Introduction
Laryngeal cancer consists mostly of squamous cell
carcinomas (also known as the larynx or laryngeal carcinoma),
indicating their origin from human skin of larynx (1,2).
According to GLOBOCAN 2012, there were an estimated 156,877 new
cases and 83,376 deaths in the world, and the adjusted incidence
and mortality rates were 2.1/100,000 and 1.1/100,000, respectively
(3). Cancer could develop in any
part of the larynx according to a previous study. However, the cure
rate presently is influenced by the tumor location (4). The origin of laryngeal cancer is a
specialised area, which needs the coordinated expertise of ear,
nose and throat surgeons and oncologists (5,6). The
laryngeal cancer symptoms rely on the tumor size and location.
Specific treatment has a close relationship with the type,
location, as well as the stage of laryngeal cancer (7,8).
Treatment includes radiotherapy, surgery and chemotherapy, alone or
in combination. However, a patient affected severely may need a
laryngectomy, the total or the partial remove of the vocal cords.
In 2013, data indicated that laryngeal cancer has led to a large
number of deaths since 1990 (9).
Furthermore, the survival rate of patients with laryngeal cancer
has decreased compared with the survival rate of patients with all
other types of cancers as a whole (4). Hence, finding effective and safe
therapeutic strategy is urgenyly needed to improve cancer
treatments. It has been indicated that the treatment of a variety
of drugs targeting different signaling pathways can provide
effective strategy for various cancer cell mutations, and postpone
cancer adaptation procedure subsequently.
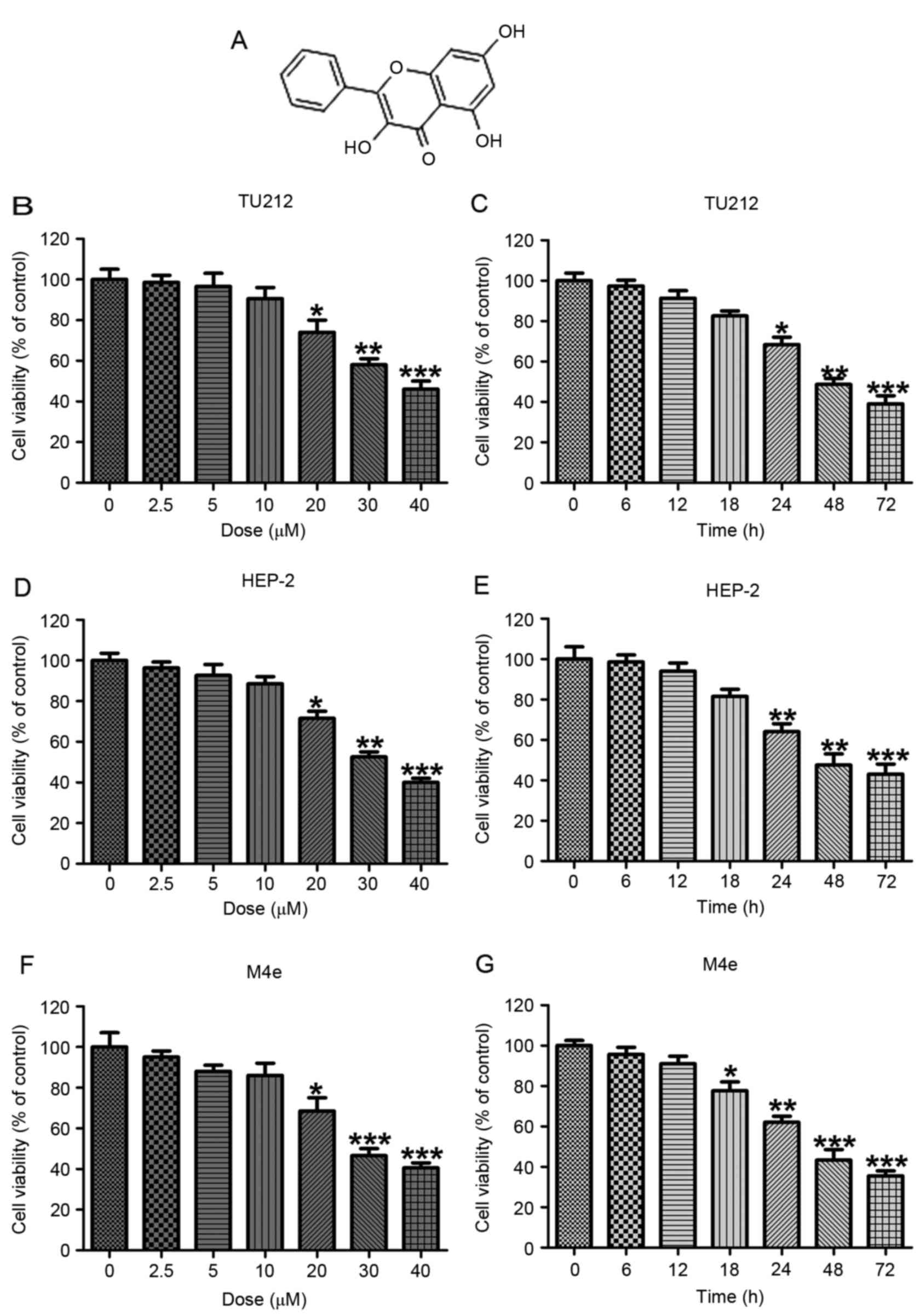
Galangin (Fig. 1A)
is a member of the flavonol class of flavonoids, which occurs at
high concentrations in the rhizome of Alpinia officinarum
Hance, as well as in propolis, applied in China for centuries
as a spice and a traditional Chinese medicine for various diseases
(10,11). As a potential scavenger of free
radicals, including singlet oxygen and superoxide anion, galangin
has various bioactivities and influences many cellular processes
(12). In addition to its
anti-mutagenic, anti-oxidant, as well as anti-inflammatory
functions, galangin has been reported to possess antitumor role in
a number of in vitro and in vivo systems, including
melanoma, hepatoma and leukaemia (13,14).
Furthermore, a previous study suggested that galangin possesses
therapeutic potential as an antitumor agent for liver cancer
(15). Galangin induced apoptosis
by suppressing tumor cell migration, promoting caspase-3 and
increasing ROS production (16).
However, the effect of galangin on human laryngeal cancer
progression has not been investigated yet. In the present study, we
investigated the anticancer effects of galangin on two types of
human laryngeal cancer cells.
Apoptosis is a distinct genetic and biochemical
pathway of cell death necessary for cell growth, development and
maintenance of homeostasis in organisms. Caspase-9 was activated
with the elevated level of cleaved caspase-9, which in turn cleaved
caspase-3 and ultimately induced apoptosis (17,18).
Autophagy is a cellular process of catabolic degradation in which
damaged, dysfunctional, or superfluous organelles and proteins are
sequestered, engulfed, and recycled to maintain cellular
metabolism, viability and homeostasis (19,20).
The mTOR kinase, which is activated by signaling pathway
originating from growth factors, plays a critical role in
regulating autophagy progression (21). There are diverse signaling pathways
implicated in the regulation of mTOR signaling, including positive
regulation of mTOR (PI3K/Akt and p38 MAPK signaling) suppressing
autophagy (22,23). The PI3K/Akt signaling pathway is
implicated in cell migration and invasion, which induces the
expression of NF-κB transcription factor, resulting in cancer cell
proliferation (24).
This study is the first time that galangin was used
to treat human laryngeal cancer TU212 and HEP-2 cells, to prove if
galangin had a potential effect on inhibiting laryngeal cancer
progression in the future. In the present study, TU212 and HEP-2
cells, exposed to galangin, suppressed cancer cell proliferation,
migration and invasion. Caspase-3, PI3K/AKT and p38 signaling
pathways were investigated. These results indicated that galangin
performed its anticancer effect on human laryngeal cancer possibly
via inactivating PI3K/AKT- and p38-signaling pathways.
Materials and methods
Cells and treatment
Human laryngeal cancer cell lines, TU212 and M4e,
human normal larynx epithelial HBE cells, and human normal liver
HHL-5 cells, were purchased from the American Type Culture
Collection (ATCC; Manassas, VA USA). The mouse normal larynx
epithelial RTE cells, and human laryngeal cancer HEP-2 cells were
purchased from the Nanjing KeyGen Biotech, Co., Ltd. (Nanjing,
China). TU212, HEP-2 and RTE cells were routinely cultured in
RPMI-1640 medium (Gibco, Carlsbad, CA, USA), containing 10% fetal
bovine serum (FBS; Gibco), 1% penicillin/streptomycin. The cell
lines M4e, HBE and HHL-5 were cultured in Dulbeccos modified Eagles
medium (DMEM; Gibco) supplemented with 10% FBS, 100 U/ml penicillin
and 100 µg/ml streptomycin. All cells were kept in a humidified
atmosphere with 5% CO2 and 95% humidity at 37°C in an
incubator. Galangin (>98% purity), purchased from the Hangzhou
DayangChem, Co., Ltd., (Hangzhou, China) was used for the treatment
of human laryngeal cancer dissolved in dimethyl sulfoxide (DMSO)
and then stored at −20°C for experimental treatment use. The final
DMSO concentration in cells is <0.1% (v/v) in each
treatment.
Cell viability analysis
In order to calculate the growth inhibitory role of
galangin in different cell lines, ~1×103 cells/well were
seeded in plates (Corning Inc., Corning, NY, USA) with the
respectively complete growth media. The following day, the cells
were treated with different concentrations of galangin for
different time as shown in the figures and incubated at 37°C. Then,
cell viability was calculated by
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide
(MTT) analysis at 570 nm.
Wound healing analysis
TU212 and HEP-2 cells were seeded in 60-mM dishes
and incubated until confluence. After a 3-h cell pre-treatment with
50 µM mytomicin C, wounds were created by scratching cell sheets
with a sterile 200-µl pipette tip. The culture medium was replaced
with fresh medium containing either DMSO or galangin. The images of
a specific position on the scratched areas were taken by an
inverted microscope (Leica Microsystems, Wetzlar, Germany) using a
×10 objective every 24 h. The wound widths were assessed and the
relative wound widths were evaluated.
Cancer cell migration and
invasion
TU212 and HEP-2 cells were seeded into the upper
chamber of a Transwell insert pre-coated with 5 µg/ml fibronectin
for migration or a BD™ Matrigel invasion chamber for invasion.
Medium with 10% serum was put in the lower chamber as a
chemo-attractant, and cells were then incubated for 4 h of
migration invasion. Non-migratory and non-invasive cancer cells
were removed from the upper chamber using a cotton bud. The cells
on the lower insert surface were stained with Diff-Quick. Cells
were finally calculated as the number of cells observed in five
different microscope fields of three independent inserts.
Apoptosis analysis
Apoptosis assay of tissue samples was determined by
TUNEL used an In Situ Cell Death Detection kit, fluorescein (Roche
Applied Science, Indianapolis, IN, USA) following the manufacturers
protocol. Tumor tissue sections were counterstained with
hematoxylin. Then, the number of TUNEL-positive cells was evaluated
under a microscope. The number of apoptosis cells were counted by
the ratio of apoptotic cells to the total cells. The experiment was
conducted three times independently for each cell line.
TU212 and HEP-2 cell apoptosis was
determined by flow cytometric (FCM) analysis
The tumor cells were harvested, and then washed
three times with chilled phosphate-buffered saline (PBS), stained
with Annexin V-FITC and propidium iodide (PI) diluted in the
binding buffer, and tested by FACSCalibur FCM (BD Biosciences, San
Jose, CA, USA) for 15-min incubation at the room temperature in the
dark. Fluorescence was then detected at an excitation wavelength of
480 nm through 530 nm FL-1 and 585 nm FL-2 filters. The apoptotic
cells were then quantified.
Western blot analysis
For western blot analysis, the cancer cells of TU212
and HEP-2 were washed with chilled PBS and lysed on ice in modified
RIPA buffer, containing 50 mM Tris-HCl pH 7.4, 1% NP-40, 0.25%
Na-deoxycholate, 150 mM NaCl, 1 mM Na3VO4,
and 1 mM NaF, with protease inhibitors (100 µM phenylmethylsulfonyl
fluoride, 10 µg/ml leupeptin, 10 µg/ml pepstatin and 2 mM EDTA).
The lysates were centrifuged at 12,000 × g for 20 min at 4°C and
the supernatant fractions were collected. The proteins were
separated by SDS-PAGE electrophoresis and transferred to
Immobilon-P membranes. The specific proteins were detected using an
enhanced chemiluminescence (ECL) western blotting kit according to
the manufacturers instructions. The primary antibodies including
PI3K, Bcl-2, Bax, caspase-9, caspase-3, PARP, LC3, Beclin 1, Ras,
Raf, p38, AKT, p-AKT, NF-κB, p-NF-κB and GAPDH. GAPDH was used as
the loading control. The gray value of the western blotting band
was anlyzed by ImageJ software (Version 1.4.2b, Mac OS X; National
Institutes of Health, Bethesda, MD, USA USA), representing the
specific protein expression levels.
Nude mouse xenograft tumor assay
The 6-week-old SPF male BALB/c nude mice, weighed
20–25 g, and were obtained from Vital River Laboratory Animals Co.,
Ltd. (Beijing, China). Before the experiments, all mice were
required to adapt to the environment for a week. They were housed
in a specific pathogen-free, temperature and humidity-controlled
environment (25±2°C, 50±2% humidity) with a standard 12-h
light/12-h dark cycle with food and water in their cages. All
procedures were in accordance with the Regulations of Experimental
Animal Administration issued by the Ministry of Science and
Technology of China. The mice were randomly divided into 4 groups:
the control (Con) group, the galangin group (10 mg/kg), the
galangin group (20 mg/kg) and the galangin group (30 mg/kg).
Subsequently, 150 µl of the TU212 cell suspension (containing
2×107 cells) was injected subcutaneously into the right
axilla area of the mice. Ten days before the cancer cell
inoculation, the treatment groups were gavaged with galangin every
day and observed for the growth of tumors at 42 days. The control
mice were only injected with the equal volume of TU212 cells. The
growth of tumors were determined after the mice were sacrificed,
and stored for the following experiments.
Histopathological examination of
tissues
Histopathological evaluation was performed on mice
that were collected. Samples were fixed with 10% buffered formalin,
imbedded in paraffin and sliced. Samples were subjected to
immunohistochemical staining (Ki-67) according to CST Technology
Co. Introduction and performed by Shanghai Zhenda Biotechnology,
Co., Ltd. (Shanghai, China).
Immunofluorescence assay
TU212 and HEP-2 cells were cultured on sterilized
glass coverslips overnight and treated with galangin for 24 h.
After being fixed with 4% paraformaldehyde solution and blocked
with 4% BSA in PBS, cells on coverslips were incubated with TSC1,
LC3II and Bax primary antibody and anti-rabbit secondary antibody
conjugated with Alex Flour 555. Images were captured with a
fluorescence microscope (Olympus, Tokyo, Japan).
Statistical analysis
Data were expressed as mean ± SEM from three or more
experiments. Treated cells and the corresponding controls were
compared using GraphPad Prism (version 6.0; GraphPad Software,
Inc., La Jolla, CA, USA) by a one-way ANOVA with Dunns least
significant difference tests. Differences between groups were
considered significant at P<0.05.
Results
Galangin inhibits human laryngeal
carcinoma cell viability without toxicity on normal cells
The human laryngeal carcinoma cell viability treated
by galangin was calculated. As shown in Fig. 1B, TU212 cells were treated with
various concentrations of galangin for 24 h, then the MTT analysis
was used to calculate cell viability. With the increasing of
galangin concentration, we found that TU212 cell viability was
reduced, especially at >10 µM, showing significant difference
compared to the Con ones. Additionally, after the TU212 cells were
treated with 10 µM galangin for different time as indicated, we
found that the cell viability was reduced in a time-dependent
manner (Fig. 2C). Furthermore, in
human laryngeal carcinoma HEP-2 cells, the cell viability was
decreased with the upregulation of galangin concentrations,
especially at >10 µM (Fig. 1D).
Also, HEP-2 cell viability was decreased by 10 µM galangin
administration for different times. In addition, significant
difference was observed for over 24 h (Fig. 1E). Finally, M4e cells were also
included to confirm the suppressive role of galangin in human
laryngeal carcinoma. From Fig. 1F,
shows that the M4e cell viability was downregulated for galangin
treatment, which was comparable to the Con ones at >20 µM.
Similarly, the cell viability was lower by 10 µM galangin treatment
for different times (Fig. 1G). The
data indicated that galangin indeed has a potential value in
reducing human laryngeal carcinoma cell proliferation in a dose-
and time-dependent manner.
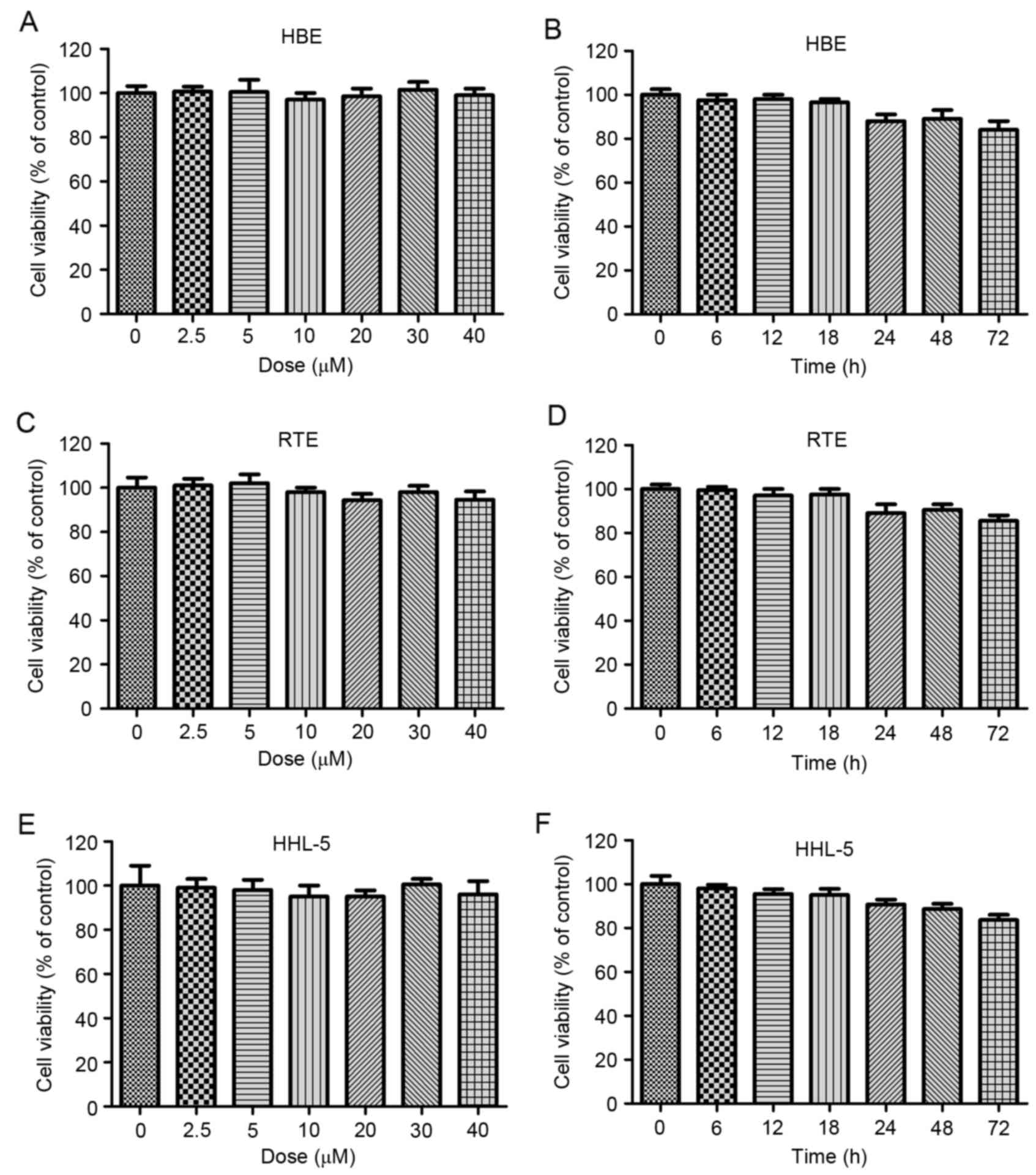 | Figure 2.Galangin shows no toxicity on normal
cells. (A) The human larynx epithelial HBE cells were treated with
different concentrations of galangin, ranged from 0 to 40 µM, for
24 h, and then the cell viability was calculated by MTT. (B) HBE
cells were treated with 40 µM galangin for the indicated time. The
cell viability was evaluated by MTT assays. (C) The mouse larynx
epithelial RTE cells were treated with different concentrations of
galangin, ranged from 0 to 40 µM, for 24 h, and the cell viability
was calculated by MTT. (D) RTE cells were treated with 40 µM
galangin for the indicated time. The cell viability was evaluated
by MTT assays. (E) The human normal liver HHL-5 cells were treated
with different concentrations of galangin, ranged from 0 to 40 µM,
for 24 h, and then the cell viability was calculated by MTT. (F)
HHL-5 cells were treated with 40 µM galangin for the indicated
time. The cell viability was evaluated by MTT assays. All analysis
were conducted in triplicate, and the results are the mean ± SEM of
three independent experiments. |
Galangin has been proved to be toxic for human
laryngeal carcinoma cells, and further study was needed to
calculate its effects on normal cells. As shown in Fig. 2A and B, no significant difference
was observed in human normal larynx epithelial HBE cells, between
the various groups treated under different concentrations of
galangin for different times. In line with the results above, in
the mouse larynx epithelial cells, no significant difference was
observed after galangin treatment under different conditions
(Fig. 2C and D). The human normal
liver HHL-5 cells were included to measure the toxicity of galangin
on normal cells. As shown in Fig. 2E
and F, galangin treatment under different conditions did not
alter the cell viability of HHL-5. The results indicated that
galangin shows no toxicity to normal cells, providing its
effectivity in human laryngeal carcinoma treatment in future.
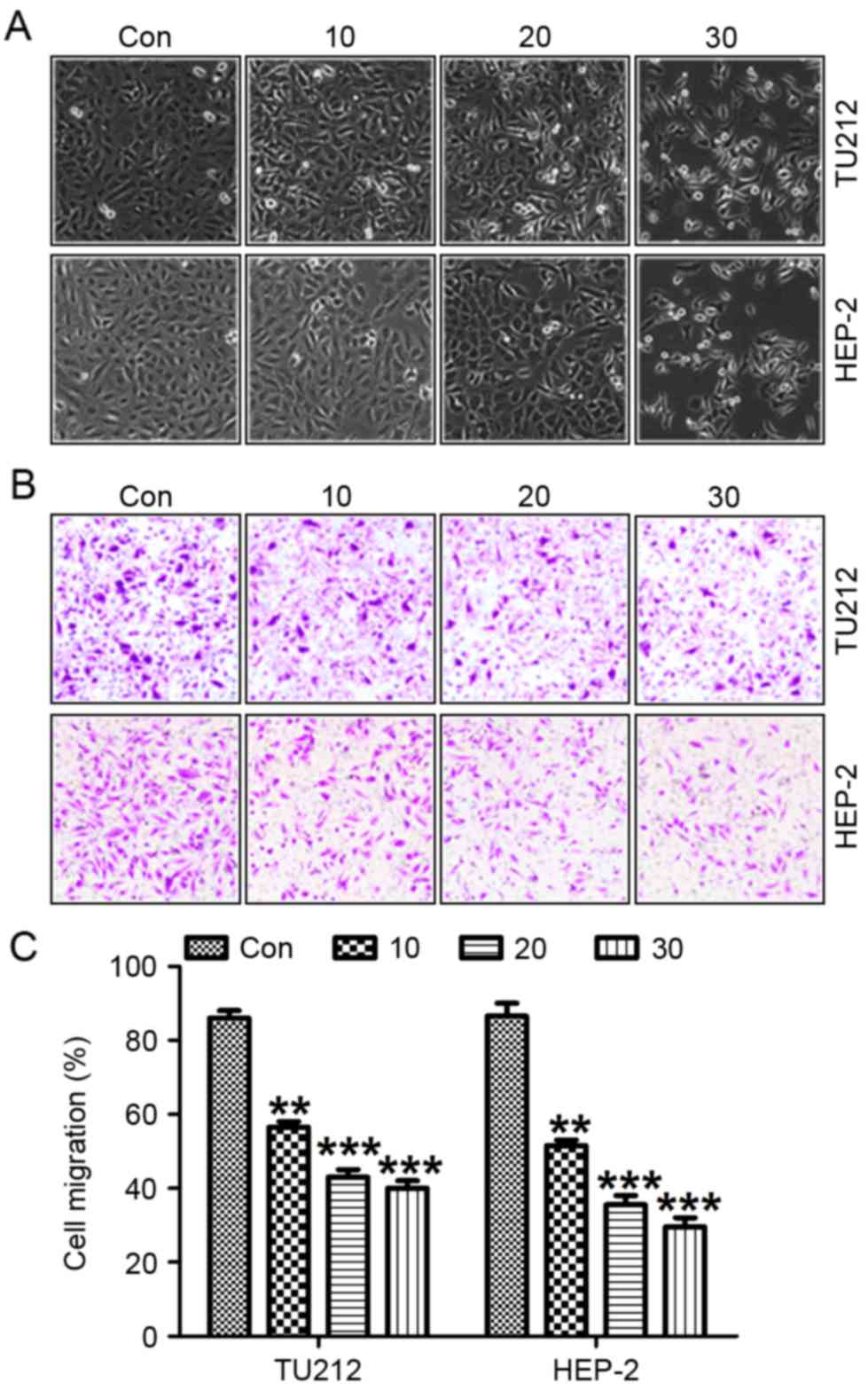
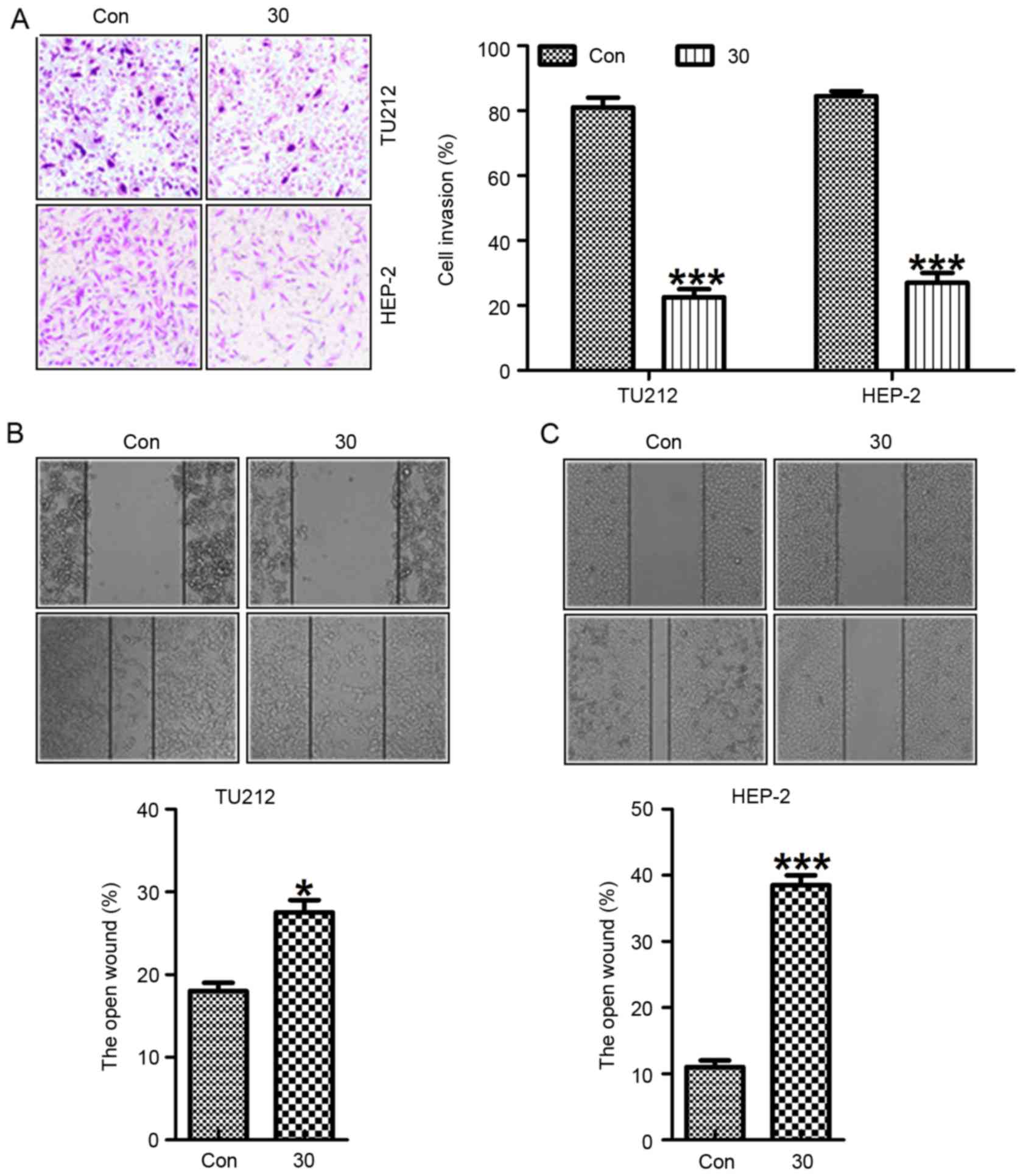
Galangin promotes laryngeal carcinoma
cell morphology alteration and suppresses migration, invasion as
well as proliferation
Exposure of TU212 and HEP-2 cells to galangin at
different concentrations resulted in necrotic morphological changes
and a downregulation in the percentage of viable cells, which was
dose-dependent (Fig. 3A).
Pretreatment with galangin considerably increased the inhibition of
migrated cancer cells (Fig. 3B and
C). In addition, we determined the invasion of TU212 and HEP-2
cells treated with galangin at the indicated dose. Fig. 4A shows that the invasion of TU212
and HEP-2 cells was markedly downregulated for galangin tretament
compared to the control ones. In addition, images were taken at 0
and 24 h after galangin treatment as shown in Fig 4B and C. TU212 and HEP-2 cells treated
with galangin for 24 h indicated that galangin suppressed the
cancer cell migration, which was comparable to the control group in
the absence of galangin. These results suggested that galangin
suppressed TU212 and HEP-2 cell proliferation.
Galangin induces apoptosis in human
laryngeal carcinoma cells of TU212 and HEP-2
In this regard, we attempted to explore the
mechanism of galangin on TU212 and HEP-2 cell growth and
progression. The data from flow cytometric assay clearly showed
that galangin induced apoptosis in TU212 cancer cells (Fig. 5A and B) and HEP-2 (Fig. 5A and C). In apoptosis process,
mitochondrial outer membrane permeabilization is known as a
‘point-of-no-return’ and is closely regulated by Bcl-2 family
proteins, especially Bax activation (25). Galangin apparently promoted Bax
protein expresssion and pre-treatment with galangin significantly
suppressed Bcl-2 protein levels (Fig.
5D-F). The results from western blot analysis showed that
galangin enhanced activation of cleaved caspase-3 and caspase-9,
resulting in increased PARP cleavage in TU212 (Fig. 5D and E) and HEP-2 cells (Fig. 5D and F). Furthermore, Bax activation
was confirmed to be increased for galangin treatment in TU212
(Fig. 5G and H) and HEP-2 cells
(Fig. 5G and I). These results
suggested that galangin treatment-triggered apoptosis was modulated
through caspase activation.
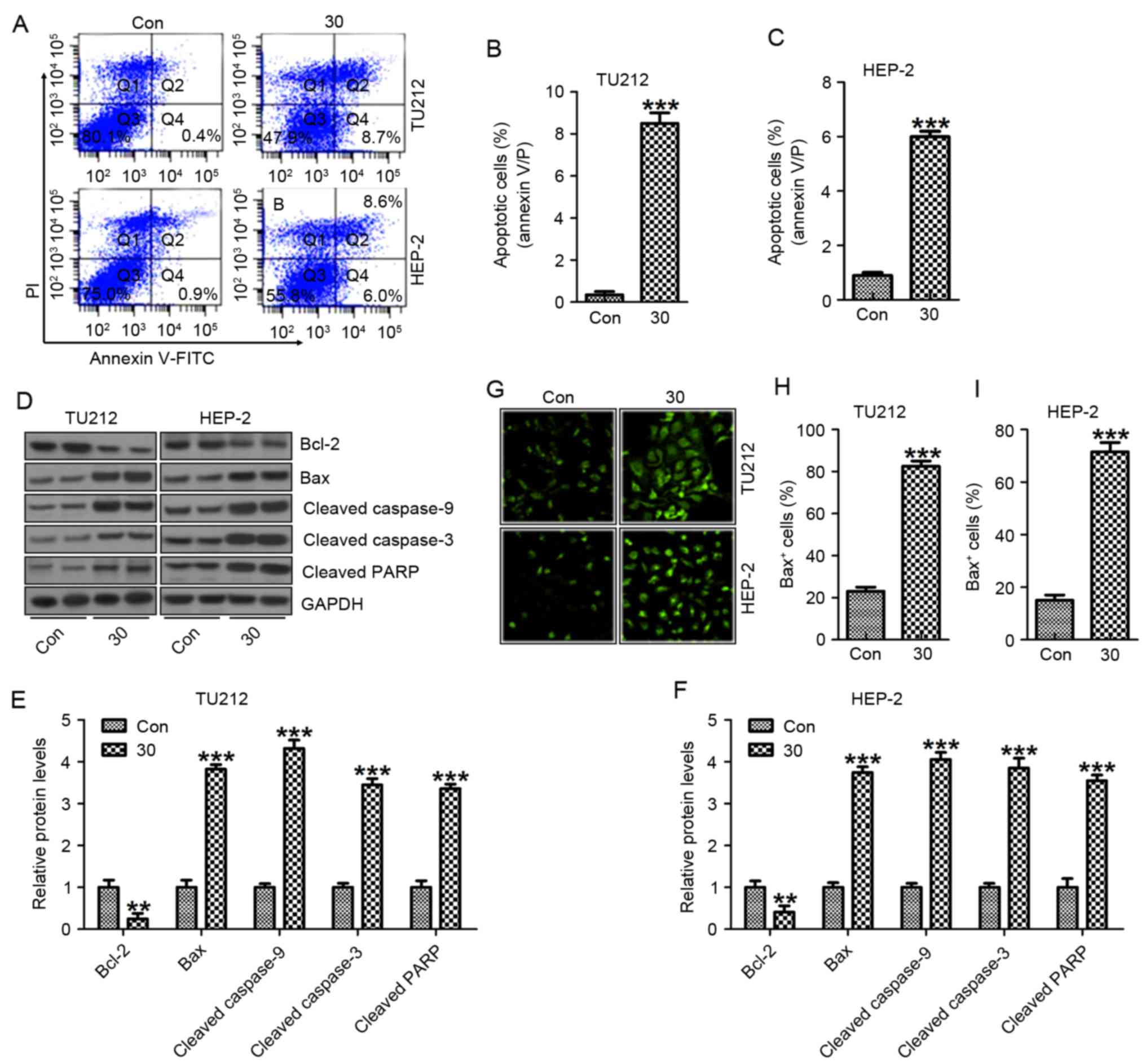 | Figure 5.Galangin induces apoptosis in human
laryngeal carcinoma TU212 and HEP-2 cells. (A) Flow cytometric
assays were applied to determine the number of apoptotic TU212 and
HEP-2 cells. (B) The quantification of TU212 apoptosis levels is
shown. (C) The quantification of HEP-2 apoptosis levels was shown.
(D) The signaling pathway, leading to apoptosis, was analyzed by
western blot analysis in TU212 and HEP-2 cells. Protein levels of
Bcl-2, Bax, cleaved caspase-9, cleaved caspase-3 and cleaved PARP
were evaluated. (E) Preotein levels of Bcl-2, Bax, cleaved
caspase-9, cleaved caspase-3 and cleaved PARP in TU212 cells were
quantified. (F) Protein levels of Bcl-2, Bax, cleaved caspase-9,
cleaved caspase-3 and cleaved PARP in HEP-2 cells were quantified.
(G) Immunofluorescence assays were carried out to determine Bax
positive TU212 and HEP-2 cells. The quantification of Bax positive
(H) TU212 and (I) HEP-2 cells are displayed. The analysis was
conducted in triplicate, and the results exhibit the mean ± SEM of
three independent experiments. **P<0.01 and ***P<0.001
(compared to the control/Con). |
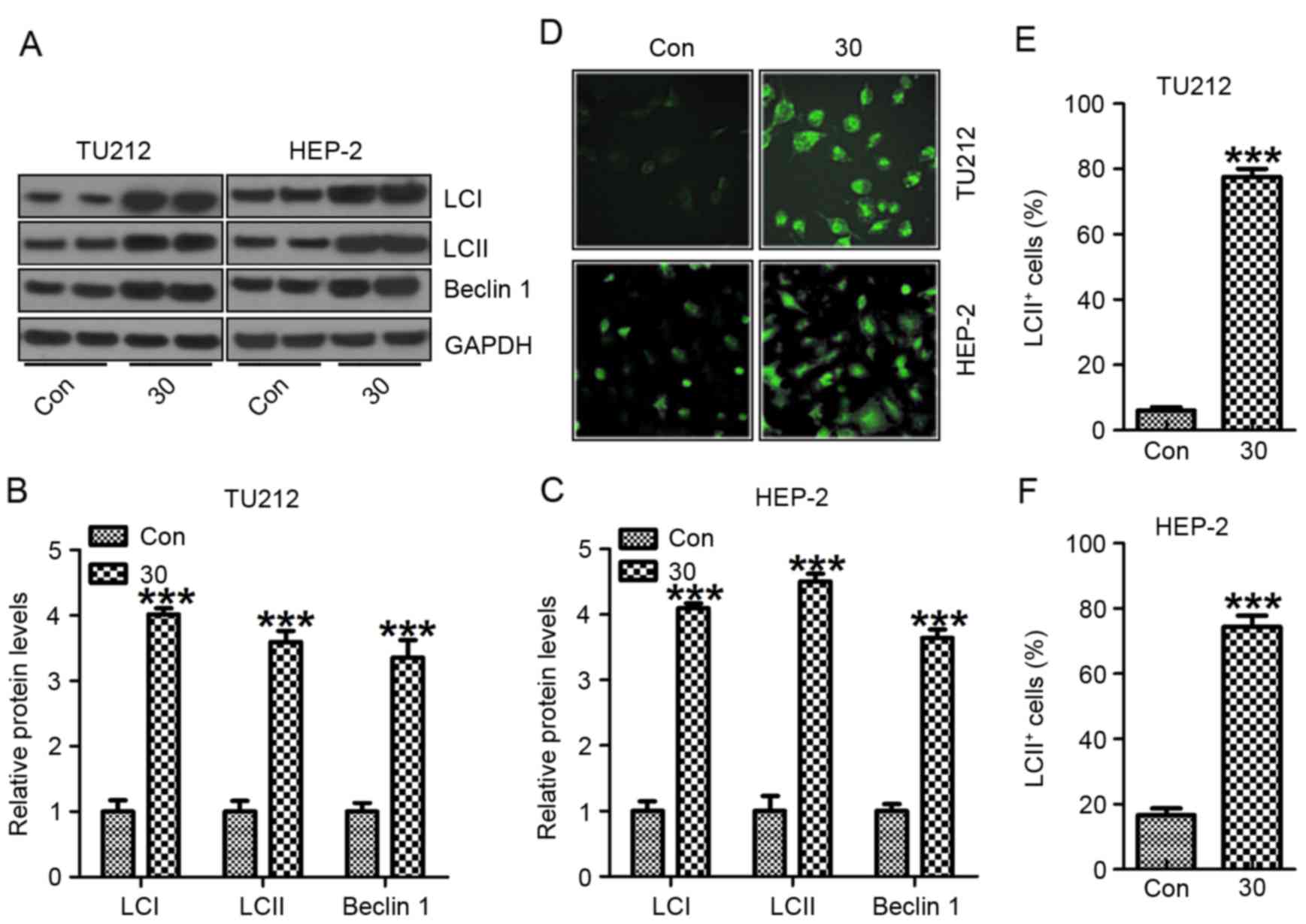
Galangin induces human laryngeal
carcinoma cell death through autophagy regulation
LCI, LC3II and Beclin 1 are required for the
autophagy-mediated elimination of unfolded ubiquinated long
half-life proteins (26). As shown
in Fig. 6A, western blot analysis
was carried out to reveal a considerable increase in LC3I, LC3II
and Beclin 1 expression induced by galangin exposure compared to
the control group without galangin treatment in TU212 (Fig. 6B) and HEP-2 (Fig. 6C) cells. The increased LC3I, LC3II
and Beclin 1 further suggested that galangin stimulated autophagy
and cell death in human laryngeal carcinoma (27). Similarly, in Fig. 6D-F, immunofluorescent assays further
evidenced that LC3II was highly induced in galangin treatment. The
data above indicate that galangin could result in human laryngeal
carcinoma cell death, contributing to tumor suppression.
Galangin inhibits human laryngeal
carcinoma TU212 and HEP-2 cell proliferation via p38 and AKT/NF-κB
suppression
To further explore the potential molecular mechanism
involved in galangin-induced TU212 and HEP-2 cell death, the
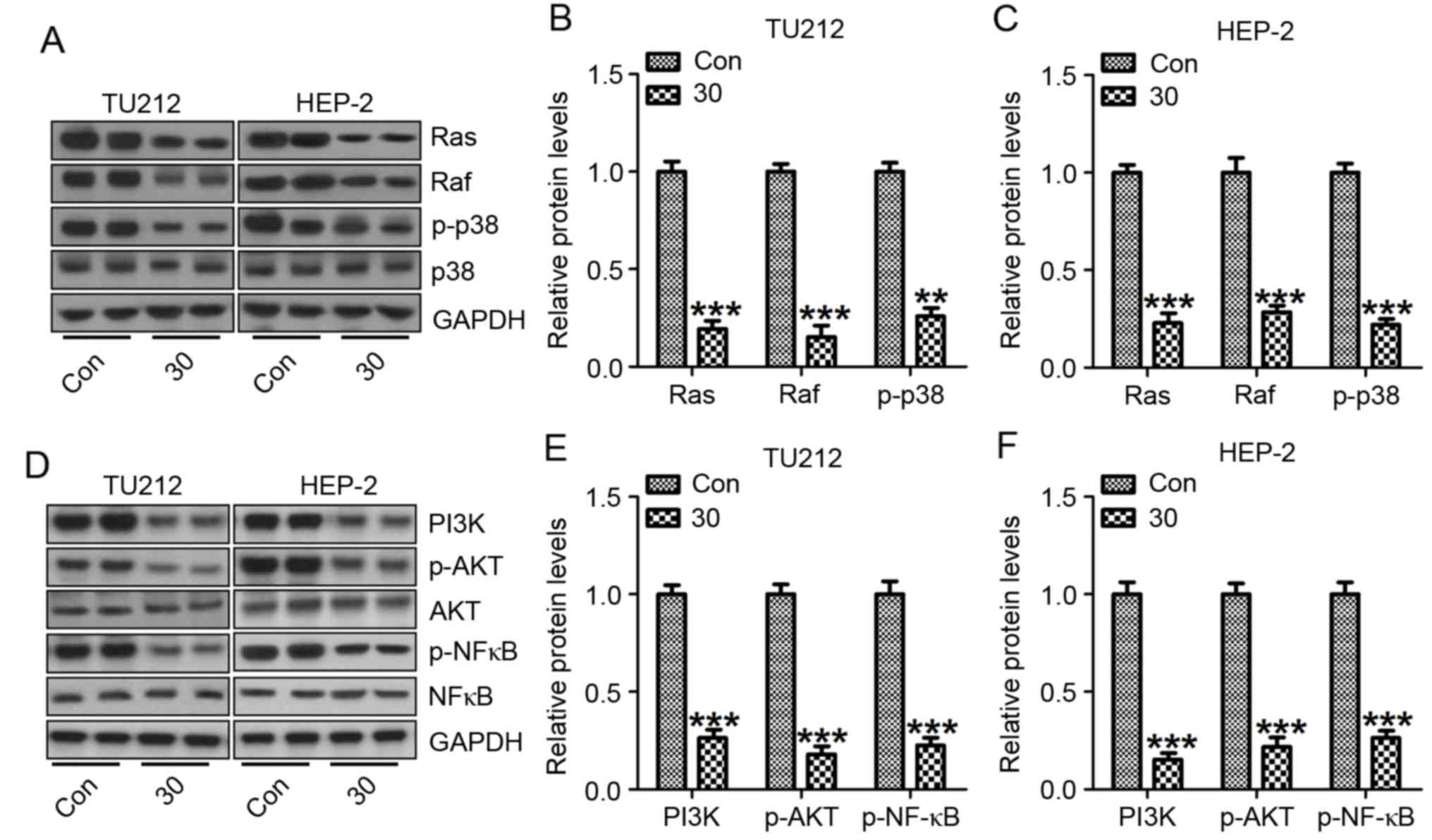
protein expression of Raf, Ras and p38 were calculated. The results
showed that Raf, Ras and p-p38 was highly expressed in the control
group without galangin treatment in both TU212 (Fig. 7A and B) and HEP-2 cells (Fig. 7A and C). Importantly, galangin
supplementation partially reversed the overexpression of Raf, Ras
and p-p38. Enhancement of p38 activation plays an important role in
modulating tumor migration, invasion as well as metastasis in some
cancer cases (28). Altogether,
galangin treatment-attenuated TU212 and HEP-2 cell progression
partially relied on p38 signaling pathway.
NF-κB is a pleiotropic transcription factor, which
is related to various biological processes, including inflammation,
apoptosis as well as autophogy (29). NF-κB activation has been detected in
>50% of tumors, regulated by PI3K/AKT signaling pathway
(30). In this study, PI3K/AKT was
expressed highly in the cancer cells without galangin treatment,
which was downregulated after galangin administration in both TU212
(Fig. 7D and E) and HEP-2 (Fig. 7D and F) cells. These results
indicated that galangin might play important roles in TU212 and
HEP-2 cells as an antitumor agent inhibiting cancer cell
proliferation by targeting p38 and NF-κB.
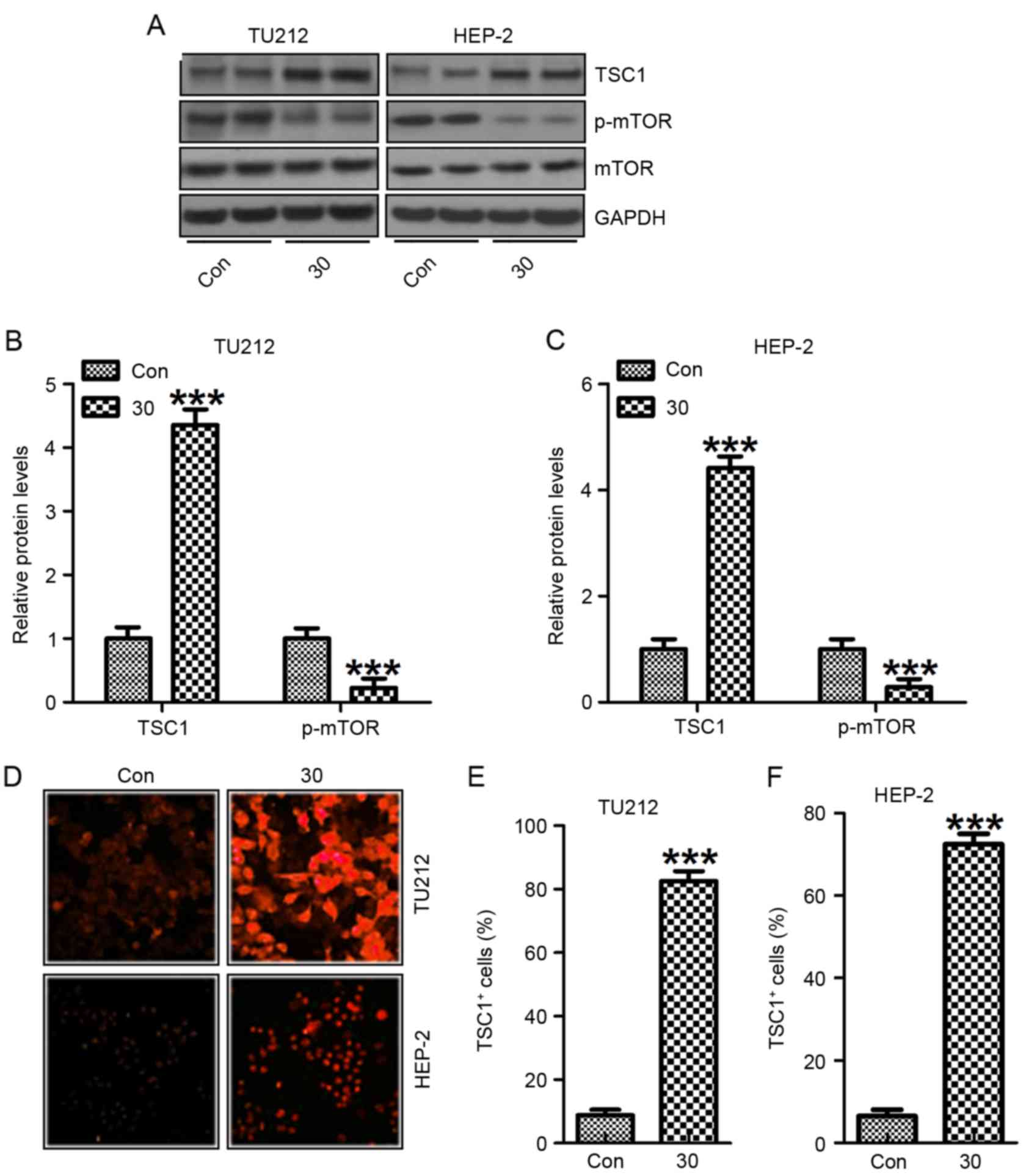
Galangin impedes human laryngeal
carcinoma TU212 and HEP-2 cell proliferation via suppressing
mTOR
Finally, suppressing the PI3K-Akt-mTOR signaling
pathway has been considered as an essential molecular mechanism
causing tumor suppression (31). In
this regard, we found that TSC1, an inhibitor of mTOR activation,
was upregulated after the treatment with galangin. On the contrary,
mTOR phosphorylated levels were reduced in TU212 and HEP-2 cells
(Fig. 8A-C). Also,
immunofluorescent analysis was performed to explore TSC1 levels in
both TU212 (Fig. 8D and E) and
HEP-2 (Fig. 8D and F) cells. TSC1
were highly expressed for galangin treatment in comparison to the
control ones. Collectively, these results indicated that galangin
suppressed human laryngeal carcinoma cell growth via mTOR signaling
pathway.
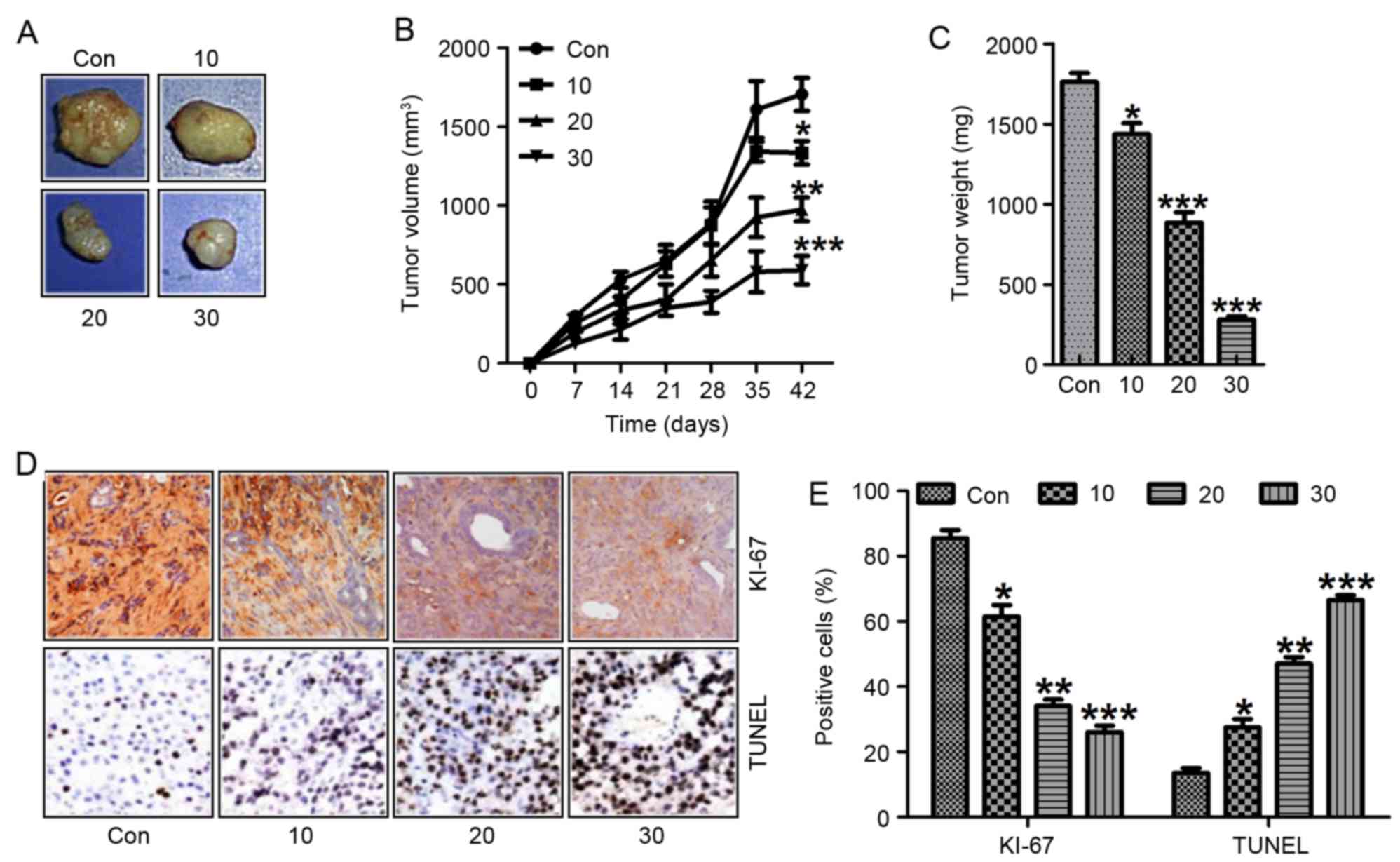
Galangin promotes human laryngeal
carcinoma growth inhibition in a xenograft tumor model in vivo
To confirm the enhanced galangin-regulated
inhibition of human laryngeal carcinoma growth, we analyzed the
effects of galangin treatment on tumorigenicity in vivo
using a TU212 xenograft mouse model. After administration with
galangin for 42 days, both the tumor volume and weight were
inhibited (Fig. 9A-C).
Additionally, reduction of tumor Ki-67 (Fig. 9D and E) and upregulation of TUNEL
(Fig. 9D and E) through IHC assays
were also noted in the galangin-treated group in a dose-dependent
manner. These results indicated that galangin could promote
suppression of xenografted human laryngeal carcinoma cell growth
in vivo, which was in line with the data in
vitro.
Discussion
Human laryngeal cancer is known as one of the most
common tumors of the head and neck region in the world (1,2,32).
Patients suffering from laryngeal cancer show a poor survival rate
with poor advance during the last decades (33). Thus, more effective treatments are
needed to be explored for preventing laryngeal cancer. Natural
compounds show an essential role in cancer and other disease
prevention and treatment worldwide (4). Galangin is known as an anti-tumor
agent, which is effective in preventing a broad range of tumors,
such as liver, breast, as well as lung cancer (14–16,34).
Galangin has been reported to perform its role in inhibiting tumor
growth through apoptosis regulation (35). However, whether galangin could be a
potential compound for human laryngeal cancer prevention is not
known. Thus, galangin was used here to provide possible therapeutic
strategy for suppressing human laryngeal cancer. Notably, it is the
first study to explore the molecular mechanisms of
galangin-triggered human laryngeal cancer cell death. In our study,
we found that exposure of galangin into TU212 and HEP-2 cells could
partially suppress cancer cell proliferation, invasion and
migration. Autophagy is a regulated process of degradation and
recycling of cellular constituents; the process is important in
organelle turnover and the bioenergetics management of starvation
(36). During autophagy, the
precursor form of LC3 is post-modified into LC3-I and LC3-II
(37,38). LC3-I is localized in the cytosol,
and LC3-II is a membrane-associated and a key hallmark for
autophagosome formation (39).
LC3-II can be used to estimate the abundance of autophagosomes
before they are destroyed through fusion with lysosomes (40). Here, augmented LC3II was found after
galangin treatment, indicating that the autophagy was induced.
Obviously in in vivo study, the tumor size and weight was
reduced for galangin administration. The results suggested that
galangin has potential, to be developed as a therapeutic strategy
for human laryngeal carcinoma.
PI3K/AKT signal pathway is crucial for regulating
various cell activities, such as proliferation, cell growth,
survival, chemotaxis, the inflammatory response and apoptosis
(41). Enhanced activation of
PI3K/AKT pathway is linked to the development and progression of
many cancers, as well as resistance to chemotherapy (42). A previous study reported that
inactivation of PI3K/AKT signaling pathway is involved in the
success of chemotherapy-caused apoptosis in some cancer cells
(43,44). In the present study, PI3K was
reduced in galangin treatment, subsequently downregulating the
protein levels of phosphorylated AKT, being in line with previous
results, indicating that galangin might perform its role in
suppressing human laryngeal carcinoma through PI3K/AKT signaling
pathway inactivation. Apoptosis constitutes a fundamental intrinsic
mechanism of tumor suppression, as the resistance of apoptosis is a
well-established hallmark of cancer (45,46).
It is known that Bcl-2 protein is also a key regulator for
apoptosis and its tumorigenic potential is supported by the finding
of overexpression of Bcl-2 in various types of tumor, which is
related to the activation of AKT (47). AKT is a key player in regulating
cell signals that are important for cell death and survival.
Activation of the AKT pathway promotes cell survival and is
involved in the upregulation of Bcl-2 (48,49).
In this study, we found that with the altered trend of AKT
expressed levels, Bcl-2 was reduced in galangin treatment. The
caspase-3 and PARP cleavage were highly improved, contributing to
the death of TU212 and HEP-2 cells and leading to cell apoptosis
and cell death due to galangin tretament. The results above
suggested that galangin suppressed human laryngeal cancer
development and progression through apoptosis induction.
NF-κB takes part in the information transfer
process, including tissue damage, apoptosis, as well as stress,
tumor suppression and cell differentiation, thus, it is an
important nuclear transcription factor (50,51).
NF-κB in different tumors or cancers is modulated by PI3K/AKT
pathway (52). P-AKT activates IκB
kinase (IKKα), leading to inhibition of NF-κB degradation by IκB,
allowing NF-κB to be transferred into the nucleus from the
cytoplasm, where it activates its target genes and promotes cell
survival (53). In this study,
NF-κB was markedly activated accompanied by AKT phosphorylation in
the absence of galangin treatment. However, galangin administration
downregulated NF-κB phosphorylated activity, leading to the
upregulation of caspase-9, caspase-3 and PARP cleavage, indicating
that galangin could inhibit human laryngeal cancer via AKT-mediated
NF-κB signaling pathway. In addition, PI3K/AKT pathway is of great
importance for the cell proliferation. One of the best-known
downstream substrates of PI3K/AKT is the mammalian target of
rapamycin (mTOR), inducing mammalian autophagy significantly,
regulating protein translation (54). Previously it was indicated that the
activation of PI3K-AKT-mTOR signal pathway may be involved in
autophagy suppression (55).
Impeding PI3K-AKT-mTOR signal pathway has been indicated to be
effective for various cancers treatment (56,57).
In our investigation, here the phosphorylated mTOR in high
expression was suppressed in TU212 and HEP-2 cells for galangin
administration, and in contrast, TSC1, an inhibitor of mTOR, was
upregulated for galangin. The results illustrated that galangin
prevented human laryngeal cancer through inhibiting laryngeal
cancer cells proliferation and inducing autophagy via
PI3K/AKT-regulated mTOR activity.
Collectively, our findings above demonstrated that
galangin prevented human laryngeal cancer proliferation, invasion
and migration by PI3K/AKT and p38 suppression, resulting in caspase
activation, NF-κB dephosphorylation as well as mTOR inactivation
with reduced Ki-67 expression and enhanced TUNEL levels. The
present study indicated that the use of the dietary compound
galangin might be a potential therapeutic strategy for human
laryngeal carcinoma treatment.
References
|
1
|
Zhang Y, Chen Y, Yu J, Liu G and Huang Z:
Integrated transcriptome analysis reveals miRNA-mRNA crosstalk in
laryngeal squamous cell carcinoma. Genomics. 104:249–256. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liu M, Wu H, Liu T, Li Y, Wang F, Wan H,
Li X and Tang H: Regulation of the cell cycle gene, BTG2, by miR-21
in human laryngeal carcinoma. Cell Res. 19:828–837. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
van Dijk BA, Karim-Kos HE, Coebergh JW,
Marres HA and de Vries E: Progress against laryngeal cancer in The
Netherlands between 1989 and 2010. Int J Cancer. 134:674–681. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hoffman HT, Porter K, Karnell LH, Cooper
JS, Weber RS, Langer CJ, Ang KK, Gay G, Stewart A and Robinson RA:
Laryngeal cancer in the United States: Changes in demographics,
patterns of care, and survival. Laryngoscope. 116:(Suppl 111).
1–13. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ouyang D, Liu TR, Liu XW, Chen YF, Wang J,
Su X and Yang AK: Combined hyoid bone flap in laryngeal
reconstruction after extensive partial laryngectomy for laryngeal
cancer. Eur Arch Otorhinolaryngol. 270:1455–1462. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Diab S, Pascoe J, Shahriar M, Read D,
Kinde H, Moore J, Odani J and Uzal F: Study of laryngopharyngeal
pathology in Thoroughbred horses in southern California. Equine Vet
J. 41:903–907. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Silver CE, Beitler JJ, Shaha AR, Rinaldo A
and Ferlito A: Current trends in initial management of laryngeal
cancer: The declining use of open surgery. Eur Arch
Otorhinolaryngol. 266:1333–1352. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ulualp SO: Mapping regional
laryngopharyngeal mechanoreceptor response. Clin Exp
Otorhinolaryngol. 7:319–323. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang SY, Lu ZM, Chen LS, Luo XN, Ge PJ,
Song XH, Chen SH and Wu YL: Supracricoid partial laryngectomy
cricohyoidoepiglottopexy (SCPL-CHEP) versus vertical partial
laryngectomy for the treatment of glottic carcinoma. Eur Arch
Otorhinolaryngol. 270:1027–1034. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang HT, Luo H, Wu J, Lan LB, Fan DH, Zhu
KD, Chen XY, Wen M and Liu HM: Galangin induces apoptosis of
hepatocellular carcinoma cells via the mitochondrial pathway. World
J Gastroenterol. 16:3377–3384. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Capasso R and Mascolo N: Inhibitory effect
of the plant flavonoid galangin on rat vas deferens in vitro. Life
Sci. 72:2993–3001. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kim DA, Jeon YK and Nam MJ: Galangin
induces apoptosis in gastric cancer cells via regulation of
ubiquitin carboxy-terminal hydrolase isozyme L1 and glutathione
S-transferase P. Food Chem Toxicol. 50:684–688. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang W, Tang B, Huang Q and Hua Z:
Galangin inhibits tumor growth and metastasis of B16F10 melanoma. J
Cell Biochem. 114:152–161. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Heo MY, Sohn SJ and Au WW:
Anti-genotoxicity of galangin as a cancer chemopreventive agent
candidate. Mutat Res. 488:135–150. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Agati G, Azzarello E, Pollastri S and
Tattini M: Flavonoids as antioxidants in plants: Location and
functional significance. Plant Sci. 196:67–76. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Parhiz H, Roohbakhsh A, Soltani F, Rezaee
R and Iranshahi M: Antioxidant and anti-inflammatory properties of
the citrus flavonoids hesperidin and hesperetin: An updated review
of their molecular mechanisms and experimental models. Phytother
Res. 29:323–331. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Schultz DR and Harrington WJ Jr:
Apoptosis: Programmed cell death at a molecular level. Semin
Arthritis Rheum. 32:345–369. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tomita T: Cleaved caspase-3
immunocytochemical staining for pancreatic islets and pancreatic
endocrine tumors: A potential marker for biological malignancy.
Islets. 2:82–88. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
McCormick J, Knight RA, Barry SP,
Scarabelli TM, Abounit K, Latchman DS and Stephanou A: Autophagy in
the stress-induced myocardium. Front Biosci (Elite Ed).
4:2131–2141. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li Y, Zhang Q, Tian R, Wang Q, Zhao JJ,
Iglehart JD, Wang ZC and Richardson AL: Lysosomal transmembrane
protein LAPTM4B promotes autophagy and tolerance to metabolic
stress in cancer cells. Cancer Res. 71:7481–7489. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu Z, Antalek M, Nguyen L, Li X, Tian X,
Le A and Zi X: The effect of gartanin, a naturally occurring
xanthone in mangosteen juice, on the mTOR pathway, autophagy,
apoptosis, and the growth of human urinary bladder cancer cell
lines. Nutr Cancer. 65:(Suppl 1). 68–77. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ling YH, Aracil M, Zou Y, Yuan Z, Lu B,
Jimeno J, Cuervo AM and Perez-Soler R: PM02734 (elisidepsin)
induces caspase-independent cell death associated with features of
autophagy, inhibition of the Akt/mTOR signaling pathway, and
activation of death-associated protein kinase. Clin Cancer Res.
17:5353–5366. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhai B, Hu F, Jiang X, Xu J, Zhao D, Liu
B, Pan S, Dong X, Tan G, Wei Z, et al: Inhibition of Akt reverses
the acquired resistance to sorafenib by switching protective
autophagy to autophagic cell death in hepatocellular carcinoma. Mol
Cancer Ther. 13:1589–1598. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Graham TR, Odero-Marah VA, Chung LW,
Agrawal KC, Davis R and Abdel-Mageed AB: PI3K/Akt-dependent
transcriptional regulation and activation of BMP-2-Smad signaling
by NF-kappaB in metastatic prostate cancer cells. Prostate.
69:168–180. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pugazhenthi S, Nesterova A, Sable C,
Heidenreich KA, Boxer LM, Heasley LE and Reusch JE: Akt/protein
kinase B up-regulates Bcl-2 expression through cAMP-response
element-binding protein. J Biol Chem. 275:10761–10766. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hale AN, Ledbetter DJ, Gawriluk TR and
Rucker EB III: Autophagy: Regulation and role in development.
Autophagy. 9:951–972. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li C, Han X, Zhang H, Wu J and Li B: The
interplay between autophagy and apoptosis induced by tanshinone IIA
in prostate cancer cells. Tumour Biol. 37:7667–7674. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Platanias LC: Map kinase signaling
pathways and hematologic malignancies. Blood. 101:4667–4679. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rauch BH, Weber A, Braun M, Zimmermann N
and Schrör K: PDGF-induced Akt phosphorylation does not activate
NF-kappa B in human vascular smooth muscle cells and fibroblasts.
FEBS Lett. 481:3–7. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ryu HJ, Kim JE, Yeo SI and Kang TC:
p65/RelA-Ser529 NF-κB subunit phosphorylation induces autophagic
astroglial death (Clasmatodendrosis) following status epilepticus.
Cell Mol Neurobiol. 31:1071–1078. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Samuels Y and Velculescu VE: Oncogenic
mutations of PIK3CA in human cancers. Cell Cycle. 3:1221–1224.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen K, Song F, He M, Li H, Qian B, Zhang
W, Wei Q and Hao X: Trends in head and neck cancer incidence in
Tianjin, China, between 1981 and 2002. Head Neck. 31:175–182. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li XY, Guo X, Feng S, Li XT, Wei HQ, Yang
HA, Ren Z and Jiang XJ: Relationship between a family history of
malignancy and the incidence of laryngeal carcinoma in the Liaoning
province of China. Clin Otolaryngol. 34:127–131. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li S, Chou G, Hseu Y, Yang H, Kwan H and
Yu Z: Isolation of anticancer constituents from flos genkwa (Daphne
genkwa Sieb.et Zucc.) through bioassay-guided procedures. Chem Cent
J. 7:1592013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lee LT, Huang YT, Hwang JJ, Lee AY, Ke FC,
Huang CJ, Kandaswami C, Lee PP and Lee MT: Transinactivation of the
epidermal growth factor receptor tyrosine kinase and focal adhesion
kinase phosphorylation by dietary flavonoids: Effect on invasive
potential of human carcinoma cells. Biochem Pharmacol.
67:2103–2114. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Khandelwal VK, Mitrofan LM, Hyttinen JM,
Chaudhari KR, Buccione R, Kaarniranta K, Ravingerová T and
Mönkkönen J: Oxidative stress plays an important role in zoledronic
acid-induced autophagy. Physiol Res. 63:(Suppl 4). S601–S612.
2014.PubMed/NCBI
|
|
37
|
McLeland CB, Rodriguez J and Stern ST:
Autophagy monitoring assay: Qualitative analysis of MAP LC3-I to II
conversion by immunoblot. Methods Mol Biol. 697:199–206. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Giménez-Xavier P, Francisco R, Platini F,
Pérez R and Ambrosio S: LC3-I conversion to LC3-II does not
necessarily result in complete autophagy. Int J Mol Med.
22:781–785. 2008.PubMed/NCBI
|
|
39
|
Kroemer G and Levine B: Autophagic cell
death: The story of a misnomer. Nat Rev Mol Cell Biol. 9:1004–1010.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Reggiori F, Monastyrska I, Verheije MH,
Calì T, Ulasli M, Bianchi S, Bernasconi R, de Haan CA and Molinari
M: Coronaviruses Hijack the LC3-I-positive EDEMosomes, ER-derived
vesicles exporting short-lived ERAD regulators, for replication.
Cell Host Microbe. 7:500–508. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Saini KS, Loi S, de Azambuja E,
Metzger-Filho O, Saini ML, Ignatiadis M, Dancey JE and
Piccart-Gebhart MJ: Targeting the PI3K/AKT/mTOR and Raf/MEK/ERK
pathways in the treatment of breast cancer. Cancer Treat Rev.
39:935–946. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Han L, Yang Y, Yue X, Huang K, Liu X, Pu
P, Jiang H, Yan W, Jiang T and Kang C: Inactivation of PI3K/AKT
signaling inhibits glioma cell growth through modulation of
β-catenin-mediated transcription. Brain Res. 1366:9–17. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lee J, Zhang G, Wu X, Xu F, Zhou J and
Zhang X: Growth inhibitory effect of dihydroartemisinin on
Bcr/Abl+ chronic myeloid leukemia K562 cells involve
AKT, ERK and NF-κB modulation. J Cancer Res Clin Oncol.
138:2095–2102. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yip KW and Reed JC: Bcl-2 family proteins
and cancer. Oncogene. 27:6398–6406. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Noble P, Vyas M, Al-Attar A, Durrant S,
Scholefield J and Durrant L: High levels of cleaved caspase-3 in
colorectal tumour stroma predict good survival. Br J Cancer.
108:2097–2105. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Liu K, Liu PC, Liu R and Wu X: Dual AO/EB
staining to detect apoptosis in osteosarcoma cells compared with
flow cytometry. Med Sci Monit Basic Res. 21:15–20. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Sheppard KE, Cullinane C, Hannan KM, Wall
M, Chan J, Barber F, Foo J, Cameron D, Neilsen A, Ng P, et al:
Synergistic inhibition of ovarian cancer cell growth by combining
selective PI3K/mTOR and RAS/ERK pathway inhibitors. Eur J Cancer.
49:3936–3944. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kim KW, Moretti L, Mitchell LR, Jung DK
and Lu B: Combined Bcl-2/mammalian target of rapamycin inhibition
leads to enhanced radiosensitization via induction of apoptosis and
autophagy in non-small cell lung tumor xenograft model. Clin Cancer
Res. 15:6096–6105. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Levine B, Sinha S and Kroemer G: Bcl-2
family members: Dual regulators of apoptosis and autophagy.
Autophagy. 4:600–606. 2008. View Article : Google Scholar :
|
|
50
|
Go EK, Jung KJ, Kim JY, Yu BP and Chung
HY: Betaine suppresses proinflammatory signaling during aging: The
involvement of nuclear factor-kappaB via nuclear factor-inducing
kinase/IkappaB kinase and mitogen-activated protein kinases. J
Gerontol A Biol Sci Med Sci. 60:1252–1264. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Lin YG, Kunnumakkara AB, Nair A, Merritt
WM, Han LY, Armaiz-Pena GN, Kamat AA, Spannuth WA, Gershenson DM,
Lutgendorf SK, et al: Curcumin inhibits tumor growth and
angiogenesis in ovarian carcinoma by targeting the nuclear
factor-kappaB pathway. Clin Cancer Res. 13:3423–3430. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Jayasooriya RG, Choi YH, Moon SK, Kim WJ
and Kim GY: Methanol extract of Hydroclathrus clathratus suppresses
matrix metalloproteinase-9 in T24 bladder carcinoma cells by
suppressing the NF-κB and MAPK pathways. Oncol Rep. 27:541–546.
2012.PubMed/NCBI
|
|
53
|
Li W, Wang H, Kuang CY, Zhu JK, Yu Y, Qin
ZX, Liu J and Huang L: An essential role for the
Id1/PI3K/Akt/NF-κB/survivin signalling pathway in promoting the
proliferation of endothelial progenitor cells in vitro. Mol Cell
Biochem. 363:135–145. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Roberts PJ and Der CJ: Targeting the
Raf-MEK-ERK mitogen-activated protein kinase cascade for the
treatment of cancer. Oncogene. 26:3291–3310. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Sun CH, Chang YH and Pan CC: Activation of
the PI3K/Akt/mTOR pathway correlates with tumour progression and
reduced survival in patients with urothelial carcinoma of the
urinary bladder. Histopathology. 58:1054–1063. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Steelman LS, Abrams SL, Whelan J, Bertrand
FE, Ludwig DE, Bäsecke J, Libra M, Stivala F, Milella M, Tafuri A,
et al: Contributions of the Raf/MEK/ERK, PI3K/PTEN/Akt/mTOR and
Jak/STAT pathways to leukemia. Leukemia. 22:686–707. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zhang P, Pang X and Tu Y:
Thioredoxin-interacting protein as a common regulation target for
multiple drugs in clinical therapy/application. Cancer Transl Med.
1:26–30. 2015. View Article : Google Scholar
|