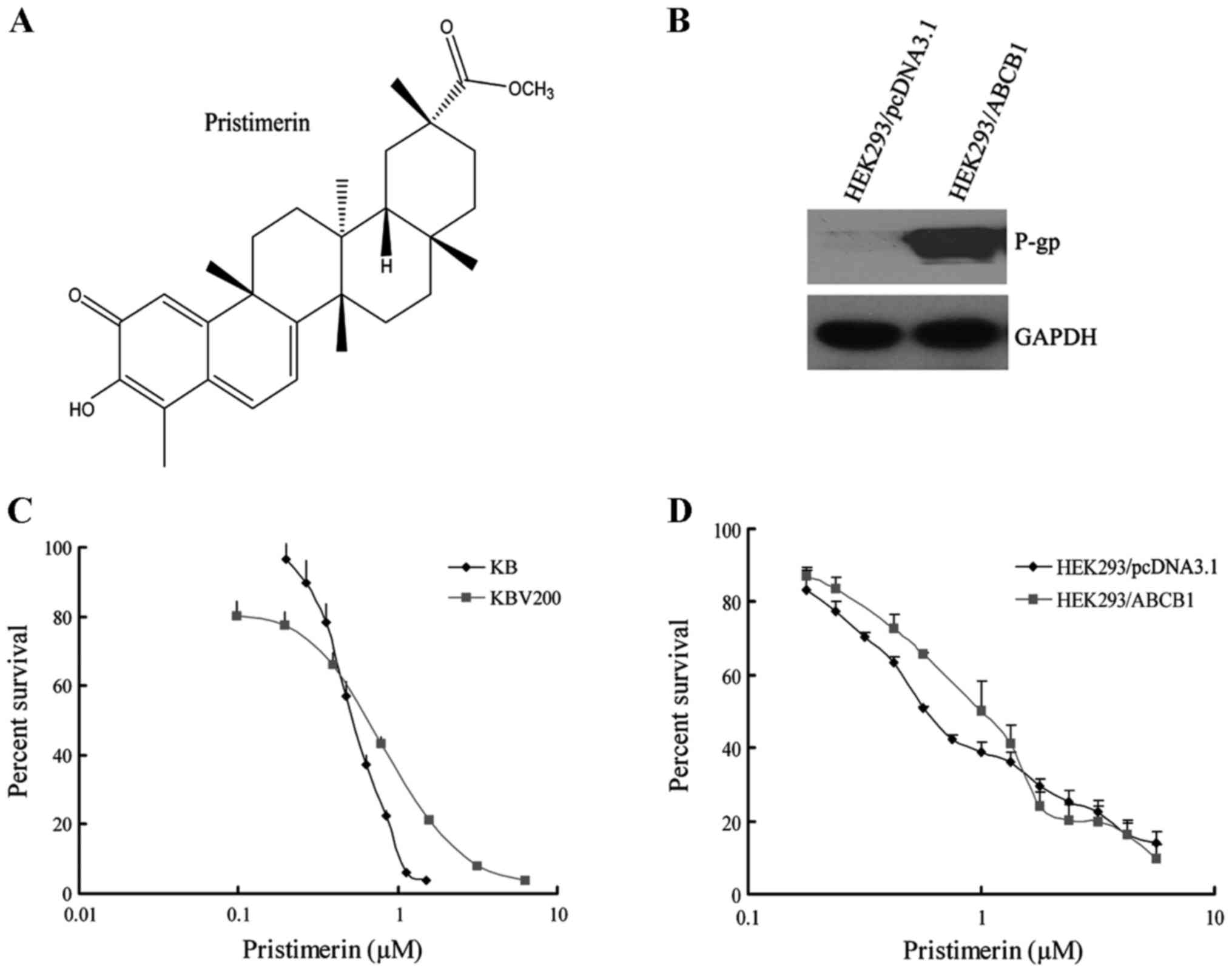
Pristimerin with a purity of >98% was purchased
from the PI & PI Technology Inc. (Guangzhou, China) and the
molecular structure is shown in Fig.
1A. Monoclonal antibodies against ABCB1 for western blotting
and immunofluorescence assay, and for flow cytometry were from
Santa Cruz Biotechnology, respectively. Antibodies against Bax,
Bcl-2, caspase-3 and PARP were obtained from Cell Signaling
Technology Inc. (Danvers, MA, USA). Antibodies against Akt, ERK1/2,
glyceraldehyde-3-phosphate dehydrogenase (GAPDH), anti-mouse and
anti-rabbit IgG-horseradish peroxidase were purchased from Kangchen
Biotechnology (Shanghai, China). DMEM and RPMI-1640 were products
of Gibco BRL. Platinum® SYBR® Green qPCR
SuperMix-UDG with ROX was obtained from Invitrogen Co. Protein
synthesis inhibitor cycloheximide and 3-(4,5)-dimethylthiazol-2-y1)-2,5-diphenyltetrazolium
bromide (MTT) were purchased from Sigma (St. Louis, MO, USA). Other
routine laboratory reagents of analytical or high-performance
liquid chromatography grade were obtained from Whiga Biotechnology
(Guangzhou, China).
Cell lines and culture
The following cell lines were cultured in Dulbecco's
modified Eagle's medium or RPMI-1640 medium supplemented with 10%
fetal bovine serum at 37°C in a humidified atmosphere of 5%
CO2. The human oral epidermoid carcinoma cell line KB
and its vincristine-selected, ABCB1-overexpressing derivative
KBv200 were gifts from Dr Xu-Yi Liu (Cancer Hospital of Beijing,
Beijing, China). The human primary embryonic kidney cell line
HEK293 and its stably pcDNA3.1- and ABCB1-transfected cell lines
HEK293/pcDNA3.1 and HEK293/ABCB1 (Fig.
1B) were obtained from Dr S.E. Bates (National Cancer
Institute, National Institutes of Health, Bethesda, MD, USA). All
of the transfected cells were cultured in medium with 2 mg/ml G418
(Geneticin) (17). All resistant
cells were authenticated through comparison of their fold
resistance with that of the parental, drug-sensitive cells and
examination of the expression levels of ABC transporters. All cells
were grown in drug-free culture medium for >2 weeks before
assays.
Cell viability assay
Cells harvested during logarithmic growth phase were
seeded in 96-well plates in a volume of 190 µl/well. After 24 h of
incubation, 10 µl of pristimerin full-range concentration was added
to the 96-well plates. After 68 h of treatment, 10 µl MTT (10 mg/ml
stock solution of saline) was added to each well for 4 h.
Subsequently, the supernatant was removed, and MTT crystals were
solubilized with 100 µl anhydrous DMSO each well. Thereafter, cell
viability was measured by model 550 microplate reader (Bio-Rad) at
540 nm with 655 nm as reference filter. The 50% inhibitory
concentration (IC50) was determined as the anticancer
drug concentration causing 50% reduction in cell viability and
calculated from the cytotoxicity curves (Bliss's software). Cell
percent survival rate was calculated using the following formula:
survival (%) = (mean experimental absorbance/mean control
absorbance) × 100%.
Assessment of apoptosis morphology by
Hoechst 33258 staining
After KB and KBv200 or HEK293/pcDNA3.1 and
HEK293/ABCB1 cells grown on coverslips were treated with 2.0 µmol/l
pristimerin for 48 h, both floating and trypsinized adherent cells
were collected, washed once with ice-cold phosphate-buffered saline
(PBS), fixed with 1 ml 4% paraformaldehyde for 20 min, and washed
once with ice-cold PBS. Then the cells were incubated in 1 ml PBS
containing 10 µmol/l Hoechst 33258 at 37°C for 30 min, washed
twice, and observed using fluorescence microscopy with standard
excitation filters (Leica Dmirb, Germany) in random microscopic
fields at ×400 magnification (18).
Annexin V/PI double-staining
assay
Annexin V and PI staining was performed using
ApopNexin™ FITC Apoptosis Detection kit. Cells (6×105)
were seeded in 25-cm2 flasks and allowed to attach for
24 h. After HEK293/pcDNA3.1 and HEK293/ABCB1 were treated with
0.5–2.0 µmol/l pristimerin for 48 h, both floating and attached
cells were collected, washed with ice-cold PBS twice and
resuspended in 200 µl 1X binding buffer containing Annexin V (1:50
according to the manufacturer's instructions) and 40 ng/sample of
PI for 15 min at 37°C in the dark. Then the number of viable,
apoptotic and necrotic cells were quantified by flow cytometer
(Becton-Dickinson, USA) and analysed by CellQuest software. Cells
were excited at 488 nm and the emissions of Annexin V at 525 nm and
PI were collected through 610 nm band pass filters, respectively.
At least 10,000 cells were analyzed for each sample. The percent
apoptosis (%) = (the number of apoptotic cells/the number of total
cells observed) × 100%.
Expression of ABCB1 analyzed by flow
cytometry
Expression of ABCB1 in the cell lines KB, KBv200,
HEK293/pcDNA3.1, and HEK293/ABCB1 were assessed through flow
cytometry. After KBv200 cells were treated with 2.0 µmol/l
pristimerin for indicated time, and HEK293/ABCB1 were treated with
0.5–4.0 µmol/l pristimerin for 48 h, single-cell suspensions were
prepared and washed three times with isotonic PBS (supplemented
with 0.5% bovine serum albumin). Then, 10 µl of
phycoerythrin-conjugated, mouse anti-human ABCB1 antibody was mixed
with 25 µl of cells (4×106 cells per ml). After
incubation for 45 min at 4°C in the dark, the cells were washed
twice with PBS (supplemented with 0.5% bovine serum albumin) and
were resuspended in 400 µl of PBS for flow cytometric analysis.
Isotype control samples were treated in an identical manner with
phycoerythrin-conjugated mouse IgG2a (19).
Immunofluorescence assay
After KBv200 or HEK293/ABCB1 cells grown on
coverslips were treated with 2.0 µmol/l pristimerin for 48 h, cells
were fixed with 4% polyoxymethylene for 20 min, and Triton X-100
was added for 10 min at room temperature. Then, the cells were
rinsed with PBS three times, and the non-specific binding sites
were blocked in PBS with 1% bovine serum albumin for 1 h. The cells
were incubated overnight at 4°C with ABCB1 antibody followed by
FITC-conjugated secondary antibody at room temperature for 1 h.
Subcellular distribution of P-gp protein was observed under
fluorescence microscopy with standard excitation filters (Leica
Dmirb) in random microscopic field at ×400 magnification (20).
RT-PCR and real-time RT-PCR
ABCB1 expression was assayed as described previously
(21). After pristimerin treatment
for indicated time, total cellular RNA was isolated with a TRIzol
reagent RNA extraction kit (Molecular Research Center, Cincinnati,
OH, USA), by following the manufacturer's instructions. The
first-strand cDNA was synthesized with oligo(dT) primers with
reverse transcriptase (Promega, Madison, WI, USA). The PCR primers
were 5′-cccatcattgcaatagcagg-3′ (forward) and
5′-gttcaaacttctgctcctga-3′ (reverse) for ABCB1 and
5′-ctttggtatcgtggaagga-3′ (forward) and 5′-caccctgttgctgtagcc-3′
(reverse) for GAPDH. With the use of a GeneAmp 9700 PCR system
(Applied Biosystems, Foster City, CA, USA), reactions were
performed at 94°C for 2 min for initial denaturation and then at
94°C for 30 sec, 58°C for 30 sec and 72°C for 1 min. After 32
cycles of amplification, additional extension was performed at 72°C
for 10 min. Products were resolved and examined through 1.5%
agarose gel electrophoresis. Expected PCR products were 157 bp for
ABCB1 and 475 bp for GAPDH.
Real-time RT-PCR was performed with a Bio-Rad CFX96
real-time system (Applied Biosystems). The geometric mean of GAPDH
levels was used as an internal control, to normalize the
variability in expression levels. The forward primer for GAPDH was
5′-gagtcaacggatttggtcgt-3′, and the reverse primer was
5′-gatctcgctcctggaagatg-3′. The forward primer for ABCB1 was
5′-gtggggcaagtcagttcatt-3′, and the reverse primer was
5′-tcttcacctccaggctcagt-3′. PCR was performed at 50°C for 2 min, at
95°C for 5 min, and then at 95°C for 15 sec and 60°C for 30 sec for
40 cycles. Relative quantification of ABCB1 was performed by using
the threshold cycle difference method (22). To ensure reproducibility of the
results, all genes were tested in triplicate in three independent
experiments (19).
Protein turnover assay
After KBv200 cells were treated with 2.0 µmol/l
pristimerin or DMSO for 12 h, the protein synthesis inhibitor
cycloheximide (CHX, 10 µg/ml) was added to the cells. After treated
for additional indicated time, the whole cells were harvested at
different time-points and whole cell lysates were analyzed by
western blotting (20).
Western blot analysis
Cells were harvested and washed twice with ice-cold
PBS, and the pellets were collected in 1X lysis buffer [50 mmol/l
Tris-HCl (pH 6.8), 10% glycerol, 2% SDS, 0.25‰ bromophenol blue,
and 0.1 mol/l DTT] was added for 100 µl/5×106 cells.
After heated at 95°C for 20 min, the lysates were centrifuged at
12,000 rpm for 10 min and the supernatant was collected. The
protein concentration was determined by nucleic acid-protein
analyzer (Beckman). Equal amount of lysate protein was separated on
8–12% SDS-PAGE and transferred onto polyvinylidene difluoride
membrane (Pall). The non-specific binding sites were blocked with
TBST buffer containing 5% non-fat dry milk for 2 h at room
temperature. The membranes were incubated overnight at 4°C with
specific primary antibodies, and the membranes were then washed
thrice with TBST buffer and incubated at room temperature for 1 h
with horseradish peroxidase-conjugated secondary antibody. After
three washes with TBST buffer, the immuno-blots were visualized by
the enhanced Phototope-Horseradish Peroxidase Detection kit
purchased from Cell Signaling Technology and exposed to Kodak
medical X-ray processor (Kodak, Rochester, NY, USA) (23).
Statistical analysis
For each protocol, three independent experiments
were performed. Results were expressed as the mean ± standard error
of the mean (SEM). Statistical calculations were performed by using
SPSS16.0 software. Differences in measured variables between
experimental and control groups were assessed by the Student's
t-test. P<0.05 was indicative of significant difference and
P<0.01 was indicative of very significant difference.
Results
Pristimerin shows equally potent
anticancer effect on parental and ABCB1-mediated MDR cell
lines
The cytotoxicity of pristimerin in different cell
lines was determined with the MTT assay. As shown in Fig. 1, the IC50 values were
0.54±0.01 and 0.52±0.01 µmol/l for KB and KBv200 cell lines,
respectively (P>0.05) (Fig. 1C).
Moreover, the IC50 values were 0.69±0.04 and 0.93±0.20
µmol/l for HEK293/pcDNA3.1 and HEK293/ABCB1 cells, respectively
(P>0.05) (Fig. 1D). The data
suggested that pristimerin exerted potent and similar cytotoxicity
to both parental and ABCB1-mediated MDR cell lines.
Pristimerin induces apoptosis in pairs
of parental and ABCB1-mediated MDR cell lines
To observe the morphological characteristics of
apoptosis, cells were stained with Hoechst 33258 after KB, KBv200,
HEK293/pcDNA3.1 and HEK293/ABCB1 cells were exposed to 2.0 µmol/l
pristimerin for 48 h respectively, and detected by fluorescence
microscopy. Control cells showed even distribution of the stain and
round homogeneous nuclei feature. Apoptotic cells displayed typical
changes including reduction of cellular volume, staining bright and
condensed or fragmented nucleus (Fig.
2A). For a further assessment of apoptosis induced by
pristimerin, we examined the exposure of phosphatidylserine on the
cell surface by using Annexin V and PI double staining. Flow
cytometric analysis revealed that the percentage of apoptotic cells
with Annexin V-positive but PI-negative cells increased gradually
with concentration in pristimerin treated cells (Fig. 2B). The early percent apoptosis was
1.30±1.20, 1.50±1.85, 20.20±2.45 and 39.17±3.15% in HEK293/pcDNA3.1
cells and 1.60±1.06, 4.57±1.11, 19.47±0.93 and 36.33±2.12% in
HEK293/ABCB1 cells, respectively (Fig.
2C). Intriguingly, we found that the early apoptosis rate was
approximately equal between HEK293/pcDNA3.1 and HEK293/ABCB1 cells
treated by the same concentration of pristimeirin (P>0.05).
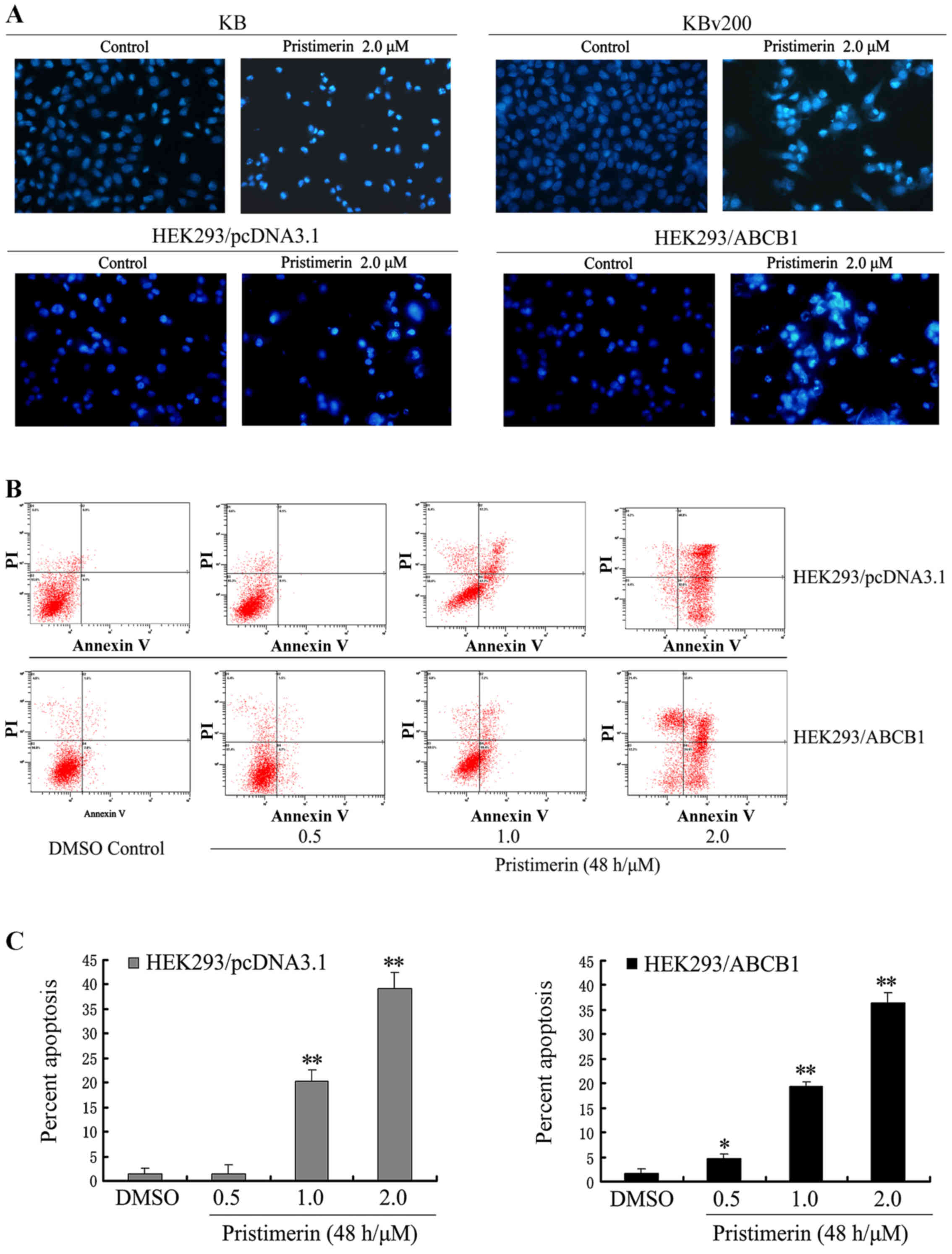 | Figure 2.(A) Cell apoptosis induced by
pristimerin was examined by Hoechst 33258 staining and observed
under fluorescence microscope at ×400 magnification. KB and KBv200
or HEK293/pcDNA3.1 and HEK293/ABCB1 cells grown on coverslips were
treated with 2.0 µmol/l pristimerin for 48 h. The apoptotic cells
detected by the fluorescence microscopy displayed condensed and
fragmented nuclei, shrinkage of cell volume, showed the
morphological changes of the above cells after exposure to
pristimerin. (B and C) Apoptosis analysis in HEK293/pcDNA3.1 and
HEK293/ABCB1 cells were assessed by Annexin V/PI double staining.
After cells were exposed to 0.5–2.0 µmol/l pristimerin for 48 h,
respectively, the attached and detached cells were collected.
Following staining with Annexin V and PI, cells were subjected to
flow cytometer analysis. (B) Bottom right quadrant, cells stained
mainly by Annexin V (early apoptotic cells); top right quadrant,
cells stained by both PI and Annexin V (late apoptotic/necrotic
secondary necrosis); top left quadrant, cells stained mainly by PI
viable cells; bottom left quadrant, cells negative for both Annexin
V and PI. (C) Percent apoptosis. Early apoptotic cell population
with Annexin V-positive but PI-negative cells increased gradually
from 1.30 to 39.17% and from 1.60 to 36.33% in HEK293/pcDNA3.1 and
HEK293/ABCB1 cells, respectively. Means ± SD of three assays.
*P<0.05; **P<0.01 versus control. |
Pristimerin inhibits cell
proliferation and induces cell apoptosis in pairs of MDR and
corresponding parental cell lines
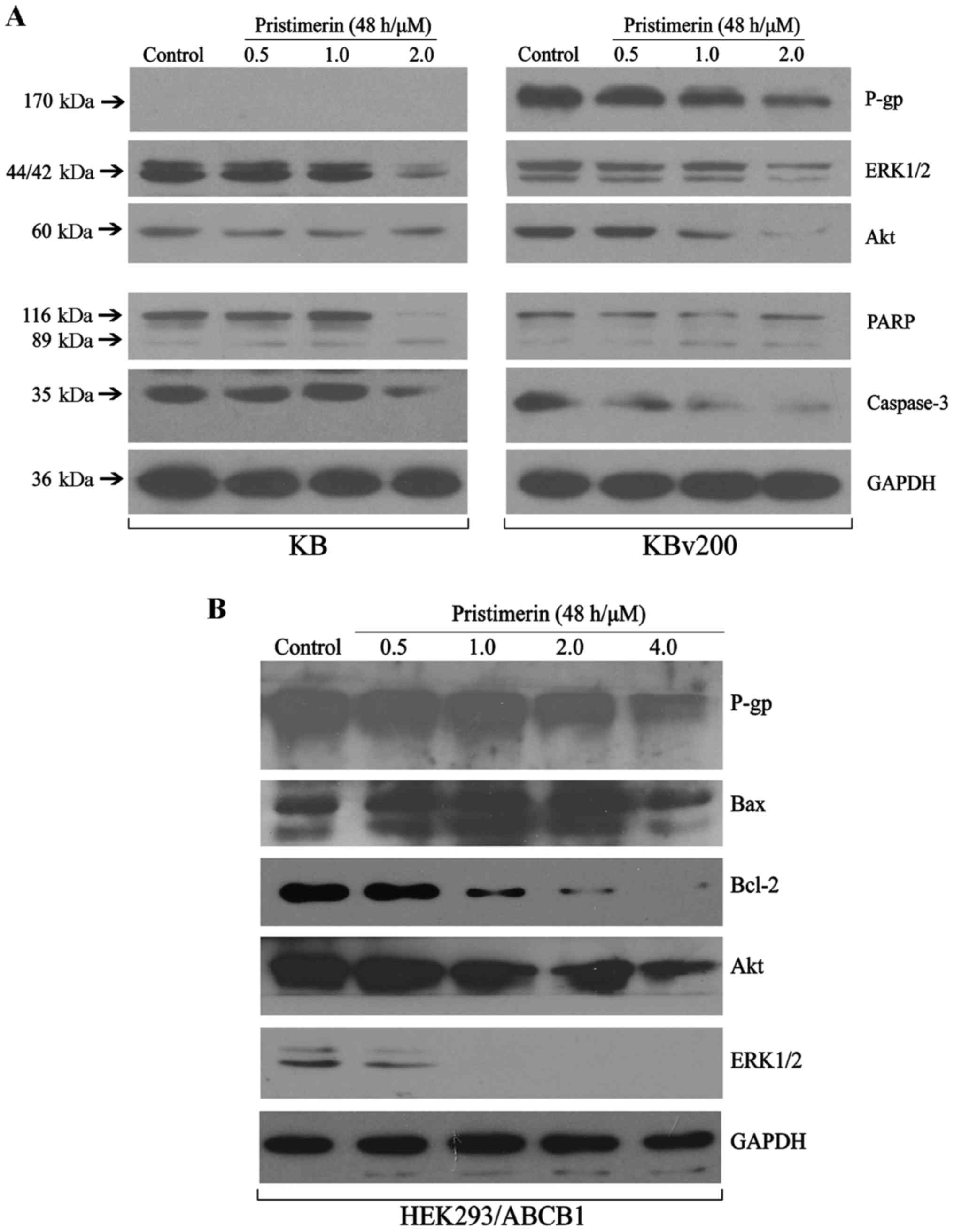
To further explore the mechanisms of pristimerin
inducing apoptosis of ABCB1-mediated MDR cell lines, western
blotting was done to detected the related protein expression. The
MAPK (mitogen-activated protein kinase) and PI3K
(phosphatidylinositol 3-kinase)/Akt signaling pathway are the two
important signalling pathway controlling cancer cell proliferation.
As shown in Fig. 3A, the expression
of total Akt and ERK1/2 proteins were inhibited in a dose-dependent
manner in KB and KBv200 cells. The results conformed that
pristimerin induced apoptosis similarly in both parental and its
corresponding ABCB1-mediated MDR cell lines. Western blotting also
revealed pristimerin induced growth inhibition and apoptosis in
stable transfected HEK293/ABCB1 cell lines, showing inhibition of
Akt and ERK1/2 proteins, and downregulation of Bcl-2/Bax in a
dose-dependent manner after pristimerin exposure in HEK293/ABCB1
cells (Fig. 3B), which suggested
that intrinsic pathways might take part in pristimerin-induced
apoptosis in ABCB1-overexpressing cell lines.
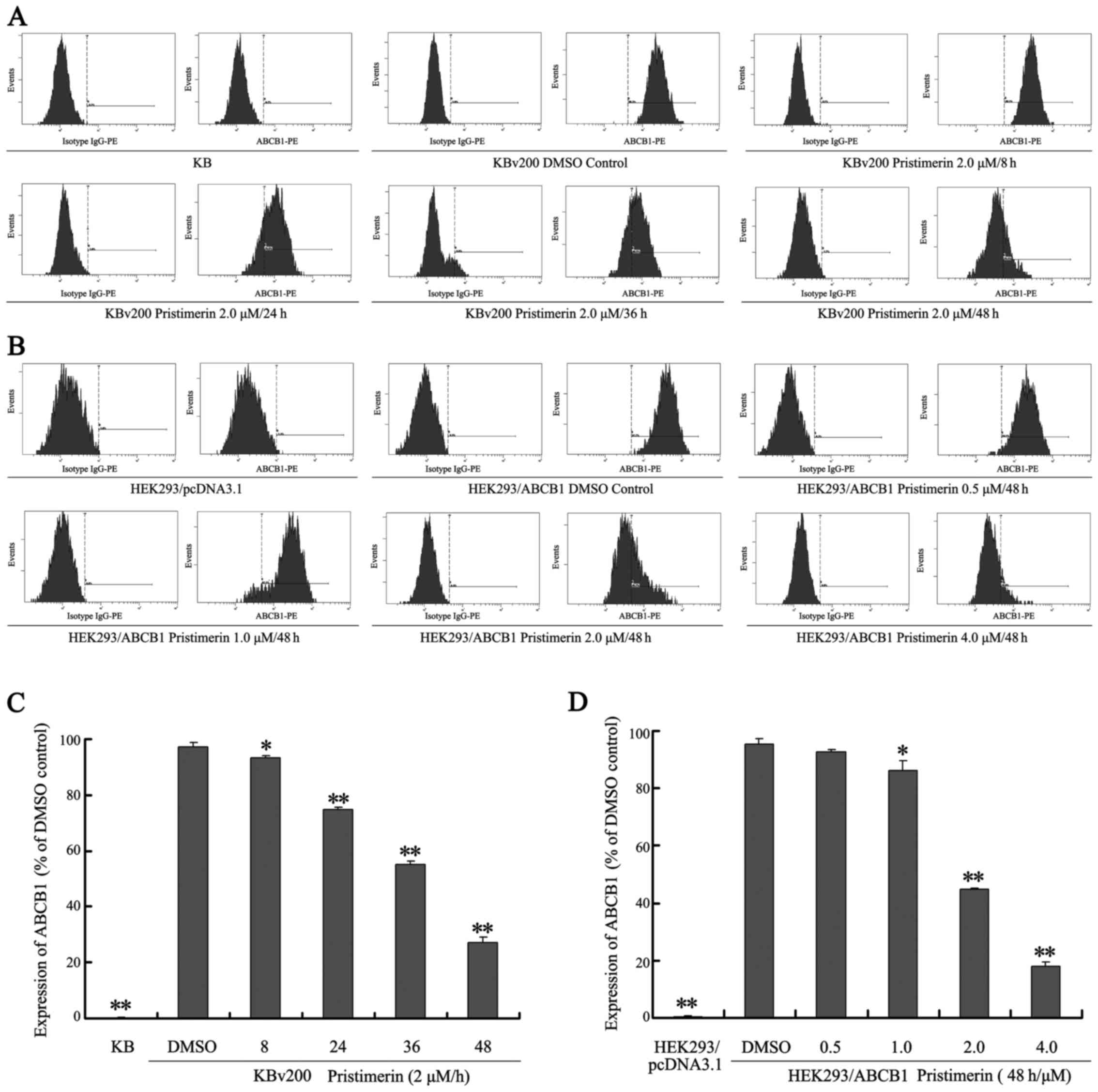
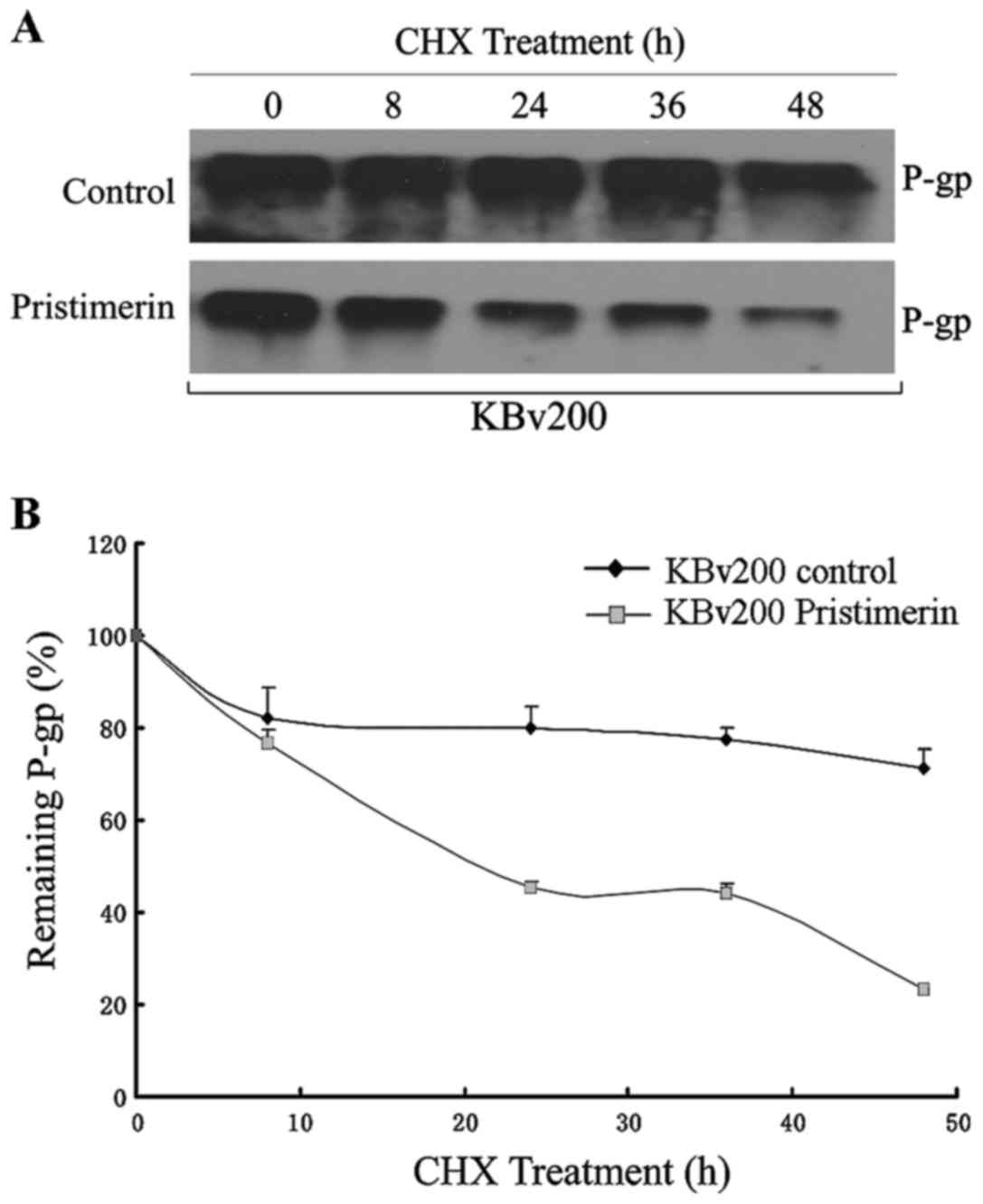
Downregulation of P-gp by pristimerin
is primarily dependent of P-gp protein stability
Our results show that pristimerin significantly
downregulated P-gp expression in a dose-dependent manner in both
drug-resistant KBv200 and stable transfected HEK293/ABCB1 cell
lines (Fig. 3A and B). Moreover,
after KBv200 cells were treated with 2.0 µmol/l pristimerin for the
indicated time, and HEK293/ABCB1 cells were treated with 0.5–4.0
µmol/l pristimerin for 48 h, pristimerin induced decrease of P-gp
membrane protein in a dose- and time-dependent manner detected by
flow cytometry (Fig. 4). The
expression of P-gp membrane protein was 0.17±0.15% for KB cells,
97.37±1.60, 93.23±1.00, 75.00±0.78, 55.17±1.27 and 27.17±1.96% for
KBv200 with 2.0 µmol/l prestimerin treatment for 0, 8, 24, 36 and
48 h, respectively (Fig. 4A and C).
The expression of P-gp membrane protein was 0.37±0.21% for
HEK293/pcDNA3.1 cells, 95.23±1.91, 92.53±1.14, 86.10±3.50,
44.70±0.56 and 17.97±1.65% for HEK293/ABCB1 cells with 0, 0.5, 1.0,
2.0 and 4.0 µmol/l prestimerin treatment for 48 h, respectively
(Fig. 4B and D).
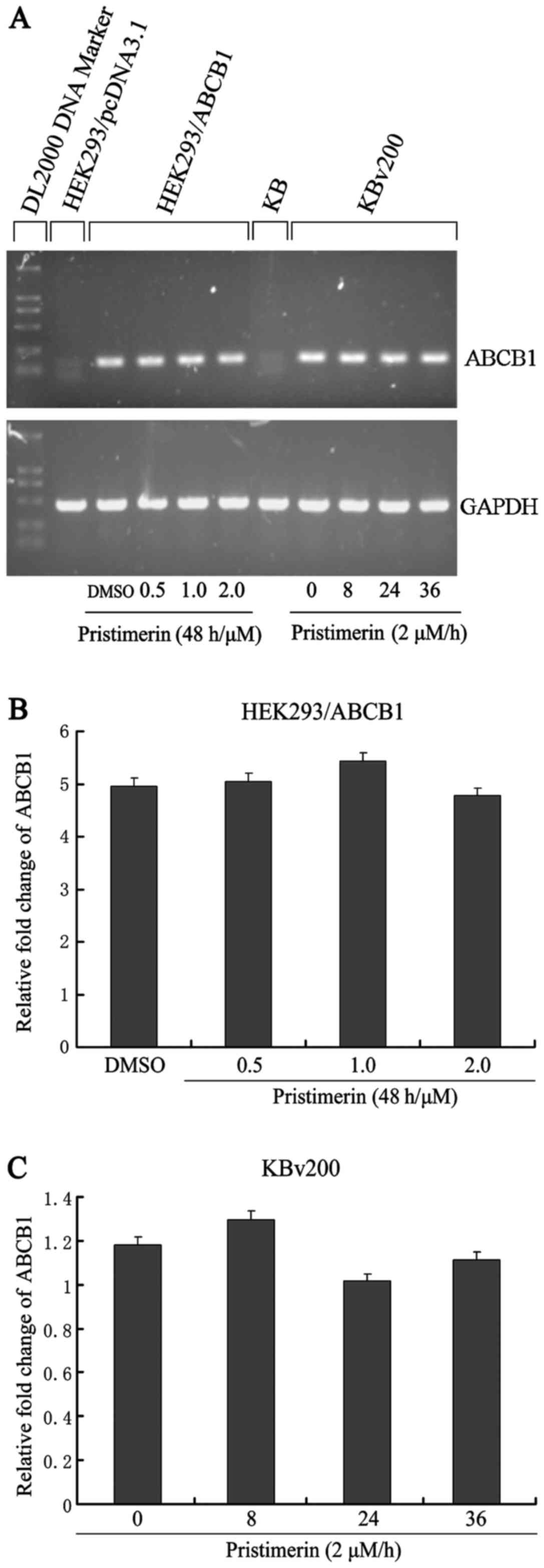
To examine whether P-gp was downregulated by
pristimerin at the mRNA level, we analyzed ABCB1 mRNA expression.
As shown in Fig. 5A, after KBv200
and HEK293/ABCB1 cells were exposed to pristimerin, no significant
change in ABCB1 mRNA expression was observed by semiquantitative
RT-PCR. This result comfirmed that only a slight change (P>0.05)
was detected by quantitative real-time RT-PCR analysis (Fig. 5B and C). These results indicated
that the decrease of P-gp protein caused by pristimerin was
independent of its mRNA level.
Like several other membrane proteins, the level of
P-gp seems to be regulated by protein stability (20). To investigate whether pristimerin
could downregulate P-gp through this mechanism, P-gp protein
stability was evaluated by using cycloheximide to stop protein
synthesis and measuring the amount of remaining P-gp protein at
various time-points after 2.0 µmol/l pristimerin treatment by
western blot analysis (Fig. 6A).
The quantitative analyses shown that a half-life of 744.63 h for
P-gp was significantly reduced to 22.09 h by pristimerin in KBv200
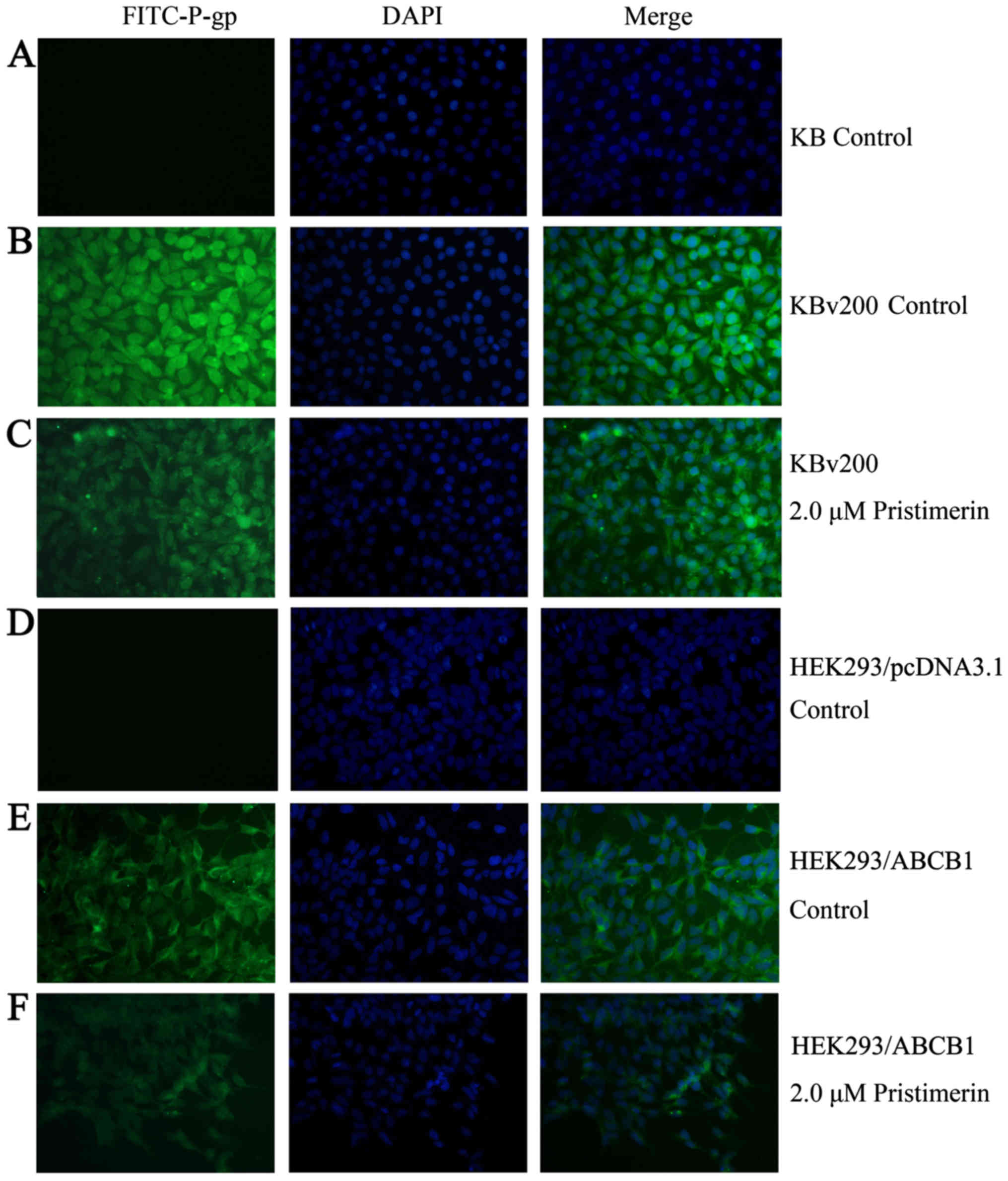
cells (Fig. 6B). Immunofluorescence
study with anti-P-gp antibody was also performed. As shown in
Fig. 7, the control cells had
strong immunofluorescence at the plasma membrane. After 2.0 µmol/l
pristimerin treatment for 48 h, the immunofluorescence at the
plasma membrane was attenuated and replaced with diffuse
cytoplasmic and nuclear punctate staining in drug-resistant KBv200
cells (Fig. 7B and C). Similar
findings were observed in stable transfected HEK293/ABCB1 cells
(Fig. 7E and F).
Discussion
Cancer has become an increasing public health
problem due to its high rates of morbidity and mortality.
Conventional cancer chemotherapy is limited by MDR caused by
overexpression of integral membrane transporters, such as P-gp,
which can efflux intracellular anticancer drugs thus decrease drug
accumulation. Developing new anticancer drugs which are efficient
to MDR cells is a feasible strategy to overcome MDR. Our previous
studies demonstrated that pristimerin induced HepG2 cell apoptosis
through ROS-mediated mitochondrial dysfunction (16). This study further showed that
pristimerin exhibited equally cytotoxicity and apoptosis in both
MDR and parental cell lines (Figs.
1–3). These data seem to
beautifully interpret the anti-MDR activities of pristimerin.
Pristimerin could not be the substrate of the drug transporter
P-gp. Therefore, there could be the same amount of pristimerin
inside the cell to induce apoptosis, which will be finally
translated into similar biological effects in MDR and corresponding
parental cells.
It is noteworthy that we observed that pristimerin
achieved effective downregulation of P-gp in a dose-dependent
manner (Fig. 3). Moreover,
pristimerin induced decrease of P-gp membrane protein in
drug-resistant KBv200 cells and stable transfected HEK293/ABCB1
cells detected by flow cytometry (Fig.
4). Further investigation of the signal molecules revealed that
components of the proliferation- and survival-associated MAPK and
PI3K/Akt pathways were markedly inhibited by pristimerin (Fig. 3). We observed that P-gp
downregulation contributed to equal cytotoxicity and apoptosis
induced by pristimerin in KB and KBv200 cells (Fig. 3A). Therefore, it is plausible that
P-gp interferes with DNA damage signaling to apoptotic machinery or
with signal transductions within apoptotic cascades, which is
further supported by the findings of the apoptosis inhibition of
P-gp (24–26).
How is the expression of P-gp downregulated by
pristimerin? Two possible mechanisms may be involved. First,
pristimerin downregulated P-gp protein at the mRNA level; and
second, ubiquitin-proteasome-, lysosome-, and calpain-mediated
protein degradation systems regulated the degradation of P-gp by
pristimerin. Interestingly, our data revealed that pristimerin
decreased P-gp protein expression independent of its mRNA level
(Fig. 5). However, protein turnover
assay revealed that the half-life of P-gp protein was significantly
reduced by pristimerin (Fig. 6),
suggesting that downregulation of P-gp by pristimerin was primarily
dependent of P-gp protein stability. We will further explore which
protein degradation systems are involved in the degradation of P-gp
protein caused by pristimerin.
In conclusion, we presented cell biological evidence
that pristimerin inhibited cell proliferation and induced cell
apoptosis in ABCB1-overexpressing cancer cells and was a potent
cytotoxic agent to ABCB1-mediated MDR cells. Importantly, we
demonstrated for the first time that pristimerin treatment led to
downregulation of P-gp, and intracellular redistribution, which may
represent a novel approach for the targeted therapy of
ABCB1-overexpressing cancers.
Acknowledgements
This study was supported by the National Science
Foundation for Young Scientists of Shanxi Province (2014021037-6),
the Research Foundation of Collaborative Innovation Center for
Cancer and the Priming Scientific Research Foundation for Ph.D. in
Shanxi Datong University (no. 2010-B-11).
Glossary
Abbreviations
Abbreviations:
|
ABCB1
|
ATP-binding cassette subfamily B
member 1
|
|
P-gp
|
P-glycoprotein
|
|
MTT
|
3-(4,5)-dimethylthiazol-2-y1)-2,5-diphenyltetrazolium
bromide
|
|
MAPK
|
mitogen-activated protein kinase
|
|
PI3K
|
phosphatidylinositol 3-kinase
|
|
ERK
|
extracellular signal-regulated
kinase
|
|
GAPDH
|
glyceraldehyde-3-phosphate
dehydrogenase
|
|
v/v
|
volume per volume
|
|
RT-PCR
|
reverse transcription-polymerase chain
reaction
|
|
PBS
|
phosphate-buffered saline
|
|
TBST
|
Tris-buffered saline/Tween-20
|
References
|
1
|
Pérez-Tomás R: Multidrug resistance:
Retrospect and prospects in anti-cancer drug treatment. Curr Med
Chem. 13:1859–1876. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Eytan GD: Mechanism of multidrug
resistance in relation to passive membrane permeation. Biomed
Pharmacother. 59:90–97. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Johnson WW: P-glycoprotein-mediated efflux
as a major factor in the variance of absorption and distribution of
drugs: Modulation of chemotherapy resistance. Methods Find Exp Clin
Pharmacol. 24:501–514. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fojo T and Bates S: Strategies for
reversing drug resistance. Oncogene. 22:7512–7523. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dean M, Rzhetsky A and Allikmets R: The
human ATP-binding cassette (ABC) transporter superfamily. Genome
Res. 11:1156–1166. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mi YJ, Liang YJ, Huang HB, Zhao HY, Wu CP,
Wang F, Tao LY, Zhang CZ, Dai CL, Tiwari AK, et al: Apatinib
(YN968D1) reverses multidrug resistance by inhibiting the efflux
function of multiple ATP-binding cassette transporters. Cancer Res.
70:7981–7991. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tong XZ, Wang F, Liang S, Zhang X, He JH,
Chen XG, Liang YJ, Mi YJ, To KK and Fu LW: Apatinib (YN968D1)
enhances the efficacy of conventional chemotherapeutical drugs in
side population cells and ABCB1-overexpressing leukemia cells.
Biochem Pharmacol. 83:586–597. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang F, Mi YJ, Chen XG, Wu XP, Liu Z, Chen
SP, Liang YJ, Cheng C, To KK and Fu LW: Axitinib targeted cancer
stemlike cells to enhance efficacy of chemotherapeutic drugs via
inhibiting the drug transport function of ABCG2. Mol Med.
18:887–898. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhou WJ, Zhang X, Cheng C, Wang F, Wang
XK, Liang YJ, To KK, Zhou W, Huang HB and Fu LW: Crizotinib
(PF-02341066) reverses multidrug resistance in cancer cells by
inhibiting the function of P-glycoprotein. Br J Pharmacol.
166:1669–1683. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zheng LS, Wang F, Li YH, Zhang X, Chen LM,
Liang YJ, Dai CL, Yan YY, Tao LY, Mi YJ, et al: Vandetanib
(Zactima, ZD6474) antagonizes ABCC1- and ABCG2-mediated multidrug
resistance by inhibition of their transport function. PLoS One.
4:e51722009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gao JM, Wu WJ, Zhang JW and Konishi Y: The
dihydro-beta-agarofuran sesquiterpenoids. Nat Prod Rep.
24:1153–1189. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Brinker AM, Ma J, Lipsky PE and Raskin I:
Medicinal chemistry and pharmacology of genus Tripterygium
(Celastraceae). Phytochemistry. 68:732–766. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Salminen A, Lehtonen M, Suuronen T,
Kaarniranta K and Huuskonen J: Terpenoids: Natural inhibitors of
NF-kappaB signaling with anti-inflammatory and anticancer
potential. Cell Mol Life Sci. 65:2979–2999. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wu CC, Chan ML, Chen WY, Tsai CY, Chang FR
and Wu YC: Pristimerin induces caspase-dependent apoptosis in
MDA-MB-231 cells via direct effects on mitochondria. Mol Cancer
Ther. 4:1277–1285. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nagase M, Oto J, Sugiyama S, Yube K,
Takaishi Y and Sakato N: Apoptosis induction in HL-60 cells and
inhibition of topoisomerase II by triterpene celastrol. Biosci
Biotechnol Biochem. 67:1883–1887. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Guo Y, Zhang W, Yan YY, Ma CG, Wang X,
Wang C and Zhao JL: Triterpenoid pristimerin induced HepG2 cells
apoptosis through ROS-mediated mitochondrial dysfunction. J BUON.
18:477–485. 2013.PubMed/NCBI
|
|
17
|
Robey RW, Honjo Y, Morisaki K, Nadjem TA,
Runge S, Risbood M, Poruchynsky MS and Bates SE: Mutations at
amino-acid 482 in the ABCG2 gene affect substrate and antagonist
specificity. Br J Cancer. 89:1971–1978. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wu QL, Wu XP, Liang YJ, Chen LM, Ding Y
and Fu LW: P-glycoprotein is not involved in pathway of
anti-Fas/Fas-induced apoptosis in KBv200 cells. World J
Gastroenterol. 11:3544–3548. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhao XQ, Xie JD, Chen XG, Sim HM, Zhang X,
Liang YJ, Singh S, Talele TT, Sun Y, Ambudkar SV, et al: Neratinib
reverses ATP-binding cassette B1-mediated chemotherapeutic drug
resistance in vitro, in vivo, and ex vivo. Mol Pharmacol. 82:47–58.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yan YY, Zheng LS, Zhang X, Chen LK, Singh
S, Wang F, Zhang JY, Liang YJ, Dai CL, Gu LQ, et al: Blockade of
Her2/neu binding to Hsp90 by emodin azide methyl anthraquinone
derivative induces proteasomal degradation of Her2/neu. Mol Pharm.
8:1687–1697. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dai CL, Tiwari AK, Wu CP, Su XD, Wang SR,
Liu DG, Ashby CR Jr, Huang Y, Robey RW, Liang YJ, et al: Lapatinib
(Tykerb, GW572016) reverses multidrug resistance in cancer cells by
inhibiting the activity of ATP-binding cassette subfamily B member
1 and G member 2. Cancer Res. 68:7905–7914. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yan Y, Su X, Liang Y, Zhang J, Shi C, Lu
Y, Gu L and Fu L: Emodin azide methyl anthraquinone derivative
triggers mitochondrial-dependent cell apoptosis involving in
caspase-8-mediated Bid cleavage. Mol Cancer Ther. 7:1688–1697.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Johnstone RW, Cretney E and Smyth MJ:
P-glycoprotein protects leukemia cells against caspase-dependent,
but not caspase-independent, cell death. Blood. 93:1075–1085.
1999.PubMed/NCBI
|
|
25
|
Johnstone RW, Ruefli AA, Tainton KM and
Smyth MJ: A role for P-glycoprotein in regulating cell death. Leuk
Lymphoma. 38:1–11. 2000.PubMed/NCBI
|
|
26
|
Ruefli AA, Smyth MJ and Johnstone RW: HMBA
induces activation of a caspase-independent cell death pathway to
overcome P-glycoprotein-mediated multidrug resistance. Blood.
95:2378–2385. 2000.PubMed/NCBI
|